
Establishment of an ELISA Based on a Recombinant Antigenic Protein Containing Multiple Prominent Epitopes for Detection of African Swine Fever Virus Antibodies

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Standard Controls
2.2. Serum Samples Collection
2.3. Epitopes Prediction
2.4. Construction of the Multi-Epitope Protein k3
2.5. Expression of Multi-Epitope Protein k3 in E. coli
2.6. Purification of Multi-Epitope Protein k3
2.7. Establishment and Evaluation of Indirect ELISA of k3 Protein
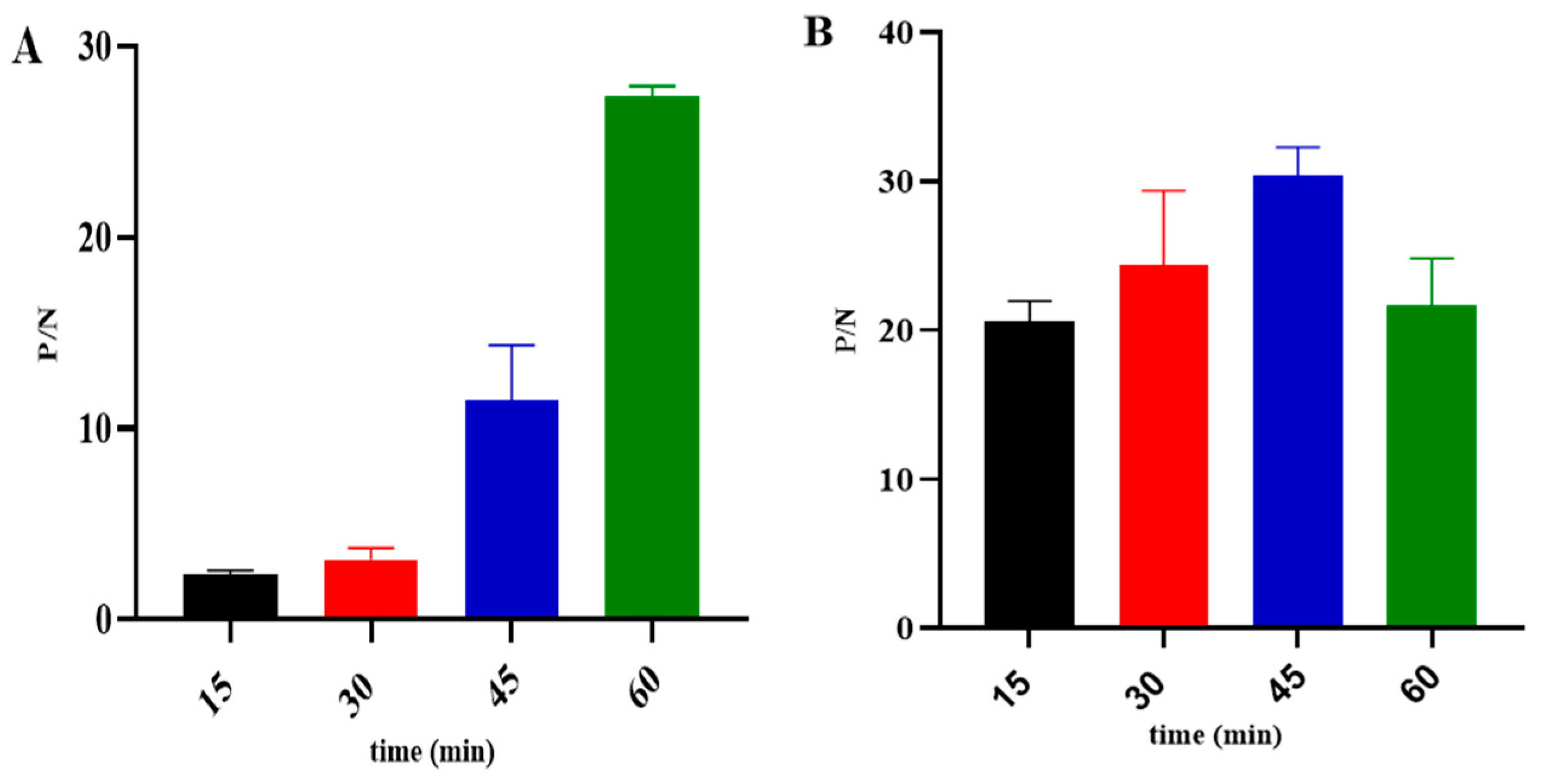
2.7.1. Matrix Titration
2.7.2. Optimization of Experimental Parameters
2.7.3. Determination of the Cutoff Value in k3-iELISA
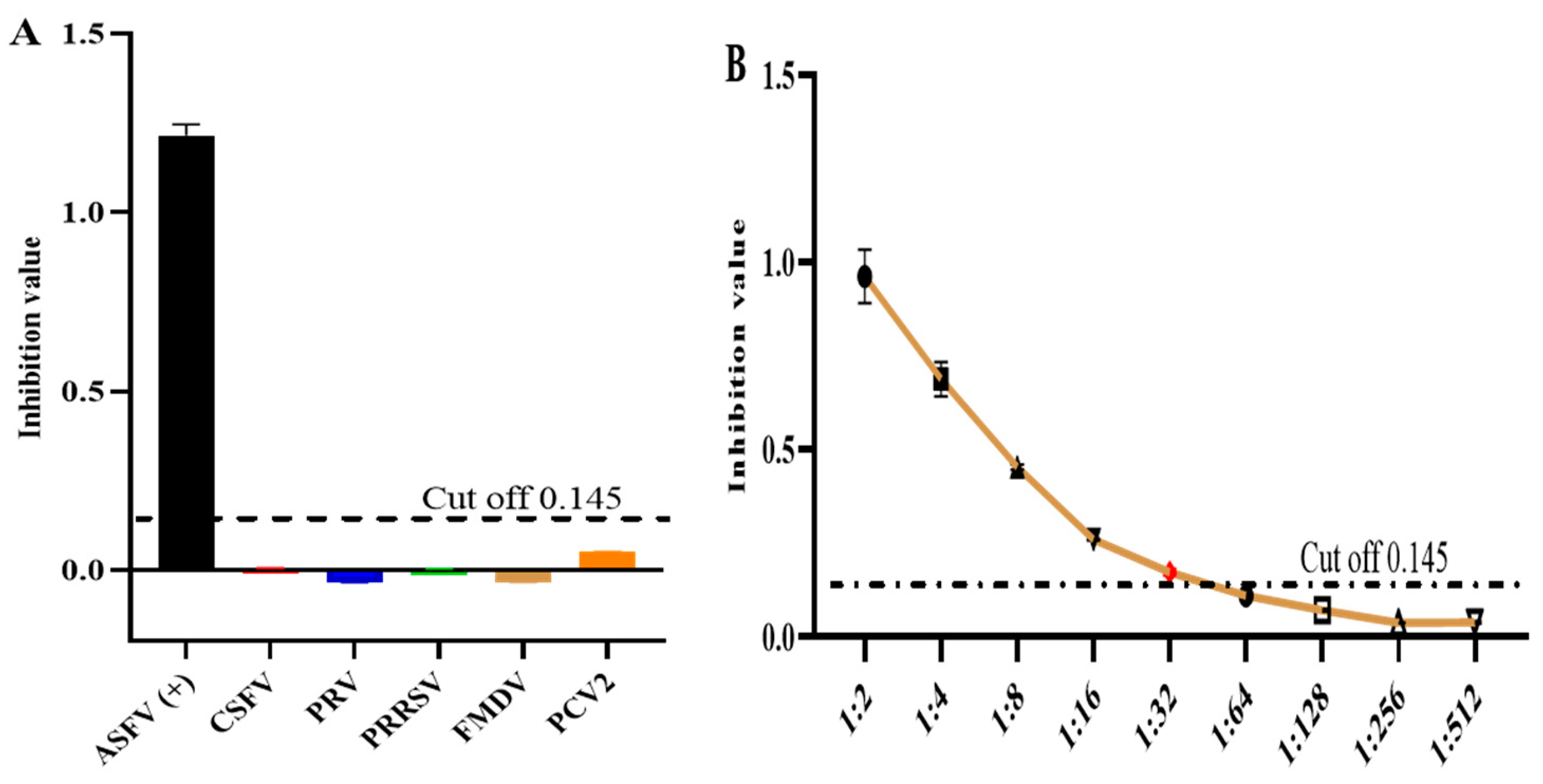
2.7.4. Specificity and Sensitivity Tests
2.8. Detection of Clinical Samples and Comparison with the Commercial Kit
2.9. Analyses
3. Results
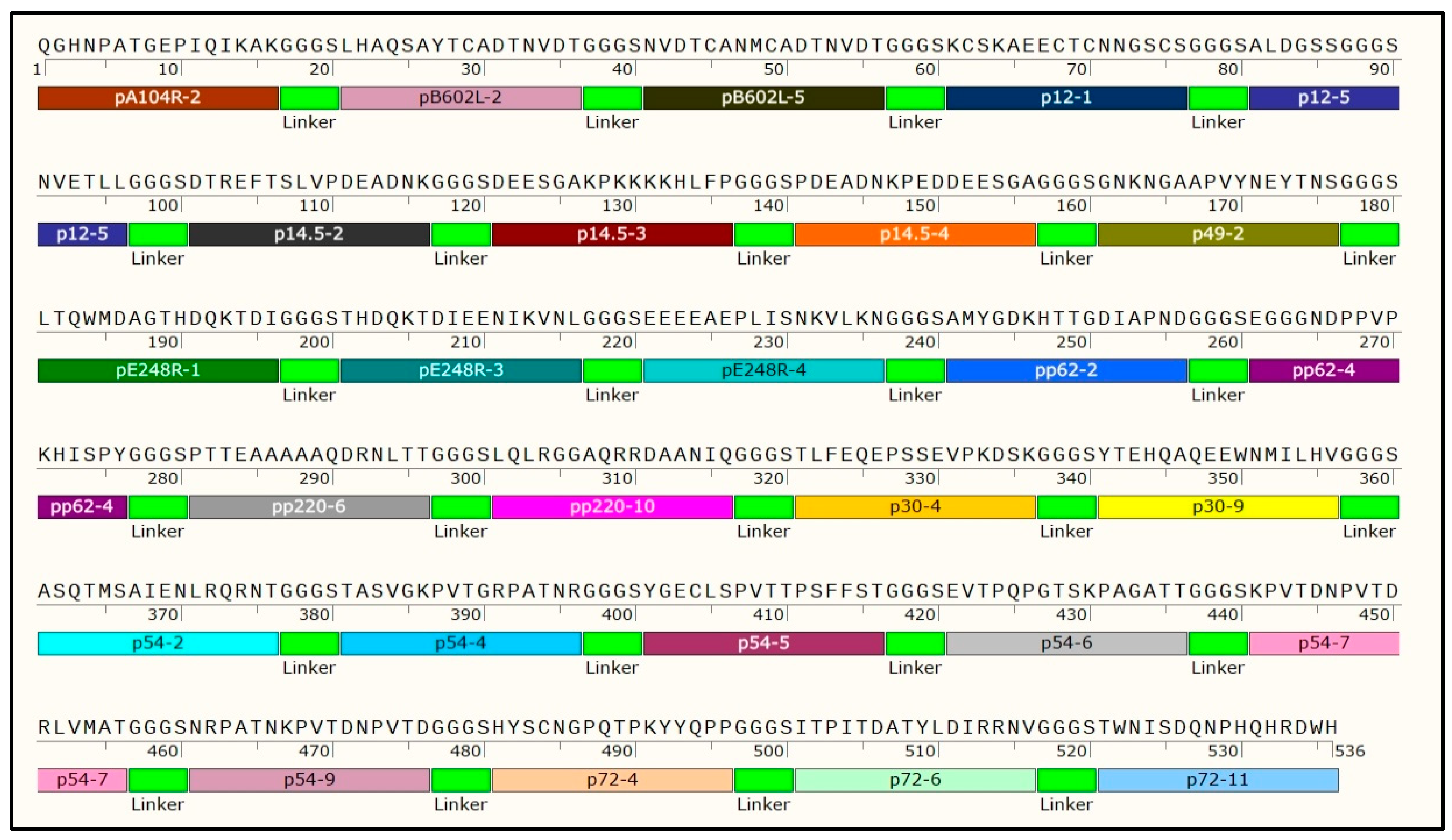
3.1. Screening of the Predominant Peptides
3.2. Expression, Purification, and Characterization of Recombinant Protein k3
3.3. Development of the k3-ELISA Method
3.4. Cutoff Value Evaluation
3.5. Assessment of Specificity and Sensitivity in k3-ELISA
3.6. Comparison of the k3-ELISA Method to Commercial Kit
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, D.; Zhang, Q.; Liu, Y.; Wang, M.; Zhang, L.; Han, L.; Chu, X.; Ding, G.; Li, Y.; Hou, Y.; et al. Indirect ELISA Using Multi-Antigenic Dominants of p30, p54 and p72 Recombinant Proteins to Detect Antibodies against African Swine Fever Virus in Pigs. Viruses 2022, 14, 2660. [Google Scholar] [CrossRef] [PubMed]
- Dixon, L.K.; Chapman, D.A.G.; Netherton, C.L.; Upton, C. African swine fever virus replication and genomics. Virus Res. 2013, 173, 3–14. [Google Scholar] [CrossRef]
- Gallardo, M.C.; Reoyo, A.T.; Fernández-Pinero, J.; Iglesias, I.; Muñoz, M.J.; Arias, M.L. African swine fever: A global view of the current challenge. Porc. Health Manag. 2015, 1, 21. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, S.; Zhang, H.; Qin, Z.; Shan, H.; Cai, X. Vaccines for African swine fever: An update. Front. Microbiol. 2023, 14, 1139494. [Google Scholar] [CrossRef] [PubMed]
- Jia, N.; Ou, Y.; Pejsak, Z.; Zhang, Y.; Zhang, J. Roles of African Swine Fever Virus Structural Proteins in Viral Infection. J. Vet. Res. 2017, 61, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, C.; Fernández-Pinero, J.; Arias, M. African swine fever (ASF) diagnosis, an essential tool in the epidemiological investigation. Virus Res. 2019, 271, 197676. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Qiao, S.; Li, G.; Tong, W.; Dong, S.; Liu, J.; Guo, Z.; Zheng, H.; Zhao, R.; Tong, G.; et al. The Indirect ELISA and Monoclonal Antibody against African Swine Fever Virus p17 Revealed Efficient Detection and Application Prospects. Viruses 2022, 15, 50. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.S.; Shao, N.; Chen, J.W.; Qi, W.B.; Li, Y.; Liu, P.; Chen, Y.J.; Bian, S.Y.; Zhang, Y.; Tao, S.C. Multiplex and visual detection of African Swine Fever Virus (ASFV) based on Hive-Chip and direct loop-mediated isothermal amplification. Anal. Chim. Acta 2020, 1140, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Q.; Guo, J.; Li, D.; Wang, L.; Wang, X.; Xing, G.; Deng, R.; Zhang, G. An Isothermal Molecular Point of Care Testing for African Swine Fever Virus Using Recombinase-Aided Amplification and Lateral Flow Assay without the Need to Extract Nucleic Acids in Blood. Front. Cell. Infect. Microbiol. 2021, 11, 633763. [Google Scholar] [CrossRef] [PubMed]
- Ward, M.P.; Tian, K.; Nowotny, N. African Swine Fever, the forgotten pandemic. Trans. Emerg. Dis. 2021, 68, 2637–2639. [Google Scholar] [CrossRef]
- Hu, L.; Lin, X.Y.; Yang, Z.X.; Yao, X.P.; Li, G.L.; Peng, S.Z.; Wang, Y. A multiplex PCR for simultaneous detection of classical swine fever virus, African swine fever virus, highly pathogenic porcine reproductive and respiratory syndrome virus, porcine reproductive and respiratory syndrome virus and pseudorabies in swines. Pol. J. Vet. Sci. 2015, 18, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Carmina, G.; Nieto, R.; Arias, M. African Swine Fever Virus (ASFV) Indirect ELISA Test Based on the Use of the Soluble Cytoplasmic Semi- purified Antigen (ASFV CP-Ag). Methods Mol. Biol. 2022, 2503, 133–145. [Google Scholar] [PubMed]
- Oh, T.; Do, D.T.; Lai, D.C.; Nguyen, L.T.; Lee, J.Y.; Van Le, P.; Chae, C. Chronological expression and distribution of African swine fever virus p30 and p72 proteins in experimentally infected pigs. Sci. Rep. 2022, 12, 4151. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Kitamura, T.; Nagata, K.; Ikezawa, M.; Kameyama, K.I.; Masujin, K.; Kokuho, T. Development of a Novel Indirect ELISA for the Serological Diagnosis of African Swine Fever Using p11.5 Protein as a Target Antigen. Pathogens 2023, 12, 774. [Google Scholar] [CrossRef]
- Muñoz, A.L.; Tabarés, E. Characteristics of the major structural proteins of African swine fever virus: Role as antigens in the induction of neutralizing antibodies. A review. Virology 2022, 571, 46–51. [Google Scholar] [CrossRef]
- Wu, Z.; Lu, H.; Zhu, D.; Xie, J.; Sun, F.; Xu, Y.; Zhang, H.; Wu, Z.; Xia, W.; Zhu, S. Developing an Indirect ELISA for the Detection of African Swine Fever Virus Antibodies Using a Tag-Free p15 Protein Antigen. Viruses 2023, 15, 1939. [Google Scholar] [CrossRef]
- Yang, Y.; Xia, Q.; Sun, Q.; Zhang, Y.; Li, Y.; Ma, X.; Guan, Z.; Zhang, J.; Li, Z.; Liu, K.; et al. Detection of African swine fever virus antibodies in serum using a pB602L protein-based indirect ELISA. Front. Vet. Sci. 2022, 9, 971841. [Google Scholar] [CrossRef]
- Giménez-Lirola, L.G.; Mur, L.; Rivera, B.; Mogler, M.; Sun, Y.; Lizano, S.; Goodell, C.; Harris, D.L.; Rowland, R.R.; Gallardo, C.; et al. Detection of African Swine Fever Virus Antibodies in Serum and Oral Fluid Specimens Using a Recombinant Protein 30 (p30) Dual Matrix Indirect ELISA. PLoS ONE 2016, 11, e0161230. [Google Scholar] [CrossRef]
- Nah, J.-J.; Kwon, O.-K.; Choi, J.-D.; Jang, S.-H.; Lee, H.J.; Ahn, D.-G.; Lee, K.; Kang, B.; Kang, H.-E.; Shin, Y.-K. Development of an indirect ELISA against African swine fever virus using two recombinant antigens, partial p22 and p30. J. Virol. Methods 2022, 309, 114611. [Google Scholar] [CrossRef]
- Wang, T.; Sun, Y.; Huang, S.; Qiu, H.J. Multifaceted Immune Responses to African Swine Fever Virus: Implications for Vaccine Development. Vet. Microbiol. 2020, 249, 108832. [Google Scholar] [CrossRef]
- Ros-Lucas, A.; Correa-Fiz, F.; Bosch-Camós, L.; Rodriguez, F.; Alonso-Padilla, J. Computational Analysis of African Swine Fever Virus Protein Space for the Design of an Epitope-Based Vaccine Ensemble. Pathogens 2020, 9, 1078. [Google Scholar] [CrossRef]
- Teklue, T.; Sun, Y.; Abid, M.; Luo, Y.; Qiu, H.J. Current status and evolving approaches to African swine fever vaccine development. Trans. Emerg. Dis. 2020, 67, 529–542. [Google Scholar] [CrossRef]
- Mallapaty, S. Spread of deadly pig virus in China hastens vaccine research. Nature 2019, 569, 13–14. [Google Scholar] [CrossRef]
- Wang, Y.; Kang, W.; Yang, W.; Zhang, J.; Li, D.; Zheng, H. Structure of African Swine Fever Virus and Associated Molecular Mechanisms Underlying Infection and Immunosuppression: A Review. Front. Immunol. 2021, 12, 715582. [Google Scholar] [CrossRef]
- Gaudreault, N.N.; Richt, J.A. Subunit Vaccine Approaches for African Swine Fever Virus. Vaccines 2019, 7, 56. [Google Scholar] [CrossRef]
- Jiang, W.; Jiang, D.; Li, L.; Wan, B.; Wang, J.; Wang, P.; Shi, X.; Zhao, Q.; Song, J.; Zhu, Z.; et al. Development of an indirect ELISA for the identification of African swine fever virus wild-type strains and CD2v-deleted strains. Front. Vet. Sci. 2022, 9, 1006895. [Google Scholar] [CrossRef]
- Gaudreault, N.N.; Madden, D.W.; Wilson, W.C.; Trujillo, J.D.; Richt, J.A. African Swine Fever Virus: An Emerging DNA Arbovirus. Front. Vet. Sci. 2020, 7, 215. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Pinero, J.; Gallardo, C.; Elizalde, M.; Robles, A.; Gómez, C.; Bishop, R.; Heath, L.; Couacy-Hymann, E.; Fasina, F.O.; Pelayo, V.; et al. Molecular diagnosis of African Swine Fever by a new real-time PCR using universal probe library. Trans. Emerg. Dis. 2013, 60, 48–58. [Google Scholar] [CrossRef]
- Sattler, T.; Wodak, E.; Revilla-Fernández, S.; Schmoll, F. Comparison of different commercial ELISAs for detection of antibodies against porcine respiratory and reproductive syndrome virus in serum. BMC Vet. Res. 2014, 10, 300. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Epitope Name | Protein Name | Sequence of Dominant Linear Epitope | Position | Score |
|---|---|---|---|---|---|
| 1 | pA104R-2 | pA104R | QGHNPATGEPIQIKAK | 70–85 | 0.85 |
| 2 | pB602L-2 | pB602L | LHAQSAYTCADTNVDT | 20–35 | 0.94 |
| 3 | pB602L-5 | pB602L | NVDTCASMCADTNVDT | 32–47 | 0.91 |
| 4 | p12-1 | p12 | KCSKAEECTCNNGSCS | 42–57 | 0.81 |
| 5 | p12-5 | p12 | ALDGSSGGGSNVETLL | 2–17 | 0.60 |
| 6 | p14.5-2 | p14.5 | DTREFTSLVPDEADNK | 80–95 | 0.91 |
| 7 | p14.5-3 | p14.5 | DEESGAKPKKKKHLFP | 99–114 | 0.89 |
| 8 | p14.5-4 | p14.5 | PDEADNKPEDDEESGA | 99–114 | 0.83 |
| 9 | p49-2 | p49 | GNKNGAAPVYNEYTNS | 41–56 | 0.93 |
| 10 | pE248R-1 | pE248R | LTQWMDAGTHDQKTDI | 91–106 | 0.94 |
| 11 | pE248R-3 | pE248R | THDQKTDIEENIKVNL | 99–114 | 0.89 |
| 12 | pE248R-4 | pE248R | EEEEAEPLISNKVLKN | 229–244 | 0.85 |
| 13 | p72-4 | p72 | HYSCNGPQTPKYYQPP | 311–326 | 0.95 |
| 14 | p72-6 | p72 | ITPITDATYLDIRRNV | 295–310 | 0.94 |
| 15 | p72-11 | p72 | TWNISDQNPHQHRDWH | 473–488 | 0.9 |
| 16 | p30-4 | p30 | TLFEQEPSSEVPKDSK | 121–136 | 0.82 |
| 17 | p30-9 | p30 | YTEHQAQEEWNMILHV | 74–89 | 0.77 |
| 18 | p54-2 | p54 | ASQTMSAIENLRQRNT | 159–174 | 0.87 |
| 19 | p54-4 | p54 | TASVGKPVTGRPATNR | 95–110 | 0.85 |
| 20 | p54-5 | p54 | YGECLSPVTTPSFFST | 14–29 | 0.83 |
| 21 | p54-6 | p54 | EVTPQPGTSKPAGATT | 80–95 | 0.82 |
| 22 | p54-7 | p54 | KPVTDNPVTDRLVMAT | 115–130 | 0.81 |
| 23 | p54-9 | p54 | NRPATNKPVTDNPVTD | 109–124 | 0.8 |
| 24 | pp62-2 | pp62 | AMYGDKHTTGDIAPND | 422–437 | 0.89 |
| 25 | pp62-4 | pp62 | EGGGNDPPVPKHISPY | 156–171 | 0.88 |
| 26 | pp220-6 | pp220 | PTTEAAAAAQDRNLTT | 54–69 | 0.84 |
| 27 | pp220-10 | pp220 | LQLRGGAQRRDAANIQ | 233–248 | 0.81 |
| INGENASA-Indirect ELISA | Total | |||
|---|---|---|---|---|
| Positive | Negative | |||
| k3-iELISA | positive | 145 | 4 | 149 |
| negative | 0 | 230 | 230 | |
| Total | 145 | 234 | 379 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Afayibo, D.J.A.; Zhang, Z.; Sun, H.; Fu, J.; Zhao, Y.; Amuda, T.O.; Wu, M.; Du, J.; Guan, G.; Niu, Q.; et al. Establishment of an ELISA Based on a Recombinant Antigenic Protein Containing Multiple Prominent Epitopes for Detection of African Swine Fever Virus Antibodies. Microorganisms 2024, 12, 943. https://doi.org/10.3390/microorganisms12050943
Afayibo DJA, Zhang Z, Sun H, Fu J, Zhao Y, Amuda TO, Wu M, Du J, Guan G, Niu Q, et al. Establishment of an ELISA Based on a Recombinant Antigenic Protein Containing Multiple Prominent Epitopes for Detection of African Swine Fever Virus Antibodies. Microorganisms. 2024; 12(5):943. https://doi.org/10.3390/microorganisms12050943
Chicago/Turabian StyleAfayibo, Dossêh Jean Apôtre, Zhonghui Zhang, Hualin Sun, Jingsheng Fu, Yaru Zhao, Tharheer Oluwashola Amuda, Mengli Wu, Junzheng Du, Guiquan Guan, Qingli Niu, and et al. 2024. "Establishment of an ELISA Based on a Recombinant Antigenic Protein Containing Multiple Prominent Epitopes for Detection of African Swine Fever Virus Antibodies" Microorganisms 12, no. 5: 943. https://doi.org/10.3390/microorganisms12050943
APA StyleAfayibo, D. J. A., Zhang, Z., Sun, H., Fu, J., Zhao, Y., Amuda, T. O., Wu, M., Du, J., Guan, G., Niu, Q., Yang, J., & Yin, H. (2024). Establishment of an ELISA Based on a Recombinant Antigenic Protein Containing Multiple Prominent Epitopes for Detection of African Swine Fever Virus Antibodies. Microorganisms, 12(5), 943. https://doi.org/10.3390/microorganisms12050943