
NcSWP8, a New Spore Wall Protein, Interacts with Polar Tube Proteins in the Parasitic Microsporidia Vairimorpha (Nosema) ceranae

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Sequence Analysis
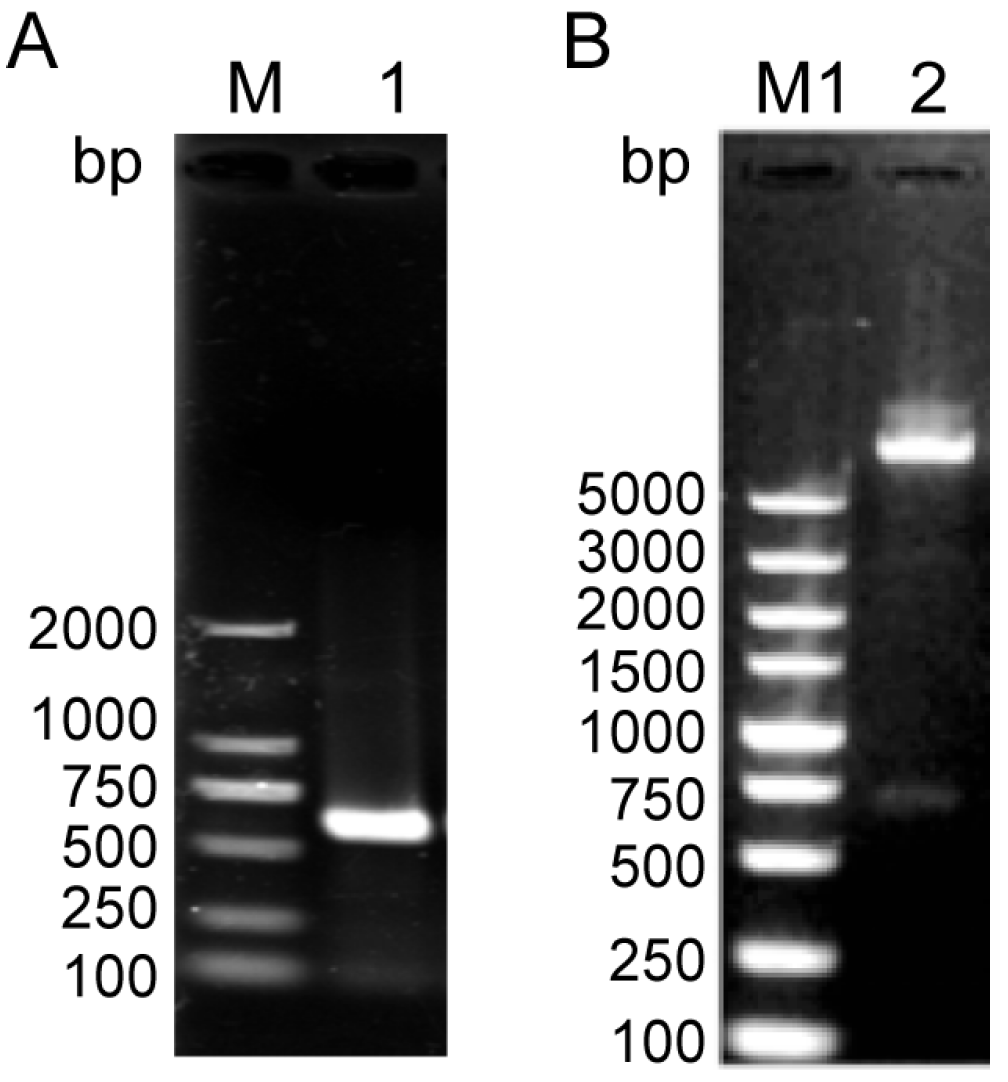
2.2. RNA Extraction, Polymerase Chain Reaction (PCR), Semi-Quantitative Reverse Transcription Polymerase Chain Reaction (sqRT-PCR)
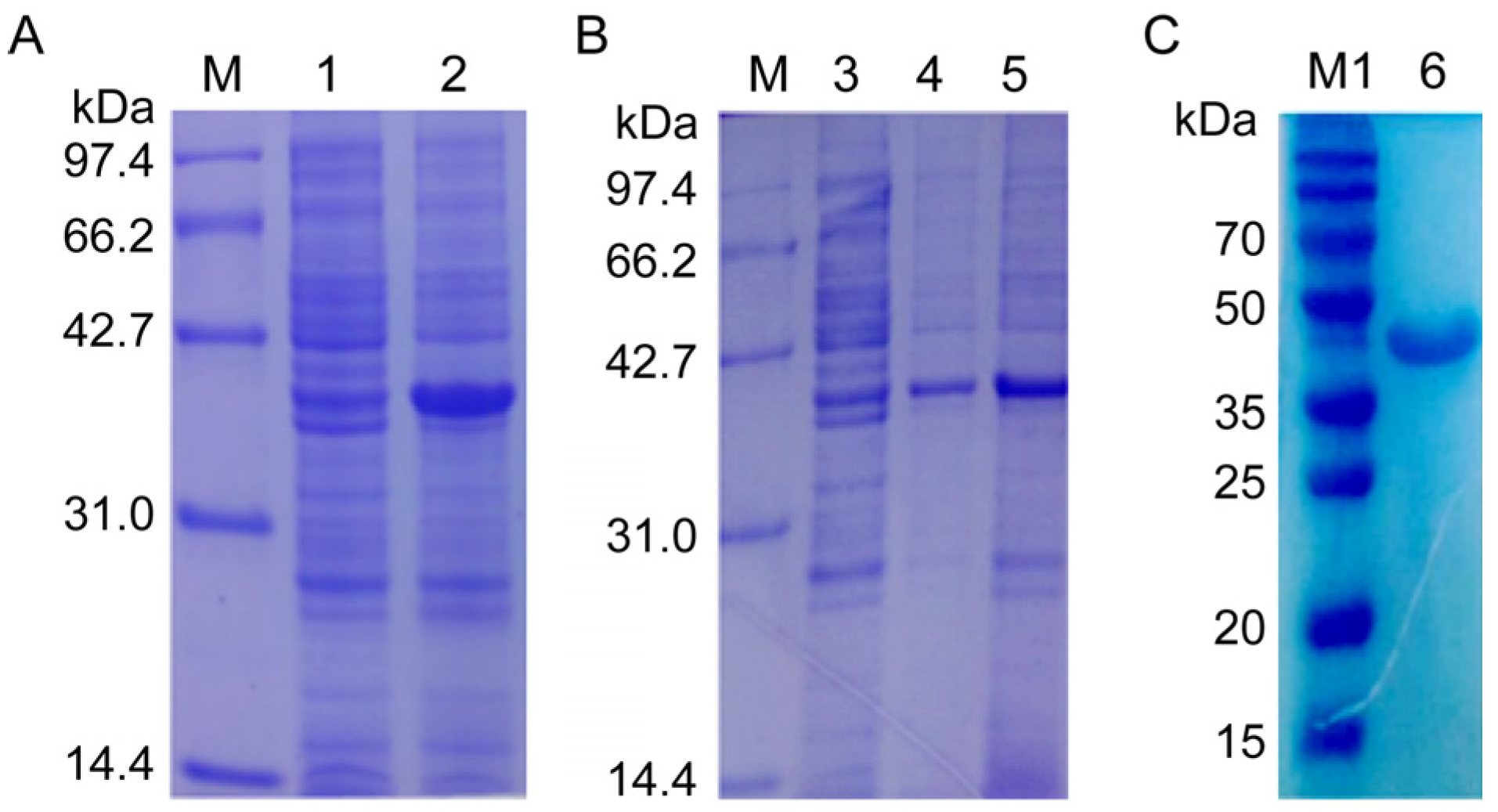
2.3. Cloning, Expression, Purification of Recombinant NcSWP8, and Preparation of Anti-NcSWP8 Antiserum
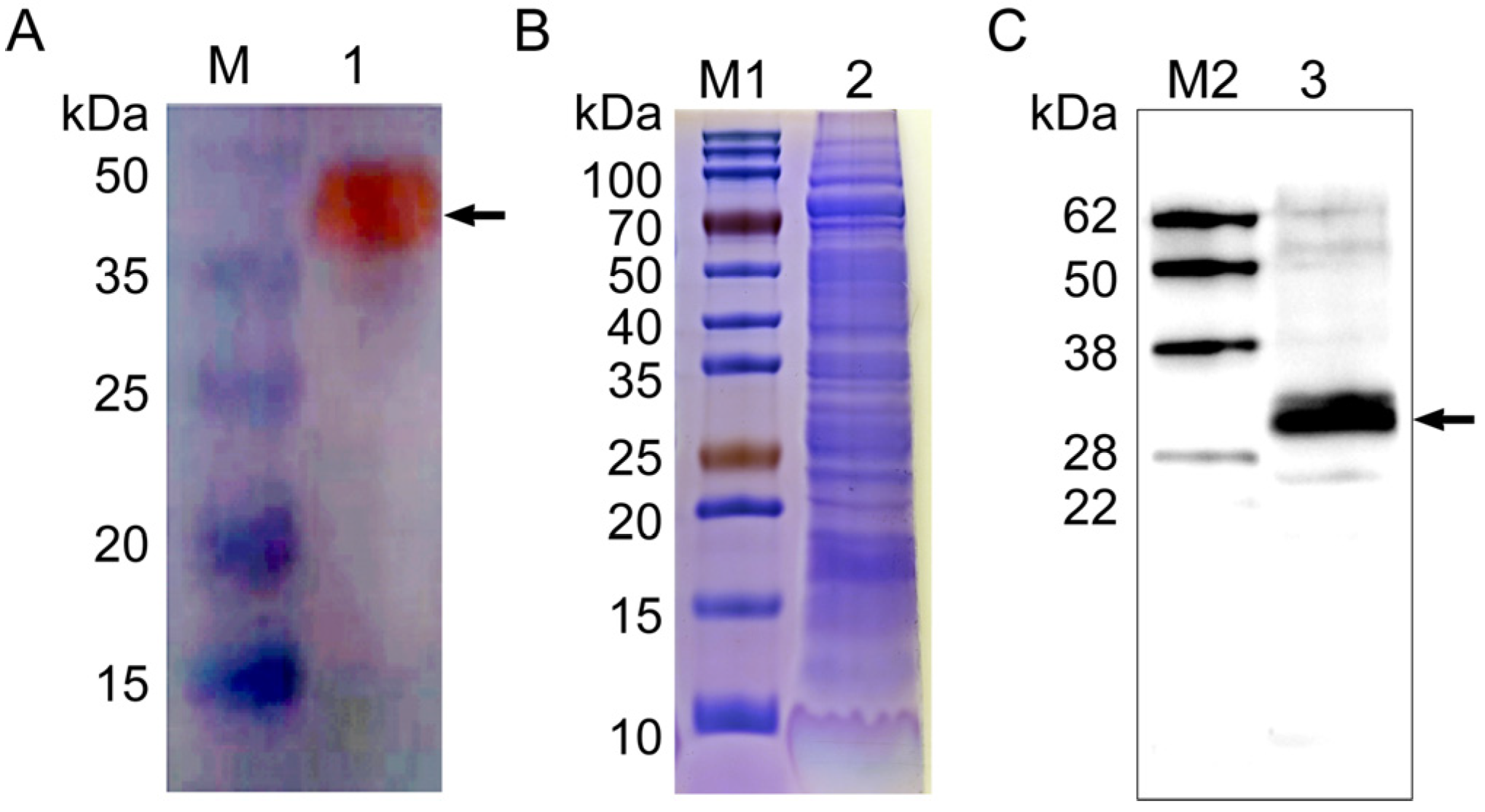
2.4. Specificity Analysis of Polyclonal Antibody Anti-NcSWP8
2.5. Subcellular Localization Analysis of NcSWP8
2.5.1. Immunofluorescence Analysis (IFA)
2.5.2. Immunoelectron Microscopy Analysis (IEM)
2.5.3. Immunohistochemical Analysis
2.6. Biological Function Research of NcSWP8
2.6.1. Co-Immunoprecipitation (Co-IP)
2.6.2. Far-Western Blotting
2.6.3. Antibody Blocking Assay
3. Results
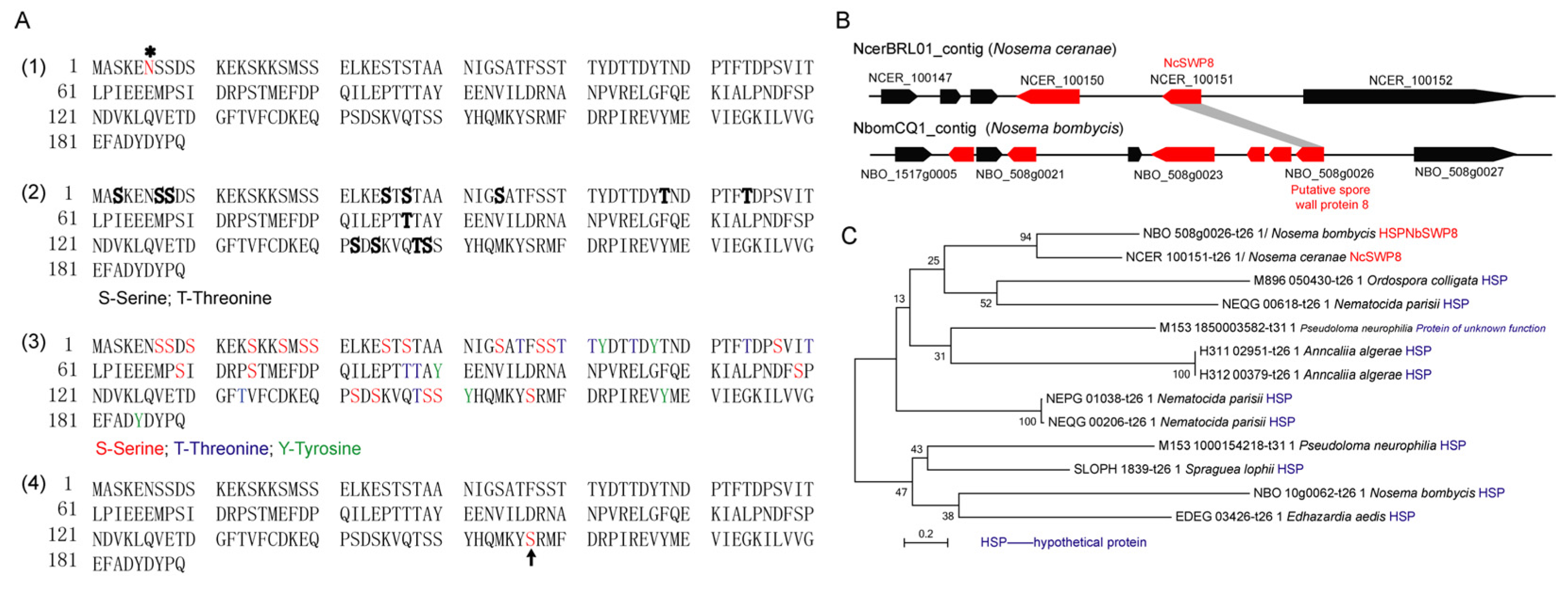
3.1. Characterization of the Gene Encoding NcSWP8
3.2. Expression and Detection of NcSWP8
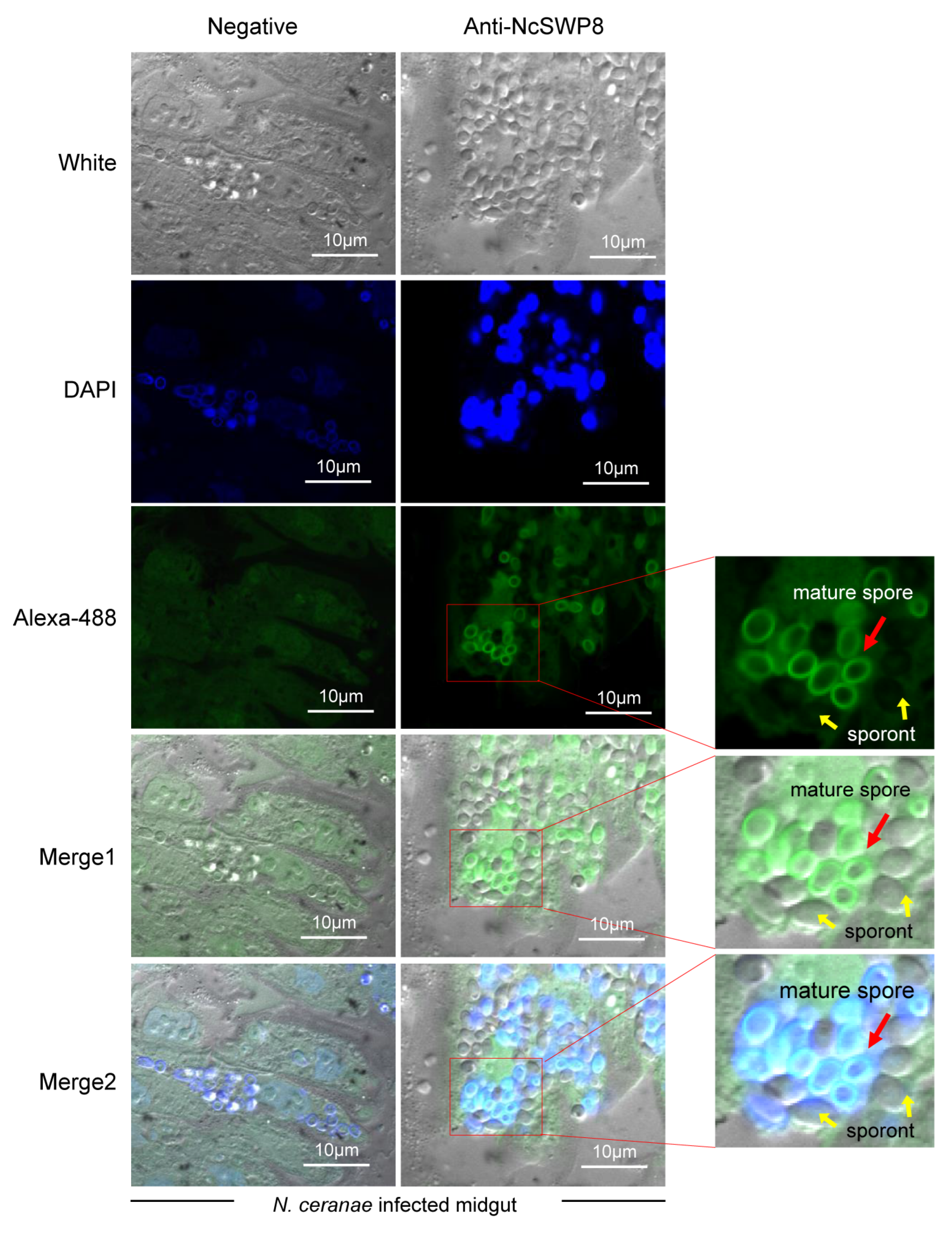
3.3. Subcellular Localization of NcSWP8 in V. ceranae
3.4. Biological Functions of NcSWP8
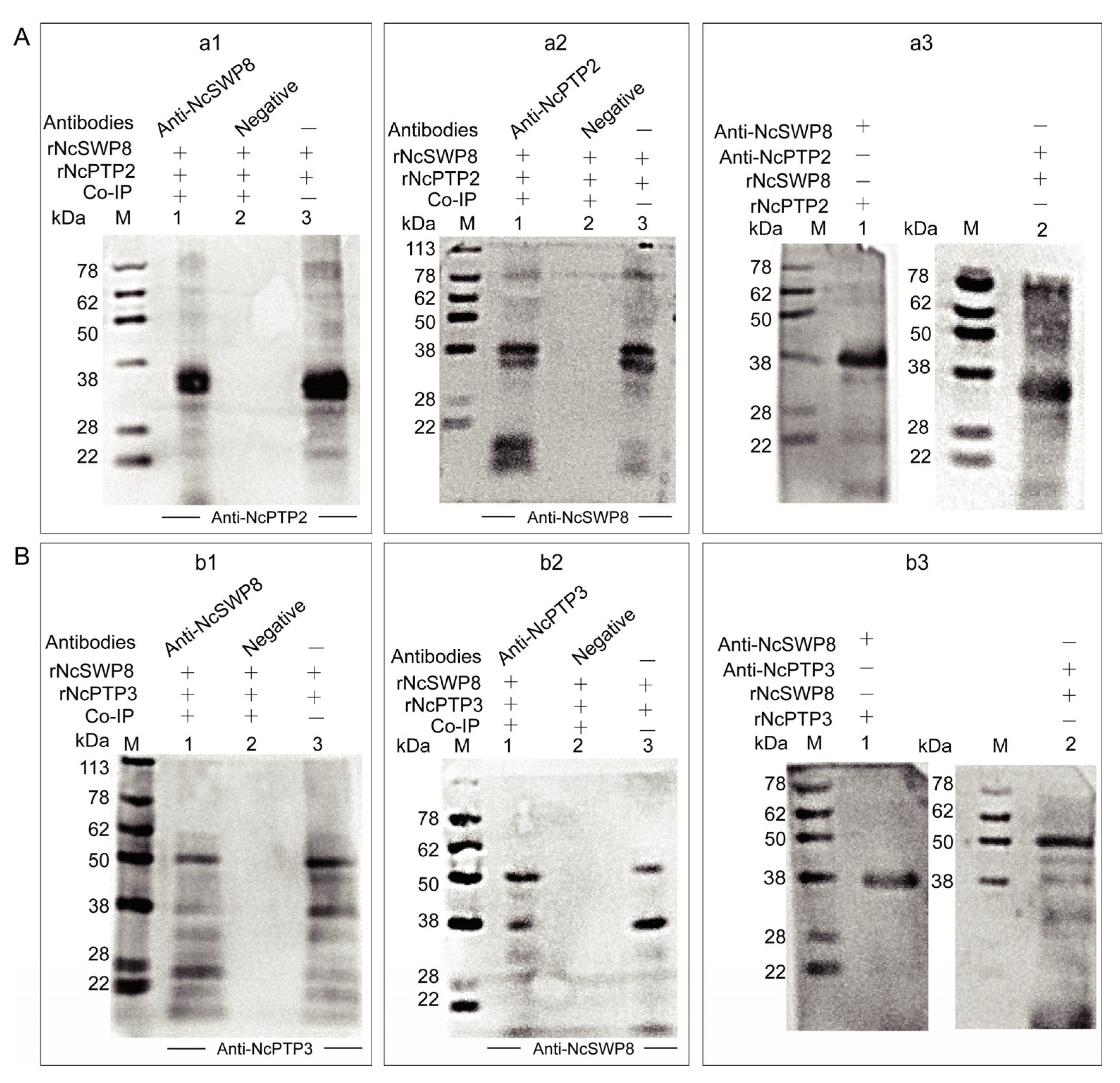
3.4.1. Interaction Between NcSWP8 and Polar Tube Proteins
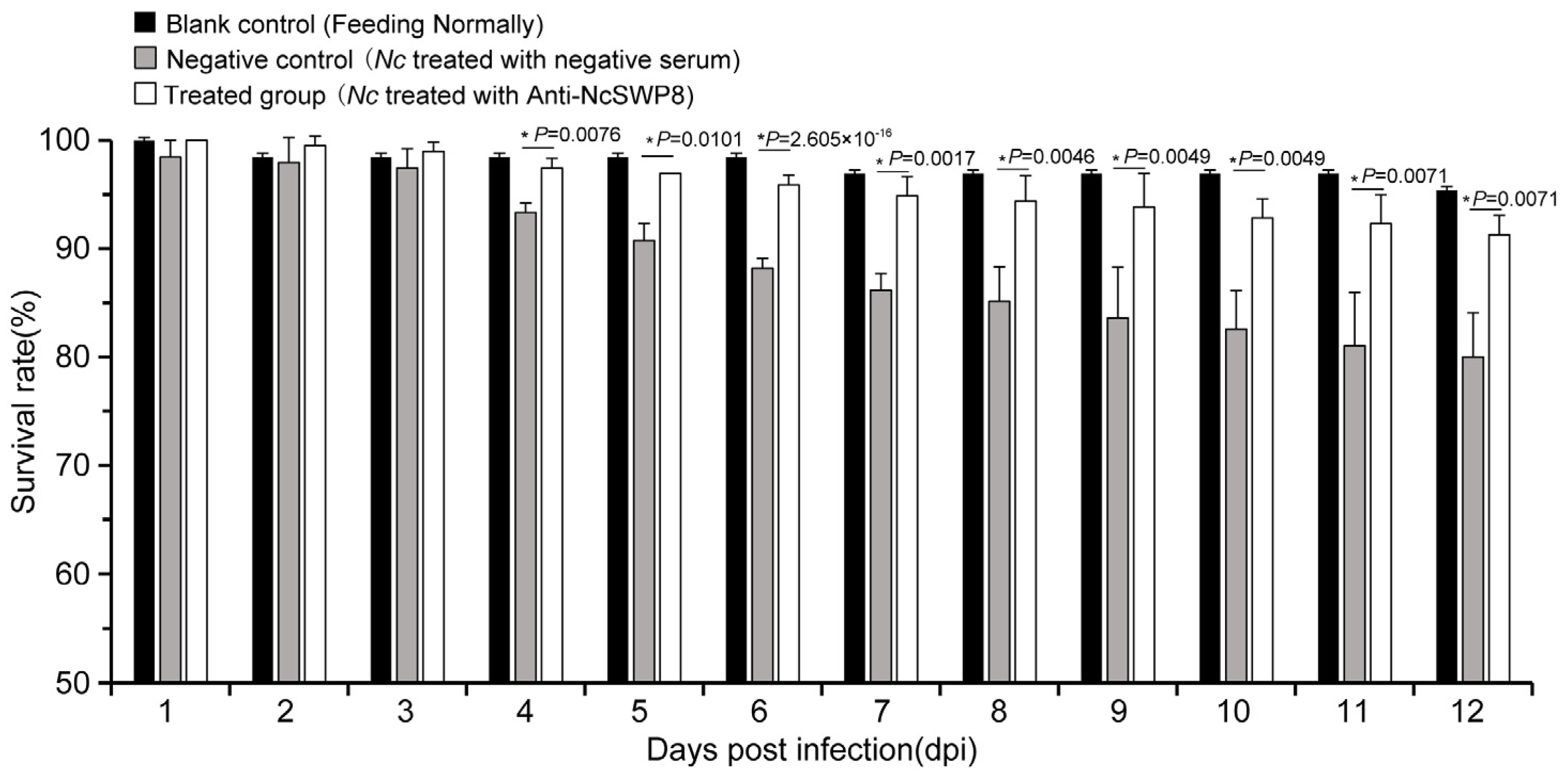
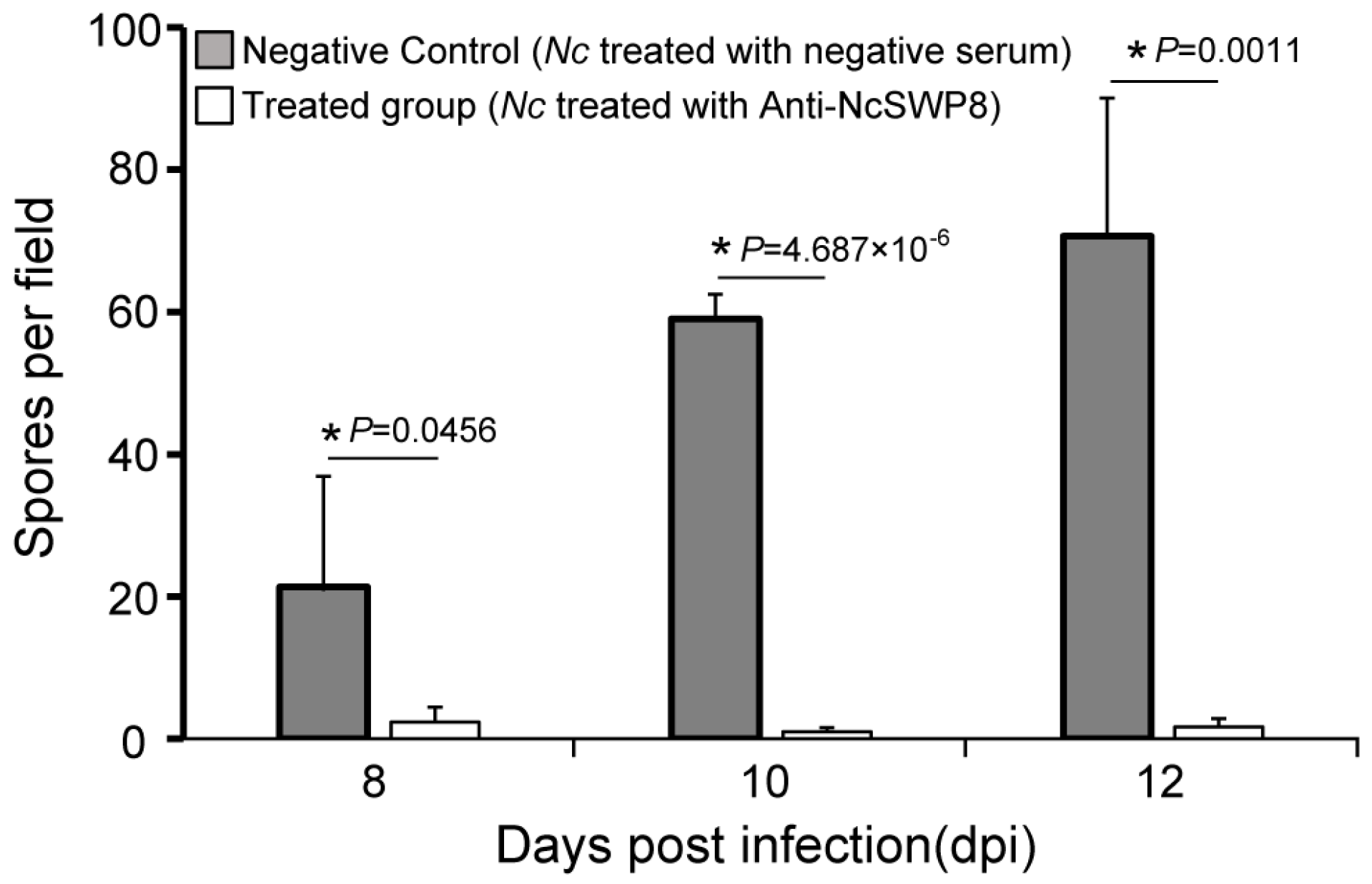
3.4.2. Effect of Anti-NcSWP8 Serum on Infection Ability
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Snowden, K. Zoonotic microsporidia from animals and arthropods with a discussion of human infections. In Opportunistic Infections: Toxoplasma, Sarcocystis, and Microsporidia; Springer: Boston, MA, USA, 2004; pp. 123–134. [Google Scholar]
- Fries, I. Nosema apis—A parasite in the honey bee colony. Bee World 1993, 74, 5–19. [Google Scholar] [CrossRef]
- Fries, I.; Feng, F.; da Silva, A.; Slemenda, S.B.; Pieniazek, N.J. Nosema ceranae n. sp. (Microspora, Nosematidae), morphological and molecular characterization of a microsporidian parasite of the Asian honey bee Apis cerana (Hymenoptera, Apidae). Eur. J. Protistol. 1996, 32, 356–365. [Google Scholar] [CrossRef]
- Chen, Y.P.; Huang, Z.Y. Nosema ceranae, a newly identified pathogen of Apis mellifera in the USA and Asia. Apidologie 2010, 41, 364–374. [Google Scholar] [CrossRef]
- Higes, M.; García-Palencia, P.; Martín-Hernández, R.; Meana, A. Experimental infection of Apis mellifera honeybees with Nosema ceranae (Microsporidia). J. Invertebr. Pathol. 2007, 94, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Higes, M.; Martín-Hernández, R.; Botías, C.; Bailón, E.G.; González-Porto, A.V.; Barrios, L.; Del Nozal, M.J.; Bernal, J.L.; Jiménez, J.J.; Palencia, P.G. How natural infection by Nosema ceranae causes honeybee colony collapse. Environ. Microbiol. 2008, 10, 2659–2669. [Google Scholar] [CrossRef]
- Katznelson, H.; Jamieson, C. Control of nosema disease of honeybees with fumagillin. Science 1952, 115, 70–71. [Google Scholar] [CrossRef]
- Mendoza, Y.; Diaz-Cetti, S.; Ramallo, G.; Santos, E.; Porrini, M.; Invernizzi, C. Nosema ceranae winter control: Study of the effectiveness of different fumagillin treatments and consequences on the strength of honey bee (Hymenoptera: Apidae) colonies. J. Econ. Entomol. 2017, 110, 1–5. [Google Scholar]
- Glavinic, U.; Stevanovic, J.; Ristanic, M.; Rajkovic, M.; Davitkov, D.; Lakic, N.; Stanimirovic, Z. Potential of fumagillin and Agaricus blazei mushroom extract to reduce Nosema ceranae in honey bees. Insects 2021, 12, 282. [Google Scholar] [CrossRef]
- Williams, G.R.; Sampson, M.A.; Shutler, D.; Rogers, R.E. Does fumagillin control the recently detected invasive parasite Nosema ceranae in western honey bees (Apis mellifera)? J. Invertebr. Pathol. 2008, 99, 342–344. [Google Scholar] [CrossRef]
- Burnham, A.J. Scientific advances in controlling Nosema ceranae (Microsporidia) infections in honey bees (Apis mellifera). Front. Vet. Sci. 2019, 6, 79. [Google Scholar] [CrossRef]
- Shumkova, R.; Balkanska, R.; Hristov, P. The herbal supplements NOZEMAT HERB® and NOZEMAT HERB PLUS®: An alternative therapy for N. ceranae infection and its effects on honey bee strength and production traits. Pathogens 2021, 10, 234. [Google Scholar] [CrossRef] [PubMed]
- Vávra, J.; Ronny Larsson, J. Structure of microsporidia. In Microsporidia: Pathogens Of Opportunity; Wiley Publisher: Hoboken, NJ, USA, 2014; pp. 1–70. [Google Scholar]
- Xu, Y.; Takvorian, P.; Cali, A.; Wang, F.; Zhang, H.; Orr, G.; Weiss, L.M. Identification of a new spore wall protein from Encephalitozoon cuniculi. Infect. Immun. 2006, 74, 239–247. [Google Scholar] [CrossRef] [PubMed]
- UM, S.; Karunakar, P.; Krishnamurthy, V. Homology modeling, virtual screening and dynamics study of proteins involved in Pebrine-Serine protease inhibitor 106 and spore wall protein 26. J. Biomol. Struct. Dyn. 2020, 38, 5148–5158. [Google Scholar]
- Southern, T.R.; Jolly, C.E.; Lester, M.E.; Hayman, J.R. EnP1, a microsporidian spore wall protein that enables spores to adhere to and infect host cells in vitro. Eukaryot. Cell 2007, 6, 1354–1362. [Google Scholar] [CrossRef] [PubMed]
- Hayman, J.R.; Southern, T.R.; Nash, T.E. Role of sulfated glycans in adherence of the microsporidian Encephalitozoon intestinalis to host cells in vitro. Infect. Immun. 2005, 73, 841–848. [Google Scholar] [CrossRef]
- Ao, T.Y.; Wang, J.L.; Ma, Z.G.; Zhou, Z.Y. Identification and biological functions of a novel spore wall protein NcER_100148 from Nosema cerana. Acta Entomol. Sin. 2023, 6, 747–758. [Google Scholar]
- Yang, D.; Pan, L.; Chen, Z.; Du, H.; Luo, B.; Luo, J.; Pan, G. The roles of microsporidia spore wall proteins in the spore wall formation and polar tube anchorage to spore wall during development and infection processes. Exp. Parasitol. 2018, 187, 93–100. [Google Scholar] [CrossRef]
- Polonais, V.; Mazet, M.; Wawrzyniak, I.; Texier, C.; Blot, N.; El Alaoui, H.; Delbac, F. The human microsporidian Encephalitozoon hellem synthesizes two spore wall polymorphic proteins useful for epidemiological studies. Infect. Immun. 2010, 78, 2221–2230. [Google Scholar] [CrossRef]
- Peuvel-Fanget, I.; Polonais, V.; Brosson, D.; Texier, C.; Kuhn, L.; Peyret, P.; Vivarès, C.; Delbac, F. EnP1 and EnP2, two proteins associated with the Encephalitozoon cuniculi endospore, the chitin-rich inner layer of the microsporidian spore wall. Int. J. Parasitol. 2006, 36, 309–318. [Google Scholar] [CrossRef]
- Hayman, J.R.; Hayes, S.F.; Amon, J.; Nash, T.E. Developmental expression of two spore wall proteins during maturation of the microsporidian Encephalitozoon intestinalis. Infect. Immun. 2001, 69, 7057–7066. [Google Scholar] [CrossRef]
- Brosson, D.; Kuhn, L.; Prensier, G.; Vivarès, C.P.; Texier, C. The putative chitin deacetylase of Encephalitozoon cuniculi: A surface protein implicated in microsporidian spore-wall formation. FEMS Microbiol. Lett. 2005, 247, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Bohne, W.; Ferguson, D.J.; Kohler, K.; Gross, U. Developmental expression of a tandemly repeated, glycine-and serine-rich spore wall protein in the microsporidian pathogen Encephalitozoon cuniculi. Infect. Immun. 2000, 68, 2268–2275. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Pan, L.; Peng, P.; Dang, X.; Li, C.; Li, T.; Long, M.; Chen, J.; Wu, Y.; Du, H. Interaction between SWP9 and polar tube proteins of the microsporidian Nosema bombycis and function of SWP9 as a scaffolding protein contribute to polar tube tethering to the spore wall. Infect. Immun. 2017, 85, 10–1128. [Google Scholar] [CrossRef]
- Yang, D.; Pan, G.; Dang, X.; Shi, Y.; Li, C.; Peng, P.; Luo, B.; Bian, M.; Song, Y.; Ma, C. Interaction and assembly of two novel proteins in the spore wall of the microsporidian species Nosema bombycis and their roles in adherence to and infection of host cells. Infect. Immun. 2015, 83, 1715–1731. [Google Scholar] [CrossRef]
- Yang, D.; Dang, X.; Peng, P.; Long, M.; Ma, C.; Qin, J.J.G.; Wu, H.; Liu, T.; Zhou, X.; Pan, G. NbHSWP11, a microsporidia Nosema bombycis protein, localizing in the spore wall and membranes, reduces spore adherence to host cell BME. J. Parasitol. 2014, 100, 623–632. [Google Scholar] [CrossRef]
- Wu, Z.; Li, Y.; Pan, G.; Zhou, Z.; Xiang, Z. SWP25, a novel protein associated with the Nosema bombycis endospore 1. J. Eukaryot. Microbiol. 2009, 56, 113–118. [Google Scholar] [CrossRef]
- Wu, Z.; Li, Y.; Pan, G.; Tan, X.; Hu, J.; Zhou, Z.; Xiang, Z. Proteomic analysis of spore wall proteins and identification of two spore wall proteins from Nosema bombycis (Microsporidia). Proteomics 2008, 8, 2447–2461. [Google Scholar] [CrossRef]
- Wang, Y.; Geng, H.; Dang, X.; Xiang, H.; Li, T.; Pan, G.; Zhou, Z. Comparative analysis of the proteins with tandem repeats from 8 microsporidia and characterization of a novel endospore wall protein colocalizing with polar tube from Nosema bombycis. J. Eukaryot. Microbiol. 2017, 64, 707–715. [Google Scholar] [CrossRef]
- Wang, Y.; Dang, X.; Ma, Q.; Liu, F.; Pan, G.; Li, T.; Zhou, Z. Characterization of a novel spore wall protein NbSWP16 with proline-rich tandem repeats from Nosema bombycis (microsporidia). Parasitology 2015, 142, 534–542. [Google Scholar] [CrossRef]
- Li, Y.; Wu, Z.; Pan, G.; He, W.; Zhang, R.; Hu, J.; Zhou, Z. Identification of a novel spore wall protein (SWP26) from microsporidia Nosema bombycis. Int. J. Parasitol. 2009, 39, 391–398. [Google Scholar] [CrossRef]
- Chen, J.; Geng, L.; Long, M.; Li, T.; Li, Z.; Yang, D.; Ma, C.; Wu, H.; Ma, Z.; Li, C. Identification of a novel chitin-binding spore wall protein (NbSWP12) with a BAR-2 domain from Nosema bombycis (microsporidia). Parasitology 2013, 140, 1394–1402. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Pan, G.; Li, T.; Huang, W.; Chen, J.; Geng, L.; Yang, D.; Wang, L.; Zhou, Z. SWP5, a spore wall protein, interacts with polar tube proteins in the parasitic microsporidian Nosema bombycis. Eukaryot. Cell 2012, 11, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, R.; You, Y.; Zhang, K.; Zhang, L. A novel spore wall protein from Antonospora locustae (Microsporidia: Nosematidae) contributes to sporulation. J. Eukaryot. Microbiol. 2017, 64, 779–791. [Google Scholar] [CrossRef] [PubMed]
- Jaroenlak, P.; Boakye, D.W.; Vanichviriyakit, R.; Williams, B.A.; Sritunyalucksana, K.; Itsathitphaisarn, O. Identification, characterization and heparin binding capacity of a spore-wall, virulence protein from the shrimp microsporidian, Enterocytozoon hepatopenaei (EHP). Parasites Vectors 2018, 11, 177. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Ao, T.Y.; Zhang, Z.; Ma, Z.G.; Zhou, Z.Y. Gene cloning, expression and subcellular localization of a novel spore wall protein of Nosema ceranae (Microsporidia). Acta Entomol. Sin. 2021, 9, 1070–1079. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Murray, M.; Thompson, W. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef]
- He, Y.L. Study on Histopathology of Pebrine and Preliminary Construction of Digital Silkworm. Master’s Thesis, Southwest University, Chongqing, China, 2012. [Google Scholar]
- Krüger, A.T.; Engel, J.; Buettner, F.F.; Routier, F.H. Aspergillus fumigatus Cap59-like protein A is involved in α1, 3-mannosylation of GPI-anchors. Glycobiology 2016, 26, 30–38. [Google Scholar]
- Wang, D. Functional Analysis of CFEM Domain-Containing Small Secreted Cysteine-Rich Proteins in Verticillium dahliae. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2020. [Google Scholar]
- Zhu, W.; Wei, W.; Wu, Y.; Zhou, Y.; Peng, F.; Zhang, S.; Chen, P.; Xu, X. BcCFEM1, a CFEM domain-containing protein with putative GPI-anchored site, is involved in pathogenicity, conidial production, and stress tolerance in Botrytis cinerea. Front. Microbiol. 2017, 8, 1807. [Google Scholar] [CrossRef]
- Zhao, X.; Oh, S.-H.; Hoyer, L.L. Deletion of ALS5, ALS6 or ALS7 increases adhesion of Candida albicans to human vascular endothelial and buccal epithelial cells. Sabouraudia 2007, 45, 429–434. [Google Scholar] [CrossRef]
- Li, D.D.; Wang, Y.; Li, X.X.; Zhao, L.X.; Jiang, Y.Y. GPI-anchored proteins in Candida albicans: Recent progress. Acad. J. Second. Mil. Med. Univ. 2012, 12, 1368–1373. [Google Scholar] [CrossRef]
- Fan, X.; Wei, C.; Yang, X.; Xiao, A.; Tan, N.; Chen, J.; Long, M.; Pan, G.; Wan, Y.; Zhou, Z. Proteomic analysis of spore surface proteins and characteristics of a novel spore wall protein and biomarker, EhSWP3, from the shrimp microsporidium Enterocytozoon hepatopenaei (EHP). Microorganisms 2022, 10, 367. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Geng, L.; Xu, J.; Jiang, P.; An, Q.; Pu, Y.; Jiang, Y.; He, S.; Tao, X.; Luo, J. Expression and identification of a novel spore wall protein in microsporidian Nosema bombycis. J. Eukaryot. Microbiol. 2020, 67, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Cali, A.; Takvorian, P.M. Developmental morphology and life cycles of the microsporidia. In Microsporidia: Pathogens of Opportunity; Wiley Publisher: Hoboken, NJ, USA, 2014; pp. 71–133. [Google Scholar]
- Yang, D.; Dang, X.; Tian, R.; Long, M.; Li, C.; Li, T.; Chen, J.; Li, Z.; Pan, G.; Zhou, Z. Development of an approach to analyze the interaction between Nosema bombycis (microsporidia) deproteinated chitin spore coats and spore wall proteins. J. Invertebr. Pathol. 2014, 115, 1–7. [Google Scholar] [CrossRef]
- Zheng, S.; Huang, Y.; Chen, J.; Wei, J.; Pan, G.; Li, C.; Zhou, Z. A specific molecular label for identifying mature Nosema bombycis spores. J. Invertebr. Pathol. 2020, 170, 107322. [Google Scholar] [CrossRef]
- Chen, J.; Guo, W.; Dang, X.; Huang, Y.; Liu, F.; Meng, X.; An, Y.; Long, M.; Bao, J.; Zhou, Z. Easy labeling of proliferative phase and sporogonic phase of microsporidia Nosema bombycis in host cells. PLoS ONE 2017, 12, e0179618. [Google Scholar] [CrossRef]
- Prudêncio, M.; Rodriguez, A.; Mota, M.M. The silent path to thousands of merozoites: The Plasmodium liver stage. Nat. Rev. Microbiol. 2006, 4, 849–856. [Google Scholar] [CrossRef]
- Zhu, F.; Shen, Z.; Hou, J.; Zhang, J.; Geng, T.; Tang, X.; Xu, L.; Guo, X. Identification of a protein interacting with the spore wall protein SWP26 of Nosema bombycis in a cultured BmN cell line of silkworm. Infect. Genet. Evol. 2013, 17, 38–45. [Google Scholar] [CrossRef]
- Southern, T.R.; Jolly, C.E.; Russell Hayman, J. Augmentation of microsporidia adherence and host cell infection by divalent cations. FEMS Microbiol. Lett. 2006, 260, 143–149. [Google Scholar] [CrossRef]
- Kappe, S.H.; Buscaglia, C.A.; Nussenzweig, V. Plasmodium sporozoite molecular cell biology. Annu. Rev. Cell Dev. Biol. 2004, 20, 29–59. [Google Scholar] [CrossRef]
- Dubey, J.; Lindsay, D.; Speer, C. Structures of Toxoplasma gondii tachyzoites, bradyzoites, and sporozoites and biology and development of tissue cysts. Clin. Microbiol. Rev. 1998, 11, 267–299. [Google Scholar] [CrossRef] [PubMed]
- Rostand, K.S.; Esko, J.D. Microbial adherence to and invasion through proteoglycans. Infect. Immun. 1997, 65, 1–8. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, P.; Li, D.; Hai, Q.; Liu, S.; Zhang, Y.; Zhang, J.; Xu, J.; Ma, Z.; Zhou, Z. NcSWP8, a New Spore Wall Protein, Interacts with Polar Tube Proteins in the Parasitic Microsporidia Vairimorpha (Nosema) ceranae. Microorganisms 2025, 13, 142. https://doi.org/10.3390/microorganisms13010142
Wang P, Li D, Hai Q, Liu S, Zhang Y, Zhang J, Xu J, Ma Z, Zhou Z. NcSWP8, a New Spore Wall Protein, Interacts with Polar Tube Proteins in the Parasitic Microsporidia Vairimorpha (Nosema) ceranae. Microorganisms. 2025; 13(1):142. https://doi.org/10.3390/microorganisms13010142
Chicago/Turabian StyleWang, Pengfei, Dufu Li, Qianmin Hai, Siming Liu, Yueyue Zhang, Jun Zhang, Jinshan Xu, Zhengang Ma, and Zeyang Zhou. 2025. "NcSWP8, a New Spore Wall Protein, Interacts with Polar Tube Proteins in the Parasitic Microsporidia Vairimorpha (Nosema) ceranae" Microorganisms 13, no. 1: 142. https://doi.org/10.3390/microorganisms13010142
APA StyleWang, P., Li, D., Hai, Q., Liu, S., Zhang, Y., Zhang, J., Xu, J., Ma, Z., & Zhou, Z. (2025). NcSWP8, a New Spore Wall Protein, Interacts with Polar Tube Proteins in the Parasitic Microsporidia Vairimorpha (Nosema) ceranae. Microorganisms, 13(1), 142. https://doi.org/10.3390/microorganisms13010142