
The Combination of Buchloe dactyloides Engelm and Biochar Promotes the Remediation of Soil Contaminated with Polycyclic Aromatic Hydrocarbons

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Pot Experiment
2.2. Determination of PAHs
2.3. Determination of Enzyme Activity
2.4. High-Throughput Sequencing and Quantitative Polymerase Chain Reaction (qPCR)
2.5. Soil Metabolite Assay
2.6. Date Analysis
3. Results
3.1. Degradation Rate of PAHs and Enzymatic Activity
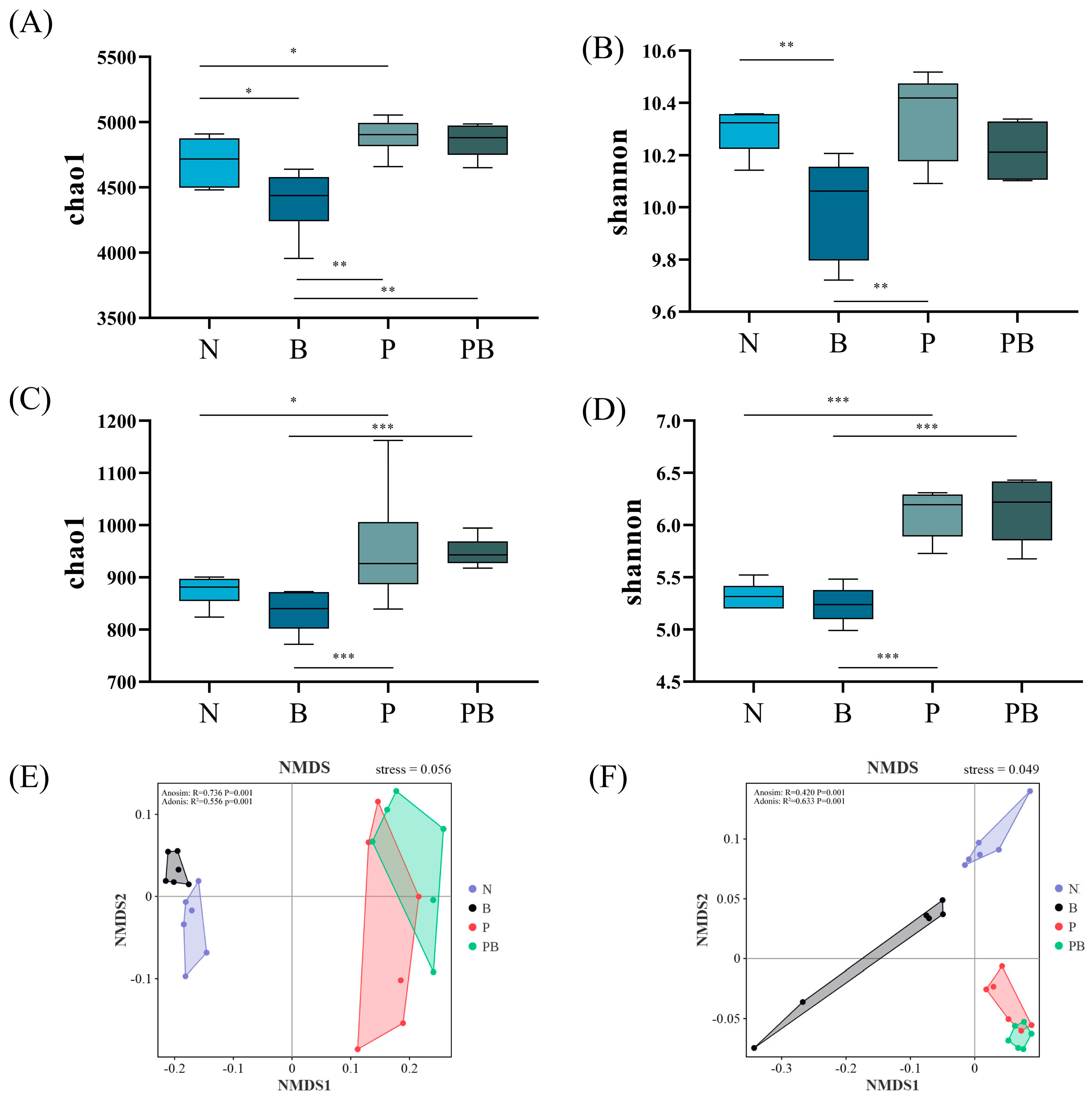
3.2. Shifts in Microbial Community and qPCR
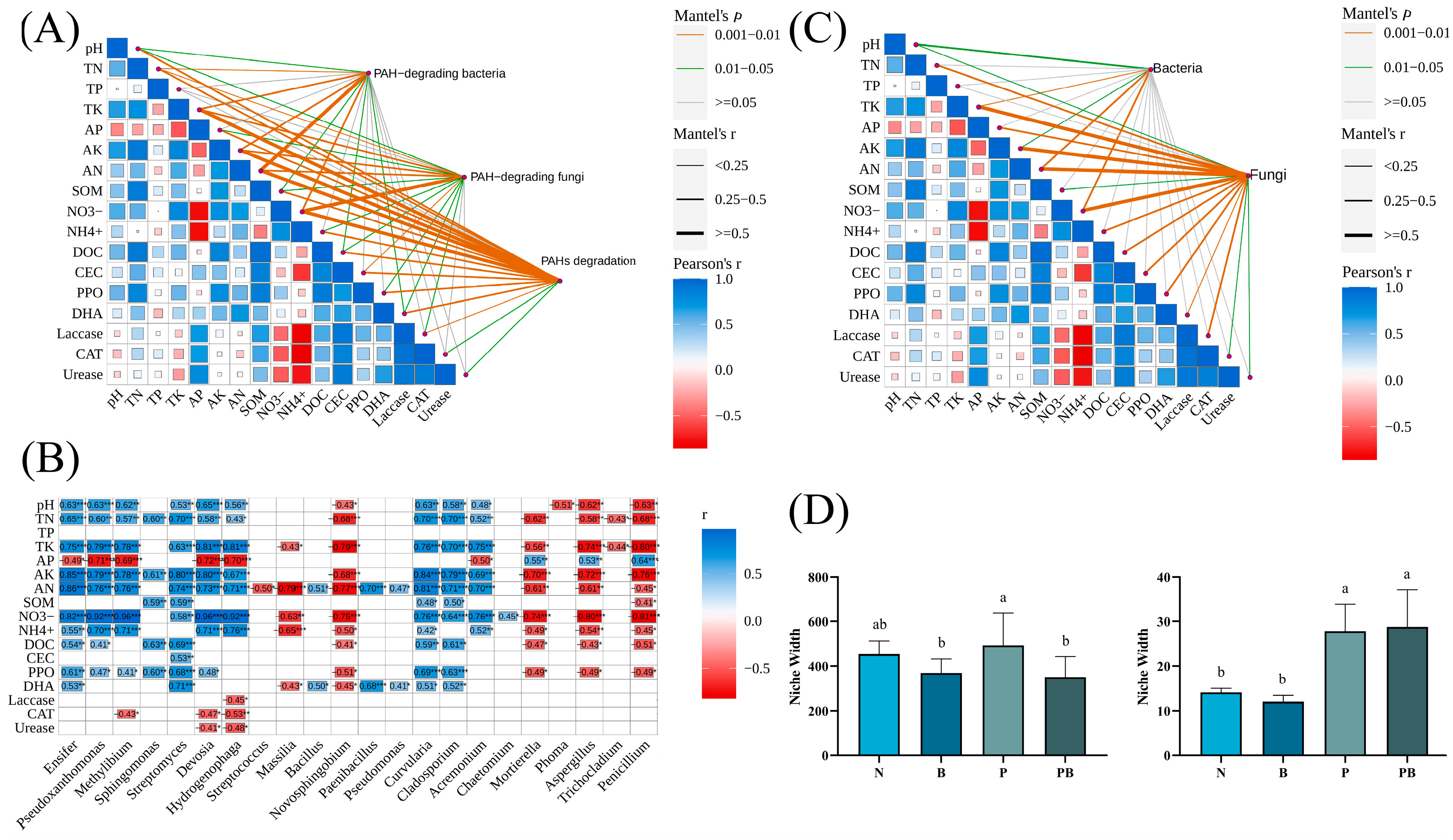
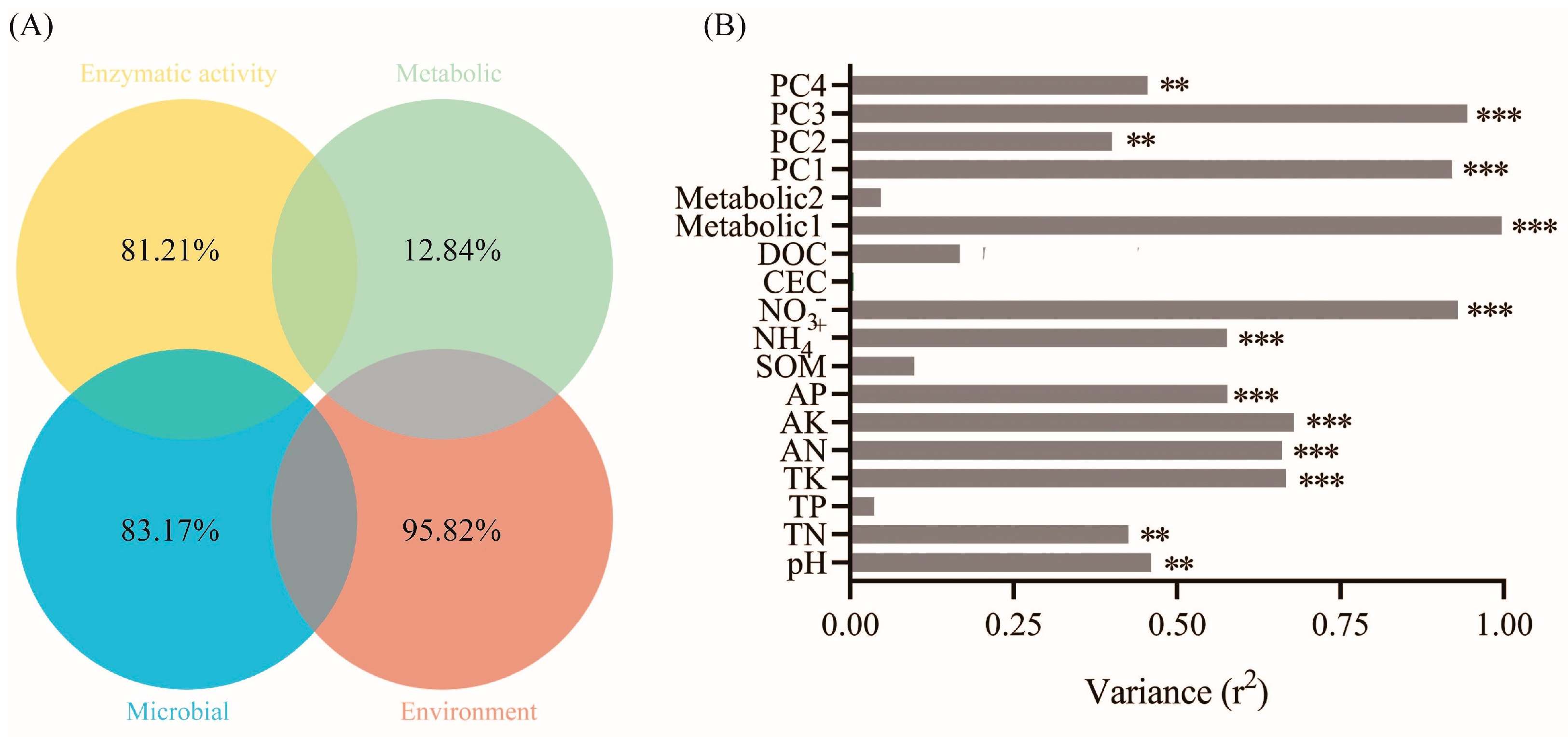
3.3. Environmental Factors and Function of Microbial Community
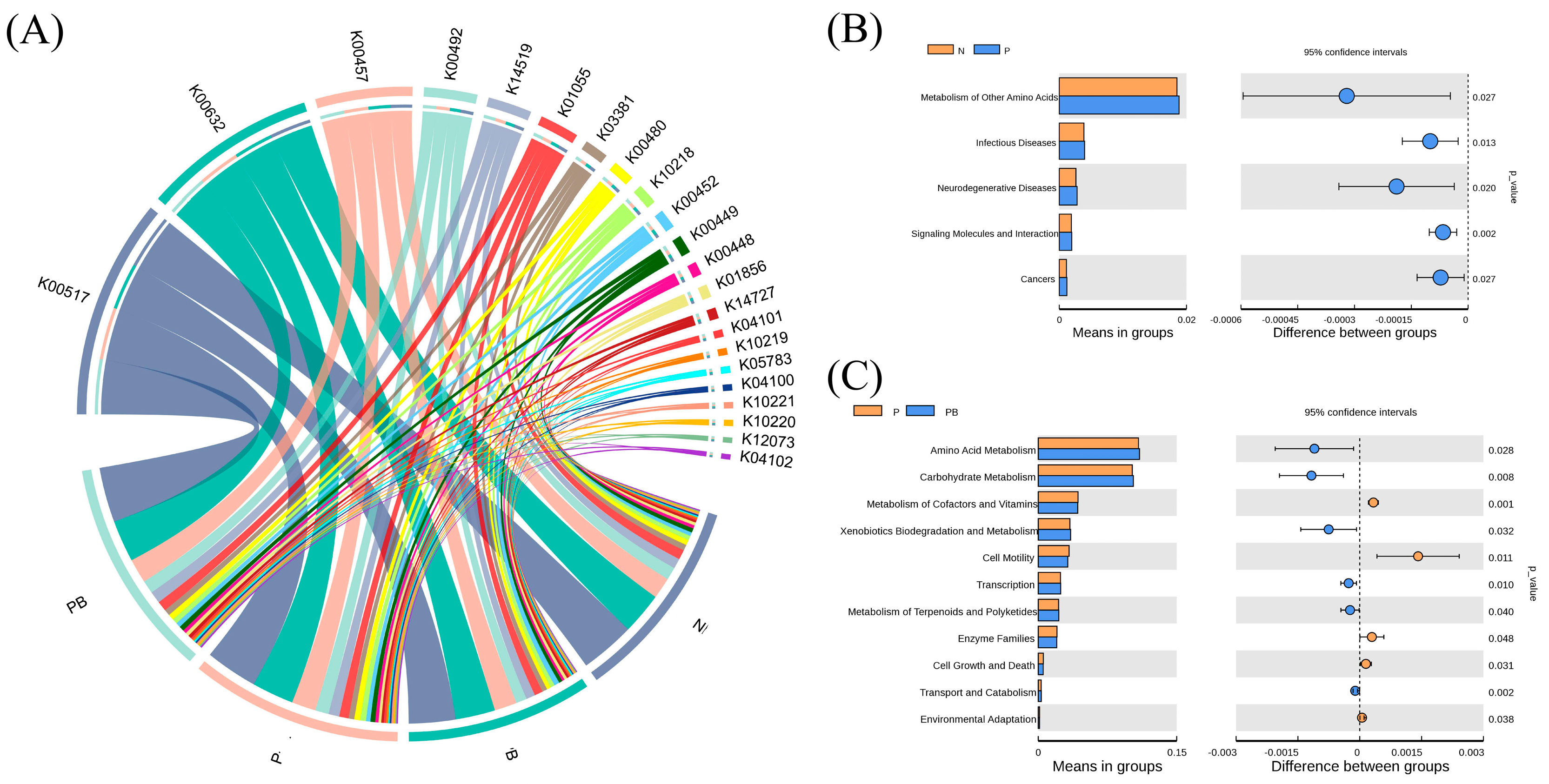
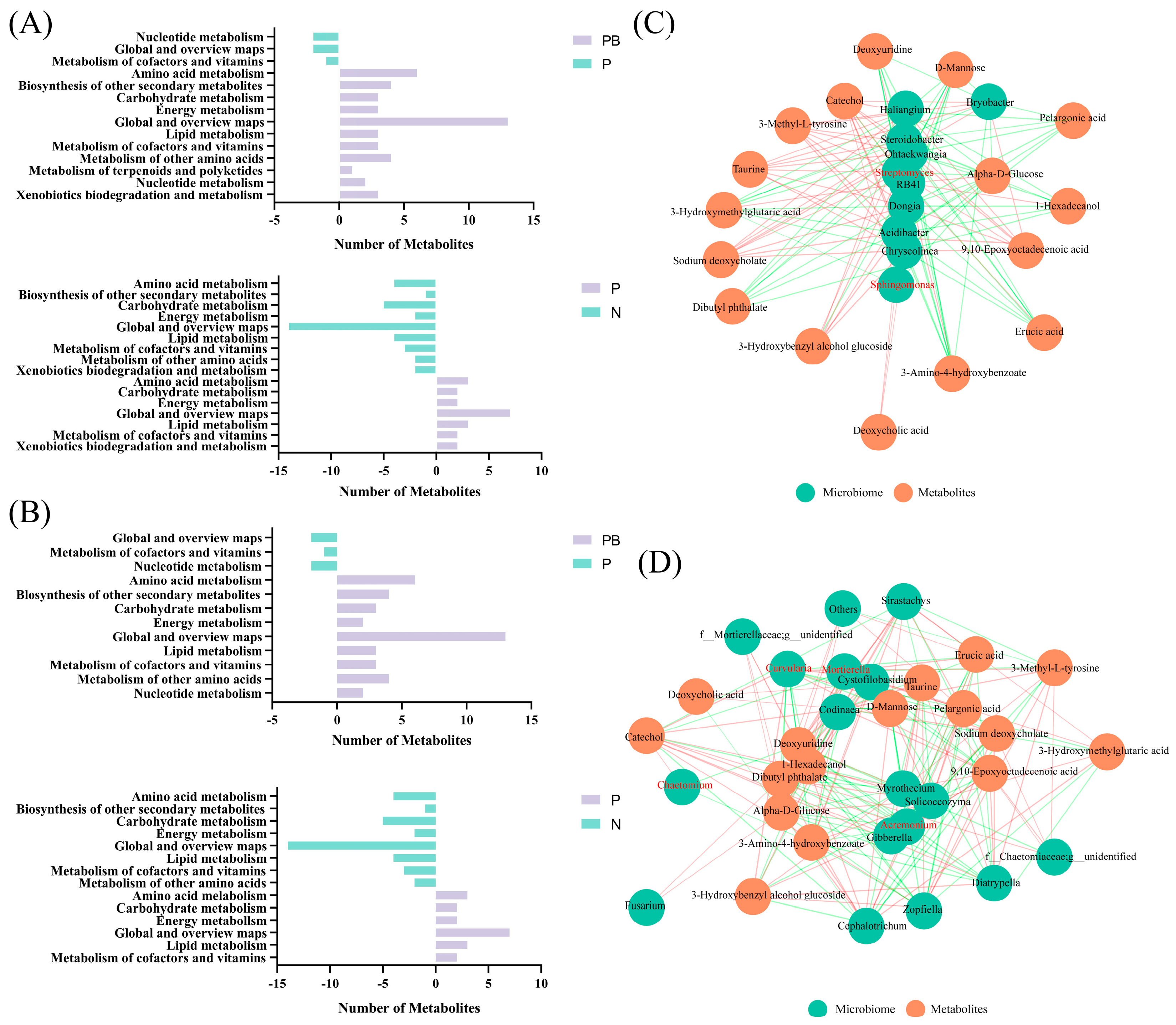
3.4. Soil Metabolism and Co-Occurrence Network Analysis
4. Discussion
4.1. The Effects of Biochar and Plant Roots on PAH Contaminated Soil and Microorganisms
4.2. The Effects of Biochar and Plant Roots on Soil Carbon Metabolism Associated with PAH Degradation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thacharodi, A.; Hassan, S.; Singh, T.; Mandal, R.; Chinnadurai, J.; Khan, H.A.; Hussain, M.A.; Brindhadevi, K.; Pugazhendhi, A. Bioremediation of Polycyclic Aromatic Hydrocarbons: An Updated Microbiological Review. Chemosphere 2023, 328, 138498. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qu, C.; Bian, Y.; Gu, C.; Jiang, X.; Song, Y. New Insights into the Responses of Soil Microorganisms to Polycyclic Aromatic Hydrocarbon Stress by Combining Enzyme Activity and Sequencing Analysis with Metabolomics. Environ. Pollut. 2019, 255, 113312. [Google Scholar] [CrossRef]
- Singha, L.P.; Pandey, P. Rhizosphere Assisted Bioengineering Approaches for the Mitigation of Petroleum Hydrocarbons Contamination in Soil. Crit. Rev. Biotechnol. 2021, 41, 749–766. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, M.; Tateno, R. Rhizosphere Effects on Soil Extracellular Enzymatic Activity and Microbial Abundance during the Low-Temperature Dormant Season in a Northern Hardwood Forest. Rhizosphere-Neth. 2022, 21, 100465. [Google Scholar] [CrossRef]
- Haichar, F.E.Z.; Santaella, C.; Heulin, T.; Achouak, W. Root Exudates Mediated Interactions Belowground. Soil Biol. Biochem. 2014, 77, 69–80. [Google Scholar] [CrossRef]
- Zhong, Y.; Xun, W.; Wang, X.; Tian, S.; Zhang, Y.; Li, D.; Zhou, Y.; Qin, Y.; Zhang, B.; Zhao, G.; et al. Root-Secreted Bitter Triterpene Modulates the Rhizosphere Microbiota to Improve Plant Fitness. Nat. Plants 2022, 8, 887–896. [Google Scholar] [CrossRef]
- Qi, D.; Feng, F.; Lu, C.; Fu, Y. C:N:P Stoichiometry of Different Soil Components after the Transition of Temperate Primary Coniferous and Broad-Leaved Mixed Forests to Secondary Forests. Soil Tillage Res. 2022, 216, 105260. [Google Scholar] [CrossRef]
- Hamid, Y.; Tang, L.; Hussain, B.; Usman, M.; Lin, Q.; Rashid, M.S.; He, Z.; Yang, X. Organic Soil Additives for the Remediation of Cadmium Contaminated Soils and Their Impact on the Soil-Plant System: A Review. Sci. Total Environ. 2020, 707, 136121. [Google Scholar] [CrossRef]
- Ji, M.; Wang, X.; Usman, M.; Liu, F.; Dan, Y.; Zhou, L.; Campanaro, S.; Luo, G.; Sang, W. Effects of Different Feedstocks-Based Biochar on Soil Remediation: A Review. Environ. Pollut. 2022, 294, 118655. [Google Scholar] [CrossRef]
- Li, X.; Yao, S.; Bian, Y.; Jiang, X.; Song, Y. The Combination of Biochar and Plant Roots Improves Soil Bacterial Adaptation to PAH Stress: Insights from Soil Enzymes, Microbiome, and Metabolome. J. Hazard. Mater. 2020, 400, 123227. [Google Scholar] [CrossRef]
- Quilliam, R.S.; Rangecroft, S.; Emmett, B.A.; Deluca, T.H.; Jones, D.L. Is Biochar a Source or Sink for Polycyclic Aromatic Hydrocarbon (PAH) Compounds in Agricultural Soils? GCB Bioenergy 2013, 5, 96–103. [Google Scholar] [CrossRef]
- Hao, Z.; Wang, Q.; Yan, Z.; Jiang, H. Novel Magnetic Loofah Sponge Biochar Enhancing Microbial Responses for the Remediation of Polycyclic Aromatic Hydrocarbons-Contaminated Sediment. J. Hazard. Mater. 2021, 401, 123859. [Google Scholar] [CrossRef]
- Ali, S.; Rizwan, M.; Qayyum, M.F.; Ok, Y.S.; Ibrahim, M.; Riaz, M.; Arif, M.S.; Hafeez, F.; Al-Wabel, M.I.; Shahzad, A.N. Biochar Soil Amendment on Alleviation of Drought and Salt Stress in Plants: A Critical Review. Environ. Sci. Pollut. Res. 2017, 24, 12700–12712. [Google Scholar] [CrossRef]
- Li, X.; Song, Y.; Bian, Y.; Gu, C.; Yang, X.; Wang, F.; Jiang, X. Insights into the Mechanisms Underlying Efficient Rhizodegradation of PAHs in Biochar-Amended Soil: From Microbial Communities to Soil Metabolomics. Ecol. Indic. 2020, 144, 105995. [Google Scholar] [CrossRef] [PubMed]
- Bornø, M.L.; Müller-Stöver, D.S.; Liu, F. Biochar Modifies the Content of Primary Metabolites in the Rhizosphere of Well-Watered and Drought-Stressed Zea Mays L. (Maize). Biol. Fertil. Soils 2022, 58, 633–647. [Google Scholar] [CrossRef]
- Wang, Y.; Fang, L.; Lin, L.; Luan, T.; Tam, N.F.Y. Effects of Low Molecular-Weight Organic Acids and Dehydrogenase Activity in Rhizosphere Sediments of Mangrove Plants on Phytoremediation of Polycyclic Aromatic Hydrocarbons. Chemosphere 2014, 99, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Guo, X.; Zhu, Y.; Liu, X.; Han, Z.; Sun, K.; Ji, L.; He, Q.; Han, L. The Effects of Different Biochars on Microbial Quantity, Microbial Community Shift, Enzyme Activity, and Biodegradation of Polycyclic Aromatic Hydrocarbons in Soil. Geoderma 2018, 328, 100–108. [Google Scholar] [CrossRef]
- Kramer-Walter, K.R.; Laughlin, D.C. Root Nutrient Concentration and Biomass Allocation Are More Plastic than Morphological Traits in Response to Nutrient Limitation. Plant Soil 2017, 416, 539–550. [Google Scholar] [CrossRef]
- Song, X.; Razavi, B.S.; Ludwig, B.; Zamanian, K.; Zang, H.; Kuzyakov, Y.; Dippold, M.A.; Gunina, A. Combined Biochar and Nitrogen Application Stimulates Enzyme Activity and Root Plasticity. Sci. Total Environ. 2020, 735, 139393. [Google Scholar] [CrossRef]
- Wan, H.; Liu, X.; Shi, Q.; Chen, Y.; Jiang, M.; Zhang, J.; Cui, B.; Hou, J.; Wei, Z.; Hossain, M.A.; et al. Biochar Amendment Alters Root Morphology of Maize Plant: Its Implications in Enhancing Nutrient Uptake and Shoot Growth under Reduced Irrigation Regimes. Front. Plant Sci. 2023, 14, 1122742. [Google Scholar] [CrossRef]
- Zhu, H.; Gao, Y.; Li, D. Germination of Grass Species in Soil Affected by Crude Oil Contamination. Int. J. Phytoremediat. 2018, 20, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Li, X.; Liu, J.; Cheng, Y.; Zou, J.; Zhai, F.; Sun, Z.; Han, L. Soil Microbial Community Succession and Interactions during Combined Plant/White-Rot Fungus Remediation of Polycyclic Aromatic Hydrocarbons. Sci. Total Environ. 2021, 752, 142224. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.; Hernandez, T.; Costa, F. Potential Use of Dehydrogenase Activity as an Index of Microbial Activity in Degraded Soils. Commun. Soil Sci. Plant Anal. 1997, 28, 123–134. [Google Scholar] [CrossRef]
- Zelena, E.; Dunn, W.B.; Broadhurst, D.; Francis-McIntyre, S.; Carroll, K.M.; Begley, P.; O’Hagan, S.; Knowles, J.D.; Halsall, A.; HUSERMET Consortium; et al. Development of a Robust and Repeatable UPLC−MS Method for the Long-Term Metabolomic Study of Human Serum. Anal. Chem. 2009, 81, 1357–1364. [Google Scholar] [CrossRef] [PubMed]
- Want, E.J.; Masson, P.; Michopoulos, F.; Wilson, I.D.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Loftus, N.; Holmes, E.; Nicholson, J.K. Global Metabolic Profiling of Animal and Human Tissues via UPLC-MS. Nat. Protoc. 2013, 8, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Vasilev, N.; Boccard, J.; Lang, G.; Grömping, U.; Fischer, R.; Goepfert, S.; Rudaz, S.; Schillberg, S. Structured Plant Metabolomics for the Simultaneous Exploration of Multiple Factors. Sci. Rep. 2016, 6, 37390. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Liu, R.; Zhou, Y.; Li, N.; Hou, L.; Ma, Q.; Gao, B. Fire Phoenix Facilitates Phytoremediation of PAH-Cd Co-Contaminated Soil through Promotion of Beneficial Rhizosphere Bacterial Communities. Environ. Int. 2020, 136, 105421. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Yang, B.; Song, Z.; Wang, H.; He, F.; Han, X. Wheat Straw Biochar Amendments on the Removal of Polycyclic Aromatic Hydrocarbons (PAHs) in Contaminated Soil. Ecotoxicol. Environ. Saf. 2016, 130, 248–255. [Google Scholar] [CrossRef]
- Shi, W.; Zhang, X.; Jia, H.; Feng, S.; Yang, Z.; Zhao, O.; Li, Y. Effective Remediation of Aged HMW-PAHs Polluted Agricultural Soil by the Combination of Fusarium sp. and Smooth Bromegrass (Bromus inermis Leyss.). J. Integr. Agric. 2017, 16, 199–209. [Google Scholar] [CrossRef]
- Wang, X.; Chen, J.; Wang, Y.; Li, Y.; Chen, S.; Zhang, Q. Promoting Resuscitation of the VBNC Bacteria and Enrichment of Naphthalene-Degrading Bacteria from Oil-Contaminated Soils with the Rpf4 from Rhodococcus Erythropolis KB1. Geomicrobiol. J. 2022, 39, 916–924. [Google Scholar] [CrossRef]
- Lin, X.; Qiao, B.; Chang, R.; Li, Y.; Zheng, W.; He, Z.; Tian, Y. Characterization of Two Keystone Taxa, Sulfur-Oxidizing, and Nitrate-Reducing Bacteria, by Tracking Their Role Transitions in the Benzo[a]Pyrene Degradative Microbiome. Microbiome 2023, 11, 139. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Niu, X.; Zhang, N.; Li, T. Effect of Biochar-Immobilized Sphingomonas Sp. PJ2 on Bioremediation of PAHs and Bacterial Community Composition in Saline Soil. Chemosphere 2021, 279, 130427. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Tang, H.; Wang, Y.Z.; Yang, C.; Gao, M.-T.; Tsang, Y.F.; Li, J. Effect of Dissolved Solids Released from Biochar on Soil Microbial Metabolism. Environ. Sci. Process. Impacts 2022, 598–608. [Google Scholar] [CrossRef] [PubMed]
- Mei, J.; Li, B.; Su, L.; Zhou, X.; Duan, E. Effects of Potassium Persulfate on Nitrogen Loss and Microbial Community during Cow Manure and Corn Straw Composting. Bioresour. Technol. 2022, 363, 127919. [Google Scholar] [CrossRef] [PubMed]
- Shakoor, N.; Hussain, M.; Adeel, M.; Azeem, I.; Ahmad, M.A.; Zain, M.; Zhang, P.; Li, Y.; Quanlong, W.; Horton, R.; et al. Lithium-Induced Alterations in Soybean Nodulation and Nitrogen Fixation through Multifunctional Mechanisms. Sci. Total Environ. 2023, 904, 166438. [Google Scholar] [CrossRef] [PubMed]
- Baoune, H.; Aparicio, J.D.; Acuña, A.; El Hadj-khelil, A.O.; Sanchez, L.; Polti, M.A.; Alvarez, A. Effectiveness of the Zea Mays-Streptomyces Association for the Phytoremediation of Petroleum Hydrocarbons Impacted Soils. Ecotoxicol. Environ. Saf. 2019, 184, 109591. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, Q.; Xie, S. Stable Isotope Probing Identifies Anthracene Degraders under Methanogenic Conditions. Biodegradation 2012, 23, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Singha, L.P.; Pandey, P. Rhizobacterial Community of Jatropha Curcas Associated with Pyrene Biodegradation by Consortium of PAH-Degrading Bacteria. Appl. Soil Ecol. 2020, 155, 103685. [Google Scholar] [CrossRef]
- Wang, X.; Teng, Y.; Wang, X.; Li, X.; Luo, Y. Microbial Diversity Drives Pyrene Dissipation in Soil. Sci. Total Environ. 2022, 819, 153082. [Google Scholar] [CrossRef]
- Zhang, X.-X.; Cheng, S.-P.; Zhu, C.-J.; Sun, S.-L. Microbial PAH-Degradation in Soil: Degradation Pathways and Contributing Factors1 1Project Supported by the National High Technology Research and Development Program (863 Program) of China (No. 2001AA214191). Pedosphere 2006, 16, 555–565. [Google Scholar] [CrossRef]
- Li, X.; Song, Y.; Wang, F.; Bian, Y.; Jiang, X. Combined Effects of Maize Straw Biochar and Oxalic Acid on the Dissipation of Polycyclic Aromatic Hydrocarbons and Microbial Community Structures in Soil: A Mechanistic Study. J. Hazard. Mater. 2018, 364, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, A.; Dennis, P.G.; Forstner, C.; Raghavendra, A.K.H.; Mathesius, U.; Richardson, A.E.; Delhaize, E.; Gilliham, M.; Watt, M.; Ryan, P.R. Manipulating Exudate Composition from Root Apices Shapes the Microbiome throughout the Root System. Plant Physiol. 2021, 187, 2279–2295. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Hu, X.; Zhou, Z.; Zhang, W.; Wang, Y.; Sun, B. Phytoavailability and Mechanism of Bound PAH Residues in Filed Contaminated Soils. Environ. Pollut. 2017, 222, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Badejo, A.C.; Choi, C.-W.; Badejo, A.O.; Shin, K.-H.; Hyun, J.-H.; Lee, Y.-G.; Kim, S.; Park, K.-S.; Kim, S.H.; Jung, K.H.; et al. A Global Proteome Study of Mycobacterium Gilvum PYR-GCK Grown on Pyrene and Glucose Reveals the Activation of Glyoxylate, Shikimate and Gluconeogenetic Pathways through the Central Carbon Metabolism Highway. Biodegradation 2013, 24, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Ressler, B.P.; Kneifel, H.; Winter, J. Bioavailability of Polycyclic Aromatic Hydrocarbons and Formation of Humic Acid-like Residues during Bacterial PAH Degradation. Appl. Microbiol. Biotechnol. 1999, 85–91. [Google Scholar] [CrossRef]
- Wang, S.; Guo, S. Effects of Soil Organic Carbon Metabolism on Electro-Bioremediation of Petroleum-Contaminated Soil. J. Hazard. Mater. 2023, 459, 132180. [Google Scholar] [CrossRef] [PubMed]
- Techer, D.; D’Innocenzo, M.; Laval-Gilly, P.; Henry, S.; Bennasroune, A.; Martinez-Chois, C.; Falla, J. Assessment of Miscanthus×giganteus Secondary Root Metabolites for the Biostimulation of PAH-Utilizing Soil Bacteria. Appl. Soil Ecol. 2012, 62, 142–146. [Google Scholar] [CrossRef]
- Zhao, W.; Li, S.; Dong, L.; Wang, P.; Lu, X.; Zhang, X.; Su, Z.; Guo, Q.; Ma, P. Effects of Different Crop Rotations on the Incidence of Cotton Verticillium Wilt and Structure and Function of the Rhizospheric Microbial Community. Plant Soil 2023, 485, 457–474. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Li, A.; Zou, B.; Qian, Y.; Li, X.; Sun, Z. The Combination of Buchloe dactyloides Engelm and Biochar Promotes the Remediation of Soil Contaminated with Polycyclic Aromatic Hydrocarbons. Microorganisms 2024, 12, 968. https://doi.org/10.3390/microorganisms12050968
Wang Y, Li A, Zou B, Qian Y, Li X, Sun Z. The Combination of Buchloe dactyloides Engelm and Biochar Promotes the Remediation of Soil Contaminated with Polycyclic Aromatic Hydrocarbons. Microorganisms. 2024; 12(5):968. https://doi.org/10.3390/microorganisms12050968
Chicago/Turabian StyleWang, Yuancheng, Ao Li, Bokun Zou, Yongqiang Qian, Xiaoxia Li, and Zhenyuan Sun. 2024. "The Combination of Buchloe dactyloides Engelm and Biochar Promotes the Remediation of Soil Contaminated with Polycyclic Aromatic Hydrocarbons" Microorganisms 12, no. 5: 968. https://doi.org/10.3390/microorganisms12050968
APA StyleWang, Y., Li, A., Zou, B., Qian, Y., Li, X., & Sun, Z. (2024). The Combination of Buchloe dactyloides Engelm and Biochar Promotes the Remediation of Soil Contaminated with Polycyclic Aromatic Hydrocarbons. Microorganisms, 12(5), 968. https://doi.org/10.3390/microorganisms12050968