
Characterizing the Role of AosfgA and AofluG in Mycelial and Conidial Development in Arthrobotrys oligospora and Their Role in Secondary Metabolism



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains, Plasmids, and Growth Conditions
2.2. Fluorescent Quantitative PCR (RT-qPCR)
2.3. Y2H Assay
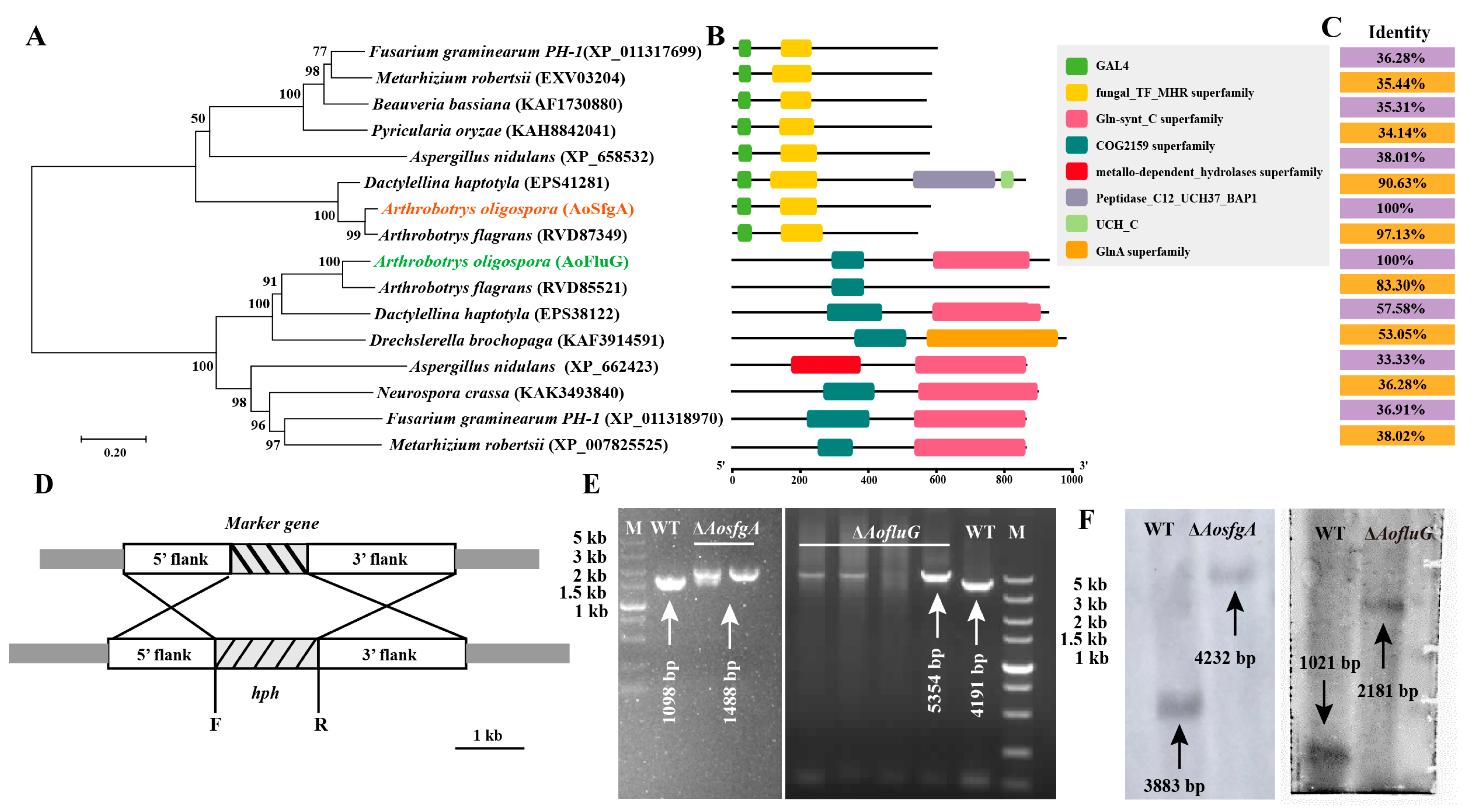
2.4. Sequence and Phylogenetic Analysis of AoSfgA and AoFluG
2.5. Targeted Gene Deletion
2.6. Colony Growth and Stress Adaption Analysis
2.7. Staining Analysis
2.8. Analysis of Conidial Production and Morphology
2.9. Observation of Trap Morphology and Determination of Pathogenicity
2.10. Liquid Chromatography–Mass Spectrometry (LC-MS) Analysis
2.11. Statistical Analysis
3. Results
3.1. AoSfgA Interacts with AoFluG
3.2. Sequence Analysis of AoSfgA and AoFluG Protein
3.3. Deletion and Validation of the AosfgA and AofluG Genes
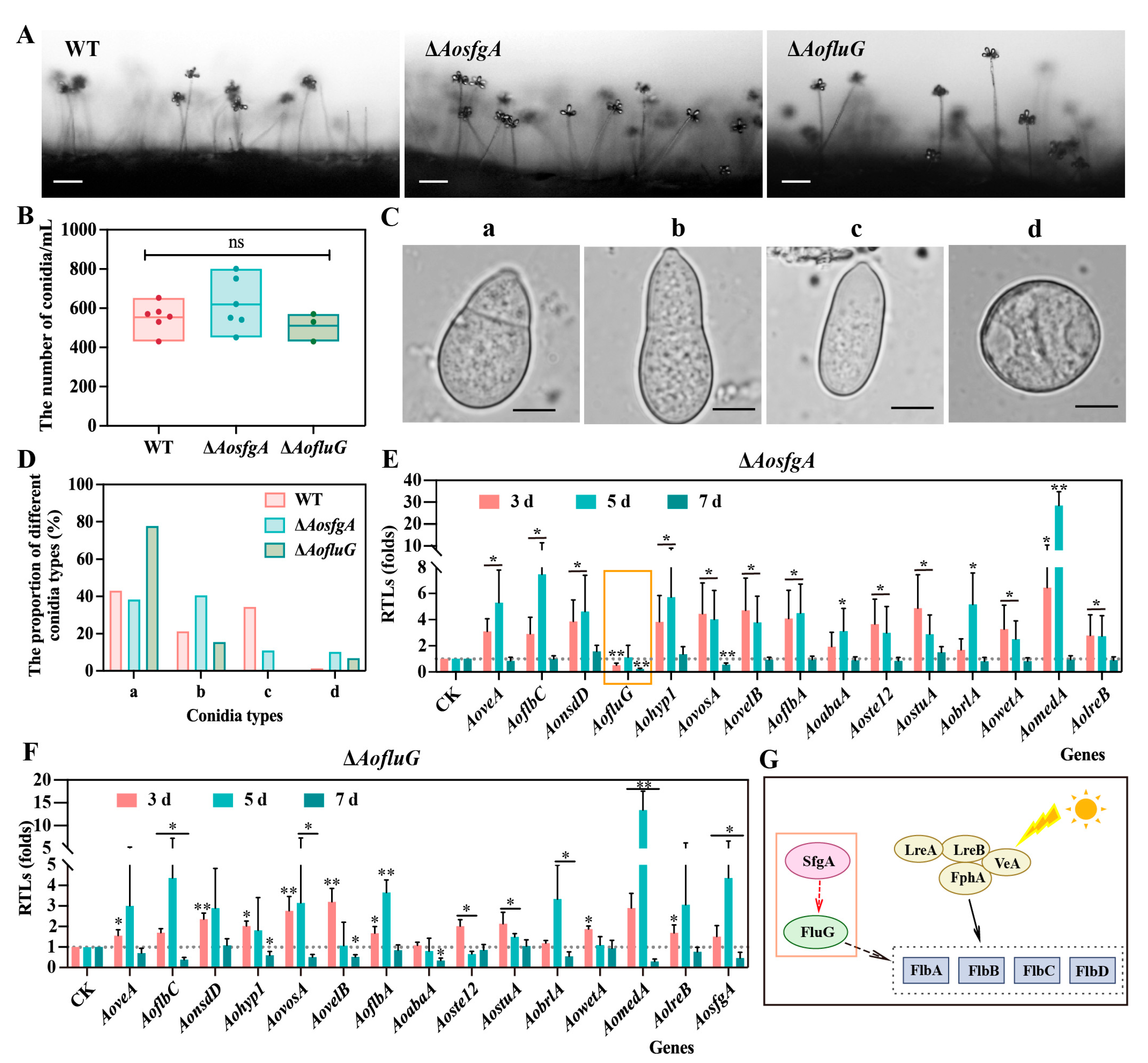
3.4. AosfgA and AofluG Genes Regulate Mycelial and Nuclei Development
3.5. AosfgA and AofluG Genes Impair Conidial Growth
3.6. AosfgA Positively Regulates AofluG in A. oligospora
3.7. AosfgA and AofluG Genes Do Not Affect Trap Formation but Regulate Vacuole Assembly
3.8. AosfgA Responds to Heat and Chemical Stresses
3.9. AosfgA and AofluG Genes Contribute to Secondary Metabolite Synthesis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flores Francisco, B.G.; Ponce, I.M.; Plascencia Espinosa, M.Á.; Mendieta Moctezuma, A.; López Y López, V.E. Advances in the biological control of phytoparasitic nematodes via the use of nematophagous fungi. World J. Microbiol. Biotechnol. 2021, 37, 180. [Google Scholar] [CrossRef]
- Lahm, G.P.; Desaeger, J.; Smith, B.K.; Pahutski, T.F.; Rivera, M.A.; Meloro, T.; Kucharczyk, R.; Lett, R.M.; Daly, A.; Smith, B.T.; et al. The discovery of fluazaindolizine: A new product for the control of plant parasitic nematodes. Bioorg. Med. Chem. Lett. 2017, 27, 1572–1575. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Xiang, M.; Liu, X. Nematode-trapping fungi. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Jiang, K.; Duan, S.; Zhao, N.; Shen, Y.; Zhu, L.; Zhang, K.Q.; Yang, J. Identification of a transcription factor AoMsn2 of the Hog1 signaling pathway contributes to fungal growth, development and pathogenicity in Arthrobotrys oligospora. J. Adv. Res. 2024. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.; Xu, L.; Yang, Y.; Zhang, X.; Xiang, M.; Wang, C.; An, Z.; Liu, X. Origin and evolution of carnivorism in the Ascomycota (fungi). Proc. Natl. Acad. Sci. USA 2012, 109, 10960–10965. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Li, X.; Li, G.; Pan, Y.; Zhang, K. Acanthocytes of Stropharia rugosoannulata function as a nematode-attacking device. Appl. Environ. Microbiol. 2006, 72, 2982–2987. [Google Scholar] [CrossRef]
- Luo, H.; Liu, Y.; Fang, L.; Li, X.; Tang, N.; Zhang, K. Coprinus comatus damages nematode cuticles mechanically with spiny balls and produces potent toxins to immobilize nematodes. Appl. Environ. Microbiol. 2007, 73, 3916–3923. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liang, L.; Li, J.; Zhang, K.Q. Nematicidal enzymes from microorganisms and their applications. Appl. Microbiol. Biotechnol. 2013, 97, 7081–7095. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.M.; Zou, C.G.; Xu, J.; Zhang, K.Q. Signal pathways involved in microbe-nematode interactions provide new insights into the biocontrol of plant-parasitic nematodes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180317. [Google Scholar] [CrossRef]
- Yang, J.; Wang, L.; Ji, X.; Feng, Y.; Li, X.; Zou, C.; Xu, J.; Ren, Y.; Mi, Q.; Wu, J.; et al. Genomic and proteomic analyses of the fungus Arthrobotrys oligospora provide insights into nematode-trap formation. PLoS Pathog. 2011, 7, e1002179. [Google Scholar] [CrossRef]
- Hsueh, Y.P.; Gronquist, M.R.; Schwarz, E.M.; Nath, R.D.; Lee, C.H.; Gharib, S.; Schroeder, F.C.; Sternberg, P.W. Nematophagous fungus Arthrobotrys oligospora mimics olfactory cues of sex and food to lure its nematode prey. eLife 2017, 6, e20023. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Kumar, N.; Singh, K.P. Morphological variations in conidia of Arthrobotrys oligospora on different media. Mycobiology 2005, 33, 118–120. [Google Scholar] [CrossRef] [PubMed]
- Dackman, C.; Nordbring-Hertz, B. Conidial traps—A new survival structure of the nematode-trapping fungus Arthrobotrys oligospora. Mycol. Res. 1992, 96, 194–198. [Google Scholar] [CrossRef]
- Persmark, L.; Nordbring-Hertz, B. Conidial trap formation of nematode-trapping fungi in soil and soil extracts. FEMS Microbiol. Ecol. 1997, 22, 313–323. [Google Scholar] [CrossRef]
- Diallo, M.; Kengen, S.W.M.; Lopez-Contreras, A.M. Sporulation in solventogenic and acetogenic clostridia. Appl. Microbiol. Biotechnol. 2021, 105, 3533–3557. [Google Scholar] [CrossRef] [PubMed]
- Jung, B.; Kim, S.; Lee, J. Microcyle conidiation in filamentous fungi. Mycobiology 2014, 42, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Boylan, M.T.; Mirabito, P.M.; Willett, C.E.; Zimmerman, C.R.; Timberlake, W.E. Isolation and physical characterization of three essential conidiation genes from Aspergillus nidulans. Mol. Cell Biol. 1987, 7, 3113–3118. [Google Scholar]
- Clutterbuck, A.J. A mutational analysis of conidial development in Aspergillus nidulans. Genetics 1969, 63, 317–327. [Google Scholar] [CrossRef]
- Adams, T.H.; Boylan, M.T.; Timberlake, W.E. brlA is necessary and sufficient to direct conidiophore development in Aspergillus nidulans. Cell 1988, 54, 353–362. [Google Scholar] [CrossRef]
- Lee, B.N.; Adams, T.H. The Aspergillus nidulans fluG gene is required for production of an extracellular developmental signal and is related to prokaryotic glutamine synthetase I. Genes Dev. 1994, 8, 641–651. [Google Scholar] [CrossRef]
- Wieser, J.; Lee, B.N.; Fondon, J., 3rd; Adams, T.H. Genetic requirements for initiating asexual development in Aspergillus nidulans. Curr. Genet. 1994, 27, 62–69. [Google Scholar] [CrossRef]
- Seo, J.A.; Guan, Y.; Yu, J.H. FluG-dependent asexual development in Aspergillus nidulans occurs via derepression. Genetics 2006, 172, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Yu, J.H. Genetic control of asexual sporulation in filamentous fungi. Curr. Opin. Microbiol. 2012, 15, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Krijgsheld, P.; Nitsche, B.M.; Post, H.; Levin, A.M.; Muller, W.H.; Heck, A.J.; Ram, A.F.; Altelaar, A.F.; Wosten, H.A. Deletion of flbA results in increased secretome complexity and reduced secretion heterogeneity in colonies of Aspergillus niger. J. Proteome Res. 2013, 12, 1808–1819. [Google Scholar] [CrossRef] [PubMed]
- Krijgsheld, P.; Bleichrodt, R.; van Veluw, G.J.; Wang, F.; Muller, W.H.; Dijksterhuis, J.; Wosten, H.A. Development in Aspergillus. Stud. Mycol. 2013, 74, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Ruger-Herreros, C.; Rodriguez-Romero, J.; Fernandez-Barranco, R.; Olmedo, M.; Fischer, R.; Corrochano, L.M.; Canovas, D. Regulation of conidiation by light in Aspergillus nidulans. Genetics 2011, 188, 809–822. [Google Scholar] [CrossRef]
- D’Souza, C.A.; Lee, B.N.; Adams, T.H. Characterization of the role of the FluG protein in asexual development of Aspergillus nidulans. Genetics 2001, 158, 1027–1036. [Google Scholar] [CrossRef]
- Yang, X.; Ma, N.; Yang, L.; Zheng, Y.; Zhen, Z.; Li, Q.; Xie, M.; Li, J.; Zhang, K.Q.; Yang, J. Two Rab GTPases play different roles in conidiation, trap formation, stress resistance, and virulence in the nematode-trapping fungus Arthrobotrys oligospora. Appl. Microbiol. Biotechnol. 2018, 102, 4601–4613. [Google Scholar] [CrossRef]
- Zhu, Y.; Yang, X.; Bai, N.; Liu, Q.; Yang, J. AoRab7A interacts with AoVps35 and AoVps41 to regulate vacuole assembly, trap formation, conidiation, and functions of proteasomes and ribosomes in Arthrobotrys oligospora. Microbiol. Res. 2024, 280, 127573. [Google Scholar] [CrossRef]
- Xie, M.; Bai, N.; Yang, X.; Liu, Y.; Zhang, K.Q.; Yang, J. Fus3 regulates asexual development and trap morphogenesis in the nematode-trapping fungus Arthrobotrys oligospora. iScience 2023, 26, 107404. [Google Scholar] [CrossRef]
- Bai, N.; Xie, M.; Liu, Q.; Zhu, Y.; Yang, X.; Zhang, K.Q.; Yang, J. AoMedA has a complex regulatory relationship with AoBrlA, AoAbaA, and AoWetA in conidiation, trap formation, and secondary metabolism in the nematode-trapping fungus Arthrobotrys oligospora. Appl. Environ. Microbiol. 2023, 89, e0098323. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, D.; Bai, N.; Zhu, Y.; Yang, J. Peroxin Pex14/17 is required for trap Formation, and plays pleiotropic roles in mycelial development, stress response, and secondary metabolism in Arthrobotrys oligospora. mSphere 2023, 8, e0001223. [Google Scholar] [CrossRef] [PubMed]
- Cui, P.; Tian, M.; Huang, J.; Zheng, X.; Guo, Y.; Li, G.; Wang, X. Amphiphysin AoRvs167-mediated membrane curvature facilitates trap formation, endocytosis, and stress resistance in Arthrobotrys oligospora. Pathogens 2022, 11, 997. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zientara-Rytter, K.; Ozeki, K.; Nazarko, T.Y.; Subramani, S. Pex3 and Atg37 compete to regulate the interaction between the pexophagy receptor, Atg30, and the Hrr25 kinase. Autophagy 2018, 14, 368–384. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.A.; Lin, H.C.; Hsueh, Y.P. The cAMP-PKA pathway regulates prey sensing and trap morphogenesis in the nematode-trapping fungus Arthrobotrys oligospora. G3 2022, 12, jkac217. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Xu, J.; Li, J.; Lin, L.; Liu, Q.; Sun, W.; Huang, B.; Tian, C. Development of genetic tools for Myceliophthora thermophila. BMC Biotechnol. 2015, 15, 35. [Google Scholar] [CrossRef]
- Zhao, X.; Fan, Y.; Xiang, M.; Kang, S.; Wang, S.; Liu, X. DdaCrz1, a C(2)H(2)-type transcription factor, regulates growth, conidiation, and stress resistance in the nematode-trapping fungus Drechslerella dactyloides. J. Fungi 2022, 8, 750. [Google Scholar] [CrossRef]
- Xiao, J.; Zhang, Y.; Yang, K.; Tang, Y.; Wei, L.; Liu, E.; Liang, Z. Protein kinase Ime2 is associated with mycelial growth, conidiation, osmoregulation, and pathogenicity in Fusarium oxysporum. Arch. Microbiol. 2022, 204, 455. [Google Scholar] [CrossRef]
- Liu, X.; Miao, Q.; Zhou, Z.; Lu, S.; Li, J. Identification of three novel conidiogenesis-related genes in the nematode-trapping fungus Arthrobotrys oligospora. Pathogens 2022, 11, 717. [Google Scholar] [CrossRef]
- Zhou, L.; Li, M.; Cui, P.; Tian, M.; Xu, Y.; Zheng, X.; Zhang, K.; Li, G.; Wang, X. Arrestin-coding genes regulate endocytosis, sporulation, pathogenicity, and stress resistance in Arthrobotrys oligospora. Front. Cell. Infect. Microbiol. 2022, 12, 754333. [Google Scholar] [CrossRef]
- Gu, T.; Lu, H.; Liu, H.; Zhang, G.; Wang, Y. Function discovery of a non-ribosomal peptide synthetase-like encoding gene in the nematode-trapping fungus Arthrobotrys oligospora. Front. Microbiol. 2023, 14, 1210288. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Ran, Y.; Zhang, K.Q.; Li, G.H. AfLaeA, a global regulator of mycelial growth, chlamydospore production, pathogenicity, secondary metabolism, and energy metabolism in the nematode-trapping fungus Arthrobotrys flagrans. Microbiol. Spectr. 2023, 11, e0018623. [Google Scholar] [CrossRef]
- Yang, X.; Chen, Y.; Zhang, L.; He, J.; Wu, Q.; Li, S.; Wang, D.; Gou, J.; Wu, Z.; Zhang, K.; et al. Melanin precursors mediated adaption to temperature changes in fungus and animal via inhibition of lipid-mediated ferroptosis. Sci. China Life Sci. 2023, 66, 1800–1817. [Google Scholar] [CrossRef]
- He, Z.Q.; Tan, J.L.; Li, N.; Zhang, H.X.; Chen, Y.H.; Wang, L.J.; Zhang, K.Q.; Niu, X.M. Sesquiterpenyl epoxy-cyclohexenoids and their signaling functions in nematode-trapping fungus Arthrobotrys oligospora. J. Agric. Food Chem. 2019, 67, 13061–13072. [Google Scholar] [CrossRef]
- Todd, R.B.; Andrianopoulos, A. Evolution of a fungal regulatory gene family: The Zn(II)2Cys6 binuclear cluster DNA binding motif. Fungal Genet. Biol. 1997, 21, 388–405. [Google Scholar] [CrossRef] [PubMed]
- Ojeda-Lopez, M.; Chen, W.; Eagle, C.E.; Gutierrez, G.; Jia, W.L.; Swilaiman, S.S.; Huang, Z.; Park, H.S.; Yu, J.H.; Canovas, D.; et al. Evolution of asexual and sexual reproduction in the aspergilli. Stud. Mycol. 2018, 91, 37–59. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Kwon, N.J.; Lee, I.S.; Jung, S.; Kim, S.C.; Yu, J.H. Negative regulation and developmental competence in Aspergillus. Sci. Rep. 2016, 6, 28874. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Urra, A.B.; Jimenez, C.; Nieto, M.I.; Rodriguez, J.; Hayashi, H.; Ugalde, U. Signaling the induction of sporulation involves the interaction of two secondary metabolites in Aspergillus nidulans. ACS Chem. Biol. 2012, 7, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.Y.; Li, J.Y.; Zhi, Q.Q.; Chi, S.D.; Qu, S.; Luo, Y.F.; He, Z.M. SfgA renders Aspergillus flavus more stable to the external environment. J. Fungi 2022, 8, 638. [Google Scholar] [CrossRef] [PubMed]
- Adams, T.H.; Hide, W.A.; Yager, L.N.; Lee, B.N. Isolation of a gene required for programmed initiation of development by Aspergillus nidulans. Mol. Cell Biol. 1992, 12, 3827–3833. [Google Scholar] [PubMed]
- Chavrier, P.; Parton, R.G.; Hauri, H.P.; Simons, K.; Zerial, M. Localization of low molecular weight GTP binding proteins to exocytic and endocytic compartments. Cell 1990, 62, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J.; Herman, P.K.; Emr, S.D. The fungal vacuole: Composition, function, and biogenesis. Microbiol. Rev. 1990, 54, 266–292. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.M.; Liu, X.G.; Shi, H.B.; Lu, J.P.; Yang, J.; Lin, F.C.; Liu, X.H. MoMon1 is required for vacuolar assembly, conidiogenesis and pathogenicity in the rice blast fungus Magnaporthe oryzae. Res. Microbiol. 2013, 164, 300–309. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, J.; Fan, Y.; Xiang, M.; Kang, S.; Wei, D.; Liu, X. SNARE protein DdVam7 of the nematode-trapping fungus Drechslerella dactyloides regulates vegetative growth, conidiation, and the predatory process via vacuole assembly. Microbiol. Spectr. 2022, 10, e0187222. [Google Scholar] [CrossRef]
- Yu, X.; Hu, X.; Pop, M.; Wernet, N.; Kirschhöfer, F.; Brenner-Weiß, G.; Keller, J.; Bunzel, M.; Fischer, R. Fatal attraction of Caenorhabditis elegans to predatory fungi through 6-methyl-salicylic acid. Nat. Commun. 2021, 12, 5462. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Q.; Bai, N.; Duan, S.; Shen, Y.; Zhu, L.; Yang, J. Characterizing the Role of AosfgA and AofluG in Mycelial and Conidial Development in Arthrobotrys oligospora and Their Role in Secondary Metabolism. Microorganisms 2024, 12, 615. https://doi.org/10.3390/microorganisms12030615
Liu Q, Bai N, Duan S, Shen Y, Zhu L, Yang J. Characterizing the Role of AosfgA and AofluG in Mycelial and Conidial Development in Arthrobotrys oligospora and Their Role in Secondary Metabolism. Microorganisms. 2024; 12(3):615. https://doi.org/10.3390/microorganisms12030615
Chicago/Turabian StyleLiu, Qianqian, Na Bai, Shipeng Duan, Yanmei Shen, Lirong Zhu, and Jinkui Yang. 2024. "Characterizing the Role of AosfgA and AofluG in Mycelial and Conidial Development in Arthrobotrys oligospora and Their Role in Secondary Metabolism" Microorganisms 12, no. 3: 615. https://doi.org/10.3390/microorganisms12030615
APA StyleLiu, Q., Bai, N., Duan, S., Shen, Y., Zhu, L., & Yang, J. (2024). Characterizing the Role of AosfgA and AofluG in Mycelial and Conidial Development in Arthrobotrys oligospora and Their Role in Secondary Metabolism. Microorganisms, 12(3), 615. https://doi.org/10.3390/microorganisms12030615