
Secreted Metabolites from Pseudomonas, Staphylococcus, and Borrelia Biofilm: Modulation of Immunogenicity by a Nutraceutical Enzyme and Botanical Blend


Abstract
1. Introduction
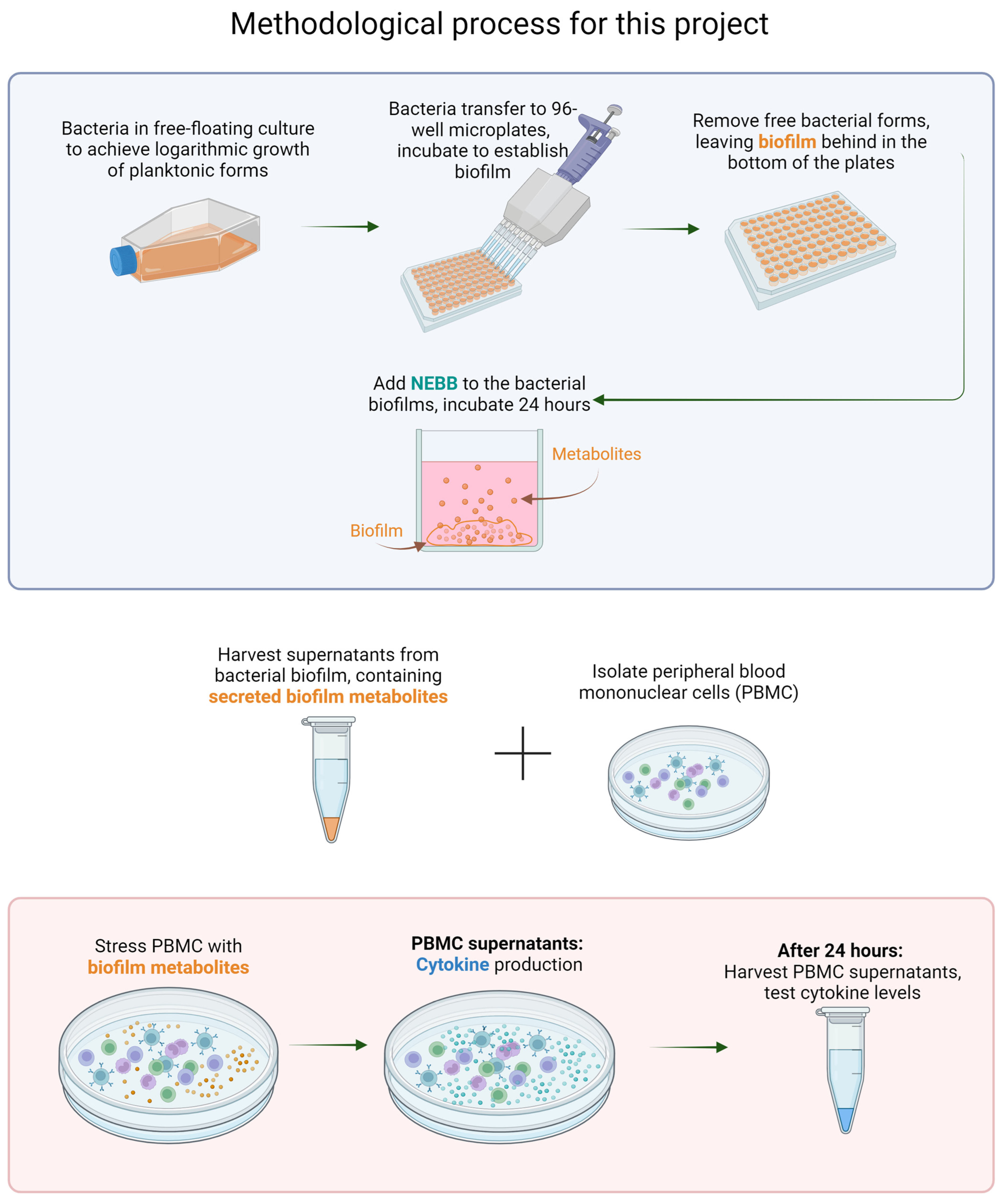
2. Materials and Methods
2.1. Reagents
2.2. Bacterial Strains
2.3. Nutraceutical Enzyme and Botanical Blend
2.4. Removal of Planktonic Forms
2.5. Harvesting of Biofilm Metabolites
2.6. Treatment of Human Immune Cells with Bacterial Biofilm Metabolites
2.7. Immune Cell Activation
2.8. Quantification of Cytokine Production by the Immune Cells in Reaction to Biofilm Metabolites
2.9. Statistical Analysis
3. Results
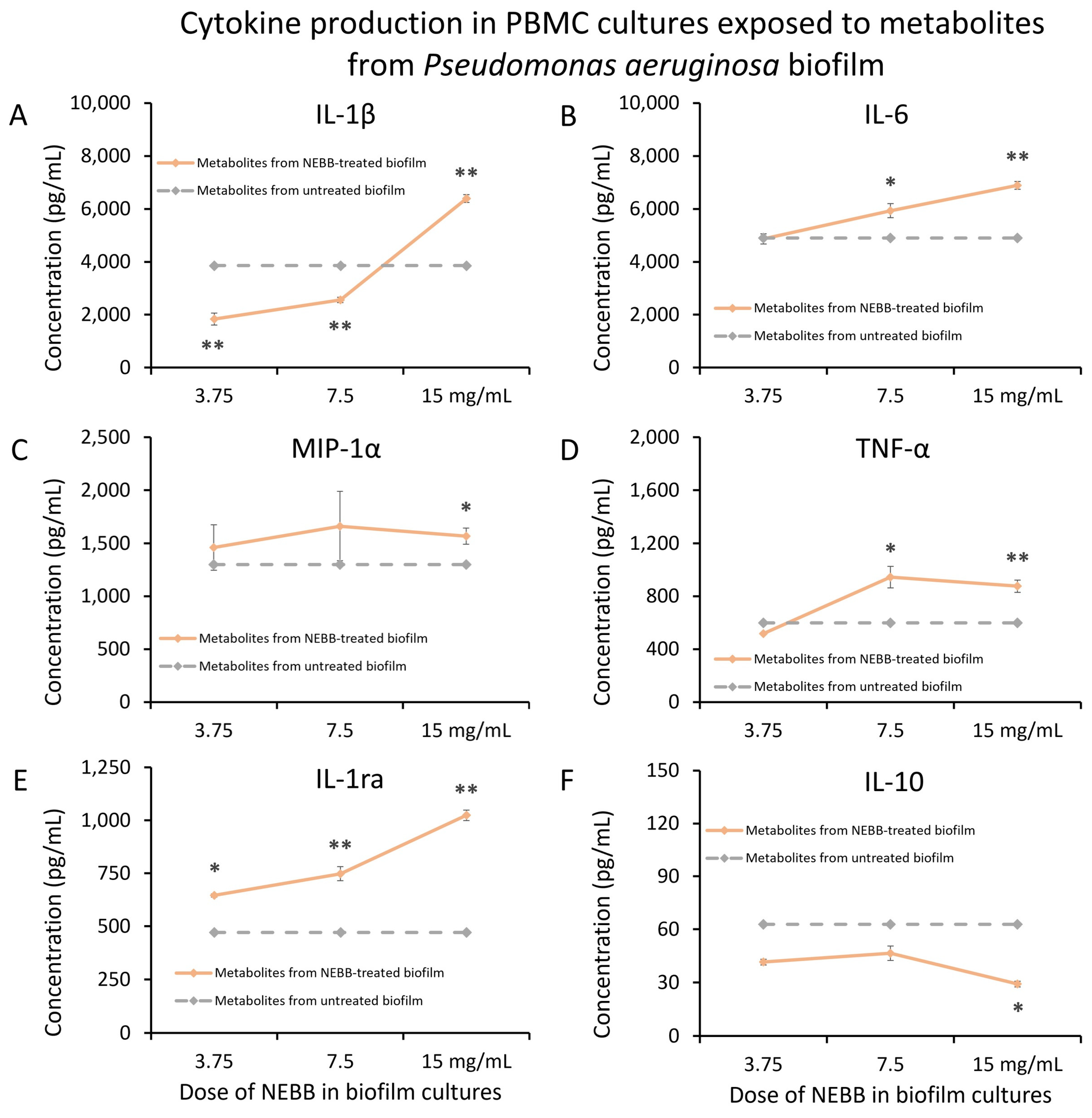
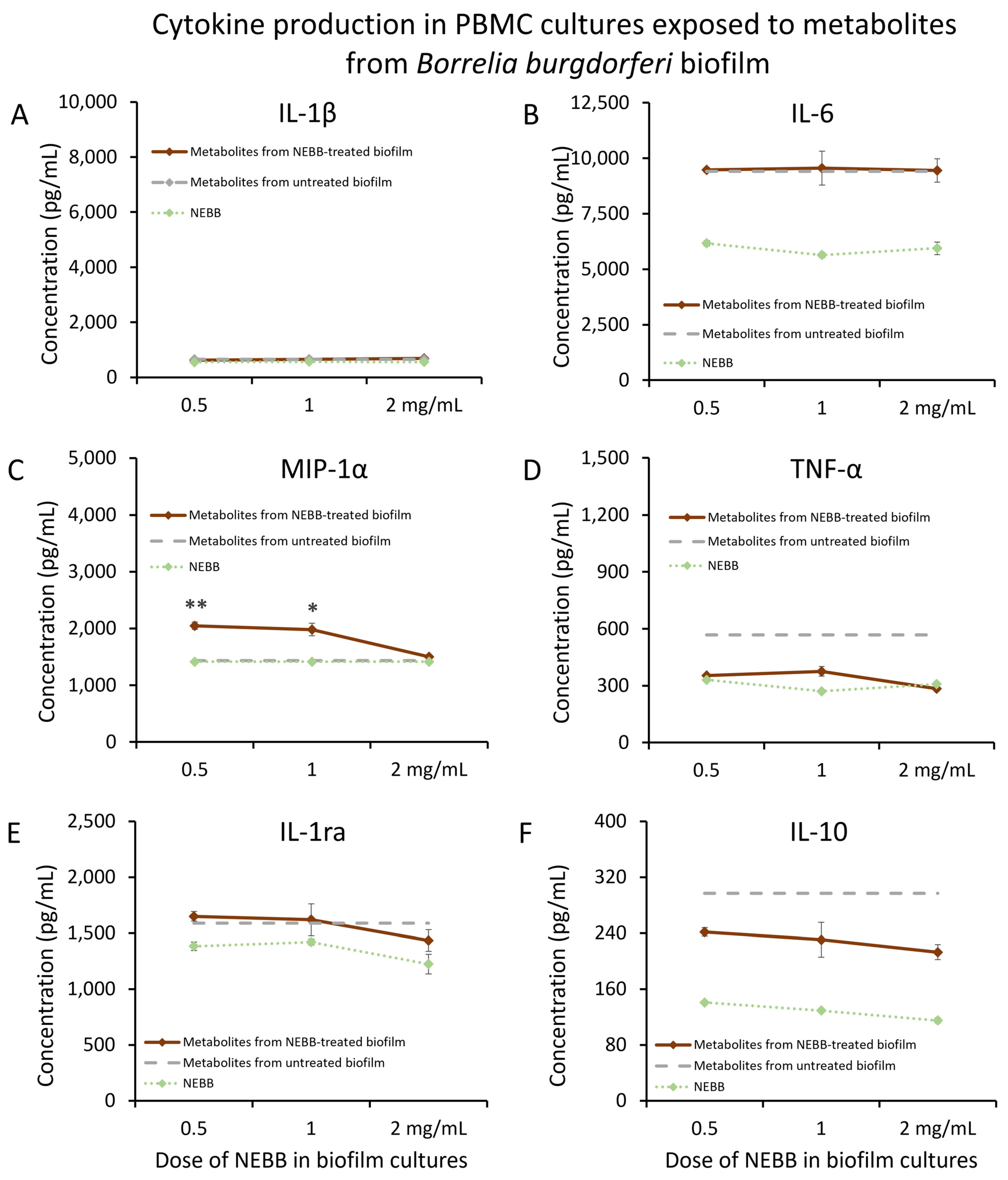
3.1. Immunomodulatory Activity of Secreted Biofilm Metabolites
3.2. Secreted Metabolites from NEBB-Treated Biofilm Have Immune-Modulating Properties
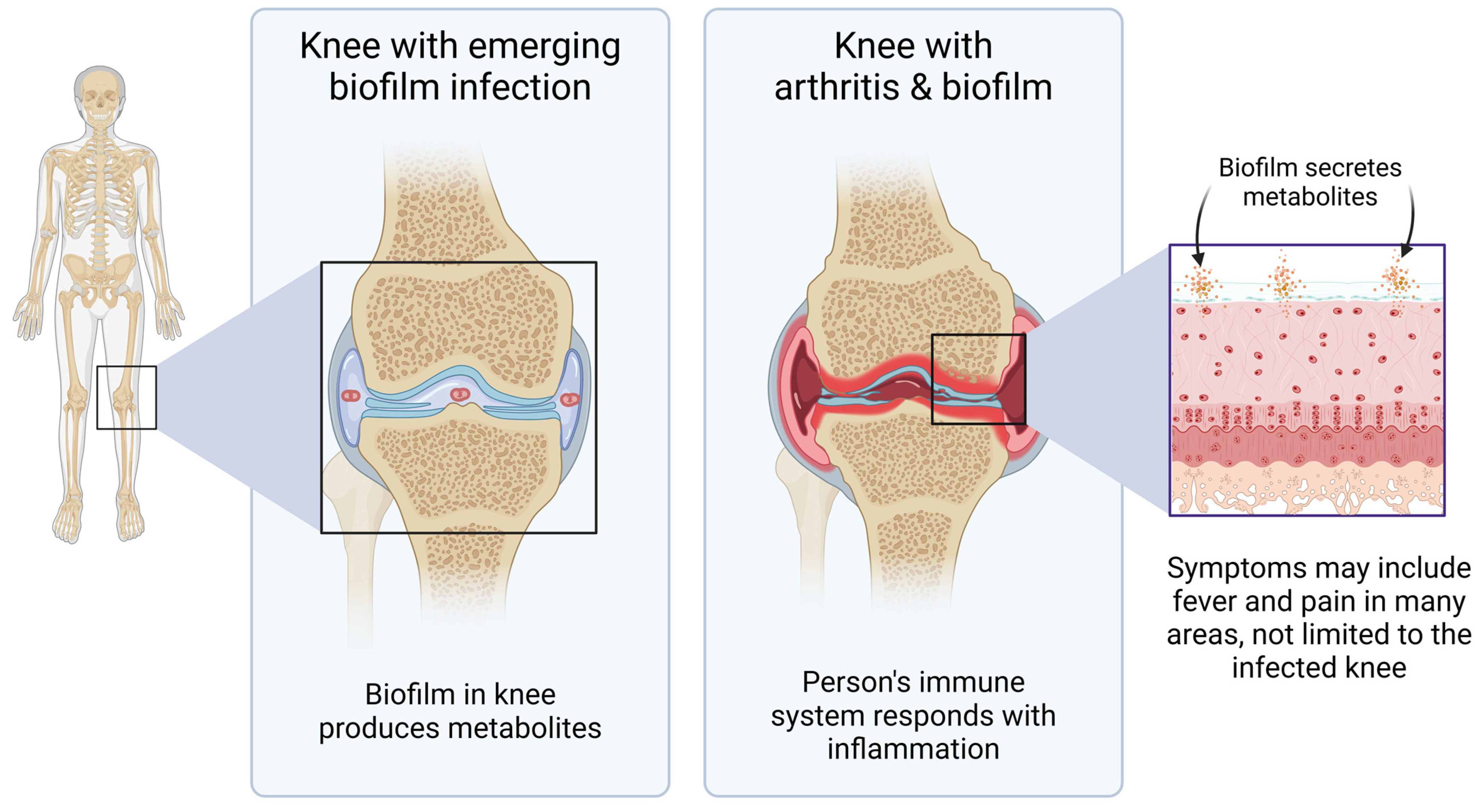
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De, S.; Hakansson, A.P. Measuring Niche-Associated Metabolic Activity in Planktonic and Biofilm Bacteria. Methods Mol. Biol. 2023, 2674, 3–32. [Google Scholar] [CrossRef] [PubMed]
- Hyde, J.A. Borrelia burgdorferi Keeps Moving and Carries on: A Review of Borrelial Dissemination and Invasion. Front. Immunol. 2017, 8, 114. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef]
- de Oliveira, A.; Cataneli Pereira, V.; Pinheiro, L.; Moraes Riboli, D.F.; Benini Martins, K.; de Souza da Cunha Mde, L.R. Antimicrobial Resistance Profile of Planktonic and Biofilm Cells of Staphylococcus aureus and Coagulase-Negative Staphylococci. Int. J. Mol. Sci. 2016, 17, 1423. [Google Scholar] [CrossRef] [PubMed]
- Olsen, I. Biofilm-specific antibiotic tolerance and resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 877–886. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, N.; Golovchenko, M.; Kybicova, K.; Vancova, M. Metamorphoses of Lyme disease spirochetes: Phenomenon of Borrelia persisters. Parasit. Vectors 2019, 12, 237. [Google Scholar] [CrossRef]
- Meriläinen, L.; Herranen, A.; Schwarzbach, A.; Gilbert, L. Morphological and biochemical features of Borrelia burgdorferi pleomorphic forms. Microbiol. Read. 2015, 161 Pt 3, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Hunfeld, K.P.; Kraiczy, P.; Norris, D.E.; Lohr, B. The In Vitro Antimicrobial Susceptibility of Borrelia burgdorferi sensu lato: Shedding Light on the Known Unknowns. Pathogens 2023, 12, 1204. [Google Scholar] [CrossRef] [PubMed]
- Di Domenico, E.G.; Cavallo, I.; Bordignon, V.; D’Agosto, G.; Pontone, M.; Trento, E.; Gallo, M.T.; Prignano, G.; Pimpinelli, F.; Toma, L.; et al. The Emerging Role of Microbial Biofilm in Lyme Neuroborreliosis. Front. Neurol. 2018, 9, 1048. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Akira, S. Toll-like receptors. Curr. Protoc. Immunol. 2015, 109, 14.12.1–14.12.10. [Google Scholar] [CrossRef] [PubMed]
- Vestby, L.K.; Grønseth, T.; Simm, R.; Nesse, L.L. Bacterial Biofilm and its Role in the Pathogenesis of Disease. Antibiotics 2020, 9, 59. [Google Scholar] [CrossRef] [PubMed]
- Sapi, E.; Bastian, S.L.; Mpoy, C.M.; Scott, S.; Rattelle, A.; Pabbati, N.; Poruri, A.; Burugu, D.; Theophilus, P.A.; Pham, T.V.; et al. Characterization of biofilm formation by Borrelia burgdorferi in vitro. PLoS ONE 2012, 7, e48277. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, S.; Hayes, D., Jr.; Wozniak, D.J. Cystic Fibrosis and Pseudomonas aeruginosa: The Host-Microbe Interface. Clin. Microbiol. Rev. 2019, 32, e00138-18. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Levin, J.M.; Nelson, J.A.; Segreti, J.; Harrison, B.; Benson, C.A.; Strle, F. In vitro susceptibility of Borrelia burgdorferi to 11 antimicrobial agents. Antimicrob. Agents Chemother. 1993, 37, 1444–1446. [Google Scholar] [CrossRef] [PubMed]
- Sapi, E.; Kaur, N.; Anyanwu, S.; Luecke, D.F.; Datar, A.; Patel, S.; Rossi, M.; Stricker, R.B. Evaluation of in-vitro antibiotic susceptibility of different morphological forms of Borrelia burgdorferi. Infect. Drug Resist. 2011, 4, 97–113. [Google Scholar] [PubMed]
- Roy, R.; Tiwari, M.; Donelli, G.; Tiwari, V. Strategies for combating bacterial biofilms: A focus on anti-biofilm agents and their mechanisms of action. Virulence 2018, 9, 522–554. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Zhang, S.; Shi, W.; Zubcevik, N.; Miklossy, J.; Zhang, Y. Selective Essential Oils from Spice or Culinary Herbs Have High Activity against Stationary Phase and Biofilm Borrelia burgdorferi. Front. Med. 2017, 4, 169. [Google Scholar] [CrossRef] [PubMed]
- Socarras, K.M.; Theophilus, P.A.S.; Torres, J.P.; Gupta, K.; Sapi, E. Antimicrobial Activity of Bee Venom and Melittin against Borrelia burgdorferi. Antibiotics 2017, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Rane, H.S.; Bernardo, S.M.; Howell, A.B.; Lee, S.A. Cranberry-derived proanthocyanidins prevent formation of Candida albicans biofilms in artificial urine through biofilm- and adherence-specific mechanisms. J. Antimicrob. Chemother. 2014, 69, 428–436. [Google Scholar] [CrossRef] [PubMed]
- Greene, A.C.; Acharya, A.P.; Lee, S.B.; Gottardi, R.; Zaleski, E.; Little, S.R. Cranberry extract-based formulations for preventing bacterial biofilms. Drug Deliv. Transl. Res. 2021, 11, 1144–1155. [Google Scholar] [CrossRef] [PubMed]
- Sandasi, M.; Leonard, C.M.; Viljoen, A.M. The in vitro antibiofilm activity of selected culinary herbs and medicinal plants against Listeria monocytogenes. Lett. Appl. Microbiol. 2010, 50, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Aswathanarayan, J.B.; Vittal, R.R. Inhibition of biofilm formation and quorum sensing mediated phenotypes by berberine in Pseudomonas aeruginosa and Salmonella typhimurium. RSC Adv. 2018, 8, 36133–36141. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yao, X.; Zhu, Z.; Tang, T.; Dai, K.; Sadovskaya, I.; Flahaut, S.; Jabbouri, S. Effect of berberine on Staphylococcus epidermidis biofilm formation. Int. J. Antimicrob. Agents 2009, 34, 60–66. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, W.; Cai, L.; Yang, T. Potentiation and Mechanism of Berberine as an Antibiotic Adjuvant Against Multidrug-Resistant Bacteria. Infect. Drug Resist. 2023, 16, 7313–7326. [Google Scholar] [CrossRef] [PubMed]
- Dinicola, S.; De Grazia, S.; Carlomagno, G.; Pintucci, J.P. N-acetylcysteine as powerful molecule to destroy bacterial biofilms. A systematic review. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 2942–2948. [Google Scholar] [PubMed]
- Zhao, T.; Liu, Y. N-acetylcysteine inhibit biofilms produced by Pseudomonas aeruginosa. BMC Microbiol. 2010, 10, 140. [Google Scholar] [CrossRef] [PubMed]
- Selan, L.; Papa, R.; Tilotta, M.; Vrenna, G.; Carpentieri, A.; Amoresano, A.; Pucci, P.; Artini, M. Serratiopeptidase: A well-known metalloprotease with a new non-proteolytic activity against S. aureus biofilm. BMC Microbiol. 2015, 15, 207. [Google Scholar] [CrossRef] [PubMed]
- Katsipis, G.; Pantazaki, A.A. Serrapeptase impairs biofilm, wall, and phospho-homeostasis of resistant and susceptible Staphylococcus aureus. Appl. Microbiol. Biotechnol. 2023, 107, 1373–1389. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Pan, S.; Erdmann, M.; Terrell, J.; Cabeen, M.T. A putative lipase affects Pseudomonas aeruginosa biofilm matrix production. mSphere 2023, 8, e0037423. [Google Scholar] [CrossRef] [PubMed]
- Kamali, E.; Jamali, A.; Izanloo, A.; Ardebili, A. In vitro activities of cellulase and ceftazidime, alone and in combination against Pseudomonas aeruginosa biofilms. BMC Microbiol. 2021, 21, 347. [Google Scholar] [CrossRef]
- Artini, M.; Vrenna, G.; Trecca, M.; Tuccio Guarna Assanti, V.; Fiscarelli, E.V.; Papa, R.; Selan, L. Serratiopeptidase Affects the Physiology of Pseudomonas aeruginosa Isolates from Cystic Fibrosis Patients. Int. J. Mol. Sci. 2022, 23, 12645. [Google Scholar] [CrossRef] [PubMed]
- Ferraboschi, P.; Ciceri, S.; Grisenti, P. Applications of Lysozyme, an Innate Immune Defense Factor, as an Alternative Antibiotic. Antibiotics 2021, 10, 1534. [Google Scholar] [CrossRef] [PubMed]
- Araújo, P.A.; Machado, I.; Meireles, A.; Leiknes, T.; Mergulhão, F.; Melo, L.F.; Simões, M. Combination of selected enzymes with cetyltrimethylammonium bromide in biofilm inactivation, removal and regrowth. Food Res. Int. 2017, 95, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Thallinger, B.; Prasetyo, E.N.; Nyanhongo, G.S.; Guebitz, G.M. Antimicrobial enzymes: An emerging strategy to fight microbes and microbial biofilms. Biotechnol. J. 2013, 8, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Jensen, G.S.; Cruickshank, D.; Hamilton, D.E. Disruption of Established Bacterial and Fungal Biofilms by a Blend of Enzymes and Botanical Extracts. J. Microbiol. Biotechnol. 2023, 33, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Glader, O.; Puljula, E.; Jokioja, J.; Karonen, M.; Sinkkonen, J.; Hytönen, J. NMR metabolome of Borrelia burgdorferi in vitro and in vivo in mice. Sci. Rep. 2019, 9, 8049. [Google Scholar] [CrossRef] [PubMed]
- Mielko, K.A.; Jabłoński, S.J.; Milczewska, J.; Sands, D.; Łukaszewicz, M.; Młynarz, P. Metabolomic studies of Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 2019, 35, 178. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.J.; Kielian, T. Biofilm-Leukocyte Cross-Talk: Impact on Immune Polarization and Immunometabolism. J. Innate Immun. 2019, 11, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Heim, C.E.; Vidlak, D.; Kielian, T. Interleukin-10 production by myeloid-derived suppressor cells contributes to bacterial persistence during Staphylococcus aureus orthopedic biofilm infection. J. Leukoc. Biol. 2015, 98, 1003–1013. [Google Scholar] [CrossRef] [PubMed]
- Macion, A.; Wyszyńska, A.; Godlewska, R. Delivery of Toxins and Effectors by Bacterial Membrane Vesicles. Toxins 2021, 13, 845. [Google Scholar] [CrossRef] [PubMed]
- Koenigs, A.; Hammerschmidt, C.; Jutras, B.L.; Pogoryelov, D.; Barthel, D.; Skerka, C.; Kugelstadt, D.; Wallich, R.; Stevenson, B.; Zipfel, P.F.; et al. BBA70 of Borrelia burgdorferi is a novel plasminogen-binding protein. J. Biol. Chem. 2013, 288, 25229–25243. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.R.; LeBlanc, K.T.; Leong, J.M. Fibronectin binding protein BBK32 of the Lyme disease spirochete promotes bacterial attachment to glycosaminoglycans. Infect. Immun. 2006, 74, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Scherr, T.D.; Hanke, M.L.; Huang, O.; James, D.B.; Horswill, A.R.; Bayles, K.W.; Fey, P.D.; Torres, V.J.; Kielian, T. Staphylococcus aureus Biofilms Induce Macrophage Dysfunction Through Leukocidin AB and Alpha-Toxin. mBio 2015, 6, e01021-15. [Google Scholar] [CrossRef] [PubMed]
- Medina-Rojas, M.; Stribling, W.; Snesrud, E.; Garry, B.I.; Li, Y.; Gann, P.M.; Demons, S.T.; Tyner, S.D.; Zurawski, D.V.; Antonic, V. Comparison of Pseudomonas aeruginosa strains reveals that Exolysin A toxin plays an additive role in virulence. Pathog. Dis. 2020, 78, ftaa010. [Google Scholar] [CrossRef] [PubMed]
- Iloba, I.; McGarry, S.V.; Yu, L.; Cruickshank, D.; Jensen, G.S. Differential Immune-Modulating Activities of Cell Walls and Secreted Metabolites from Probiotic Bacillus coagulans JBI-YZ6.3 under Normal versus Inflamed Culture Conditions. Microorganisms 2023, 11, 2564. [Google Scholar] [CrossRef] [PubMed]
- Schildberger, A.; Rossmanith, E.; Eichhorn, T.; Strassl, K.; Weber, V. Monocytes, peripheral blood mononuclear cells, and THP-1 cells exhibit different cytokine expression patterns following stimulation with lipopolysaccharide. Mediators Inflamm. 2013, 2013, 697972. [Google Scholar] [CrossRef]
- Phuong, M.S.; Hernandez, R.E.; Wolter, D.J.; Hoffman, L.R.; Sad, S. Impairment in inflammasome signaling by the chronic Pseudomonas aeruginosa isolates from cystic fibrosis patients results in an increase in inflammatory response. Cell Death Dis. 2021, 12, 241. [Google Scholar] [CrossRef] [PubMed]
- Butler, T. The Jarisch-Herxheimer Reaction After Antibiotic Treatment of Spirochetal Infections: A Review of Recent Cases and Our Understanding of Pathogenesis. Am. J. Trop. Med. Hyg. 2017, 96, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.; Zhang, N.; Wooten, R.M. Borrelia burgdorferi elicited-IL-10 suppresses the production of inflammatory mediators, phagocytosis, and expression of co-stimulatory receptors by murine macrophages and/or dendritic cells. PLoS ONE 2013, 8, e84980. [Google Scholar] [CrossRef] [PubMed]
- Janský, L.; Reymanová, P.; Kopecký, J. Dynamics of cytokine production in human peripheral blood mononuclear cells stimulated by LPS or infected by Borrelia. Physiol. Res. 2003, 52, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.; Brissette, C.A. The Brilliance of Borrelia: Mechanisms of Host Immune Evasion by Lyme Disease-Causing Spirochetes. Pathogens 2021, 10, 281. [Google Scholar] [CrossRef] [PubMed]
- Kroken, A.R.; Klein, K.A.; Mitchell, P.S.; Nieto, V.; Jedel, E.J.; Evans, D.J.; Fleiszig, S.M.J. Intracellular replication of Pseudomonas aeruginosa in epithelial cells requires suppression of the caspase-4 inflammasome. mSphere 2023, 8, e0035123. [Google Scholar] [CrossRef] [PubMed]
- Girschick, H.J.; Huppertz, H.I.; Rüssmann, H.; Krenn, V.; Karch, H. Intracellular persistence of Borrelia burgdorferi in human synovial cells. Rheumatol. Int. 1996, 16, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Jensen, G.S.; Benson, K.F. The blood as a diagnostic tool in chronic illness with obscure microbial involvement: A critical review. Int. J. Complement. Alt. Med. 2019, 12, 203–212. [Google Scholar] [CrossRef]
- Benson, K.F.; Jensen, G.S. Bacteria in blood from fibromyalgia patients include the Aquabacterium genus, producing metabolites with inflammatory properties in vitro. Results from a pilot study. Int. J. Complement. Alt. Med. 2019, 12, 232–239. [Google Scholar]
- Middelveen, M.J.; Filush, K.R.; Bandoski, C.; Kasliwala, R.S.; Melillo, A.; Stricker, R.B.; Sapi, E. Mixed Borrelia burgdorferi and Helicobacter pylori Biofilms in Morgellons Disease Dermatological Specimens. Healthcare 2019, 7, 70. [Google Scholar] [CrossRef] [PubMed]
- Sapi, E.; Gupta, K.; Wawrzeniak, K.; Gaur, G.; Torres, J.; Filush, K.; Melillo, A.; Zelger, B. Borrelia and Chlamydia Can Form Mixed Biofilms in Infected Human Skin Tissues. Eur. J. Microbiol. Immunol. 2019, 9, 46–55. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microbial Strain | Gram Staining | LPS 1 |
|---|---|---|
| Pseudomonas aeruginosa (Schroeter) Migula (ATCC® 9027) | Negative | Yes |
| Staphylococcus simulans Kloos and Schleif-er (ATCC® 11631) 2 | Positive | |
| Borrelia burgdorferi Strain B31 (ATCC® 35210) | Negative | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruickshank, D.; Hamilton, D.E.; Iloba, I.; Jensen, G.S. Secreted Metabolites from Pseudomonas, Staphylococcus, and Borrelia Biofilm: Modulation of Immunogenicity by a Nutraceutical Enzyme and Botanical Blend. Microorganisms 2024, 12, 991. https://doi.org/10.3390/microorganisms12050991
Cruickshank D, Hamilton DE, Iloba I, Jensen GS. Secreted Metabolites from Pseudomonas, Staphylococcus, and Borrelia Biofilm: Modulation of Immunogenicity by a Nutraceutical Enzyme and Botanical Blend. Microorganisms. 2024; 12(5):991. https://doi.org/10.3390/microorganisms12050991
Chicago/Turabian StyleCruickshank, Dina, Debby E. Hamilton, Ifeanyi Iloba, and Gitte S. Jensen. 2024. "Secreted Metabolites from Pseudomonas, Staphylococcus, and Borrelia Biofilm: Modulation of Immunogenicity by a Nutraceutical Enzyme and Botanical Blend" Microorganisms 12, no. 5: 991. https://doi.org/10.3390/microorganisms12050991
APA StyleCruickshank, D., Hamilton, D. E., Iloba, I., & Jensen, G. S. (2024). Secreted Metabolites from Pseudomonas, Staphylococcus, and Borrelia Biofilm: Modulation of Immunogenicity by a Nutraceutical Enzyme and Botanical Blend. Microorganisms, 12(5), 991. https://doi.org/10.3390/microorganisms12050991