

Electro-Enhanced Gas Fermentation for Bioproduction of Volatile Fatty Acids and Alcohols


Abstract
1. Introduction
2. Materials and Method
2.1. Microorganisms
2.2. Media and Additives
2.3. Experimental Procedure
2.4. Analysis
2.4.1. Potentiostat and Current Measurement Device
2.4.2. HPLC
3. Results
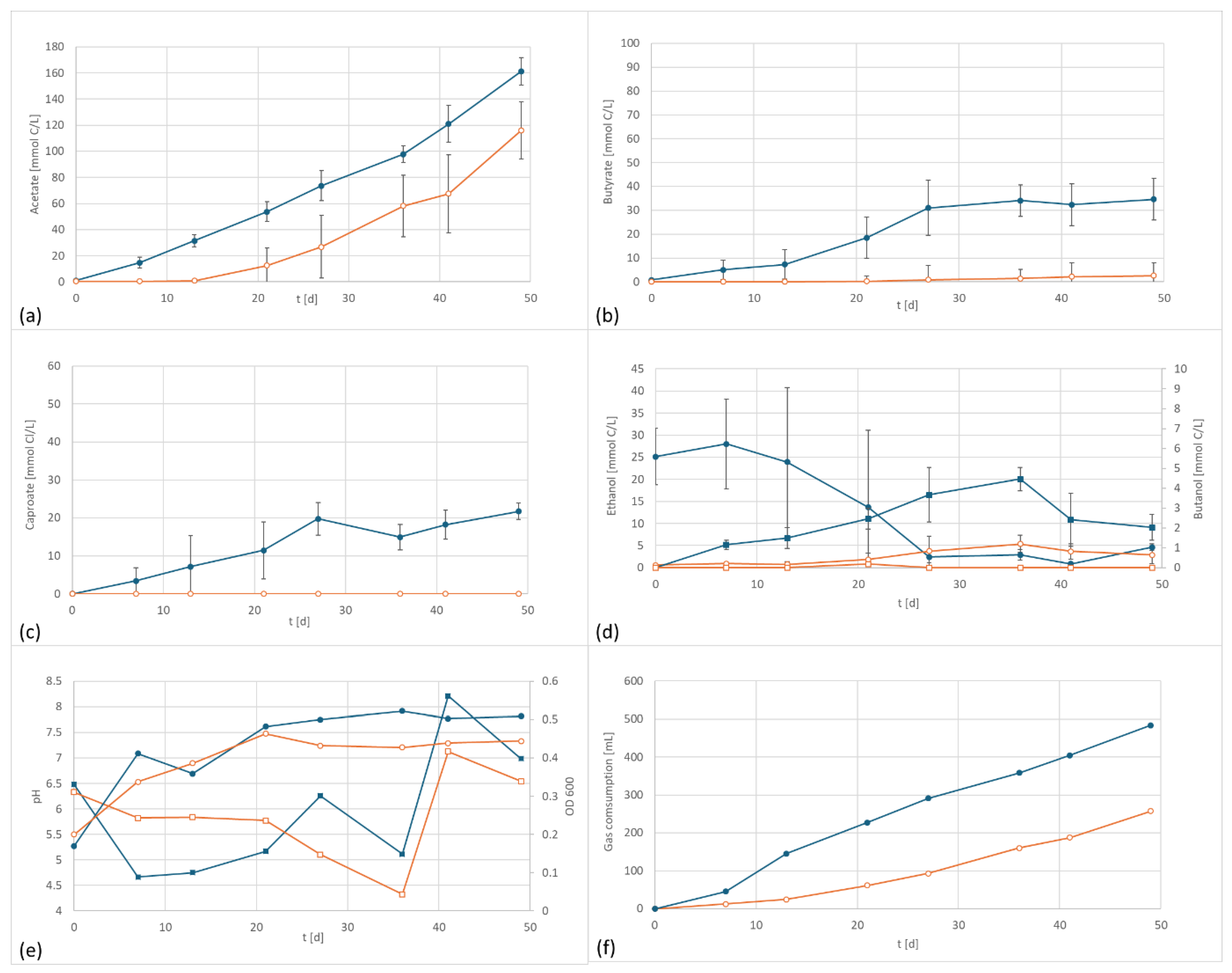
3.1. C. carboxidivorans Culture
3.2. A. bacchi Culture
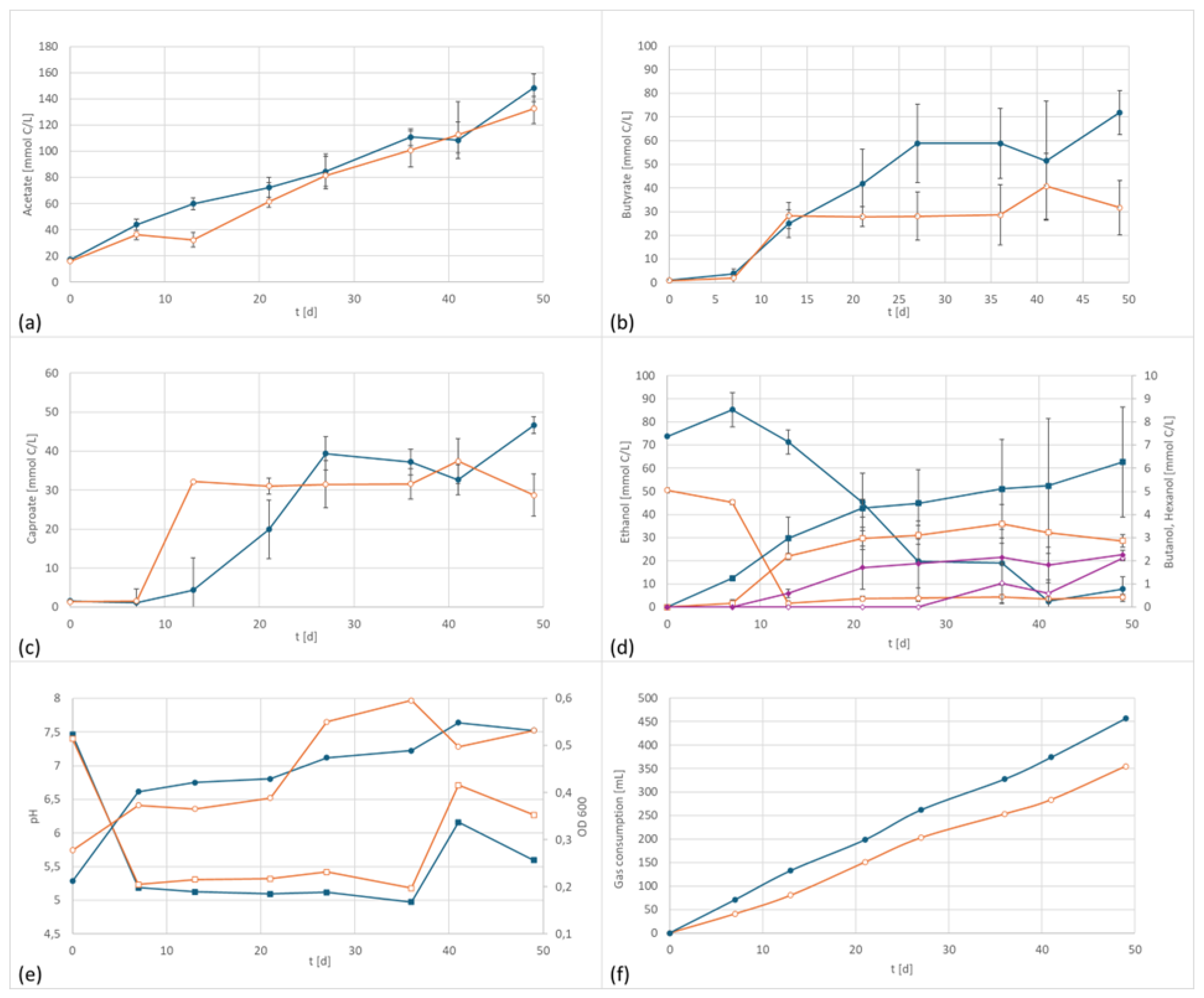
3.3. Co-Culture
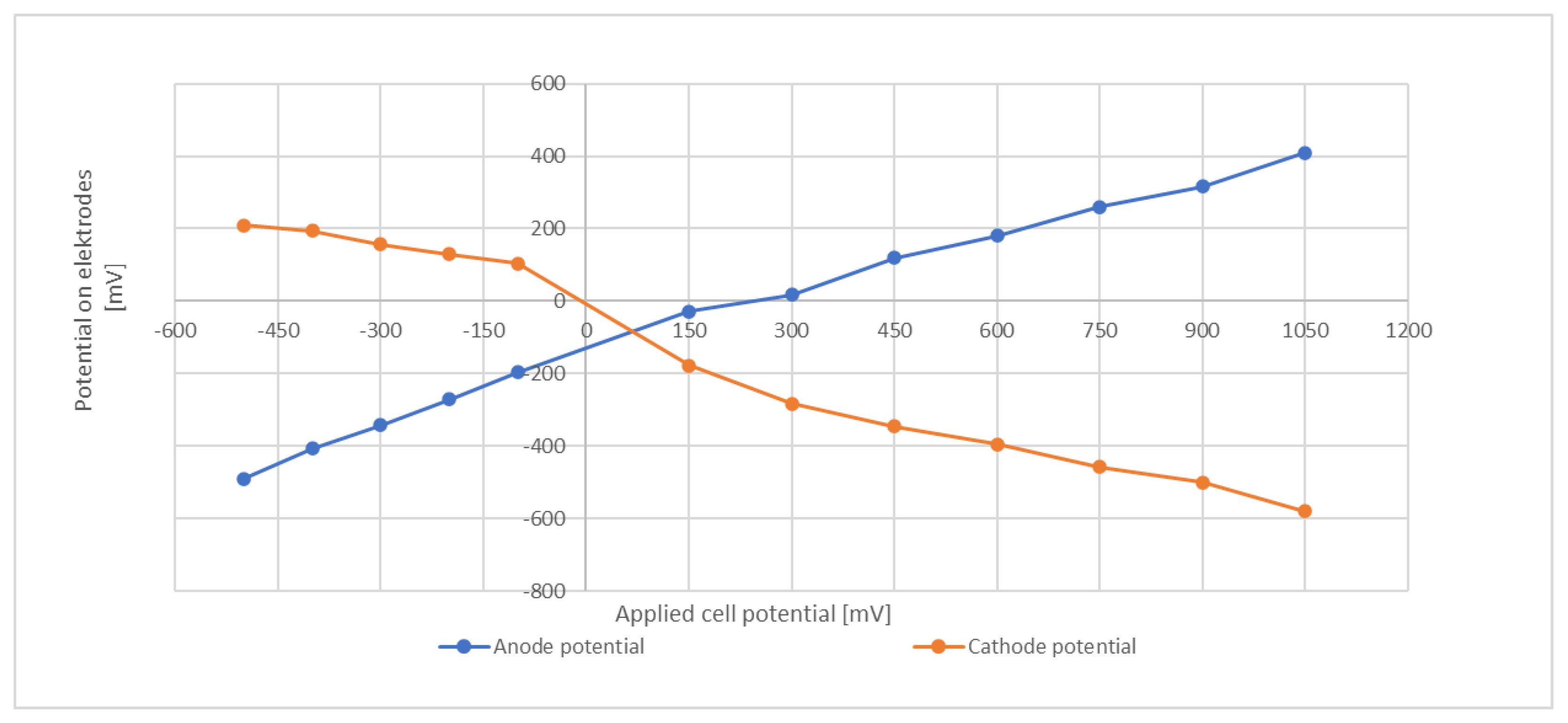
3.4. Electric Current Flow
3.5. Cathode Potential
3.6. Counterreaction
3.7. Comparison to Literature Data
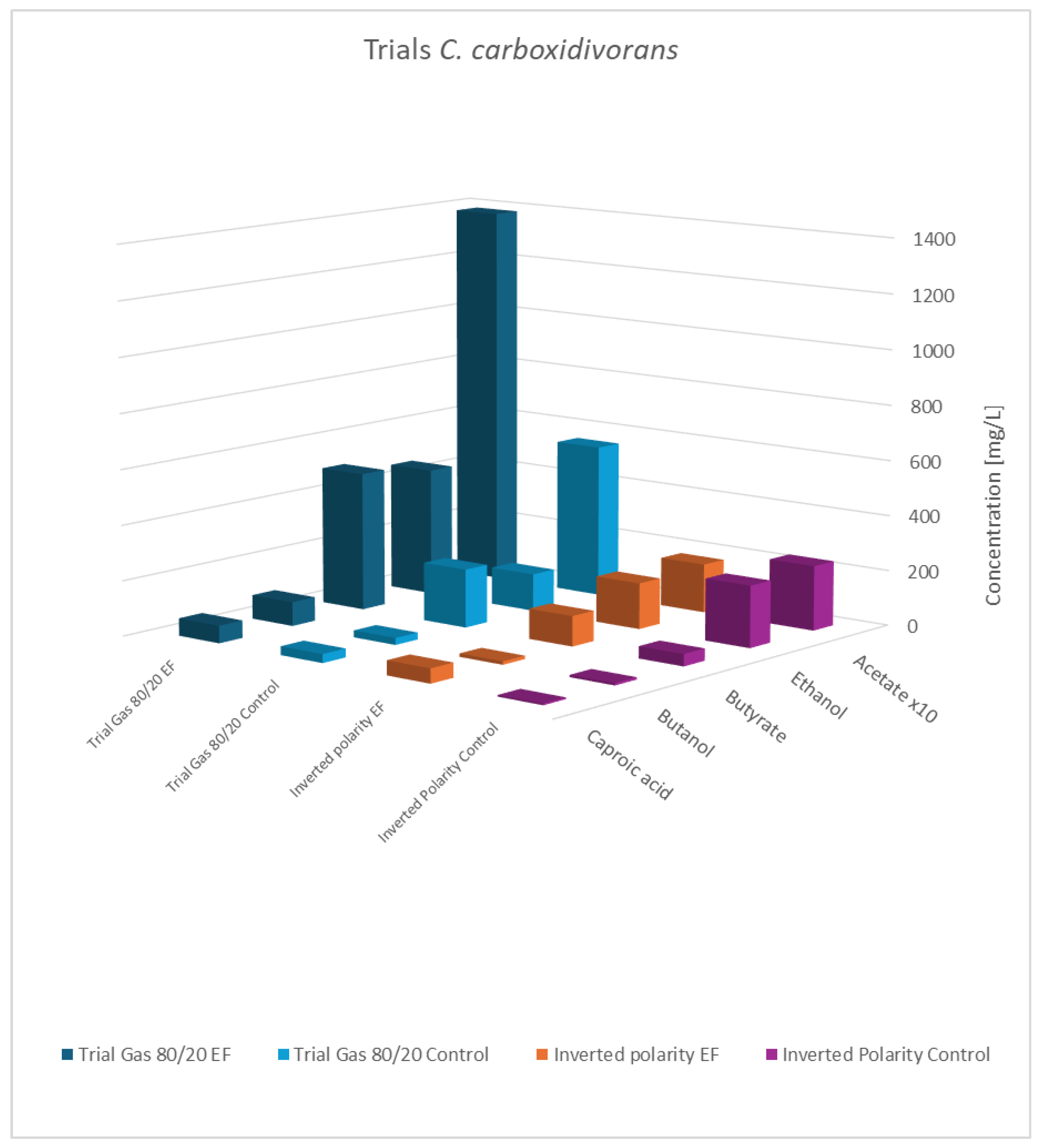
3.8. Verification Experiments
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Logan, B.E.; Rossi, R.; Ragab, A.; Saikaly, P.E. Electroactive microorganisms in bioelectrochemical systems. Nat. Rev. Microbiol. 2019, 17, 307–319. [Google Scholar] [CrossRef]
- Zhao, J.; Li, F.; Cao, Y.; Zhang, X.; Chen, T.; Song, H.; Wang, Z. Microbial extracellular electron transfer and strategies for engineering electroactive microorganisms. Biotechnol. Adv. 2021, 53, 107682. [Google Scholar] [CrossRef]
- Moscoviz, R.; Toledo-Alarcón, J.; Trably, E.; Bernet, N. Electro-Fermentation: How to Drive Fermentation Using Electrochemical Systems. Trends Biotechnol. 2016, 34, 856–865. [Google Scholar] [CrossRef]
- Liu, C.G.; Xue, C.; Lin, Y.H.; Bai, F.W. Redox potential control and applications in microaerobic and anaerobic fermentations. Biotechnol. Adv. 2013, 31, 257–265. [Google Scholar] [CrossRef]
- Fruehauf, H.M.; Enzmann, F.; Harnisch, F.; Ulber, R.; Holtmann, D. Microbial Electrosynthesis—An Inventory on Technology Readiness Level and Performance of Different Process Variants. Biotechnol. J. 2020, 15, 2000066. [Google Scholar] [CrossRef]
- Vassilev, I.; Hernandez, P.A.; Batlle-Vilanova, P.; Freguia, S.; Krömer, J.O.; Keller, J.; Ledezma, P.; Virdis, B. Microbial Electrosynthesis of Isobutyric, Butyric, Caproic Acids, and Corresponding Alcohols from Carbon Dioxide. ACS Sustain. Chem. Eng. 2018, 6, 8485–8493. [Google Scholar] [CrossRef]
- Maschio, G.; Lucchesi, A.; Stoppato, G. Production of syngas from biomass. Bioresour. Technol. 1994, 48, 119–126. [Google Scholar] [CrossRef]
- Schuchmann, K.; Müller, V. Autotrophy at the thermodynamic limit of life: A model for energy conservation in acetogenic bacteria. Nat. Rev. Microbiol. 2014, 12, 809–821. [Google Scholar] [CrossRef]
- Rachbauer, L.; Bochmann, G.; Fuchs, W. Gas fermentation. In The Autotrophic Biorefinery: Raw Materials from Biotechnology; Kourist, R., Schmidt, S., Eds.; De Gruyter: Berlin, Germany, 2021; pp. 85–112. [Google Scholar] [CrossRef]
- Devi, N.B.; Pugazhenthi, G.; Pakshirajan, K. Synthetic biology approaches and bioseparations in syngas fermentation. Trends Biotechnol. 2024, in press. [Google Scholar] [CrossRef]
- Bajracharya, S.; Krige, A.; Matsakas, L.; Rova, U.; Christakopoulos, P. Advances in cathode designs and reactor configurations of microbial electrosynthesis systems to facilitate gas electro-fermentation. Bioresour. Technol. 2022, 354, 127178. [Google Scholar] [CrossRef]
- Tharak, A.; Katakojwala, R.; Kajla, S.; Venkata Mohan, S. Chemolithoautotrophic reduction of CO2 to acetic acid in gas and gas-electro fermentation systems: Enrichment, microbial dynamics, and sustainability assessment. Chem. Eng. J. 2023, 454, 140200. [Google Scholar] [CrossRef]
- Kottenhahn, P.; Philipps, G.; Jennewein, S. Hexanol biosynthesis from syngas by Clostridium carboxidivorans P7—Product toxicity, temperature dependence and in situ extraction. Heliyon 2021, 7, e07732. [Google Scholar] [CrossRef]
- Koch, C.; Kuchenbuch, A.; Kracke, F.; Berhardt, P.; Krömer, J.; Harnisch, F. Predicting and experimental evaluating bio-electrochemical synthesis—A case study with Clostridium kluyveri. Bioelectrochemistry 2017, 118, 114–122. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, H.; Wu, P.; Li, J.; Zhang, J. Clostridium kluyveri enhances caproate production by synergistically cooperating with acetogens in mixed microbial community of electro-fermentation system. Bioresour. Technol. 2023, 369, 128436. [Google Scholar] [CrossRef] [PubMed]
- Rückel, A.; Hannemann, J.; Maierhofer, C.; Fuchs, A.; Weuster-Botz, D. Studies on Syngas Fermentation with Clostridium carboxidivorans in Stirred-Tank Reactors with Defined Gas Impurities. Front. Microbiol. 2021, 12, 655390. [Google Scholar] [CrossRef]
- Liou, J.S.C.; Balkwill, D.L.; Drake, G.R.; Tanner, R.S. Clostridium carboxidivorans sp. nov., a solvent-producing clostridium isolated from an agricultural settling lagoon, and reclassification of the acetogen Clostridium scatologenes strain SL1 as Clostridium drakei sp. nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 2085–2091. [Google Scholar] [CrossRef]
- Fernández-Naveira, Á.; Abubackar, H.N.; Veiga, M.C.; Kennes, C. Production of chemicals from C1 gases (CO, CO2) by Clostridium carboxidivorans. World J. Microbiol. Biotechnol. 2017, 33, 43. [Google Scholar] [CrossRef]
- Ukpong, M.N.; Atiyeh, H.K.; De Lorme, M.J.; Liu, K.; Zhu, X.; Tanner, R.S.; Wilkins, M.R.; Stevenson, B.S. Physiological response of Clostridium carboxidivorans during conversion of synthesis gas to solvents in a gas-fed bioreactor. Biotechnol. Bioeng. 2012, 109, 2720–2728. [Google Scholar] [CrossRef]
- Fernández-Naveira, Á.; Abubackar, H.N.; Veiga, M.C.; Kennes, C. Efficient butanol-ethanol (B-E) production from carbon monoxide fermentation by Clostridium carboxidivorans. Appl. Microbiol. Biotechnol. 2016, 100, 3361–3370. [Google Scholar] [CrossRef]
- Allen, T.D.; Caldwell, M.E.; Lawson, P.A.; Huhnke, R.L.; Tanner, R.S. Alkalibaculum bacchi gen. nov., sp. nov., a CO-oxidizing, ethanol-producing acetogen isolated from livestock-impacted soil. Int. J. Syst. Evol. Microbiol. 2010, 60, 2483–2489. [Google Scholar] [CrossRef]
- Barker, H.A.; Taha, S.M. Clostridium kluyverii, an Organism Concerned in the Formation of Caproic Acid from Ethyl Alcohol. J. Bacteriol. 1942, 43, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Berillo, D.; Al-Jwaid, A.; Caplin, J. Polymeric materials used for immobilisation of bacteria for the bioremediation of contaminants in water. Polymers 2021, 13, 1073. [Google Scholar] [CrossRef]
- Mur-Gorgas, A.; Martínez-Pellitero, S.; Joglar, T.; Escapa, A.; Mateos, R. 3D-Printed Conductive Polymers as Alternative for Bioelectrochemical Systems Electrodes: Abiotic Study and Biotic Start-Up. Appl. Sci. 2024, 14, 7199. [Google Scholar] [CrossRef]
- Pu, K.B.; Bai, J.R.; Chen, Q.Y.; Wang, Y.H. Modified Stainless Steel as Anode Materials in Bioelectrochemical Systems. ACS Symp. Ser. 2020, 1342, 165–184. [Google Scholar]
- Cheng, C.; Shao, Y.; Li, W.; Liu, J.; Liu, X.; Zhao, Y.; Li, X.; Yang, S.-T.; Xue, C. Electricity-enhanced anaerobic, non-photosynthetic mixotrophy by Clostridium carboxidivorans with increased carbon efficiency and alcohol production. Energy Convers. Manag. 2022, 252, 115118. [Google Scholar] [CrossRef]
- San-Valero, P.; Abubackar, H.N.; Veiga, M.C.; Kennes, C. Effect of pH, yeast extract and inorganic carbon on chain elongation for hexanoic acid production. Bioresour. Technol. 2020, 300, 122659. [Google Scholar] [CrossRef]
- Candry, P.; Radić, L.; Favere, J.; Carvajal-Arroyo, J.M.; Rabaey, K.; Ganigué, R. Mildly acidic pH selects for chain elongation to caproic acid over alternative pathways during lactic acid fermentation. Water Res. 2020, 186, 100284. [Google Scholar] [CrossRef]
- Ganigué, R.; Sánchez-Paredes, P.; Bañeras, L.; Colprim, J. Low fermentation pH is a trigger to alcohol production, but a killer to chain elongation. Front. Microbiol. 2016, 7, 702. [Google Scholar] [CrossRef]
- Patil, S.A.; Arends, J.B.A.; Vanwonterghem, I.; van Meerbergen, J.; Guo, K.; Tyson, G.W.; Rabaey, K. Selective Enrichment Establishes a Stable Performing Community for Microbial Electrosynthesis of Acetate from CO2. Environ. Sci. Technol. 2015, 49, 8833–8843. [Google Scholar] [CrossRef]
- Rovira-Alsina, L.; Balaguer, M.D.; Puig, S. Thermophilic bio-electro carbon dioxide recycling harnessing renewable energy surplus. Bioresour. Technol. 2021, 321, 122659. [Google Scholar] [CrossRef] [PubMed]
- Cardeña, R.; Valencia-Ojeda, C.; Chazaro-Ruiz, L.F.; Razo-Flores, E. Regulation of the dark fermentation products by electro-fermentation in reactors without membrane. Int. J. Hydrogen Energy 2024, 49, 107–116. [Google Scholar] [CrossRef]
- Batlle-Vilanova, P.; Ganigué, R.; Ramió-Pujol, S.; Bañeras, L.; Jiménez, G.; Hidalgo, M.; Balaguer, M.D.; Colprim, J.; Puig, S. Microbial electrosynthesis of butyrate from carbon dioxide: Production and extraction. Bioelectrochemistry 2017, 117, 57–64. [Google Scholar] [CrossRef]
- Fontmorina, J.M.; Izadi, P.; Rasul, S.; Yu, E.H. Carbon dioxide utilisation by bioelectrochemical systems through microbial electrochemical synthesis. Carbon Dioxide Util. Transform. 2019, 2, 561–581. [Google Scholar] [CrossRef]
- Jensen, W.B. Faraday’s Laws or Faraday’s Law? J. Chem. Educ. 2012, 89, 1208–1209. [Google Scholar] [CrossRef]
- Kracke, F.; Virdis, B.; Bernhardt, P.V.; Rabaey, K.; Krömer, J.O. Redox dependent metabolic shift in Clostridium autoethanogenum by extracellular electron supply. Biotechnol. Biofuels 2016, 9, 1–12. [Google Scholar] [CrossRef]
- Choi, O.; Kim, T.; Woo, H.M.; Um, Y. Electricity-driven metabolic shift through direct electron uptake by electroactive heterotroph Clostridium pasteurianum. Sci. Rep. 2014, 4, 5200. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Zhang, J.-W.; Li, F.; Zhao, X.-Q.; Bai, F.-W.; Song, H.; Liu, C.-G. Electrochemical Control of Cell Metabolism Improves Ethanol Production of Zymomonas mobilis in an Electro-Fermentation System. ACS Sustain. Chem. Eng. 2023, 11, 2364–2374. [Google Scholar] [CrossRef]
- Smith, E.T.; Tomich, J.M.; Iwamoto, T.; Richards, J.H.; Mao, Y.; Feinberg, B.A. A Totally Synthetic Histidine-2 Ferredoxin: Thermal Stability and Redox Properties. Biochemistry 1991, 30, 11669–11676. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.S.; Kim, B.H. Electron flow shift in Clostridium acetobutylicum fermentation by electrochemically introduced reducing equivalent. Biotechnol. Lett. 1988, 10, 123–128. [Google Scholar] [CrossRef]
- Li, H.; Lee, K.; Zhang, J. Electrocatalytic H2 oxidation reaction. PEM Fuel Cell Electrocatal. Catal. Layers Fundam. Appl. 2008, 135–164. [Google Scholar] [CrossRef]
- Kumar, P.; Chandrasekhar, K.; Kumari, A.; Sathiyamoorthi, E.; Kim, B.S. Electro-fermentation in aid of bioenergy and biopolymers. Energies 2018, 11, 356. [Google Scholar] [CrossRef]
- Engel, M.; Holtmann, D.; Ulber, R.; Tippkötter, N. Increased Biobutanol Production by Mediator-Less Electro-Fermentation. Biotechnol. J. 2019, 14, 1–10. [Google Scholar] [CrossRef]
- Nailwal, V.; Mukherjee, T.; Venkata Mohan, S. Decoupling acidogenic cascade via bio-electrogenic induced ABE pathway for enhanced n-butanol synthesis in Clostridium acetobutylicum ATCC-824: Bioenergetics and gene expression analysis. Chem. Eng. J. 2024, 486, 123456. [Google Scholar] [CrossRef]
- Ren, W.; Wu, Q.; Deng, L.; Wang, H.; Zhang, Y.; Guo, W. Insights into Chain Elongation Mechanisms of Weak Electric-Field-Stimulated Continuous Caproate Biosynthesis: Key Enzymes, Specific Species Functions, and Microbial Collaboration. ACS ES T Eng. 2023, 3, 1649–1660. [Google Scholar] [CrossRef]
- Arends, J.B.A.; Patil, S.A.; Roume, H.; Rabaey, K. Continuous long-term electricity-driven bioproduction of carboxylates and isopropanol from CO2 with a mixed microbial community. J. CO2 Util. 2017, 20, 141–149. [Google Scholar] [CrossRef]
- Wenzel, J.; Fiset, E.; Batlle-Vilanova, P.; Cabezas, A.; Etchebehere, C.; Balaguer, M.D.; Colprim, J.; Puig, S. Microbial community pathways for the production of volatile fatty acids from CO2 and electricity. Front. Energy Res. 2018, 6, 15. [Google Scholar] [CrossRef]
- Jourdin, L.; Winkelhorst, M.; Rawls, B.; Buisman, C.J.N.; Strik, D.P.B.T.B. Enhanced selectivity to butyrate and caproate above acetate in continuous bioelectrochemical chain elongation from CO2: Steering with CO2 loading rate and hydraulic retention time. Bioresour. Technol. Reports 2019, 7, 100284. [Google Scholar] [CrossRef]
- Romans-Casas, M.; Blasco-Gómez, R.; Colprim, J.; Balaguer, M.D.; Puig, S. Bio-electro CO2 recycling platform based on two separated steps. J. Environ. Chem. Eng. 2021, 9, 105189. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Concentration [g/L] |
|---|---|
| KH2PO4 | 1.000 |
| NaCl | 1.000 |
| NH4Cl | 1.000 |
| MgSO4·7H2O | 0.211 |
| K2CO3 | 1.000 |
| CaCl2·2H2O | 0.040 |
| Resazurin (1% w/v) | 1 mL/L |
| [mL/L] | |
| Wolin’s trace metal solution | 10 |
| Wolfe’s vitamin solution | 10 |
| Na2S·9 H2O | 0.5 g/L |
| pH = 6.3 |
| Species | Concentration [g/L] |
|---|---|
| Nitrilotriacetic acid | 1.5 |
| MgSO4·7H2O | 3 |
| MnSO4·H2O | 0.5 |
| NaCl | 1 |
| FeSO4·7H2O | 0.1 |
| CoSO4·7H2O | 0.18 |
| CaCl2·2H2O | 0.1 |
| ZnSO4·7H2O | 0.18 |
| CuSO4·5H2O | 0.01 |
| KAl(SO4)2·12H2O | 0.02 |
| H3BO3 | 0.01 |
| Na2MoO4·2H2O | 0.01 |
| NiCl2·6H2O | 0.03 |
| Na2SeO3·5H2O | 0.0003 |
| Na2WO4·2H2O | 0.0004 |
| Species | Concentration [mg/L] |
|---|---|
| Biotin | 2 |
| Folic acid | 2 |
| Pyridoxine-HCl | 10 |
| Thiamine-HCl | 5 |
| Riboflavin | 5 |
| Nicotinic acid | 5 |
| Ca-D-pantothenate | 5 |
| Vitamin B12 | 0.1 |
| p-Aminobenzoic acid | 5 |
| (±)-α-Lipoic acid | 5 |
| Acetate [C mmol/L] | Butyrate [C mmol/L] | Caproate [C mmol/L] | Ethanol [C mmol/L] | n-Butanol [C mmol/L] | ||
|---|---|---|---|---|---|---|
| C. carboxidivorans | EF | 128 | 29 | 9 | 16 | 3 |
| control | 86 | 10 | 7 | 4 | 2 | |
| A. bacchi | EF | 161 | 35 | 22 | 5 | 2 |
| control | 116 | 3 | 0 | 3 | 0 | |
| Co-culture | EF | 148 | 72 | 47 | 8 | 6 |
| control | 133 | 32 | 29 | 8 | 4 |
| Acetate [C mmol/(L·d)] | Ethanol [C mmol/(L·d)] | Butyrate [C mmol/(L·d)] | n-Butanol [C mmol/L·d)] | Caproate [C mmol/(L·d)] | ||
|---|---|---|---|---|---|---|
| C. carboxidivorans | EF | 2.622 | −0.576 | 0.598 | 0.035 | 0.279 |
| control | 1.760 | 0.063 | 0.195 | 0.052 | 0.034 | |
| A. bacchi | EF | 3.292 | −0.647 | 0.708 | 0.124 | 0.444 |
| control | 2.367 | 0.072 | 0.053 | 0.000 | 0.000 | |
| Co-culture | EF | 3.029 | −1.420 | 1.467 | 0.128 | 0.951 |
| control | 2.708 | 0.075 | 0.646 | 0.075 | 0.586 |
| Substrate | Reactor Type | Hac [mM C] | HBu [mM C] | HCa [mM C] | EtOH [mM C] | BuOH [mM C] | Current Density [mA/m2] | Applied Cathode Potential [mV] | Cell Potential [V] | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| CO2 | H-Cell | 3.74 | 0.6 | n.r. | 1.24 | - | - | −574 vs. SHE | - | [47] |
| CO2 | Tilted trickle bed | 34.7 | 87.5 | - | 3.4 (final conc) | 5.4 | 2.74 | −800 vs. SHE | - | [33] |
| N2/CO2 | Serum flask | 200 | 420 | 160 | n.r. | n.r. | −126,000 | 1.02 | - | [48] |
| H2/CO2 80/20 | Tilted trickle bed | 28.98 | 27.85 | 148.26 | 46.88 | 550 | −800 vs. SHE | 9.82 | [49] | |
| N2/H2/CO2 50/40/10 | Serum flask | 160 | 35 | 22 | 0 | 0 | 0.22 | −500 | 0.8 | * |
| H2/CO2 80/20 | Serum flask | 216 | 14 | 4.2 | 25 | 7.6 | 0.25 | −500 | 0.8 | * |
| Acetate [C mmol/(L·d)] | Ethanol [C mmol/(L·d)] | Butyrate [C mmol/(L·d)] | n-Butanol [C mmol/L·d)] | Caproate [C mmol/(L·d)] | |||
|---|---|---|---|---|---|---|---|
| Reversed polarity | C. carboxidivorans | EF | 1.052 | 0.141 | 0.093 | 0.013 | 0.056 |
| control | 1.397 | 0.174 | 0.035 | 0.010 | 0.008 | ||
| A. bacchi | EF | 1.499 | 0.126 | 0.202 | 0.031 | 0.077 | |
| control | 1.282 | 0.074 | 0.195 | 0.010 | 0.072 | ||
| 80:20 = H2:CO2 | C. carboxidivorans | EF | 9.055 | 0.428 | 0.412 | 0.060 | 0.150 |
| control | 3.622 | 0.180 | 0.120 | 0.026 | 0.031 | ||
| A. bacchi | EF | 5.651 | 0.881 | 0.509 | 0.149 | 0.175 | |
| control | 1.923 | 0.261 | 0.133 | 0.068 | 0.072 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hiebl, C.; Fuchs, W. Electro-Enhanced Gas Fermentation for Bioproduction of Volatile Fatty Acids and Alcohols. Microorganisms 2025, 13, 249. https://doi.org/10.3390/microorganisms13020249
Hiebl C, Fuchs W. Electro-Enhanced Gas Fermentation for Bioproduction of Volatile Fatty Acids and Alcohols. Microorganisms. 2025; 13(2):249. https://doi.org/10.3390/microorganisms13020249
Chicago/Turabian StyleHiebl, Clemens, and Werner Fuchs. 2025. "Electro-Enhanced Gas Fermentation for Bioproduction of Volatile Fatty Acids and Alcohols" Microorganisms 13, no. 2: 249. https://doi.org/10.3390/microorganisms13020249
APA StyleHiebl, C., & Fuchs, W. (2025). Electro-Enhanced Gas Fermentation for Bioproduction of Volatile Fatty Acids and Alcohols. Microorganisms, 13(2), 249. https://doi.org/10.3390/microorganisms13020249