
Colonic Microbiota Improves Fiber Digestion Ability and Enhances Absorption of Short-Chain Fatty Acids in Local Pigs of Hainan

 ,
,
Abstract
1. Background
2. Methods
2.1. Experimental Design and Sample Collection
2.2. Intestinal Morphology
2.3. Metagenomic Sequencing and Data Processing
2.4. Metabolite Extraction, LC-MS Analysis, and Data Processing
2.5. Bioinformatics Analysis
2.6. RNA Isolation, Reverse Transcription, and Real-Time Quantitative PCR
2.7. Statistical Analysis
3. Results
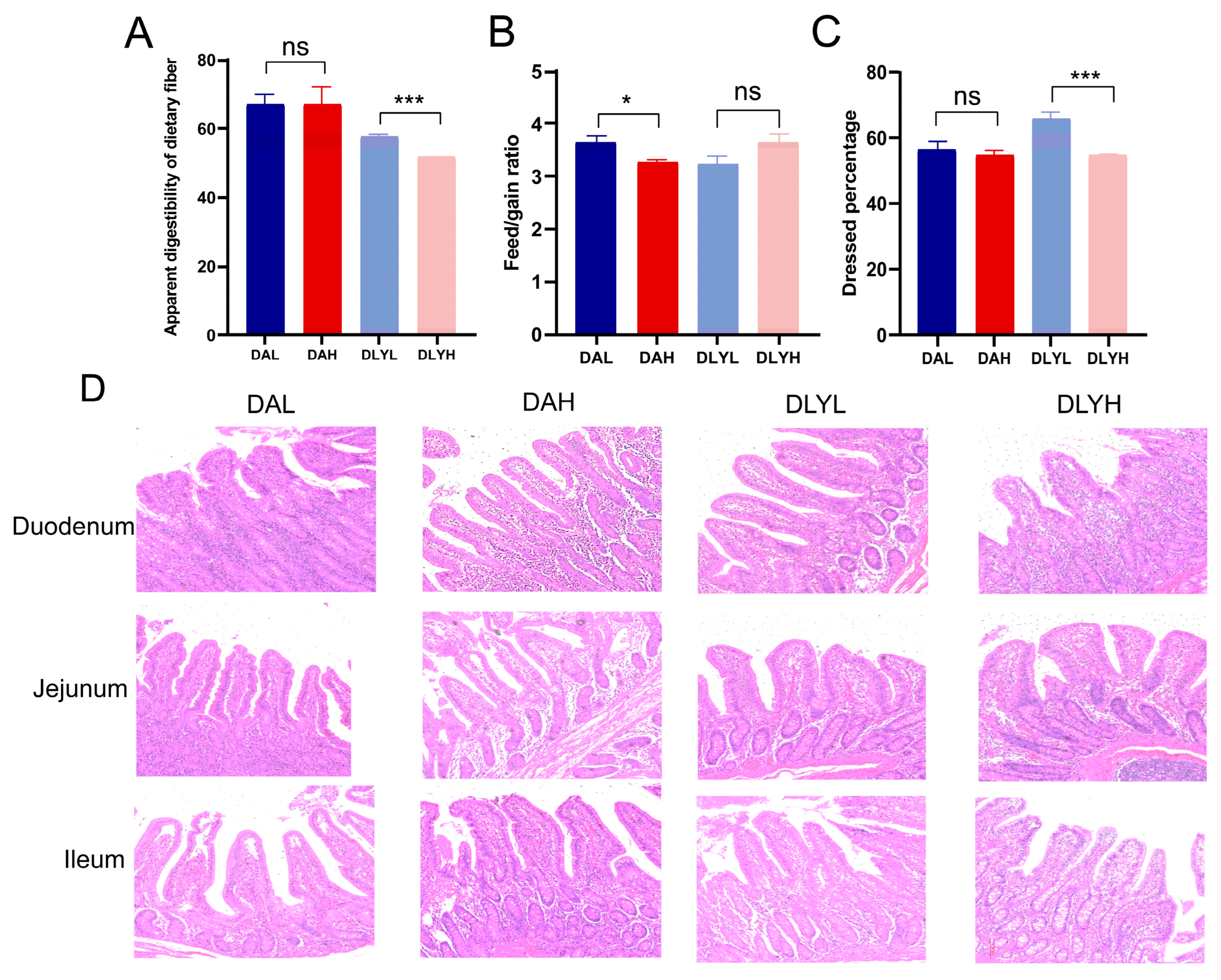
3.1. Effect of Different Fiber Contents on Phenotype of Two Breeds of Pigs
3.2. Influence of Dietary Fiber on Intestinal Morphology
3.3. Metagenomic Sequencing Data of Colonic Contents
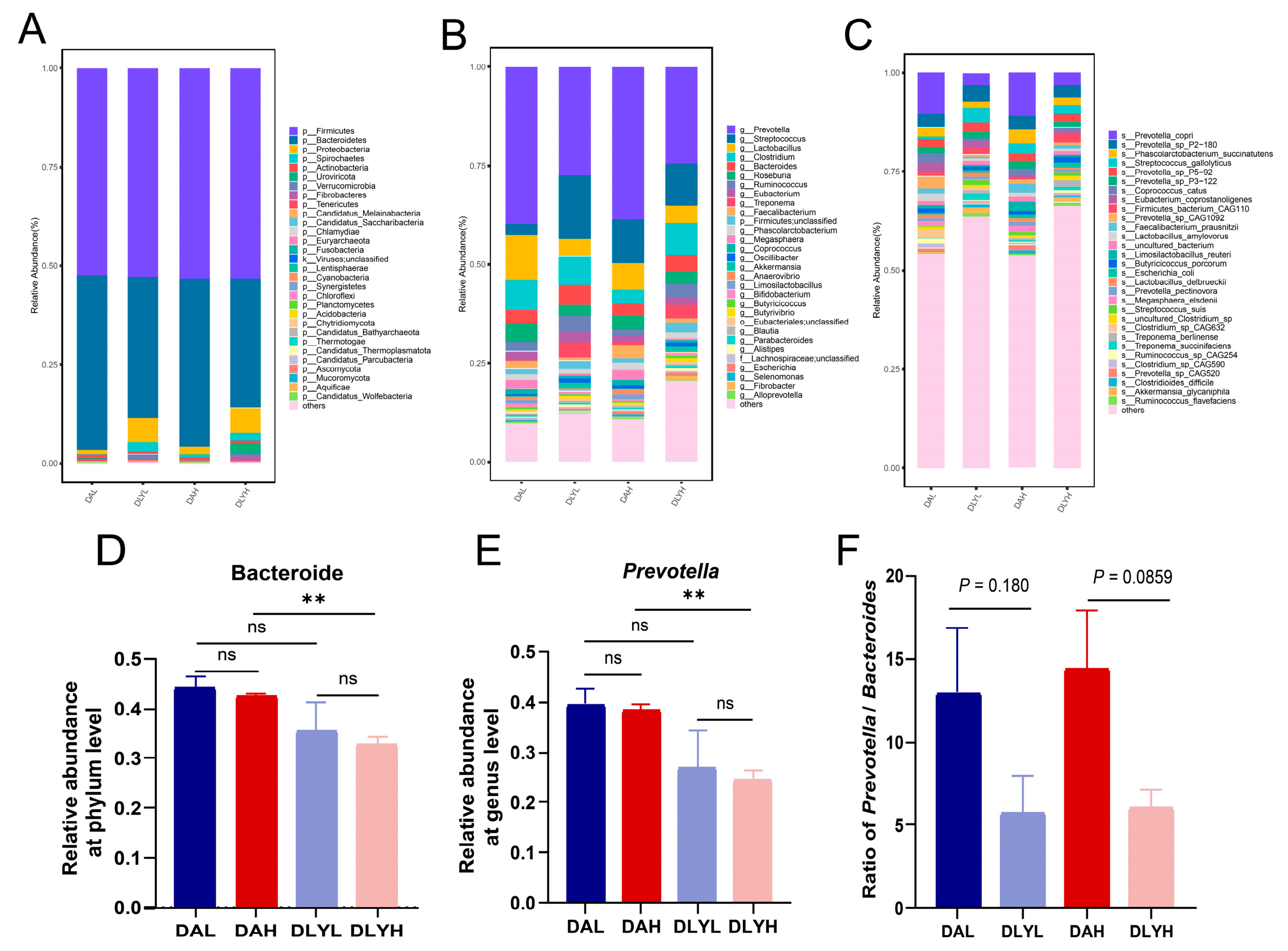
3.4. Taxonomic Composition of Colonic Microbiota in the Four Groups
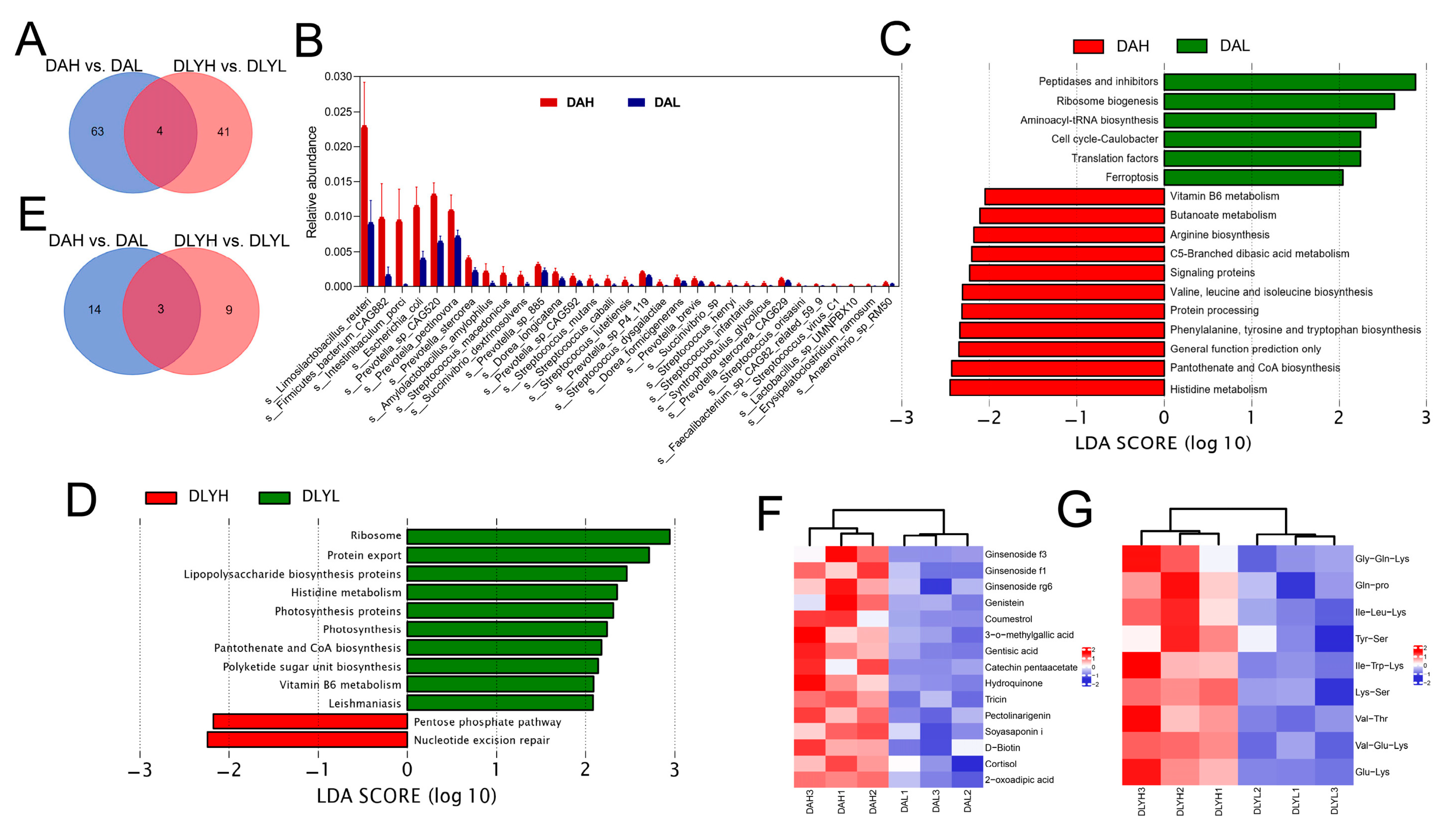
3.5. Effect of Feed Factor on the Colonic Microbiota and Metabolites
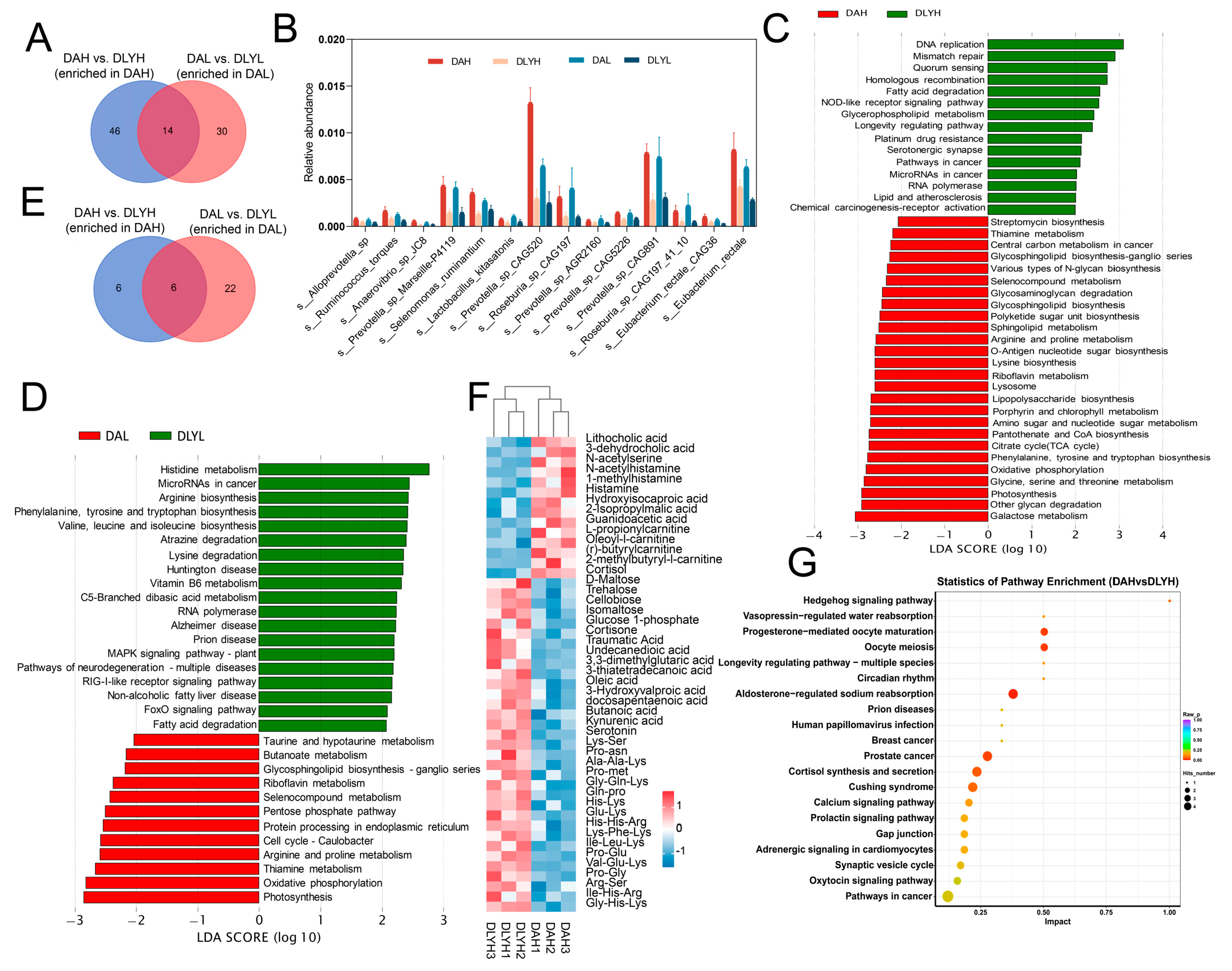
3.6. Effect of Breed Factor on the Colonic Microbiota and Metabolites
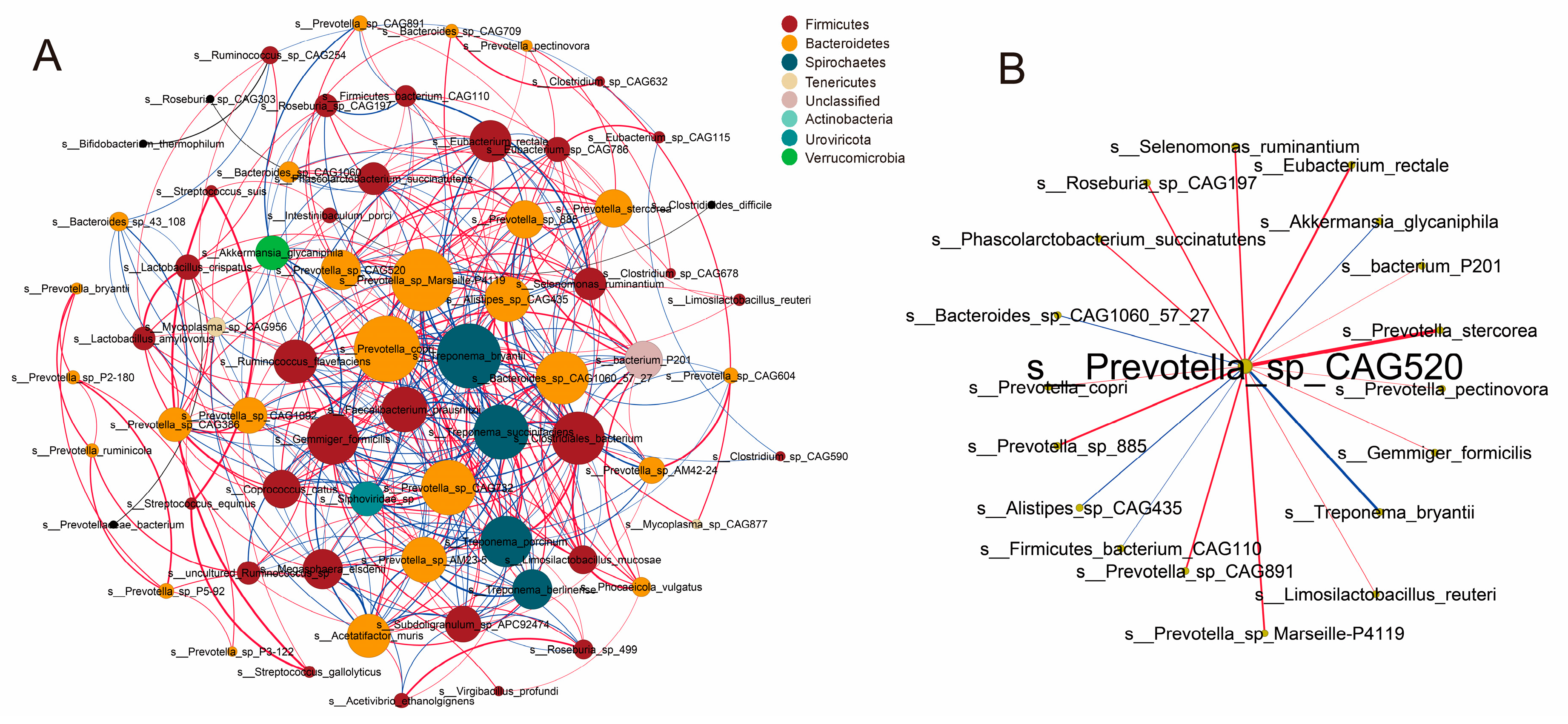
3.7. Co-Occurrence Networks of Intestinal Microbiota
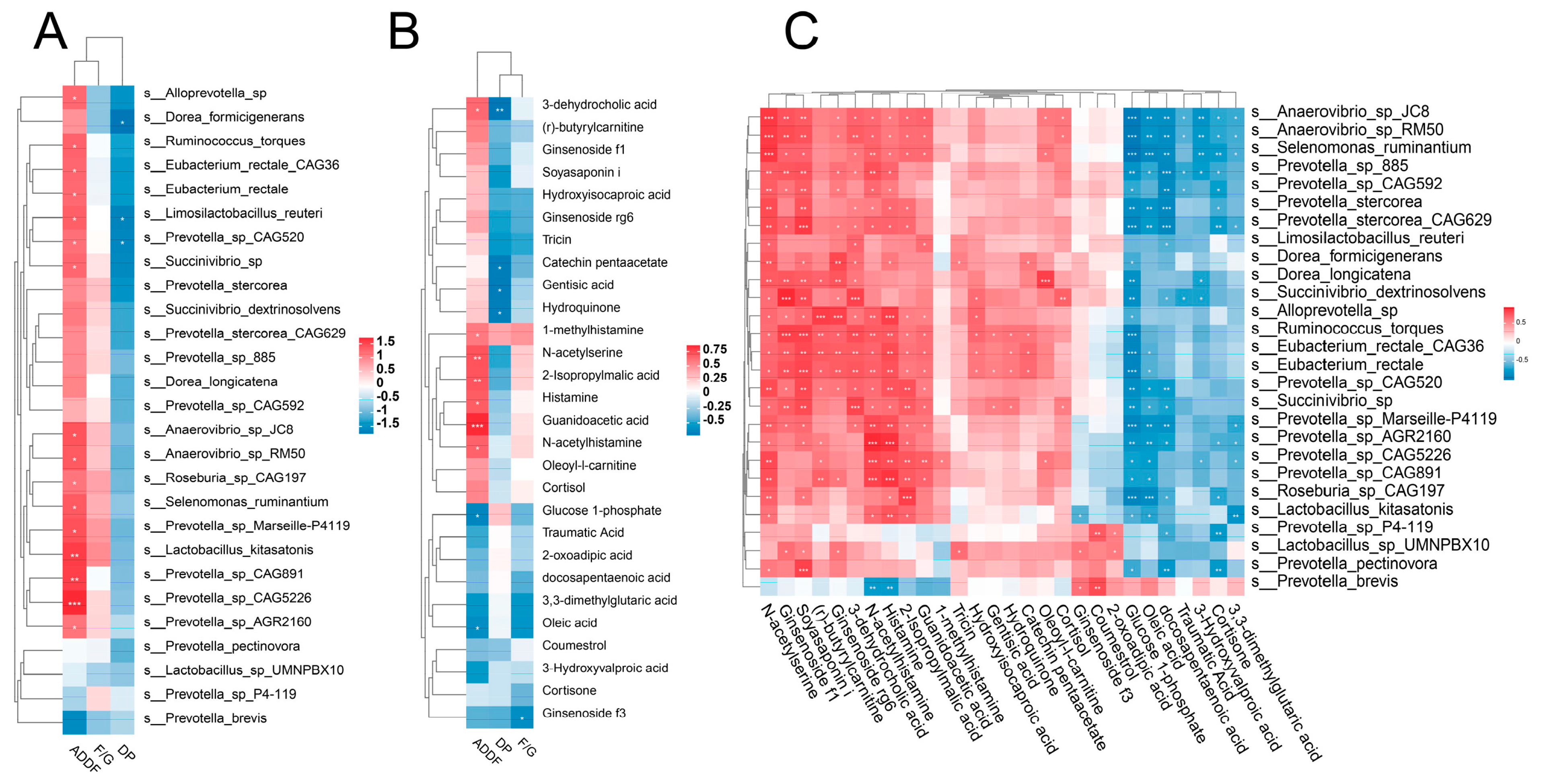
3.8. Correlation Analysis of Phenotype, Microbiota, and Metabolites
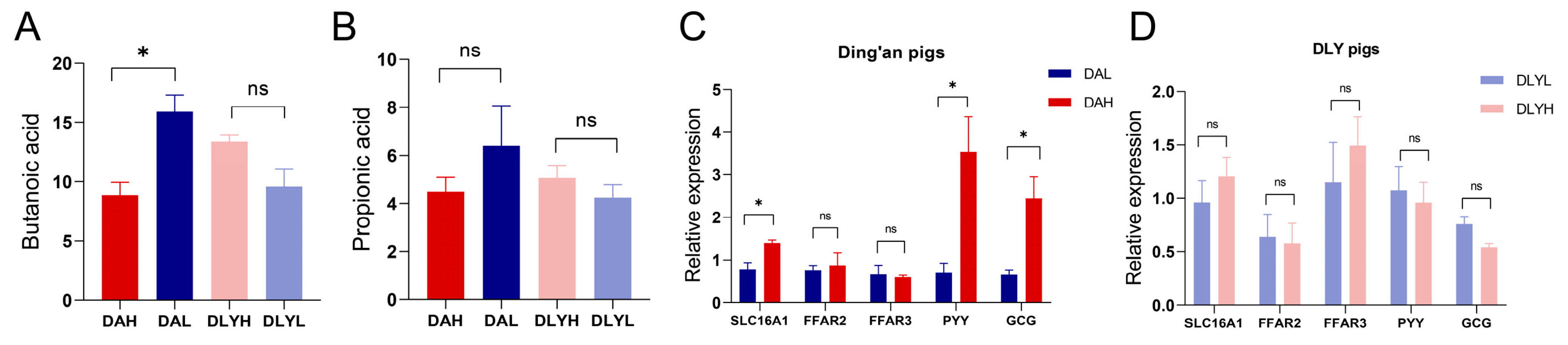
3.9. Increasing Fiber Content Can Improve the Absorption of SCFAs in Ding’an Pigs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DLY | Duroc × Landrace × Yorkshire |
| H&E | Hematoxylin–eosin |
| V/C | Villus height to crypt depth |
| PCoA | Principal co-ordinates analysis |
| LEfSe | Linear discriminate analysis effect size |
| LDA | Linear discriminant analysis |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| OPLS-DA | Orthogonal partial least squares discriminant analysis |
| VIP | Variable importance in the projection |
| SEM | Standard error of mean |
| SCFAs | Short-chain fatty acids |
References
- Tiwari, U.P.; Mandal, R.K.; Neupane, K.R.; Mishra, B.; Jha, R. Starchy and fibrous feedstuffs differ in their in vitro digestibility and fermentation characteristics and differently modulate gut microbiota of swine. J. Anim. Sci. Biotechnol. 2022, 13, 53. [Google Scholar] [CrossRef]
- Williams, B.A.; Mikkelsen, D.; Flanagan, B.M.; Gidley, M.J. “Dietary fibre”: Moving beyond the “soluble/insoluble” classification for monogastric nutrition, with an emphasis on humans and pigs. J. Anim. Sci. Biotechnol. 2019, 10, 45. [Google Scholar] [CrossRef]
- Upadhaya, S.D.; Kim, I.H. Maintenance of gut microbiome stability for optimum intestinal health in pigs—A review. J. Anim. Sci. Biotechnol. 2022, 13, 140. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, S.; Kim, E.S.; Keum, G.B.; Doo, H.; Kwak, J.; Pandey, S.; Cho, J.H.; Ryu, S.; Song, M.; et al. Comparative analysis of the pig gut microbiome associated with the pig growth performance. J. Anim. Sci. Technol. 2023, 65, 856–864. [Google Scholar] [CrossRef]
- Gardiner, G.E.; Metzler-Zebeli, B.U.; Lawlor, P.G. Impact of Intestinal Microbiota on Growth and Feed Efficiency in Pigs: A Review. Microorganisms 2020, 8, 1886. [Google Scholar] [CrossRef] [PubMed]
- Varel, V.H. Activity of fiber-degrading microorganisms in the pig large intestine. J. Anim. Sci. 1987, 65, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Li, P.; Hou, L.; Niu, Q.; Pu, G.; Wang, B.; Du, T.; Kim, S.W.; Niu, P.; Li, Q.; et al. Metagenomic Analysis Reveals New Microbiota Related to Fiber Digestion in Pigs. Front. Microbiol. 2021, 12, 746717. [Google Scholar] [CrossRef] [PubMed]
- Pond, W.G.; Varel, V.H.; Dickson, J.S.; Haschek, W.M. Comparative response of swine and rats to high-fiber or high-protein diets. J. Anim. Sci. 1989, 67, 716. [Google Scholar] [CrossRef]
- Liu, B.; Zhu, X.; Cui, Y.; Wang, W.; Liu, H.; Li, Z.; Guo, Z.; Ma, S.; Li, D.; Wang, C.; et al. Consumption of Dietary Fiber from Different Sources during Pregnancy Alters Sow Gut Microbiota and Improves Performance and Reduces Inflammation in Sows and Piglets. mSystems 2021, 6, 10–128. [Google Scholar] [CrossRef]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef]
- Morrison, K.E.; Jasarevic, E.; Howard, C.D.; Bale, T.L. It’s the fiber, not the fat: Significant effects of dietary challenge on the gut microbiome. Microbiome 2020, 8, 15. [Google Scholar] [CrossRef]
- Tang, X.; Zhang, L.; Fan, C.; Wang, L.; Fu, H.; Ren, S.; Shen, W.; Jia, S.; Wu, G.; Zhang, Y. Dietary Fiber Influences Bacterial Community Assembly Processes in the Gut Microbiota of Durco x Bamei Crossbred Pig. Front. Microbiol. 2021, 12, 688554. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Peng, L.; Li, Z.R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate enhances the intestinal barrier by facilitating tight junction assembly via activation of AMP-activated protein kinase in Caco-2 cell monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Kong, F.; Xiang, Y.; Zhou, W.; Wang, J.; Yang, H.; Zhang, G.; Zhao, J. Comparative biogeography of the gut microbiome between Jinhua and Landrace pigs. Sci. Rep. 2018, 8, 5985. [Google Scholar] [CrossRef] [PubMed]
- Pu, G.; Li, P.; Du, T.; Niu, Q.; Fan, L.; Wang, H.; Liu, H.; Li, K.; Niu, P.; Wu, C.; et al. Adding Appropriate Fiber in Diet Increases Diversity and Metabolic Capacity of Distal Gut Microbiota Without Altering Fiber Digestibility and Growth Rate of Finishing Pig. Front. Microbiol. 2020, 11, 533. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.J.; Kornegay, E.T.; Grayson, R.L.; Lindemann, M.D. Growth, nutrient utilization and intestinal morphology of pigs fed high-fiber diets. J. Anim. Sci. 1988, 66, 1570–1579. [Google Scholar] [CrossRef]
- Gutierrez, N.A.; Kerr, B.J.; Patience, J.F. Effect of insoluble-low fermentable fiber from corn-ethanol distillation origin on energy, fiber, and amino acid digestibility, hindgut degradability of fiber, and growth performance of pigs. J. Anim. Sci. 2013, 91, 5314–5325. [Google Scholar] [CrossRef]
- Cheng, P.; Wang, Y.; Liang, J.; Wu, Y.; Wright, A.; Liao, X. Exploratory Analysis of the Microbiological Potential for Efficient Utilization of Fiber Between Lantang and Duroc Pigs. Front. Microbiol. 2018, 9, 1342. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, C.; Luo, R.; Sadakane, K.; Lam, T. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, D.; Chen, G.L.; Locascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Wen, B.; Mei, Z.; Zeng, C.; Liu, S. metaX: A flexible and comprehensive software for processing metabolomics data. BMC Bioinform. 2017, 18, 183. [Google Scholar] [CrossRef] [PubMed]
- Eklund, M.; Rademacher, M.; Sauer, W.C.; Blank, R.; Mosenthin, R. Standardized ileal digestibility of amino acids in alfalfa meal, sugar beet pulp, and wheat bran compared to wheat and protein ingredients for growing pigs1. J. Anim. Sci. 2014, 92, 1037–1043. [Google Scholar] [CrossRef]
- Laudadio, V.; Ceci, E.; Lastella, N.M.B.; Introna, M.; Tufarelli, V. Low-fiber alfalfa (Medicago sativa L.) meal in the laying hen diet: Effects on productive traits and egg quality. Poult. Sci. 2014, 93, 1868–1874. [Google Scholar] [CrossRef]
- von Heimendahl, E.; Breves, G.; Abel, H.J. Fiber-related digestive processes in three different breeds of pigs. J. Anim. Sci. 2010, 88, 972–981. [Google Scholar] [CrossRef]
- Bubier, J.A.; Chesler, E.J.; Weinstock, G.M. Host genetic control of gut microbiome composition. Mamm. Genome 2021, 32, 263–281. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Xie, Y.; Zhong, Y.; Ma, X.; Sun, H.; Liu, J. Integrated meta-omics reveals new ruminal microbial features associated with feed efficiency in dairy cattle. Microbiome 2022, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, E.J.C.; Iljazovic, A.; Amend, L.; Lesker, T.R.; Renault, T.; Thiemann, S.; Hao, L.; Roy, U.; Gronow, A.; Charpentier, E.; et al. Distinct Polysaccharide Utilization Determines Interspecies Competition between Intestinal Prevotella spp. Cell Host Microbe 2020, 28, 838–852. [Google Scholar] [CrossRef] [PubMed]
- Kovatcheva-Datchary, P.; Nilsson, A.; Akrami, R.; Lee, Y.S.; De Vadder, F.; Arora, T.; Hallen, A.; Martens, E.; Björck, I.; Bäckhed, F. Dietary Fiber-Induced Improvement in Glucose Metabolism Is Associated with Increased Abundance of Prevotella. Cell Metab. 2015, 22, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Tett, A.; Pasolli, E.; Masetti, G.; Ercolini, D.; Segata, N. Prevotella diversity, niches and interactions with the human host. Nat. Rev. Microbiol. 2021, 19, 585–599. [Google Scholar] [CrossRef] [PubMed]
- De Vadder, F.; Kovatcheva-Datchary, P.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-Produced Succinate Improves Glucose Homeostasis via Intestinal Gluconeogenesis. Cell Metab. 2016, 24, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Fang, S.; Wei, H.; He, M.; Fu, H.; Xiong, X.; Zhou, Y.; Wu, J.; Gao, J.; Yang, H.; et al. Prevotella copri increases fat accumulation in pigs fed with formula diets. Microbiome 2021, 9, 175. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Diez-Echave, P.; Martín-Cabrejas, I.; Garrido-Mesa, J.; Langa, S.; Vezza, T.; Landete, J.M.; Hidalgo-García, L.; Algieri, F.; Mayer, M.J.; Narbad, A.; et al. Probiotic and Functional Properties of Limosilactobacillus reuteri INIA P572. Nutrients 2021, 13, 1860. [Google Scholar] [CrossRef]
- Mizrahi, I.; Wallace, R.J.; Moraïs, S. The rumen microbiome: Balancing food security and environmental impacts. Nat. Rev. Microbiol. 2021, 19, 553–566. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.K.; Deehan, E.C.; Zhang, Z.; Jin, M.; Baskota, N.; Perez-Muñoz, M.E.; Cole, J.; Tuncil, Y.E.; Seethaler, B.; Wang, T.; et al. Gut microbiota modulation with long-chain corn bran arabinoxylan in adults with overweight and obesity is linked to an individualized temporal increase in fecal propionate. Microbiome 2020, 8, 118. [Google Scholar] [CrossRef] [PubMed]
- Nilsen, M.; Madelen Saunders, C.; Leena Angell, I.; Arntzen, M.Ø.; Lødrup Carlsen, K.C.; Carlsen, K.; Haugen, G.; Heldal Hagen, L.; Carlsen, M.H.; Hedlin, G.; et al. Butyrate Levels in the Transition from an Infant- to an Adult-Like Gut Microbiota Correlate with Bacterial Networks Associated with Eubacterium Rectale and Ruminococcus Gnavus. Genes 2020, 11, 1245. [Google Scholar] [CrossRef] [PubMed]
- Marques, F.Z.; Nelson, E.; Chu, P.; Horlock, D.; Fiedler, A.; Ziemann, M.; Tan, J.K.; Kuruppu, S.; Rajapakse, N.W.; El-Osta, A.; et al. High-Fiber Diet and Acetate Supplementation Change the Gut Microbiota and Prevent the Development of Hypertension and Heart Failure in Hypertensive Mice. Circulation 2017, 135, 964–977. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Medina, N.; Barrios-Camacho, H.; Duran-Bedolla, J.; Garza-Ramos, U. Klebsiella variicola: An emerging pathogen in humans. Emerg. Microbes Infect. 2019, 8, 973–988. [Google Scholar] [CrossRef] [PubMed]
- Uzal, F.A.; Navarro, M.A.; Li, J.; Freedman, J.C.; Shrestha, A.; McClane, B.A. Comparative pathogenesis of enteric clostridial infections in humans and animals. Anaerobe 2018, 53, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Mu, Q.; Tavella, V.J.; Luo, X.M. Role of Lactobacillus reuteri in Human Health and Diseases. Front. Microbiol. 2018, 9, 315828. [Google Scholar] [CrossRef] [PubMed]
- Nograšek, B.; Accetto, T.; Fanedl, L.; Avguštin, G. Description of a novel pectin-degrading bacterial species Prevotella pectinovora sp. nov. based on its phenotypic and genomic traits. J. Microbiol. 2015, 53, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Yeoh, Y.K.; Sun, Y.; Ip, L.Y.T.; Wang, L.; Chan, F.K.L.; Miao, Y.; Ng, S.C. Prevotella species in the human gut is primarily comprised of Prevotella copri, Prevotella stercorea and related lineages. Sci. Rep. 2022, 12, 9055. [Google Scholar] [CrossRef]
- Jonczyk, R.; Ronconi, S.; Rychlik, M.; Genschel, U. Pantothenate synthetase is essential but not limiting for pantothenate biosynthesis in Arabidopsis. Plant Mol. Biol. 2008, 66, 1–14. [Google Scholar] [CrossRef]
- Ueland, P.M.; McCann, A.; Midttun, O.; Ulvik, A. Inflammation, vitamin B6 and related pathways. Mol. Asp. Med. 2017, 53, 10–27. [Google Scholar] [CrossRef]
- Bergamaschi, M.; Tiezzi, F.; Howard, J.; Huang, Y.J.; Gray, K.A.; Schillebeeckx, C.; McNulty, N.P.; Maltecca, C. Gut microbiome composition differences among breeds impact feed efficiency in swine. Microbiome 2020, 8, 110. [Google Scholar] [CrossRef]
- La Rosa, S.L.; Leth, M.L.; Michalak, L.; Hansen, M.E.; Pudlo, N.A.; Glowacki, R.; Pereira, G.; Workman, C.T.; Arntzen, M.Ø.; Pope, P.B.; et al. The human gut Firmicute Roseburia intestinalis is a primary degrader of dietary β-mannans. Nat. Commun. 2019, 10, 905. [Google Scholar] [CrossRef] [PubMed]
- Sawanon, S.; Koike, S.; Kobayashi, Y. Evidence for the possible involvement of Selenomonas ruminantium in rumen fiber digestion. FEMS Microbiol. Lett. 2011, 325, 170–179. [Google Scholar] [CrossRef]
- Wardman, J.F.; Bains, R.K.; Rahfeld, P.; Withers, S.G. Carbohydrate-active enzymes (CAZymes) in the gut microbiome. Nat. Rev. Microbiol. 2022, 20, 542–556. [Google Scholar] [CrossRef] [PubMed]
- Al-Khayri, J.M.; Sahana, G.R.; Nagella, P.; Joseph, B.V.; Alessa, F.M.; Al-Mssallem, M.Q. Flavonoids as Potential Anti-Inflammatory Molecules: A Review. Molecules 2022, 27, 2901. [Google Scholar] [CrossRef] [PubMed]
- Im, D. Pro-Resolving Effect of Ginsenosides as an Anti-Inflammatory Mechanism of Panax ginseng. Biomolecules 2020, 10, 444. [Google Scholar] [CrossRef]
- Zywno, H.; Bzdega, W.; Kolakowski, A.; Kurzyna, P.; Harasim-Symbor, E.; Sztolsztener, K.; Chabowski, A.; Konstantynowicz-Nowicka, K. The Influence of Coumestrol on Sphingolipid Signaling Pathway and Insulin Resistance Development in Primary Rat Hepatocytes. Biomolecules 2021, 11, 268. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, X.; Leonard, S.; Archbold, T.; Sullivan, J.A.; Duncan, A.M.; Ma, W.D.L.; Bizimungu, B.; Murphy, A.; Htoo, J.K.; et al. Responses of dietary ileal amino acid digestibility to consumption of different cultivars of potatoes and conventional fibers in grower pigs fed a high-fat basal diet1. J. Anim. Sci. 2012, 90 (Suppl. S4), 356–358. [Google Scholar] [CrossRef]
- Mekbungwan, A.; Yamauchi, K. Growth performance and histological intestinal alterations in piglets fed dietary raw and heated pigeon pea seed meal. Histol. Histopathol. 2004, 19, 381–389. [Google Scholar]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [PubMed]
- Sivaprakasam, S.; Bhutia, Y.D.; Yang, S.; Ganapathy, V. Short-Chain Fatty Acid Transporters: Role in Colonic Homeostasis. Compr. Physiol. 2017, 8, 299–314. [Google Scholar] [PubMed]
- Wichmann, A.; Allahyar, A.; Greiner, T.U.; Plovier, H.; Lunden, G.O.; Larsson, T.; Drucker, D.J.; Delzenne, N.M.; Cani, P.D.; Backhed, F. Microbial modulation of energy availability in the colon regulates intestinal transit. Cell Host Microbe 2013, 14, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Larraufie, P.; Martin-Gallausiaux, C.; Lapaque, N.; Dore, J.; Gribble, F.M.; Reimann, F.; Blottiere, H.M. SCFAs strongly stimulate PYY production in human enteroendocrine cells. Sci. Rep. 2018, 8, 74. [Google Scholar] [CrossRef] [PubMed]
- Agus, A.; Clément, K.; Sokol, H. Gut microbiota-derived metabolites as central regulators in metabolic disorders. Gut 2021, 70, 1174–1182. [Google Scholar] [CrossRef]
- Gill, S.K.; Rossi, M.; Bajka, B.; Whelan, K. Dietary fibre in gastrointestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 101–116. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Dietary Fiber 5% | Dietary Fiber 10% |
|---|---|---|
| Ingredients% | ||
| Corn | 49.4 | 32.7 |
| Alfalfa | 2.7 | 16.9 |
| Wheat bran | 13.1 | 13.1 |
| Soybean meal | 18 | 18 |
| Rice bran | 13 | 13 |
| Soybean oil | 1 | 3.5 |
| Limestone | 1.4 | 1.4 |
| Sodium chloride | 0.3 | 0.3 |
| L-Lysine | 0.1 | 0.1 |
| Premix | 1 | 1 |
| Total | 100 | 100 |
| Nutrient level * | ||
| DE, MJ·kg−1 | 13 | 12.89 |
| CP % | 16.01 | 16.3 |
| Lys % | 0.85 | 0.84 |
| Met + Cys % | 0.44 | 0.43 |
| Trp % | 0.14 | 0.14 |
| Thr % | 0.54 | 0.53 |
| Dietary fiber % | 5 | 10 |
| Calcium % | 0.64 | 0.65 |
| Phosphorus % | 0.55 | 0.54 |
| Item | Ding’an Pigs | DLY Pigs | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|
| DAL | DAH | DLYL | DLYH | Breed | DF | Interaction | ||
| Duodenum | ||||||||
| Villus height (μm) | 377 ± 1 b | 374 ± 3 b | 416 ± 6 a | 375 ± 2 b | 1.636 | 0.001 | 0.001 | 0.001 |
| Crypt depth (μm) | 171 ± 5 b | 163 ± 5 b | 200 ± 6 a | 202 ± 6 a | 2.737 | 0.001 | 0.626 | 0.354 |
| V/C | 2.25 ± 0.06 ab | 2.34 ± 0.06 a | 2.11 ± 0.06 b | 1.92 ± 0.06 c | 0.030 | 0.001 | 0.378 | 0.019 |
| Jejunum | ||||||||
| Villus height (μm) | 412 ± 6 a | 397 ± 6 ab | 389 ± 6 b | 393 ± 4 ab | 2.815 | 0.020 | 0.359 | 0.098 |
| Crypt depth (μm) | 185 ± 8 a | 179 ± 5 a | 175 ± 4 a | 172 ± 5 a | 2.874 | 0.114 | 0.474 | 0.798 |
| V/C | 2.30 ± 0.08 a | 2.25 ± 0.06 a | 2.25 ± 0.04 a | 2.33 ± 0.07 a | 0.032 | 0.726 | 0.793 | 0.318 |
| Ileum | ||||||||
| Villus height (μm) | 393 ± 6 a | 405 ± 6 a | 398 ± 5 a | 353 ± 3 b | 2.602 | 0.001 | 0.002 | 0.001 |
| Crypt depth (μm) | 194 ± 6 a | 193 ± 6 a | 185 ± 5 a | 188 ± 6 a | 2.819 | 0.227 | 0.820 | 0.744 |
| V/C | 2.06 ± 0.05 ab | 2.13 ± 0.06 a | 2.18 ± 0.05 a | 1.92 ± 0.07 b | 0.029 | 0.451 | 0.129 | 0.006 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, P.; Xue, M.; Luo, Y.; Tang, Q.; Wang, F.; Sun, R.; Song, Y.; Chao, Z.; Fang, M. Colonic Microbiota Improves Fiber Digestion Ability and Enhances Absorption of Short-Chain Fatty Acids in Local Pigs of Hainan. Microorganisms 2024, 12, 1033. https://doi.org/10.3390/microorganisms12061033
Xue P, Xue M, Luo Y, Tang Q, Wang F, Sun R, Song Y, Chao Z, Fang M. Colonic Microbiota Improves Fiber Digestion Ability and Enhances Absorption of Short-Chain Fatty Acids in Local Pigs of Hainan. Microorganisms. 2024; 12(6):1033. https://doi.org/10.3390/microorganisms12061033
Chicago/Turabian StyleXue, Pengxiang, Mingming Xue, Yabiao Luo, Qiguo Tang, Feng Wang, Ruiping Sun, Yanxia Song, Zhe Chao, and Meiying Fang. 2024. "Colonic Microbiota Improves Fiber Digestion Ability and Enhances Absorption of Short-Chain Fatty Acids in Local Pigs of Hainan" Microorganisms 12, no. 6: 1033. https://doi.org/10.3390/microorganisms12061033
APA StyleXue, P., Xue, M., Luo, Y., Tang, Q., Wang, F., Sun, R., Song, Y., Chao, Z., & Fang, M. (2024). Colonic Microbiota Improves Fiber Digestion Ability and Enhances Absorption of Short-Chain Fatty Acids in Local Pigs of Hainan. Microorganisms, 12(6), 1033. https://doi.org/10.3390/microorganisms12061033