

Exploring the Impact of Coconut Peat and Vermiculite on the Rhizosphere Microbiome of Pre-Basic Seed Potatoes under Soilless Cultivation Conditions
 ,
,
Abstract
1. Introduction
2. Materials and Methods
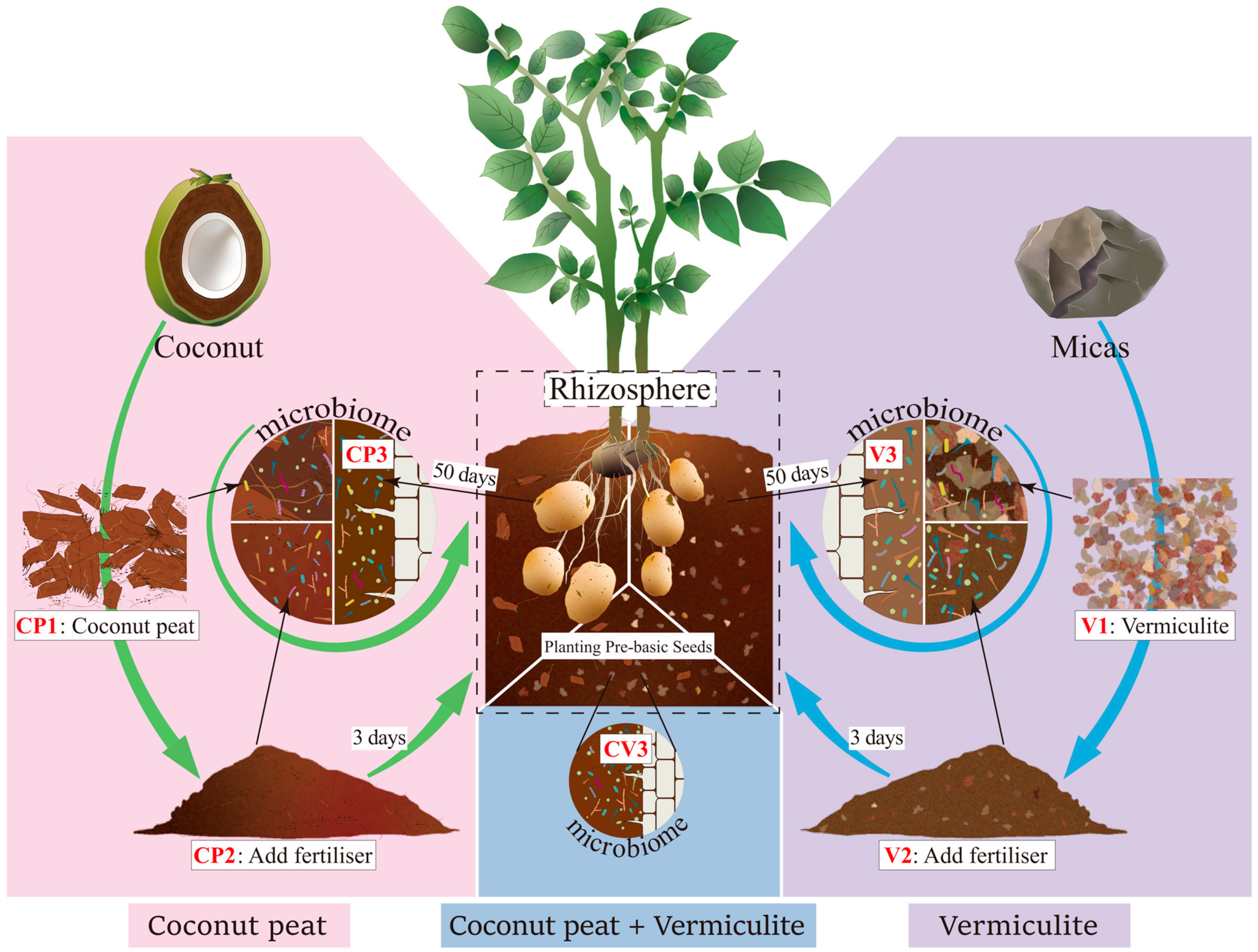
2.1. Soil Sample Collection
2.2. Determination of Substrate Physicochemical Properties
2.3. Sample DNA Extraction, Amplification, and Sequencing
2.4. Bioinformatics Analysis
2.5. Statistical Analysis
3. Results
3.1. Changes in Microbial Community Structure
3.1.1. Changes in Bacterial Community Diversity and Richness
3.1.2. Changes in Fungal Community Diversity and Richness
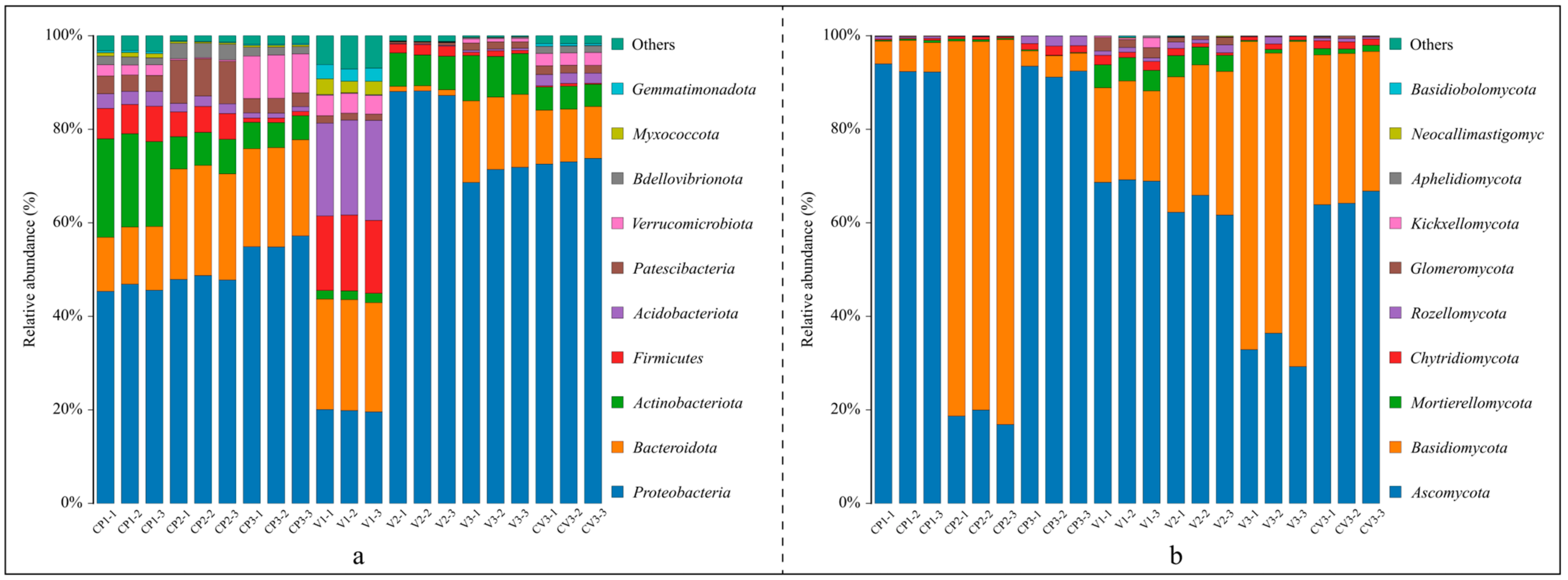
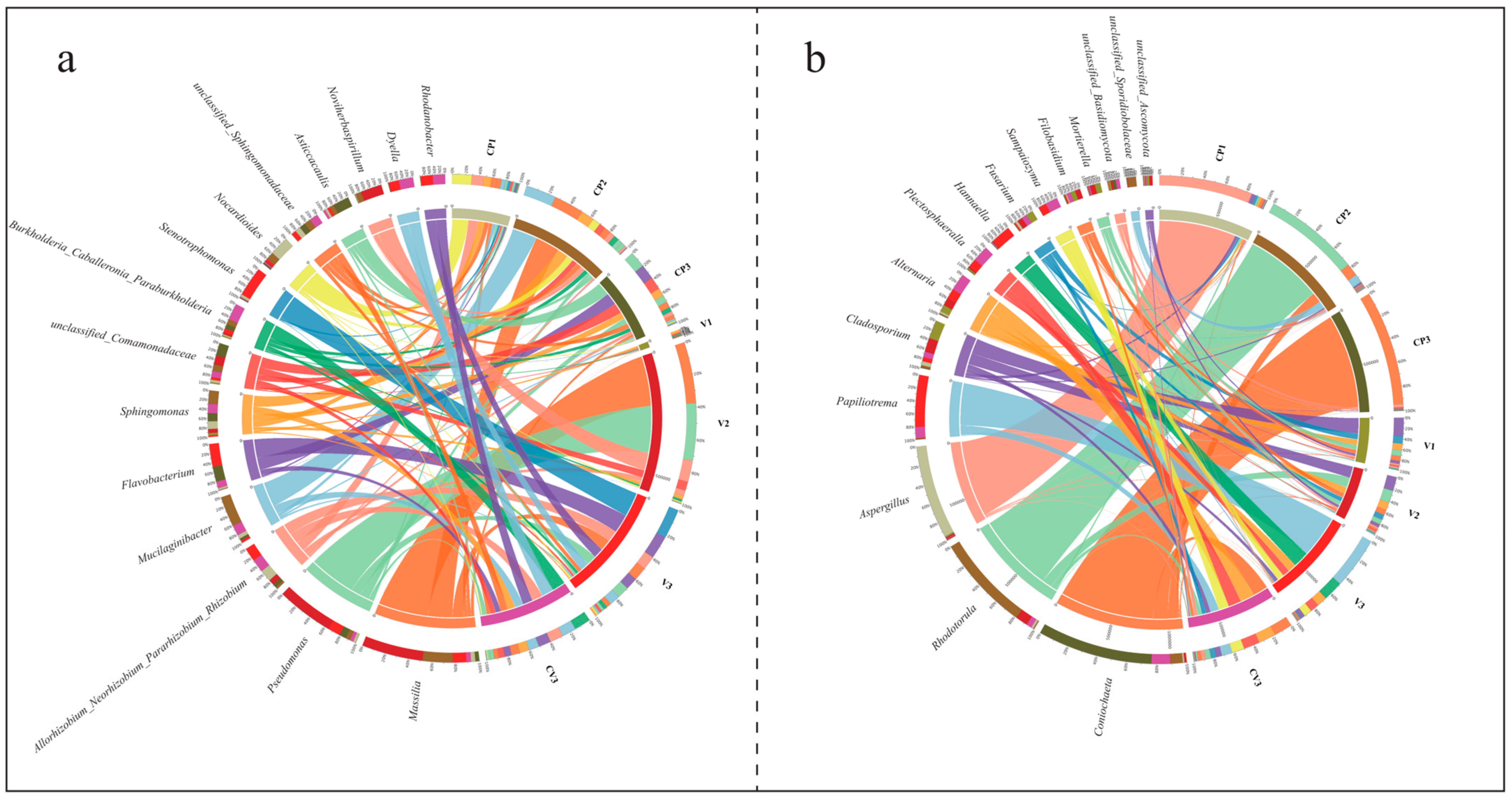
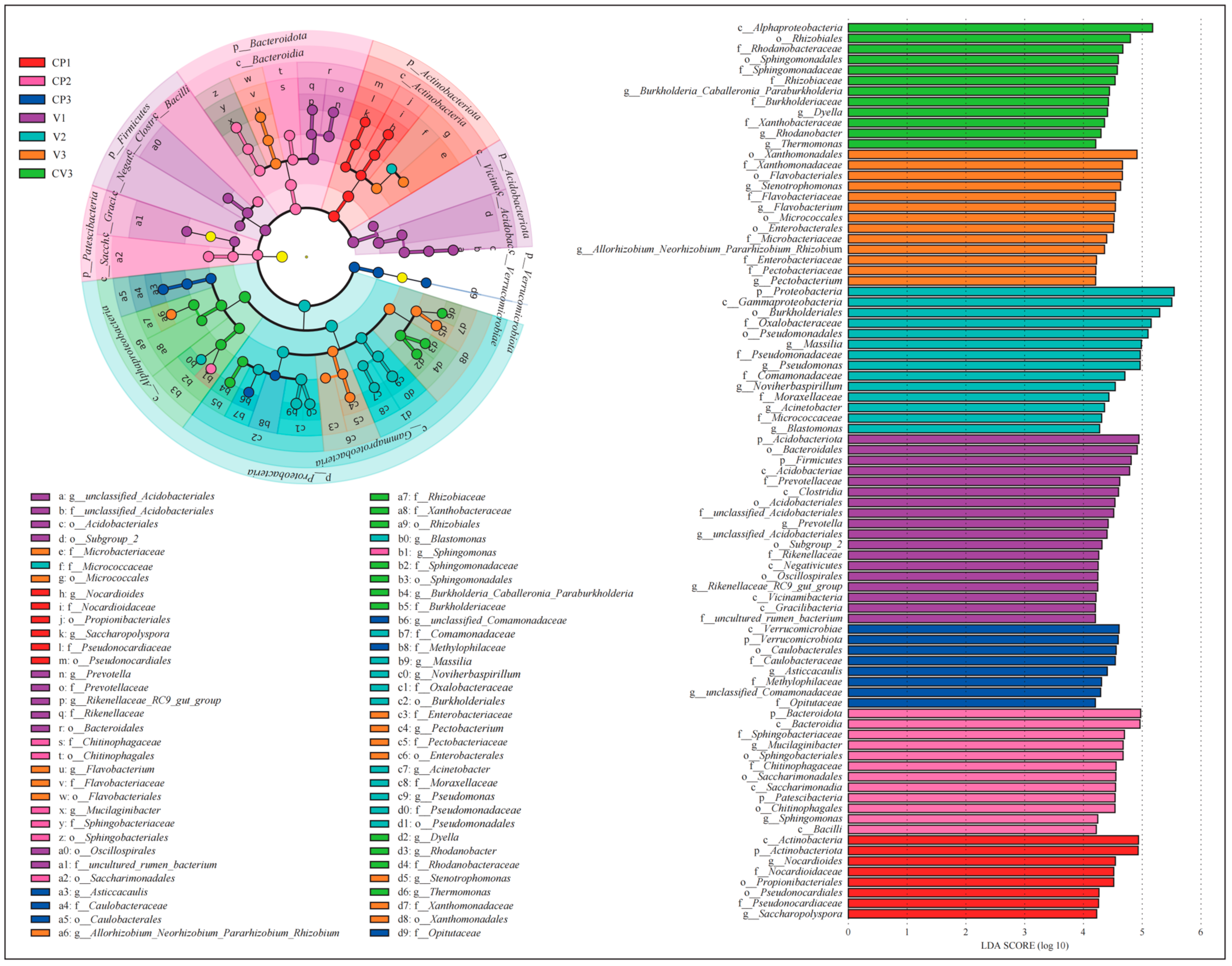
3.1.3. Changes in the Structure of Bacterial and Fungal Communities
- ASV-level analyses showed that the community compositions under different treatments were significantly different.
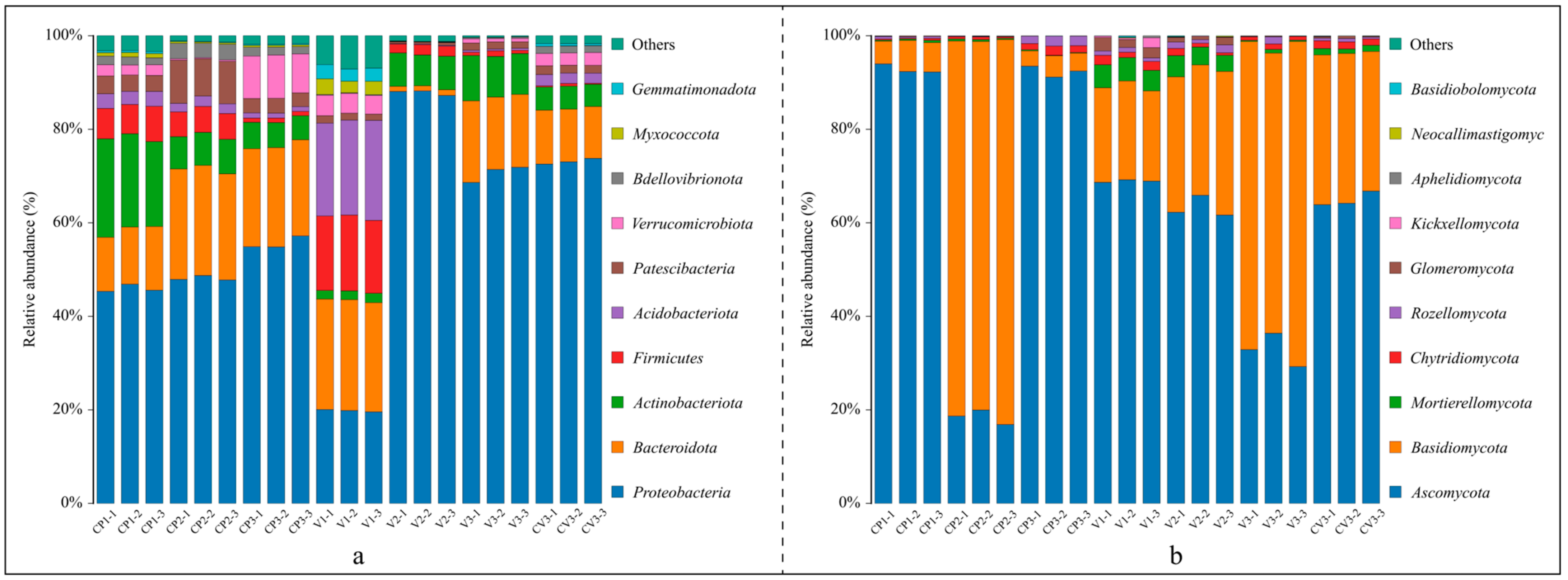
- 2.
- Trends in community changes at the phylum level.
- 3.
- Community changes at the genus level.
- 4.
- LEfSe analysis identified taxa contributing significantly to the community’s uniqueness in different groups.
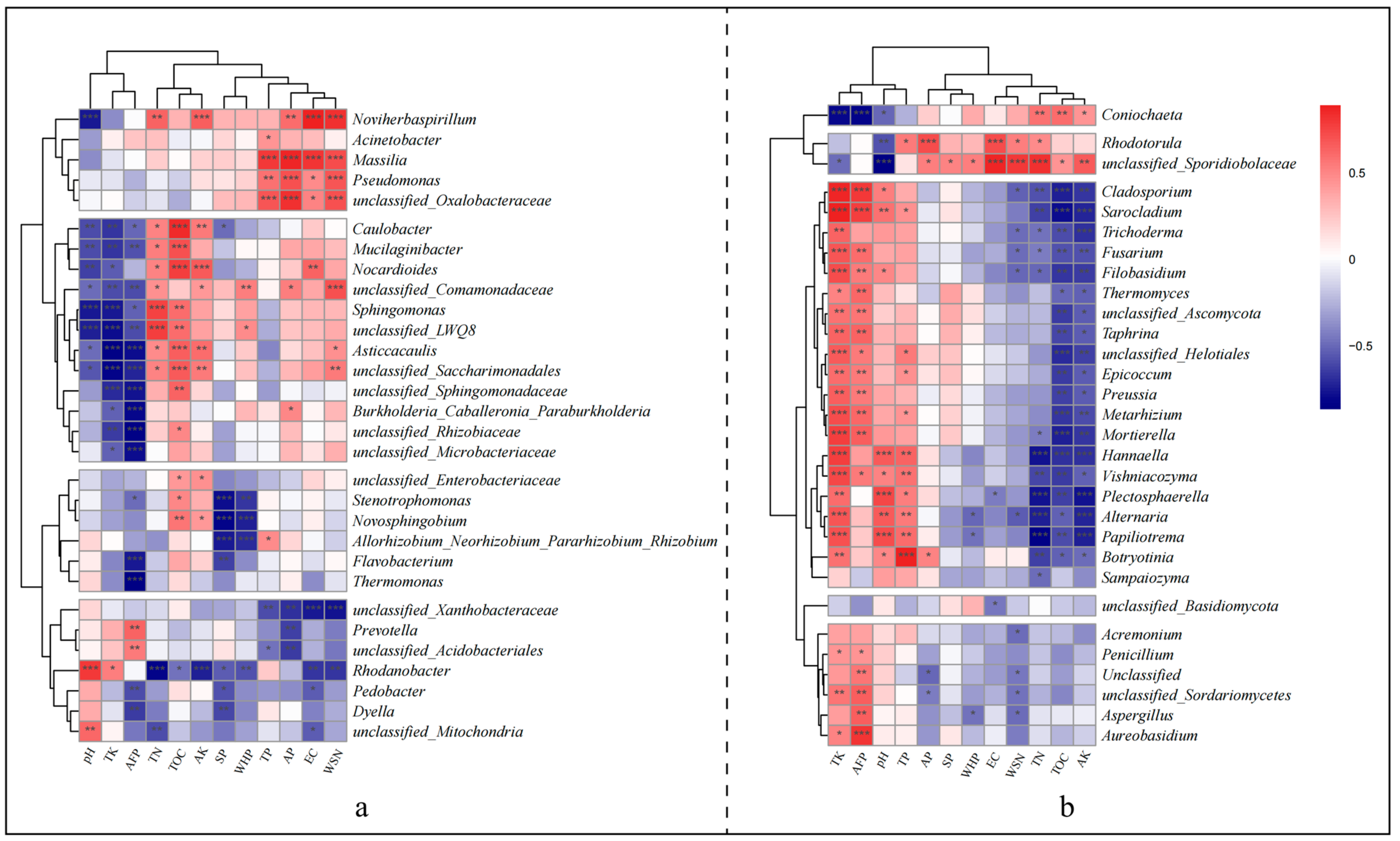
3.2. Enrichment of Plant Growth-Promoting Rhizobacteria within Groups
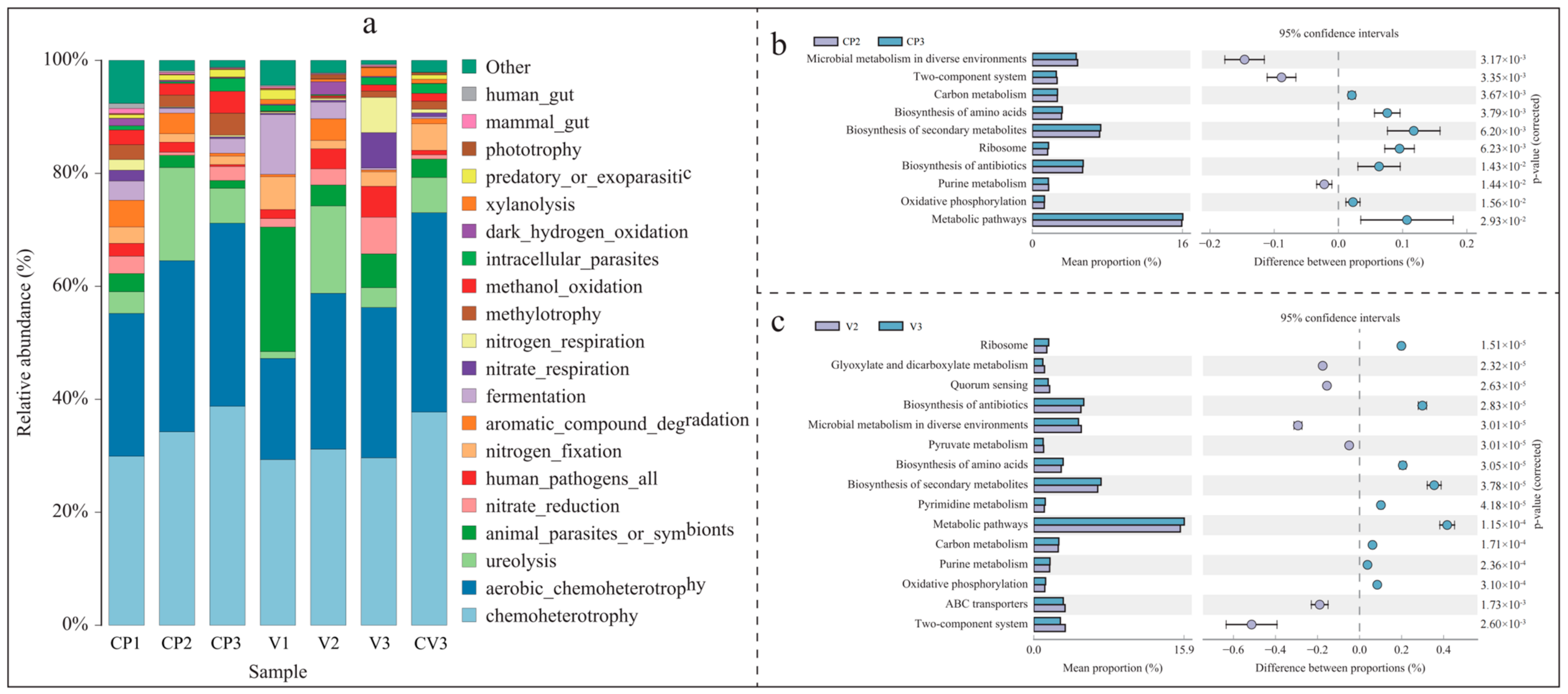
3.3. Changes in Community Function and Metabolic Pathways
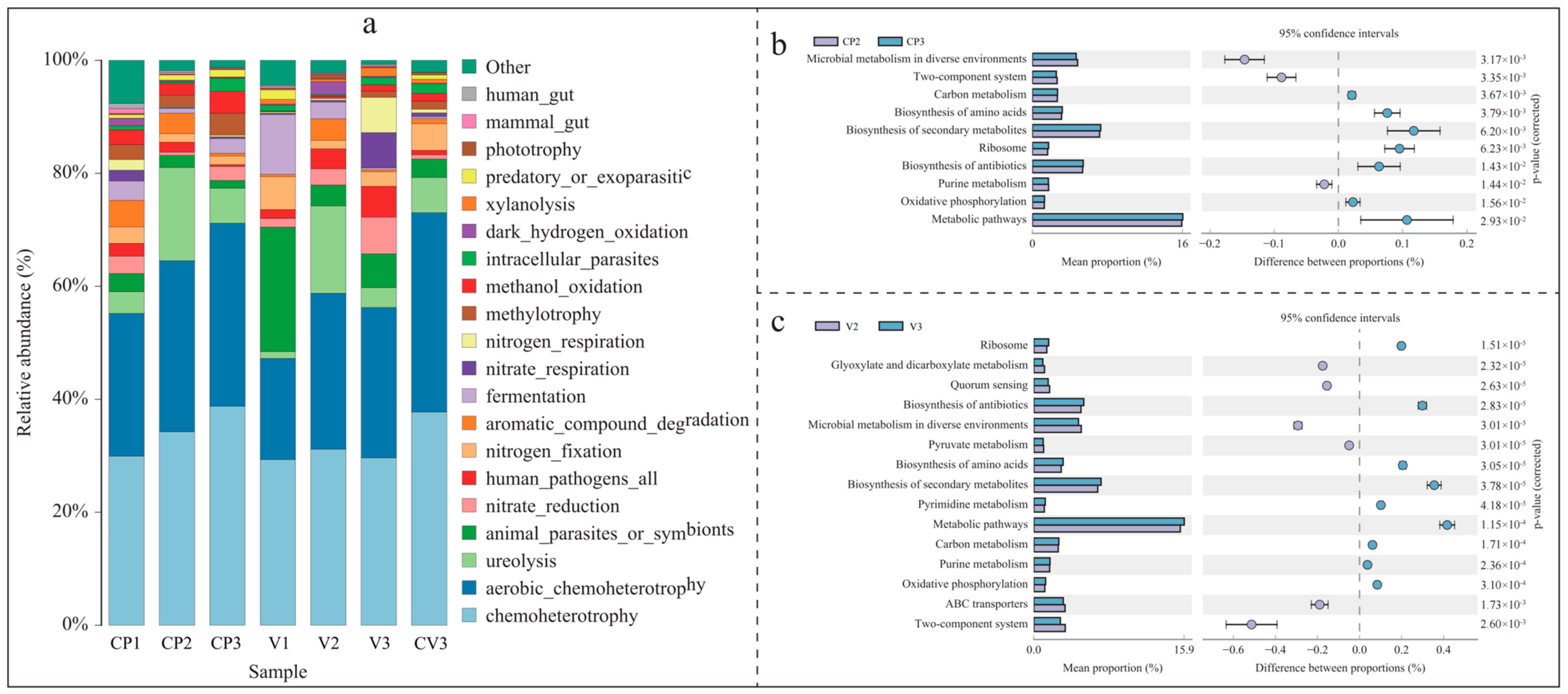
3.3.1. Bacterial FAPROTAX Function Prediction
3.3.2. Bacterial KEGG Metabolic Pathway Changes
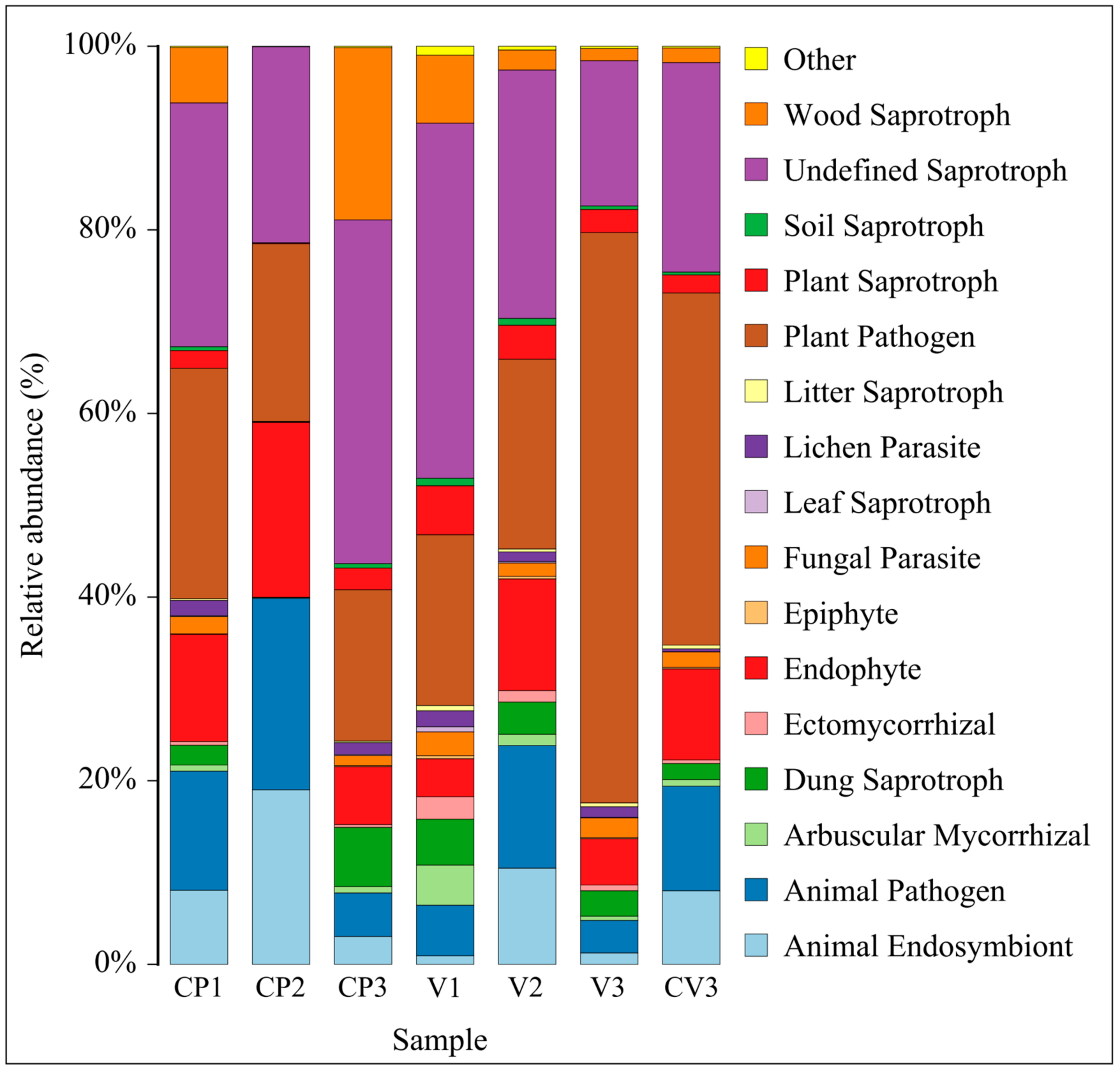
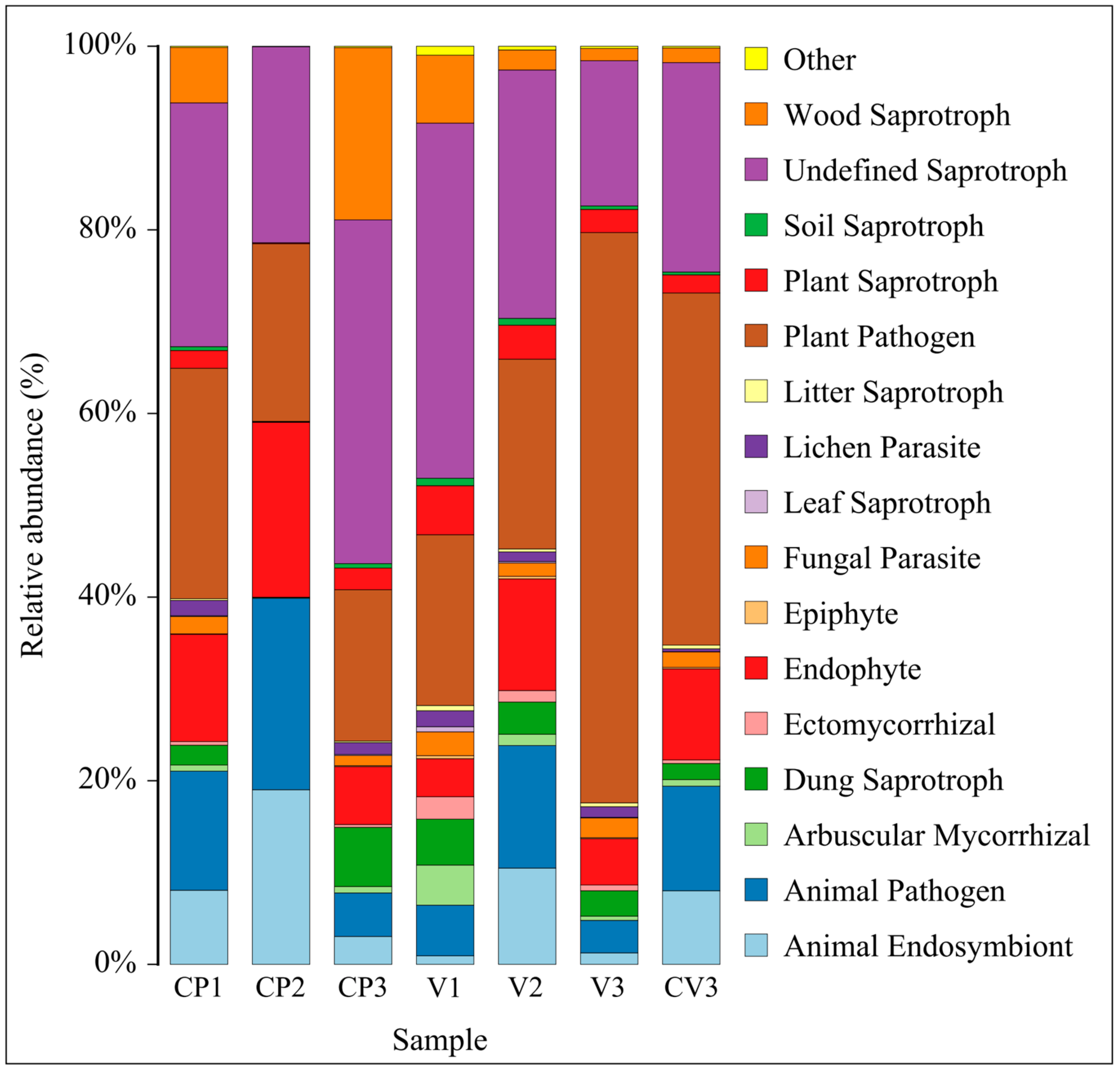
3.3.3. Changes in Fungal FunGuild Nutritional Composition
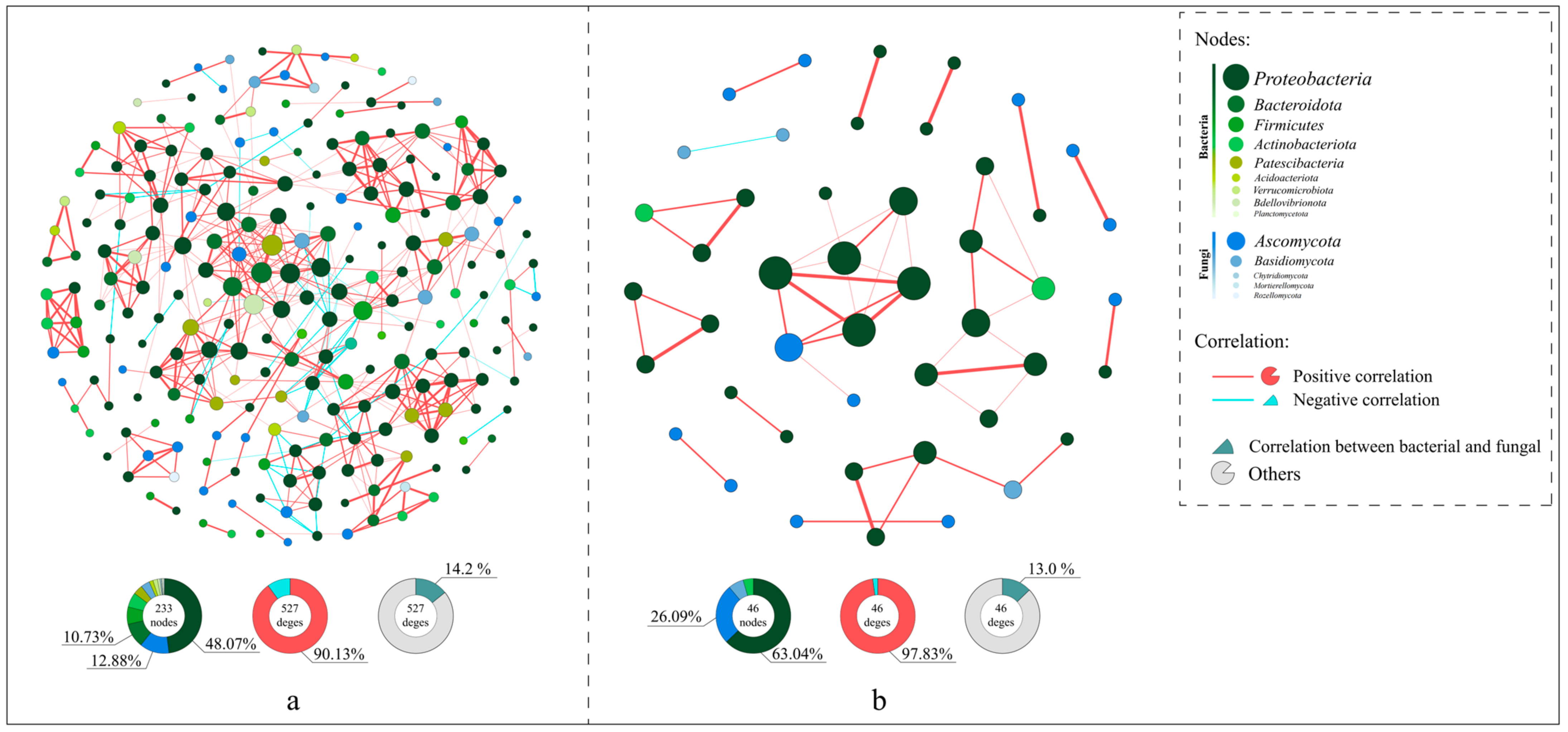
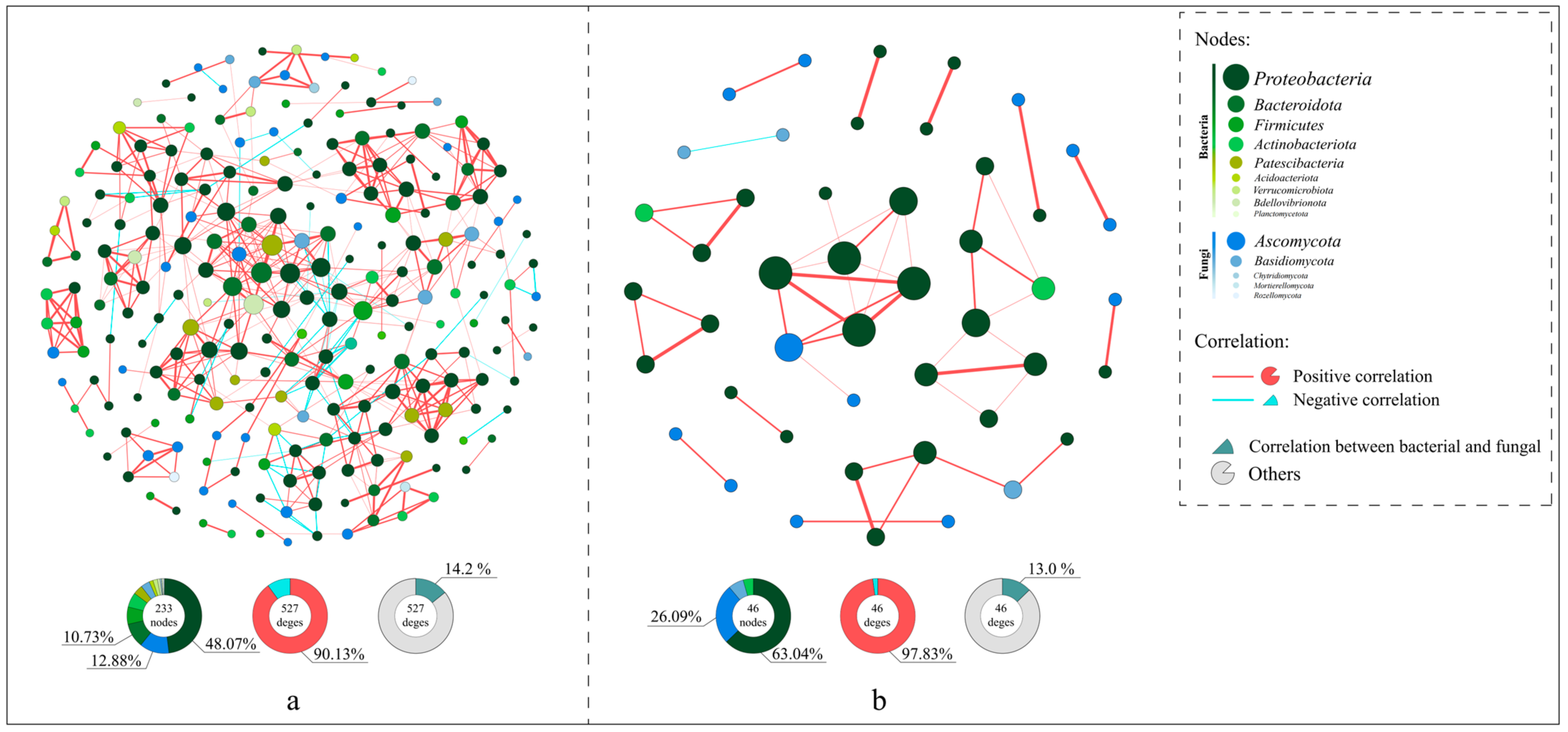
3.4. Differences in Microbial Co-Occurrence Networks
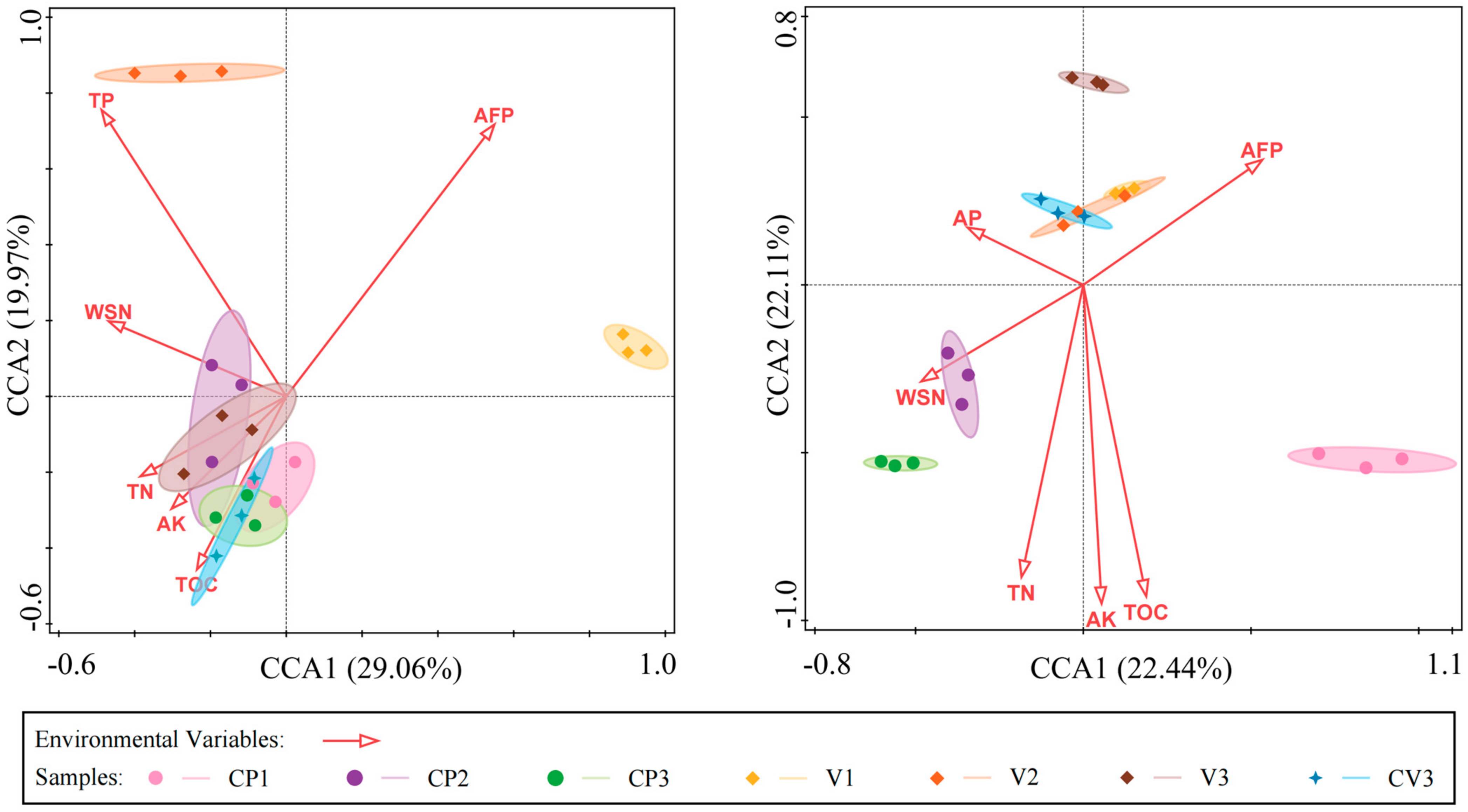
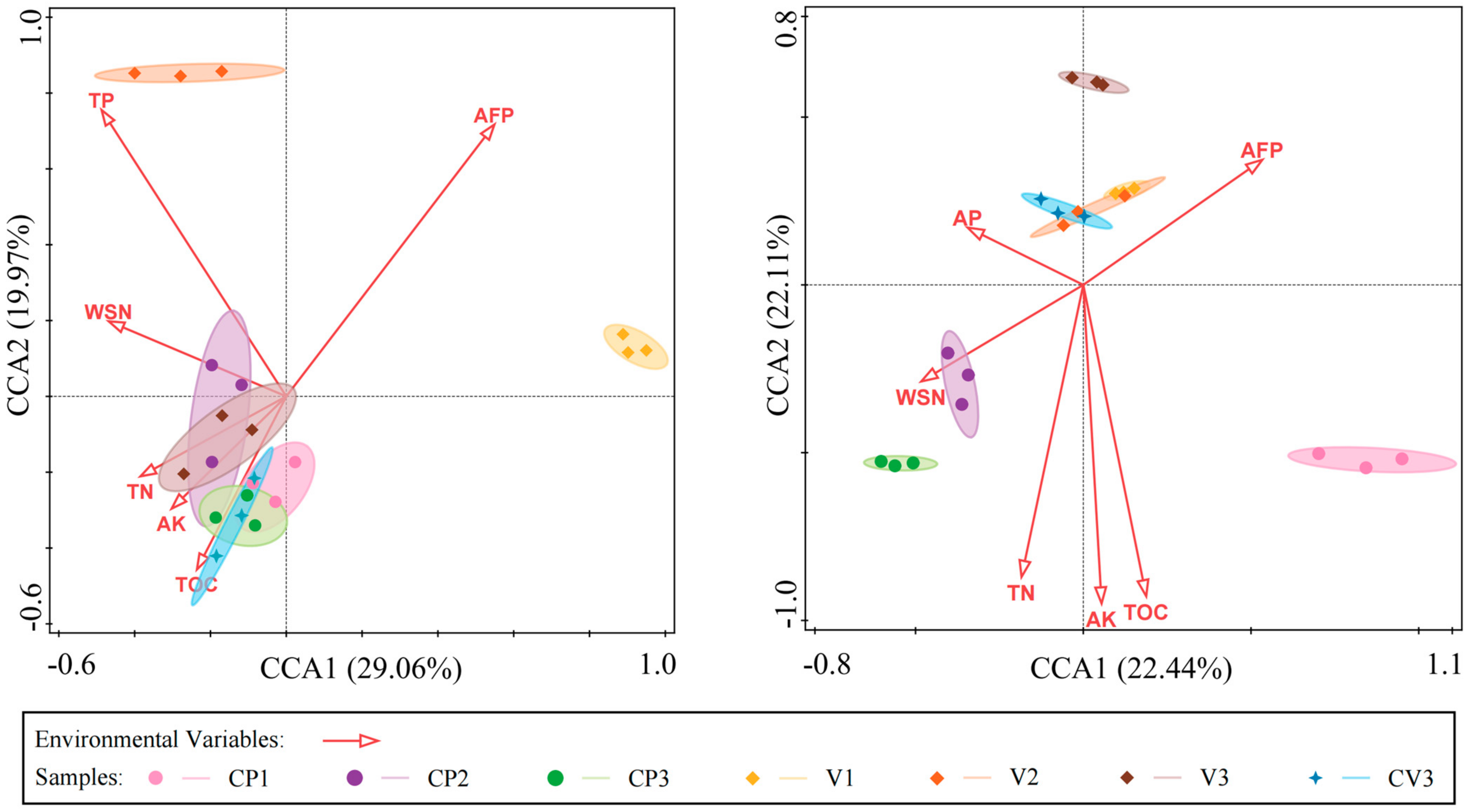
3.5. Influence of Substrate Physicochemical Properties on Microbial Communities
4. Discussion
4.1. Differential Microbial Diversity in Coconut Peat and Vermiculite
4.1.1. Differential Bacterial Diversity in Coconut Peat and Vermiculite
4.1.2. Differential Fungal Diversity in Coconut Peat and Vermiculite
4.2. Microbial Community Dynamics Associated with Substrates
4.2.1. Dynamics of Bacterial Communities Associated with Substrates
4.2.2. Dynamic Changes in Fungal Communities Associated with Substrates
4.3. Alterations in Microbial Community Functions in Substrates
4.3.1. Variations in Bacterial Community Functions in Substrates
4.3.2. Variations in Fungal Community Functionality across Substrates
4.4. Primary Environmental Factors Influencing Microbial Community Composition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Changes in Physical and Chemical Properties of Cultivation Substrates

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CP1 | CP2 | CP3 | V1 | V2 | V3 | CV3 | |
|---|---|---|---|---|---|---|---|
| pH | 6.15 ± 0.01 d | 5.96 ± 0.01 e | 6.54 ± 0.10 c | 7.09 ± 0.02 a | 6.60 ± 0.01 c | 7.07 ± 0.02 a | 6.94 ± 0.02 b |
| EC (uS/cm) | 723.33 ± 5.81 c | 1156 ± 5.13 a | 318.33 ± 2.19 e | 57.1 ± 0.66 g | 1056 ± 3.06 b | 560.33 ± 3.48 d | 208.33 ± 1.76 f |
| TOC (%) | 45.81 ± 1.31 a | 31.10 ± 0.54 b | 30.48 ± 0.70 b | 0.22 ± 0.01 f | 0.32 ± 0.01 e | 0.45 ± 0.01 d | 4.91 ± 0.01 c |
| TN (g/kg) | 8.61 ± 0.01 b | 11.34 ± 0.37 a | 8.55 ± 0.14 b | 2.83 ± 0.03 e | 5.21 ± 0.08 c | 2.40 ± 0.04 f | 4.89 ± 0.10 d |
| TP (%) | 0.13 ± 0.01 d | 0.23 ± 0.01 c | 0.07 ± 0.00 e | 0.11 ± 0.01 d | 0.55 ± 0.02 a | 0.34 ± 0.01 b | 0.25 ± 0.01 c |
| TK (%) | 0.76 ± 0.01 d | 0.66 ± 0.02 e | 0.63 ± 0.02 e | 3.22 ± 0.05 a | 2.80 ± 0.11 b | 2.77 ± 0.14 b | 2.24 ± 0.09 c |
| AP (mg/kg) | 90.83 ± 7.25 d | 1215.73 ± 96.30 a | 156.90 ± 8.88 c | 4.63 ± 0.13 e | 1129.67 ± 56.82 a | 547.67 ± 17.89 b | 507.63 ± 10.54 b |
| AK (mg/kg) | 10750.67 ± 357.45 a | 8168.33 ± 145.73 c | 8998.33 ± 349.96 b | 111.67 ± 1.45 g | 2219.33 ± 4.33 d | 313.33 ± 8.65 e | 208.67 ± 0.33 f |
| WSN (mg/kg) | 283.73 ± 2.22 e | 2038.13 ± 16.65 a | 677.67 ± 3.05 c | 10.53 ± 0.30 g | 886.23 ± 7.38 b | 520.93 ± 3.35 d | 158.37 ± 2.17 f |
| SP (%) | 64.80 ± 0.76 c | 72.83 ± 0.15 a | 70.17 ± 0.24 b | 72.27 ± 0.15 ab | 72.00 ± 1.50 ab | 64.90 ± 0.81 c | 65.30 ± 0.59 c |
| AFP (%) | 17.73 ± 0.12 b | 16.87 ± 0.15 c | 15.47 ± 0.09 e | 19.20 ± 0.06 a | 18.77 ± 0.09 a | 17.07 ± 0.23 c | 16.30 ± 0.23 d |
| WHP (%) | 47.03 ± 0.65 c | 56.00 ± 0.06 a | 54.73 ± 0.32 ab | 53.07 ± 0.12 b | 53.23 ± 1.41 b | 47.90 ± 0.64 c | 49.00 ± 0.36 c |
References
- Johnson, C.M.; Auat Cheein, F. Machinery for potato harvesting: A state-of-the-art review. Front. Plant Sci. 2023, 14, 1156734. [Google Scholar] [CrossRef]
- Hameed, A.; Zaidi, S.S.; Shakir, S.; Mansoor, S. Applications of New Breeding Technologies for Potato Improvement. Front. Plant Sci. 2018, 9, 925. [Google Scholar] [CrossRef]
- Devaux, A.; Goffart, J.-P.; Kromann, P.; Andrade-Piedra, J.; Polar, V.; Hareau, G. The Potato of the Future: Opportunities and Challenges in Sustainable Agri-food Systems. Potato Res. 2021, 64, 681–720. [Google Scholar] [CrossRef]
- Fogelman, E.; Oren-Shamir, M.; Hirschberg, J.; Mandolino, G.; Parisi, B.; Ovadia, R.; Tanami, Z.; Faigenboim, A.; Ginzberg, I. Nutritional value of potato (Solanum tuberosum) in hot climates: Anthocyanins, carotenoids, and steroidal glycoalkaloids. Planta 2019, 249, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Buono, V.; Paradiso, A.; Serio, F.; Gonnella, M.; De Gara, L.; Santamaria, P. Tuber quality and nutritional components of “early” potato subjected to chemical haulm desiccation. J. Food Comp. Analys. 2009, 22, 556–562. [Google Scholar] [CrossRef]
- Sharma, A.; Pandey, H.; Nampoothiri Devadas, V.A.S.; Kartha, B.D.; Jha, R. Production of, Factors Affecting, Gene Regulations, and Challenges in Tissue Cultured Plant through Soilless Culture. J. Agric. Food Chem. 2023, 71, 5804–5811. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, A.; Bar-Tal, A.; Silber, A.; Saha, U.; Raviv, M. Inorganic and Synthetic Organic Components of Soilless Culture and Potting Mixes. In Soilless Culture: Theory and Practice; Raviv, M., Lieth, J.H., Eds.; Academic Press: San Diego, CA, USA, 2008; pp. 505–544. [Google Scholar]
- Barrett, G.; Alexander, P.D.; Robinson, J.S.; Bragg, N.C. Achieving environmentally sustainable growing media for soilless plant cultivation systems—A review. Sci. Hortic. 2016, 212, 220–234. [Google Scholar] [CrossRef]
- Arenas, M.; Vavrina, C.; Cornell, J.; Hanlon, E.; Hochmuth, G. Coir as an Alternative to Peat in Media for Tomato Transplant Production. HortScience 2002, 37, 309–312. [Google Scholar] [CrossRef]
- Poulter, R. Quantifying differences between treated and untreated coir substrate. Acta Hortic. 2014, 1018, 557–564. [Google Scholar] [CrossRef]
- Mariotti, B.; Martini, S.; Raddi, S.; Tani, A.; Jacobs, D.F.; Oliet, J.A.; Maltoni, A. Coconut Coir as a Sustainable Nursery Growing Media for Seedling Production of the Ecologically Diverse Quercus Species. Forests 2020, 11, 522. [Google Scholar] [CrossRef]
- Wilkinson, K.M.; Landis, T.D.; Haase, D.L.; Daley, B.F.; Dumroese, R.K. Tropical Nursery Manual: A Guide to Starting and Operating a Nursery for Native and Traditional Plants; Agriculture Handbook 732; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 2014. [Google Scholar]
- Addison, J. Vermiculite: A review of the mineralogy and health effects of vermiculite exploitation. Regul. Toxicol. Pharmacol. 1995, 21, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Shen, Y.; Feng, H.; Wu, H.; Mao, R.; Ai, W.; Wu, Z. Study on primary physicochemical characteristics and nutrient adsorption of four plant cultivation substrates. Life Sci. Space Res. 2023, 36, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Kögel-Knabner, I. The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter: Fourteen years on. Soil Biol. Biochem. 2017, 105, A3–A8. [Google Scholar] [CrossRef]
- Vallance, J.; Déniel, F.; Floch, G.; Guérin-Dubrana, L.; Blancard, D.; Rey, P. Pathogenic and beneficial microorganisms in soilless cultures. Sustain. Agric. 2011, 2, 711–726. [Google Scholar] [CrossRef]
- Grunert, O.; Hernandez-Sanabria, E.; Vilchez-Vargas, R.; Jauregui, R.; Pieper, D.H.; Perneel, M.; Van Labeke, M.-C.; Reheul, D.; Boon, N. Mineral and organic growing media have distinct community structure, stability and functionality in soilless culture systems. Sci. Rep. 2016, 6, 18837. [Google Scholar] [CrossRef] [PubMed]
- Shara, S.A.; Zaharah, S.S.; Puteri, E.M.W. Physical, Chemical and Microbiological Properties of Different Combination of Soilless Media and Their Effect on the Vegetative Component and Nutrient Content of Hempedu Bumi (Andrographis paniculata). Pertanika J. Trop. Agric. Sci. 2017, 40, 35–52. [Google Scholar]
- Waller, P.; Thornton, C.; Farley, D.; Groenhof, A. Pathogens and other fungi in growing media constituents. Int. Symp. Grow. Media 2005, 779, 361–366. [Google Scholar] [CrossRef]
- Hyder, N.; Sims, J.J.; Wegulo, S.N. In vitro suppression of soilborne plant pathogens by coir. HortTechnology 2009, 19, 96–100. [Google Scholar] [CrossRef]
- Shameer, S.; Prasad, T.N.V.K.V. Plant growth promoting rhizobacteria for sustainable agricultural practices with special reference to biotic and abiotic stresses. Plant Growth Regul. 2018, 84, 603–615. [Google Scholar] [CrossRef]
- Gorbushina, A.A. Life on the rocks. Environ. Microbiol. 2007, 9, 1613–1631. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.F.; Zhao, F.; He, L.Y.; Qiu, G.; Chen, L. Isolation and characterization of silicate mineral-solubilizing Bacillus globisporus Q12 from the surfaces of weathered feldspar. Can. J. Microbiol. 2008, 54, 1064–1068. [Google Scholar] [CrossRef]
- Azua-Bustos, A.; Fairén, A.G.; González-Silva, C.; Prieto-Ballesteros, O.; Carrizo, D.; Sánchez-García, L.; Parro, V.; Fernández-Martínez, M.; Escudero, C.; Muñoz-Iglesias, V.; et al. Dark microbiome and extremely low organics in Atacama fossil delta unveil Mars life detection limits. Nat. Commun. 2023, 14, 808. [Google Scholar] [CrossRef]
- Napieralski, S.A.; Buss, H.L.; Roden, E.E. A Subsurface Fe-Silicate Weathering Microbiome. AGU Fall Meeting Abstracts. 2017. B31E-2035. Available online: https://ui.adsabs.harvard.edu/abs/2017AGUFM.B31E2035N (accessed on 2 March 2024).
- Napieralski, S.A.; Roden, E.E. The Weathering Microbiome of an Outcropping Granodiorite. Front. Microbiol. 2020, 11, 601907. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Huang, L.; Zhao, L.; Zeng, Q.; Liu, X.; Sheng, Y.; Shi, L.; Wu, G.; Jiang, H.; Li, F.; et al. A critical review of mineral-microbe interaction and co-evolution: Mechanisms and applications. Natl. Sci. Rev. 2022, 9, nwac128. [Google Scholar] [CrossRef]
- Nuppunen-Puputti, M.; Kietäväinen, R.; Kukkonen, I.; Bomberg, M. Implications of a short carbon pulse on biofilm formation on mica schist in microcosms with deep crystalline bedrock groundwater. Front. Microbiol. 2023, 14, 1054084. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, Y.; Shang, Y.; Liu, D.; Liesack, W.; Cui, Z.; Peng, J.; Zhang, F. Peat-vermiculite alters microbiota composition towards increased soil fertility and crop productivity. Plant Soil 2022, 470, 21–34. [Google Scholar] [CrossRef]
- Han, I.; Park, K.; Lee, H.; Park, B.; Lee, Y.; Kim, J. First report of root mat disease in a hydroponic tomato production system caused by rhizogenic Agrobacterium biovar 1 in South Korea. Plant Dis. 2021, 105, 1191. [Google Scholar] [CrossRef]
- Huo, C.; Cao, J.; Wu, K.; Chen, Y.; Zhao, Z. First Report of Pythium dissotocum causing root rot on hydroponically grown spinach in China. Plant Dis. 2020, 104, 3084. [Google Scholar] [CrossRef]
- McGehee, C.; Raudales, R.E.; Elmer, W.H. First report of Pythium dissotocum causing Pythium root rot on hydroponically grown lettuce in Connecticut. Plant Dis. 2018, 102, 2043. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Gilardi, G.; Pugliese, M.; Gullino, M.L. First Report of Multinucleate Rhizoctonia solani AG4 HG-I Causing Crown and Root Rot on Strawberry in Italy. Plant Dis. 2022, 106, 332. [Google Scholar] [CrossRef]
- Pitman, T.L.; Philbrook, R.N.; Warren, J.G. First Report of Pythium myriotylum Causing Root Rot in Cannabis sativa in California. Plant Dis. 2021, 105, 3766. [Google Scholar] [CrossRef]
- Martino, I.; Tabome, G.; Giordano, R.; Gullino, M.L.; Guarnaccia, V. First report of Diaporthe eres causing stem blight and dieback on highbush blueberry (Vaccinium corymbosum) in Italy. Plant Dis. 2023, 107, 1236. [Google Scholar] [CrossRef]
- Taparia, T.; Hendrix, E.; Nijhuis, E.; de Boer, W.; van der Wolf, J. Circular alternatives to peat in growing media: A microbiome perspective. J. Clean. Prod. 2021, 327, 129375. [Google Scholar] [CrossRef]
- Postma, J.; Van Os, E.; Bonants, P.J.M. Pathogen detection and management strategies in soilless plant growing systems. In Soilless Culture: Theory and Practice; Elsevier: Amsterdam, The Netherlands, 2008; pp. 425–457. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Kõljalg, U.; Larsson, K.-H.; Abarenkov, K.; Nilsson, R.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; et al. UNITE: A database providing web-based methods for the molecular identification of ectomycorrhizal fungi. New Phytol. 2005, 166, 1063–1068. [Google Scholar] [CrossRef] [PubMed]
- nmds.py—Nonmetric Multidimensional Scaling (NMDS). Available online: http://qiime.org/scripts/nmds.html (accessed on 15 August 2022).
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using CANOCO 5; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Kolde, R. pheatmap: Pretty Heatmaps. R Package Version 1.0.12. 2019. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 29 July 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 21 November 2022).
- Galaxy/Hutlab. Available online: http://huttenhower.sph.harvard.edu/galaxy/ (accessed on 8 July 2023).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.A.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Liu, Y. MicrobiomeStatPlot; GitHub: San Francisco, CA, USA, 2021; Available online: https://github.com/YongxinLiu/MicrobiomeStatPlot/tree/master/246TernaryPlot (accessed on 26 August 2022).
- Hamilton, N.E.; Ferry, M. ggtern: Ternary Diagrams Using ggplot2. J. Stat. Softw. Code Snippets 2018, 87, 1–17. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.6-2. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 29 July 2023).
- Revelle, W. psych: Procedures for Psychological, Psychometric, and Personality Research. R Package Version 1.0-95. Evanston, Illinois. 2013. Available online: https://CRAN.R-project.org/package=psych (accessed on 29 July 2023).
- Sansupa, C.; Wahdan, S.F.M.; Hossen, S.; Disayathanoowat, T.; Wubet, T.; Purahong, W. Can We Use Functional Annotation of Prokaryotic Taxa (FAPROTAX) to Assign the Ecological Functions of Soil Bacteria? Appl. Sci. 2021, 11, 688. [Google Scholar] [CrossRef]
- Functional Annotation of Prokaryotic Taxa (FAPROTAX). Available online: http://www.loucalab.com/archive/FAPROTAX/lib/php/ (accessed on 8 August 2022).
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Funguild_db. Available online: http://www.stbates.org/funguild_db.php (accessed on 8 August 2022).
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. InterJ. Complex Syst. 2006, 1695, 1–9. Available online: https://igraph.org (accessed on 13 September 2022).
- Harrell, F.E., Jr. Hmisc: Harrell Miscellaneous. R Package Version 4.7-1. 2022. Available online: https://CRAN.R-project.org/package=Hmisc (accessed on 13 September 2022).
- Jiménez-Vázquez, K.R.; García-Cárdenas, E.; Barrera-Ortiz, S.; Ortiz-Castro, R.; Ruiz-Herrera, L.F.; Ramos-Acosta, B.P.; Coria-Arellano, J.L.; Sáenz-Mata, J.; López-Bucio, J. The plant beneficial rhizobacterium Achromobacter sp. 5B1 influences root development through auxin signaling and redistribution. Plant J. 2020, 103, 1639–1654. [Google Scholar] [CrossRef]
- Samet, M.; Ghazala, I.; Karray, F.; Abid, C.; Chiab, N.; Nouri-Ellouz, O.; Sayadi, S.; Gargouri-Bouzid, R. Isolation of bacterial strains from compost teas and screening of their PGPR properties on potato plants. Environ. Sci. Pollut. Res. 2022, 29, 75365–75379. [Google Scholar] [CrossRef]
- Shahid, M.; Shah, A.A.; Basit, F.; Noman, M.; Zubair, M.; Ahmed, T.; Naqqash, T.; Manzoor, I.; Maqsood, A. Achromobacter sp. FB-14 harboring ACC deaminase activity augmented rice growth by upregulating the expression of stress-responsive CIPK genes under salinity stress. Braz. J. Microbiol. 2019, 51, 719–728. [Google Scholar] [CrossRef]
- Rokhbakhsh-Zamin, F.; Sachdev, D.; Kazemi-Pour, N.; Engineer, A.; Pardesi, K.R.; Zinjarde, S.S.; Dhakephalkar, P.K.; Chopade, B.A. Characterization of Plant-Growth-Promoting Traits of Acinetobacter Species Isolated from Rhizosphere of Pennisetum glaucum. J. Microbiol. Biotechnol. 2011, 21, 556–566. [Google Scholar] [CrossRef]
- Das, S.; Sultana, K.W.; Chandra, I. Isolation and Characterization of a Plant Growth-Promoting Bacterium Acinetobacter sp. SuKIC24 from In Vitro-Grown Basilicum polystachyon (L.) Moench. Curr. Microbiol. 2021, 78, 2961–2969. [Google Scholar] [CrossRef]
- Betoudji, F.; El Rahman, T.A.; Miller, M.J.; Ghosh, M.; Jacques, M.; Bouarab, K.; Malouin, F. A Siderophore Analog of Fimsbactin from Acinetobacter Hinders Growth of the Phytopathogen Pseudomonas syringae and Induces Systemic Priming of Immunity in Arabidopsis thaliana. Pathogens 2020, 9, 806. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.S.; Arshad, A.; Rajput, L.; Fatima, K.; Ullah, S.; Ahmad, M.; Imran, A. Growth-Stimulatory Effect of Quorum Sensing Signal Molecule N-Acyl-Homoserine Lactone-Producing Multi-Trait Aeromonas spp. on Wheat Genotypes Under Salt Stress. Front. Microbiol. 2020, 11, 553621. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Singh, S.K.; Peng, L.; Kaushal, R.; I Vílchez, J.; Shao, C.; Wu, X.; Zheng, S.; Morcillo, R.J.L.; Paré, P.W.; et al. Correction to: Flavonoid-attracted Aeromonas sp. from the Arabidopsis root microbiome enhances plant dehydration resistance. ISME J. 2022, 16, 2633. [Google Scholar] [CrossRef] [PubMed]
- Balázs, H.E.; Schmid, C.A.; Cruzeiro, C.; Podar, D.; Szatmari, P.-M.; Buegger, F.; Hufnagel, G.; Radl, V.; Schröder, P. Post-reclamation microbial diversity and functions in hexachlorocyclohexane (HCH) contaminated soil in relation to spontaneous HCH tolerant vegetation. Sci. Total. Environ. 2021, 767, 144653. [Google Scholar] [CrossRef]
- Madhaiyan, M.; Poonguzhali, S.; Senthilkumar, M.; Pragatheswari, D.; Lee, J.-S.; Lee, K.-C. Arachidicoccus rhizosphaerae gen. nov., sp. nov., a plant-growth-promoting bacterium in the family Chitinophagaceae isolated from rhizosphere soil. Int. J. Syst. Evol. Microbiol. 2015, 65, 578–586. [Google Scholar] [CrossRef]
- Vanissa, T.T.G.; Berger, B.; Patz, S.; Becker, M.; Turečková, V.; Novák, O.; Tarkowská, D.; Henri, F.; Ruppel, S. The Response of Maize to Inoculation with Arthrobacter sp. and Bacillus sp. in Phosphorus-Deficient, Salinity-Affected Soil. Microorganisms 2020, 8, 1005. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Singh, J.S.; Saxena, A.K.; Singh, D.P. Impact of PGPR inoculation on growth and antioxidant status of wheat under saline conditions. Plant Biol. 2011, 14, 605–611. [Google Scholar] [CrossRef]
- Zhao, S.; Xu, W.; Zhang, W.; Wu, H.; Guang, C.; Mu, W. In-depth biochemical identification of a novel methyl parathion hydrolase from Azohydromonas australica and its high effectiveness in the degradation of various organophosphorus pesticides. Bioresour. Technol. 2021, 323, 124641. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.-H.; Yokota, A. Reclassification of Alcaligenes latus strains IAM 12599T and IAM 12664 and Pseudomonas saccharophila as Azohydromonas lata gen. nov., comb. nov., Azohydromonas australica sp. nov. and Pelomonas saccharophila gen. nov., comb. nov., respectively. Int. J. Syst. Evol. Microbiol. 2005, 55, 2419–2425. [Google Scholar] [CrossRef] [PubMed]
- Siani, R.; Siani, R.; Stabl, G.; Stabl, G.; Gutjahr, C.; Gutjahr, C.; Schloter, M.; Schloter, M.; Radl, V.; Radl, V. Acidovorax pan-genome reveals specific functional traits for plant beneficial and pathogenic plant-associations. Microb. Genom. 2021, 7, 000666. [Google Scholar] [CrossRef]
- Hashem, A.; Tabassum, B.; Fathi Abd Allah, E. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi J. Biol. Sci. 2019, 26, 1291–1297. [Google Scholar] [CrossRef]
- Arashida, H.; Kugenuma, T.; Watanabe, M.; Maeda, I. Nitrogen fixation in Rhodopseudomonas palustris co-cultured with Bacillus subtilis in the presence of air. J. Biosci. Bioeng. 2018, 127, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Salwan, R.; Sharma, V.; Saini, R.; Pandey, M. Identification of plant beneficial Bacillus spp. for Resilient agricultural ecosystem. Curr. Res. Microb. Sci. 2021, 2, 100046. [Google Scholar] [CrossRef]
- Glick, W.H.; Miller, C.C.; Cardinal, L.B. Making a life in the field of organization science. J. Organ. Behav. 2007, 28, 817–835. [Google Scholar] [CrossRef]
- Fira, D.; Dimkić, I.; Berić, T.; Lozo, J.; Stanković, S. Biological control of plant pathogens by Bacillus species. J. Biotechnol. 2018, 285, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Narayanasamy, S.; Thangappan, S.; Uthandi, S. Plant Growth-Promoting Bacillus sp. Cahoots Moisture Stress Alleviation in Rice Genotypes by Triggering Antioxidant Defense System. Microbiol. Res. 2020, 239, 126518. [Google Scholar] [CrossRef]
- Andrews, M.; Andrews, M.E. Specificity in Legume-Rhizobia Symbioses. Int. J. Mol. Sci. 2017, 18, 705. [Google Scholar] [CrossRef] [PubMed]
- Shah, V.; Subramaniam, S. Bradyrhizobium japonicum USDA110: A representative model organism for studying the impact of pollutants on soil microbiota. Sci. Total. Environ. 2018, 624, 963–967. [Google Scholar] [CrossRef]
- Singh, N.; Marwa, N.; Mishra, S.K.; Mishra, J.; Verma, P.C.; Rathaur, S.; Singh, N. Brevundimonas diminuta mediated alleviation of arsenic toxicity and plant growth promotion in Oryza sativa L. Ecotoxicol. Environ. Saf. 2016, 125, 25–34. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, V.; Gupta, P.; Singh, A. Conjoint application of novel bacterial isolates on dynamic changes in oxidative stress responses of axenic Brassica juncea L. in Hg-stress soils. J. Hazard. Mater. 2022, 434, 128854. [Google Scholar] [CrossRef]
- Ravi, A.; Theresa, M.; Nandayipurath, V.V.T.; Rajan, S.; Khalid, N.K.; Thankappanpillai, A.C.; Krishnankutty, R.E. Plant Beneficial Features and Application of Paraburkholderia sp. NhPBG1 Isolated from Pitcher of Nepenthes hamblack. Probiotics Antimicrob. Proteins 2020, 13, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Herpell, J.B.; Vanwijnsberghe, S.; Peeters, C.; Schindler, F.; Fragner, L.; Bejtović, M.; Weckwerth, W.; Vandamme, P. Paraburkholderia dioscoreae sp. nov., a novel plant associated growth promotor. Int. J. Syst. Evol. Microbiol. 2021, 71, 004969. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Wang, L.; Deng, L.; Mei, X.; Liu, Y.; Huang, H.; Du, F.; Zhu, S.; Yang, M. Enrichment of Burkholderia in the Rhizosphere by Autotoxic Ginsenosides to Alleviate Negative Plant-Soil Feedback. Microbiol. Spectr. 2021, 9, e0140021. [Google Scholar] [CrossRef] [PubMed]
- Berrios, L. Plant-growth-promoting Caulobacter strains isolated from distinct plant hosts share conserved genetic factors involved in beneficial plant–bacteria interactions. Arch. Microbiol. 2021, 204, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Berrios, L. Complete Genome Sequence of the Plant-Growth-Promoting Bacterium Caulobacter segnis CBR1. Curr. Microbiol. 2021, 78, 2935–2942. [Google Scholar] [CrossRef]
- Berrios, L. The genus Caulobacter and its role in plant microbiomes. World J. Microbiol. Biotechnol. 2022, 38, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Suarez, C.; Ratering, S.; Kramer, I.; Schnell, S. Cellvibrio diazotrophicus sp. nov., a nitrogen-fixing bacteria isolated from the rhizosphere of salt meadow plants and emended description of the genus Cellvibrio. Int. J. Syst. Evol. Microbiol. 2014, 64, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, J.; Wang, E.; Wang, N. Mechanisms Underlying the Rhizosphere-To-Rhizoplane Enrichment of Cellvibrio Unveiled by Genome-Centric Metagenomics and Metatranscriptomics. Microorganisms 2020, 8, 583. [Google Scholar] [CrossRef]
- Kumar, V.; Sharma, N.; Maitra, S.S.; Lakkaboyana, S.K. In vivo removal of profenofos in agricultural soil and plant growth promoting activity on Vigna radiata by efficient bacterial formulation. Int. J. Phytoremediation 2019, 22, 585–593. [Google Scholar] [CrossRef]
- Khalofah, A.; Kilany, M.; Migdadi, H. Phytostimulatory Influence of Comamonas testosteroni and Silver Nanoparticles on Linum usitatissimum L. under Salinity Stress. Plants 2021, 10, 790. [Google Scholar] [CrossRef]
- Suchan, D.M.; Bergsveinson, J.; Manzon, L.; Pierce, A.; Kryachko, Y.; Korber, D.; Tan, Y.; Tambalo, D.D.; Khan, N.H.; Whiting, M.; et al. Transcriptomics reveal core activities of the plant growth-promoting bacterium Delftia acidovorans RAY209 during interaction with canola and soybean roots. Microb. Genom. 2020, 6, e000462. [Google Scholar] [CrossRef]
- Cagide, C.; Riviezzi, B.; Minteguiaga, M.; Morel, M.A.; Castro-Sowinski, S. Identification of Plant Compounds Involved in the Microbe-Plant Communication during the Coinoculation of Soybean with Bradyrhizobium elkanii and Delftia sp. strain JD2. Mol. Plant-Microbe Interact. 2018, 31, 1192–1199. [Google Scholar] [CrossRef]
- Morel, M.A.; Ubalde, M.C.; Braña, V.; Castro-Sowinski, S. Delftia sp. JD2: A potential Cr(VI)-reducing agent with plant growth-promoting activity. Arch. Microbiol. 2010, 193, 63–68. [Google Scholar] [CrossRef]
- Chhetri, G.; Kim, I.; Kang, M.; Kim, J.; So, Y.; Seo, T. Devosia rhizoryzae sp. nov., and Devosia oryziradicis sp. nov., novel plant growth promoting members of the genus Devosia, isolated from the rhizosphere of rice plants. J. Microbiol. 2021, 60, 1–10. [Google Scholar] [CrossRef]
- Haack, F.S.; Poehlein, A.; Kröger, C.; Voigt, C.A.; Piepenbring, M.; Bode, H.B.; Daniel, R.; Schäfer, W.; Streit, W.R. Molecular Keys to the Janthinobacterium and Duganella spp. Interaction with the Plant Pathogen Fusarium graminearum. Front. Microbiol. 2016, 7, 1668. [Google Scholar] [CrossRef]
- Arroyo-Herrera, I.; Román-Ponce, B.; Reséndiz-Martínez, A.L.; Santos, P.E.-D.L.; Wang, E.T.; Vásquez-Murrieta, M.S. Heavy-metal resistance mechanisms developed by bacteria from Lerma–Chapala basin. Arch. Microbiol. 2021, 203, 1807–1823. [Google Scholar] [CrossRef] [PubMed]
- de Lima, D.R.M.; dos Santos, I.B.; Oliveira, J.T.C.; da Costa, D.P.; de Queiroz, J.V.J.; Romagnoli, E.M.; Andreote, F.D.; Freire, F.J.; Kuklinsky-Sobral, J. Genetic diversity of N-fixing and plant growth-promoting bacterial community in different sugarcane genotypes, association habitat and phenological phase of the crop. Arch. Microbiol. 2020, 203, 1089–1105. [Google Scholar] [CrossRef] [PubMed]
- Tapia-García, E.Y.; Hernández-Trejo, V.; Guevara-Luna, J.; Rojas-Rojas, F.U.; Arroyo-Herrera, I.; Meza-Radilla, G.; Vásquez-Murrieta, M.S.; Santos, P.E.-D.L. Plant growth-promoting bacteria isolated from wild legume nodules and nodules of Phaseolus vulgaris L. trap plants in central and southern Mexico. Microbiol. Res. 2020, 239, 126522. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, B.; Shahid, M.; Syed, A.; Rajput, V.D.; Elgorban, A.M.; Minkina, T.; Bahkali, A.H.; Lee, J. Drought Tolerant Enterobacter sp./Leclercia adecarboxylata Secretes Indole-3-acetic Acid and Other Biomolecules and Enhances the Biological Attributes of Vigna radiata (L.) R. Wilczek in Water Deficit Conditions. Biology 2021, 10, 1149. [Google Scholar] [CrossRef] [PubMed]
- Ranawat, B.; Bachani, P.; Singh, A.; Mishra, S. Enterobacter hormaechei as Plant Growth-Promoting Bacteria for Improvement in Lycopersicum esculentum. Curr. Microbiol. 2021, 78, 1208–1217. [Google Scholar] [CrossRef]
- Synek, L.; Rawat, A.; L’Haridon, F.; Weisskopf, L.; Saad, M.M.; Hirt, H. Multiple strategies of plant colonization by beneficial endophytic Enterobacter sp. SA187. Environ. Microbiol. 2021, 23, 6223–6240. [Google Scholar] [CrossRef] [PubMed]
- Andrés-Barrao, C.; Alzubaidy, H.; Jalal, R.; Mariappan, K.G.; de Zélicourt, A.; Bokhari, A.; Artyukh, O.; Alwutayd, K.; Rawat, A.; Shekhawat, K.; et al. Coordinated bacterial and plant sulfur metabolism in Enterobacter sp. SA187–induced plant salt stress tolerance. Proc. Natl. Acad. Sci. USA 2021, 118, e2107417118. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Kumar, V.; Usmani, Z.; Rani, R.; Chandra, A.; Gupta, V.K. Implications of plant growth promoting Klebsiella sp. CPSB4 and Enterobacter sp. CPSB49 in luxuriant growth of tomato plants under chromium stress. Chemosphere 2019, 240, 124944. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.K.; Gohel, K.; Patel, H.; Solanki, T. Wheat Growth Dependent Succession of Culturable Endophytic Bacteria and Their Plant Growth Promoting Traits. Curr. Microbiol. 2021, 78, 4103–4114. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Bano, A. Role of PGPR in the reclamation and revegetation of saline land. Pak. J. Bot. 2018, 51, 27–35. [Google Scholar] [CrossRef]
- Bharti, N.; Yadav, D.; Barnawal, D.; Maji, D.; Kalra, A. Exiguobacterium oxidotolerans, a halotolerant plant growth promoting rhizobacteria, improves yield and content of secondary metabolites in Bacopa monnieri (L.) Pennell under primary and secondary salt stress. World J. Microbiol. Biotechnol. 2012, 29, 379–387. [Google Scholar] [CrossRef]
- E A Lidbury, I.D.; Borsetto, C.; Murphy, A.R.J.; Bottrill, A.; E Jones, A.M.; Bending, G.D.; Hammond, J.P.; Chen, Y.; Wellington, E.M.H.; Scanlan, D.J. Niche-adaptation in plant-associated Bacteroidetes favours specialisation in organic phosphorus mineralisation. ISME J. 2020, 15, 1040–1055. [Google Scholar] [CrossRef]
- Gontia-Mishra, I.; Sapre, S.; Sharma, A.; Tiwari, S. Amelioration of drought tolerance in wheat by the interaction of plant growth-promoting rhizobacteria. Plant Biol. 2016, 18, 992–1000. [Google Scholar] [CrossRef]
- Menon, R.R.; Kumari, S.; Viver, T.; Rameshkumar, N. Flavobacterium pokkalii sp. nov., a novel plant growth promoting native rhizobacteria isolated from pokkali rice grown in coastal saline affected agricultural regions of southern India, Kerala. Microbiol. Res. 2020, 240, 126533. [Google Scholar] [CrossRef]
- Liu, C.; Zhuang, J.; Wang, J.; Fan, G.; Feng, M.; Zhang, S. Soil bacterial communities of three types of plants from ecological restoration areas and plant-growth promotional benefits of Microbacterium invictum (strain X-18). Front. Microbiol. 2022, 13, 926037. [Google Scholar] [CrossRef]
- Shang, J.; Liu, B. Application of a microbial consortium improves the growth of Camellia sinensis and influences the indigenous rhizosphere bacterial communities. J. Appl. Microbiol. 2020, 130, 2029–2040. [Google Scholar] [CrossRef]
- Gagnon, V.; Rodrigue-Morin, M.; Tremblay, J.; Wasserscheid, J.; Champagne, J.; Bellenger, J.-P.; Greer, C.W.; Roy, S. Vegetation drives the structure of active microbial communities on an acidogenic mine tailings deposit. PeerJ 2020, 8, e10109. [Google Scholar] [CrossRef] [PubMed]
- Kundim, B.A.; Itou, Y.; Sakagami, Y.; Fudou, R.; Iizuka, T.; Yamanaka, S.; Ojika, M. New haliangicin isomers, potent antifungal metabolites produced by a marine myxobacterium. J. Antibiot. 2003, 56, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Alves, L.P.S.; do Amaral, F.P.; Kim, D.; Bom, M.T.; Gavídia, M.P.; Teixeira, C.S.; Holthman, F.; de Oliveira Pedrosa, F.; De Souza, E.M.; Chubatsu, L.S.; et al. Importance of Poly-3-Hydroxybutyrate Metabolism to the Ability ofHerbaspirillum seropedicaeTo Promote Plant Growth. Appl. Environ. Microbiol. 2019, 85, e02586-18. [Google Scholar] [CrossRef]
- Wang, X.; Zeng, X.; Luo, L.; Chen, X.; Yan, H.; Xie, Z.; Zhou, Y. Plant Growth-Promoting Activity of Herbaspirillum aquaticum ZXN111 on the Zijuan Tea Plant (Camellia sinensis var. assamica). J. Agric. Food Chem. 2022, 70, 3757–3764. [Google Scholar] [CrossRef] [PubMed]
- Grillo-Puertas, M.; Villegas, J.M.; Pankievicz, V.C.S.; Tadra-Sfeir, M.Z.; Mota, F.J.T.; Hebert, E.M.; Brusamarello-Santos, L.; Pedraza, R.O.; Pedrosa, F.O.; Rapisarda, V.A.; et al. Transcriptional Responses of Herbaspirillum seropedicae to Environmental Phosphate Concentration. Front. Microbiol. 2021, 12, 666277. [Google Scholar] [CrossRef] [PubMed]
- Chubatsu, L.S.; Monteiro, R.A.; de Souza, E.M.; de Oliveira, M.A.S.; Yates, M.G.; Wassem, R.; Bonatto, A.C.; Huergo, L.F.; Steffens, M.B.R.; Rigo, L.U.; et al. Nitrogen fixation control in Herbaspirillum seropedicae. Plant Soil 2011, 356, 197–207. [Google Scholar] [CrossRef]
- Cortés-Patiño, S.; Vargas, C.; Álvarez-Flórez, F.; Bonilla, R.; Estrada-Bonilla, G. Potential of Herbaspirillum and Azospirillum Consortium to Promote Growth of Perennial Ryegrass under Water Deficit. Microorganisms 2021, 9, 91. [Google Scholar] [CrossRef]
- Rat, A.; Naranjo, H.D.; Krigas, N.; Grigoriadou, K.; Maloupa, E.; Alonso, A.V.; Schneider, C.; Papageorgiou, V.P.; Assimopoulou, A.N.; Tsafantakis, N.; et al. Endophytic Bacteria From the Roots of the Medicinal Plant Alkanna tinctoria Tausch (Boraginaceae): Exploration of Plant Growth Promoting Properties and Potential Role in the Production of Plant Secondary Metabolites. Front. Microbiol. 2021, 12, 633488. [Google Scholar] [CrossRef]
- Guglielmetti, S.; Basilico, R.; Taverniti, V.; Arioli, S.; Piagnani, C.; Bernacchi, A. Luteibacter rhizovicinus MIMR1 promotes root development in barley (Hordeum vulgare L.) under laboratory conditions. World J. Microbiol. Biotechnol. 2013, 29, 2025–2032. [Google Scholar] [CrossRef]
- Abdelkrim, S.; Jebara, S.H.; Saadani, O.; Chiboub, M.; Abid, G.; Jebara, M. Effect of Pb-resistant plant growth-promoting rhizobacteria inoculation on growth and lead uptake by Lathyrus sativus. J. Basic Microbiol. 2018, 58, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-T.; Monkhung, S.; Lee, Y.S.; Kim, K.Y. Effects of Lysobacter antibioticus HS124, an effective biocontrol agent against Fusarium graminearum, on crown rot disease and growth promotion of wheat. Can. J. Microbiol. 2019, 65, 904–912. [Google Scholar] [CrossRef]
- Rosenzweig, N.; Tiedje, J.M.; Quensen, J.F.; Meng, Q.; Hao, J.J. Microbial Communities Associated with Potato Common Scab-Suppressive Soil Determined by Pyrosequencing Analyses. Plant Dis. 2012, 96, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Vlassi, A.; Nesler, A.; Perazzolli, M.; Lazazzara, V.; Büschl, C.; Parich, A.; Puopolo, G.; Schuhmacher, R. Volatile Organic Compounds from Lysobacter capsici AZ78 as Potential Candidates for Biological Control of Soilborne Plant Pathogens. Front. Microbiol. 2020, 11, 1748. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Xu, K.; Shen, D.; Chou, S.; Gomelsky, M.; Qian, G. Antifungal weapons of Lysobacter, a mighty biocontrol agent. Environ. Microbiol. 2021, 23, 5704–5715. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.; Pan, C.; Su, X.; Zhou, X.; Bao, Y. Combined effects of oxytetracycline and microplastic on wheat seedling growth and associated rhizosphere bacterial communities and soil metabolite profiles. Environ. Pollut. 2022, 302, 119046. [Google Scholar] [CrossRef] [PubMed]
- Chimwamurombe, P.M.; Grönemeyer, J.L.; Reinhold-Hurek, B. Isolation and characterization of culturable seed-associated bacterial endophytes from gnotobiotically grown Marama bean seedlings. FEMS Microbiol. Ecol. 2016, 92, fiw083. [Google Scholar] [CrossRef]
- Zheng, B.-X.; Bi, Q.-F.; Hao, X.-L.; Zhou, G.-W.; Yang, X.-R. Massilia phosphatilytica sp. nov., a phosphate solubilizing bacteria isolated from a long-term fertilized soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 2514–2519. [Google Scholar] [CrossRef]
- Li, C.; Cao, P.; Du, C.; Zhang, X.; Bing, H.; Li, L.; Sun, P.; Xiang, W.; Zhao, J.; Wang, X. Massilia rhizosphaerae sp. nov., a rice-associated rhizobacterium with antibacterial activity against Ralstonia solanacearum. Int. J. Syst. Evol. Microbiol. 2021, 71, 005009. [Google Scholar] [CrossRef]
- Baba, Z.A.; Hamid, B.; Sheikh, T.A.; Alotaibi, S.H.; El Enshasy, H.A.; Ansari, M.J.; Zuan, A.T.K.; Sayyed, R.Z. Psychrotolerant Mesorhizobium sp. Isolated from Temperate and Cold Desert Regions Solubilizes Potassium and Produces Multiple Plant Growth Promoting Metabolites. Molecules 2021, 26, 5758. [Google Scholar] [CrossRef]
- Vijayabharathi, R.; Gopalakrishnan, S.; Sathya, A.; Srinivas, V.; Sharma, M. Deciphering the tri-dimensional effect of endophytic Streptomyces sp. on chickpea for plant growth promotion, helper effect with Mesorhizobium ciceri and host-plant resistance induction against Botrytis cinerea. Microb. Pathog. 2018, 122, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Muleta, A.; Tesfaye, K.; Selassie, T.H.H.; Cook, D.R.; Assefa, F. Phosphate solubilization and multiple plant growth promoting properties of Mesorhizobium species nodulating chickpea from acidic soils of Ethiopia. Arch. Microbiol. 2021, 203, 2129–2137. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, F.X.; Brígido, C.; Glick, B.R.; Oliveira, S. ACC deaminase genes are conserved among Mesorhizobium species able to nodulate the same host plant. FEMS Microbiol. Lett. 2012, 336, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Khan, M.S.; Syed, A.; Marraiki, N.; Elgorban, A.M. Mesorhizobium ciceri as biological tool for improving physiological, biochemical and antioxidant state of Cicer aritienum (L.) under fungicide stress. Sci. Rep. 2021, 11, 9655. [Google Scholar] [CrossRef] [PubMed]
- Santiago, C.D.; Yagi, S.; Ijima, M.; Nashimoto, T.; Sawada, M.; Ikeda, S.; Asano, K.; Orikasa, Y.; Ohwada, T. Bacterial Compatibility in Combined Inoculations Enhances the Growth of Potato Seedlings. Microbes Environ. 2017, 32, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Agafonova, N.V.; Kaparullina, E.N.; Doronina, N.V.; Trotsenko, Y.A. Phosphate-solubilizing activity of aerobic methylobacteria. Microbiology 2013, 82, 864–867. [Google Scholar] [CrossRef]
- Agafonova, N.V.; Doronina, N.V.; Trotsenko, Y.A. Enhanced Resistance of Pea Plants to Oxidative Stress Caused by Paraquat during Colonization by Aerobic Methylobacteria. Appl. Biochem. Microbiol. 2016, 52, 199–204. [Google Scholar] [CrossRef]
- Kataoka, R.; Akashi, M.; Taniguchi, T.; Kinose, Y.; Yaprak, A.E.; Turgay, O.C. Metabolomics Analyses Reveal Metabolites Affected by Plant Growth-Promoting Endophytic Bacteria in Roots of the Halophyte Mesembryanthemum crystallinum. Int. J. Mol. Sci. 2021, 22, 11813. [Google Scholar] [CrossRef]
- Singh, T.; Singh, D.K. Rhizospheric Microbacterium sp. P27 Showing Potential of Lindane Degradation and Plant Growth Promoting Traits. Curr. Microbiol. 2019, 76, 888–895. [Google Scholar] [CrossRef]
- Madhaiyan, M.; Poonguzhali, S.; Lee, J.-S.; Senthilkumar, M.; Lee, K.C.; Sundaram, S. Mucilaginibacter gossypii sp. nov. and Mucilaginibacter gossypiicola sp. nov., plant-growth-promoting bacteria isolated from cotton rhizosphere soils. Int. J. Syst. Evol. Microbiol. 2010, 60, 2451–2457. [Google Scholar] [CrossRef]
- Fan, D.; Smith, D.L. Characterization of Selected Plant Growth-Promoting Rhizobacteria and Their Non-Host Growth Promotion Effects. Microbiol. Spectr. 2021, 9, e0027921. [Google Scholar] [CrossRef]
- Fan, D.; Smith, D.L. Mucilaginibacter sp. K Improves Growth and Induces Salt Tolerance in Nonhost Plants via Multilevel Mechanisms. Front. Plant Sci. 2022, 13, 938697. [Google Scholar] [CrossRef]
- Fan, D.; Subramanian, S.; Smith, D.L. Plant endophytes promote growth and alleviate salt stress in Arabidopsis thaliana. Sci. Rep. 2020, 10, 12740. [Google Scholar] [CrossRef]
- Liu, S.-Y.; Wei, C.-Y.; Tong, Y.; Chen, W.; Han, Z.-Y.; Zeng, D.-Q.; Tang, W.-W. Cyperus rotundus L. drives arable soil infertile by changing the structure of soil bacteria in the rhizosphere, using a maize field as an example. Environ. Sci. Pollut. Res. 2022, 29, 79579–79593. [Google Scholar] [CrossRef]
- Yan, S.; Ren, T.; Mahari, W.A.W.; Feng, H.; Xu, C.; Yun, F.; Waiho, K.; Wei, Y.; Lam, S.S.; Liu, G. Soil carbon supplementation: Improvement of root-surrounding soil bacterial communities, sugar and starch content in tobacco (N. tabacum). Sci. Total. Environ. 2021, 802, 149835. [Google Scholar] [CrossRef]
- Vives-Peris, V.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Salt stress alleviation in citrus plants by plant growth-promoting rhizobacteria Pseudomonas putida and Novosphingobium sp. Plant Cell Rep. 2018, 37, 1557–1569. [Google Scholar] [CrossRef]
- Islam, R.; Sultana, T.; Joe, M.M.; Yim, W.; Cho, J.; Sa, T. Nitrogen-fixing bacteria with multiple plant growth-promoting activities enhance growth of tomato and red pepper. J. Basic Microbiol. 2013, 53, 1004–1015. [Google Scholar] [CrossRef]
- Yu, X.; Li, Y.; Cui, Y.; Liu, R.; Chen, Q.; Gu, Y.; Zhao, K.; Xiang, Q.; Xu, K.; Zhang, X. An indoleacetic acid-producing Ochrobactrum sp. MGJ11 counteracts cadmium effect on soybean by promoting plant growth. J. Appl. Microbiol. 2016, 122, 987–996. [Google Scholar] [CrossRef]
- Lafi, F.F.; Alam, I.; Geurts, R.; Bisseling, T.; Bajic, V.B.; Hirt, H.; Saad, M.M. Draft Genome Sequence of Ochrobactrum intermedium Strain SA148, a Plant Growth-Promoting Desert Rhizobacterium. Genome Announc. 2017, 5, e01707-16. [Google Scholar] [CrossRef] [PubMed]
- Sherpa, M.T.; Bag, N.; Das, S.; Haokip, P.; Sharma, L. Isolation and characterization of plant growth promoting rhizobacteria isolated from organically grown high yielding pole type native pea (Pisum sativum L.) variety Dentami of Sikkim, India. Curr. Res. Microb. Sci. 2021, 2, 100068. [Google Scholar] [CrossRef]
- Riva, V.; Mapelli, F.; Dragonetti, G.; Elfahl, M.; Vergani, L.; Crepaldi, P.; La Maddalena, N.; Borin, S. Bacterial Inoculants Mitigating Water Scarcity in Tomato: The Importance of Long-Term in vivo Experiments. Front. Microbiol. 2021, 12, 675552. [Google Scholar] [CrossRef]
- Liu, X.; Li, Q.; Li, Y.; Guan, G.; Chen, S. Paenibacillus strains with nitrogen fixation and multiple beneficial properties for promoting plant growth. PeerJ 2019, 7, e7445. [Google Scholar] [CrossRef]
- Liu, W.; Sikora, E.; Park, S.-W. Plant growth-promoting rhizobacterium, Paenibacillus polymyxa CR1, upregulates dehydration-responsive genes, RD29A and RD29B, during priming drought tolerance in arabidopsis. Plant Physiol. Biochem. 2020, 156, 146–154. [Google Scholar] [CrossRef]
- Enquahone, S.; van Marle, G.; Simachew, A. Plant growth-promoting characteristics of halotolerant endophytic bacteria isolated from Sporobolus specatus (Vahr) Kunth and Cyperus laevigatus L. of Ethiopian rift valley lakes. Arch. Microbiol. 2022, 204, 403. [Google Scholar] [CrossRef]
- Sahoo, B.; Ningthoujam, R.; Chaudhuri, S. Isolation and characterization of a lindane degrading bacteria Paracoccus sp. NITDBR1 and evaluation of its plant growth promoting traits. Int. Microbiol. 2018, 22, 155–167. [Google Scholar] [CrossRef]
- Rai, A.; N, S.; Ch, S.; Ch, V.R. Paracoccus aeridis sp. nov., an indole-producing bacterium isolated from the rhizosphere of an orchid, Aerides maculosa. Int. J. Syst. Evol. Microbiol. 2020, 70, 1720–1728. [Google Scholar] [CrossRef]
- Morais, M.C.; Mucha, Â.; Ferreira, H.; Gonçalves, B.; Bacelar, E.; Marques, G. Comparative study of plant growth-promoting bacteria on the physiology, growth and fruit quality of strawberry. J. Sci. Food Agric. 2019, 99, 5341–5349. [Google Scholar] [CrossRef]
- Zappelini, C.; Alvarez-Lopez, V.; Capelli, N.; Guyeux, C.; Chalot, M. Streptomyces Dominate the Soil Under Betula Trees That Have Naturally Colonized a Red Gypsum Landfill. Front. Microbiol. 2018, 9, 1772. [Google Scholar] [CrossRef]
- Makino, A.; Nakai, R.; Yoneda, Y.; Toyama, T.; Tanaka, Y.; Meng, X.-Y.; Mori, K.; Ike, M.; Morikawa, M.; Kamagata, Y.; et al. Isolation of Aquatic Plant Growth-Promoting Bacteria for the Floating Plant Duckweed (Lemna minor). Microorganisms 2022, 10, 1564. [Google Scholar] [CrossRef]
- Jasim, B.; Anish, M.C.; Shimil, V.; Jyothis, M.; Radhakrishnan, E.K. Studies on Plant Growth Promoting Properties of Fruit-Associated Bacteria from Elettaria cardamomum and Molecular Analysis of ACC Deaminase Gene. Appl. Biochem. Biotechnol. 2015, 177, 175–189. [Google Scholar] [CrossRef]
- Wedage, W.M.M.; Aberathne, A.H.M.N.R.; Harischandra, I.N.; Gunawardana, D. A Nodulation-Proficient Nonrhizobial Inhabitant of Pueraria phaseoloides. Sci. World J. 2019, 2019, 1–9. [Google Scholar] [CrossRef]
- Ham, S.H.; Yoon, A.R.; Oh, H.E.; Park, Y.G. Plant Growth-Promoting Microorganism Pseudarthrobacter sp. NIBRBAC000502770 Enhances the Growth and Flavonoid Content of Geum aleppicum. Microorganisms 2022, 10, 1241. [Google Scholar] [CrossRef]
- Shi, R.; Wang, S.; Xiong, B.; Gu, H.; Wang, H.; Ji, C.; Jia, W.; Horowitz, A.R.; Zhen, W.; Ben Asher, J.; et al. Application of Bioorganic Fertilizer on Panax notoginseng Improves Plant Growth by Altering the Rhizosphere Microbiome Structure and Metabolism. Microorganisms 2022, 10, 275. [Google Scholar] [CrossRef]
- Biessy, A.; Filion, M. Phenazines in plant-beneficial Pseudomonas spp.: Biosynthesis, regulation, function and genomics. Environ. Microbiol. 2018, 20, 3905–3917. [Google Scholar] [CrossRef]
- Tang, X.; Zhong, R.; Jiang, J.; He, L.; Huang, Z.; Shi, G.; Wu, H.; Liu, J.; Xiong, F.; Han, Z.; et al. Cassava/peanut intercropping improves soil quality via rhizospheric microbes increased available nitrogen contents. BMC Biotechnol. 2020, 20, 13. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, J.; Bai, Z.; Wu, S.; Li, X.; Wang, N.; Du, X.; Fan, H.; Zhuang, G.; Bohu, T.; et al. Unraveling Mechanisms and Impact of Microbial Recruitment on Oilseed Rape (Brassica napus L.) and the Rhizosphere Mediated by Plant Growth-Promoting Rhizobacteria. Microorganisms 2021, 9, 161. [Google Scholar] [CrossRef]
- Satjarak, A.; E Graham, L.; Trest, M.T.; Zedler, J.; Knack, J.J.; Arancibia-Avila, P. Nitrogen fixation and other biogeochemically important features of Atacama Desert giant horsetail plant microbiomes inferred from metagenomic contig analysis. Ann. Bot. 2022, 130, 65–75. [Google Scholar] [CrossRef]
- Sayed, A.M.; Abdel-Wahab, N.M.; Hassan, H.M.; Abdelmohsen, U.R. Saccharopolyspora: An underexplored source for bioactive natural products. J. Appl. Microbiol. 2019, 128, 314–329. [Google Scholar] [CrossRef]
- Rattanapolsan, L.; Nakbanpote, W.; Sangdee, A. Zinc- and cadmium-tolerant endophytic bacteria from Murdannia spectabilis (Kurz) Faden. studied for plant growth-promoting properties, in vitro inoculation, and antagonism. Arch. Microbiol. 2020, 203, 1131–1148. [Google Scholar] [CrossRef]
- Vaishnav, A.; Singh, J.; Singh, P.; Rajput, R.S.; Singh, H.B.; Sarma, B.K. Sphingobacterium sp. BHU-AV3 Induces Salt Tolerance in Tomato by Enhancing Antioxidant Activities and Energy Metabolism. Front. Microbiol. 2020, 11, 443. [Google Scholar] [CrossRef]
- Boss, B.L.; Wanees, A.E.; Zaslow, S.J.; Normile, T.G.; Izquierdo, J.A. Comparative genomics of the plant-growth promoting bacterium Sphingobium sp. strain AEW4 isolated from the rhizosphere of the beachgrass Ammophila breviligulata. BMC Genom. 2022, 23, 508. [Google Scholar] [CrossRef]
- Asaf, S.; Numan, M.; Khan, A.L.; Al-Harrasi, A. Sphingomonas: From diversity and genomics to functional role in environmental remediation and plant growth. Crit. Rev. Biotechnol. 2020, 40, 138–152. [Google Scholar] [CrossRef]
- Feng, L.; Xu, N.; Qu, Q.; Zhang, Z.; Ke, M.; Lu, T.; Qian, H. Synergetic toxicity of silver nanoparticle and glyphosate on wheat (Triticum aestivum L.). Sci. Total. Environ. 2021, 797, 149200. [Google Scholar] [CrossRef]
- An, S.-Q.; Berg, G. Stenotrophomonas maltophilia. Trends Microbiol. 2018, 26, 637–638. [Google Scholar] [CrossRef]
- Aeron, A.; Dubey, R.C.; Maheshwari, D.K. Characterization of a plant-growth-promoting non-nodulating endophytic bacterium (Stenotrophomonas maltophilia) from the root nodules of Mucuna utilis var. capitata L. (Safed Kaunch). Can. J. Microbiol. 2020, 66, 670–677. [Google Scholar] [CrossRef]
- Visioli, G.; Sanangelantoni, A.M.; Vamerali, T.; Cortivo, C.D.; Blandino, M. 16S rDNA Profiling to Reveal the Influence of Seed-Applied Biostimulants on the Rhizosphere of Young Maize Plants. Molecules 2018, 23, 1461. [Google Scholar] [CrossRef]
- Natsagdorj, O.; Sakamoto, H.; Santiago, D.M.O.; Santiago, C.D.; Orikasa, Y.; Okazaki, K.; Ikeda, S.; Ohwada, T. Variovorax sp. Has an Optimum Cell Density to Fully Function as a Plant Growth Promoter. Microorganisms 2019, 7, 82. [Google Scholar] [CrossRef]
- Rameshkumar, N.; Lang, E.; Tanaka, N. Description of Vogesella oryzae sp. nov., isolated from the rhizosphere of saline tolerant pokkali rice. Syst. Appl. Microbiol. 2015, 39, 20–24. [Google Scholar] [CrossRef]
- Reboleira, A.S.; Bodawatta, K.H.; Ravn, N.M.R.; Lauritzen, S.-E.; Skoglund, R.; Poulsen, M.; Michelsen, A.; Jønsson, K.A. Nutrient-limited subarctic caves harbour more diverse and complex bacterial communities than their surface soil. Environ. Microbiome 2022, 17, 41. [Google Scholar] [CrossRef]
- Miller, J.T.; Jolley-Rogers, G.; Mishler, B.D.; Thornhill, A.H. Phylogenetic diversity is a better measure of biodiversity than taxon counting. J. Syst. Evol. 2018, 56, 663–667. [Google Scholar] [CrossRef]
- Wang, C.; Liu, D.; Bai, E. Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition. Soil Biol. Biochem. 2018, 120, 126–133. [Google Scholar] [CrossRef]
- Colin, Y.; Turpault, M.-P.; Fauchery, L.; Buée, M.; Uroz, S. Forest plant cover and mineral type determine the diversity and composition of mineral-colonizing fungal communities. Eur. J. Soil Biol. 2021, 105, 103334. [Google Scholar] [CrossRef]
- Gorbushina, A. Microcolonial fungi: Survival potential of terrestrial vegetative structures. Astrobiology 2003, 3, 543–554. [Google Scholar] [CrossRef]
- Li, T.; Liu, T.; Zheng, C.; Kang, C.; Yang, Z.; Yao, X.; Song, F.; Zhang, R.; Wang, X.; Xu, N.; et al. Changes in soil bacterial community structure as a result of incorporation of Brassica plants compared with continuous planting eggplant and chemical disinfection in greenhouses. PLoS ONE 2017, 12, e0173923. [Google Scholar] [CrossRef]
- Huang, Y.; Zheng, L.; Huang, Y.; Jia, Z.; Song, S.; Li, Z. Effects of different application methods of Bacillus subtilis agent on soil microbial diversity and growth of muskmelon. Chin. J. Biotechnol. 2020, 36, 2644–2656. [Google Scholar] [CrossRef]
- Orellana, L.H.; Ben Francis, T.; Ferraro, M.; Hehemann, J.-H.; Fuchs, B.M.; I Amann, R. Verrucomicrobiota are specialist consumers of sulfated methyl pentoses during diatom blooms. ISME J. 2022, 16, 630–641. [Google Scholar] [CrossRef]
- Verastegui, Y.; Cheng, J.; Engel, K.; Kolczynski, D.; Mortimer, S.; Lavigne, J.; Montalibet, J.; Romantsov, T.; Hall, M.; McConkey, B.J.; et al. Multisubstrate isotope labeling and metagenomic analysis of active soil bacterial communities. mBio 2014, 5, e01157-14. [Google Scholar] [CrossRef]
- Kalyuzhnaya, M.G.; Lapidus, A.; Ivanova, N.; Copeland, A.C.; McHardy, A.C.; Szeto, E.; Salamov, A.; Grigoriev, I.V.; Suciu, D.; Levine, S.R.; et al. High-resolution metagenomics targets specific functional types in complex microbial communities. Nat. Biotechnol. 2008, 26, 1029–1034. [Google Scholar] [CrossRef]
- Doronina, N.; Kaparullina, E.; Trotsenko, Y. The Family Methylophilaceae. In The Prokaryotes: Alphaproteobacteria and Betaproteobacteria; Rosenberg, E., Edward, F.D., Stephen, L., Eds.; Springer: Berlin/Heidelberg, Germay, 2014; pp. 869–880. [Google Scholar] [CrossRef]
- Kalyuhznaya, M.G.; Martens-Habbena, W.; Wang, T.; Hackett, M.; Stolyar, S.M.; Stahl, D.A.; Lidstrom, M.E.; Chistoserdova, L. Methylophilaceae link methanol oxidation to denitrification in freshwater lake sediment as suggested by stable isotope probing and pure culture analysis. Environ. Microbiol. Rep. 2009, 1, 385–392. [Google Scholar] [CrossRef]
- Eyice, Ö.; Namura, M.; Chen, Y.; Mead, A.; Samavedam, S.; Schäfer, H. SIP metagenomics identifies uncultivated Methylophilaceae as dimethylsulphide degrading bacteria in soil and lake sediment. ISME J. 2015, 9, 2336–2348. [Google Scholar] [CrossRef]
- Bending, G.D.; Lincoln, S.D. Characterisation of volatile sulphur-containing compounds produced during decomposition of Brassica juncea tissues in soil. Soil Biol. Biochem. 1999, 31, 695–703. [Google Scholar] [CrossRef]
- Brooke, J.S. Advances in the Microbiology of Stenotrophomonas maltophilia. Clin. Microbiol. Rev. 2021, 34, e00030-19. [Google Scholar] [CrossRef]
- Ryan, R.P.; Monchy, S.; Cardinale, M.; Taghavi, S.; Crossman, L.; Avison, M.B.; Berg, G.; van der Lelie, D.; Dow, J.M. The versatility and adaptation of bacteria from the genus Stenotrophomonas. Nat. Rev. Microbiol. 2009, 7, 514–525. [Google Scholar] [CrossRef]
- Kolton, M.; Erlacher, A.; Berg, G.; Cytryn, E. The Flavobacterium genus in the plant holobiont: Ecological, physiological, and applicative insights. In Microbial Models: From Environmental to Industrial Sustainability; Springer: Singapore, 2016; pp. 189–207. [Google Scholar] [CrossRef]
- Perepelov, A.V.; Shashkov, A.S.; Kim, D.; Potekhina, N.V.; Dmitrenok, A.S.; Senchenkova, S.N.; Dorofeeva, L.V.; Evtushenko, L.I.; Tul’Skaya, E.M. A highly branched novel galactofuranan in the cell wall of Clavibacter tesselarius VKM Ac-1406. Carbohydr. Res. 2023, 529, 108823. [Google Scholar] [CrossRef]
- Elhalag, K.M.; Nasr-Eldin, M.A.; Huang, Q.; Rabab, A.-E.M.; Ahmad, A.A. Lytic phages isolated from Egypt for biocontrol of potato soft rot caused by Pectobacterium carotovorum. Biol. Control 2024, 189, 105444. [Google Scholar] [CrossRef]
- Toth, I.K. Microbe Profile: Pectobacterium atrosepticum: An enemy at the door. Microbiology 2022, 168, 001221. [Google Scholar] [CrossRef]
- Gorshkov, V.; Parfirova, O. Host plant physiological transformation and microbial population heterogeneity as important determinants of the Soft Rot Pectobacteriaceae-plant interactions. Semin. Cell Dev. Biol. 2023, 148–149, 33–41. [Google Scholar] [CrossRef]
- Domingo, R.; Perez, C.; Klair, D.; Vu, H.; Candelario-Tochiki, A.; Wang, X.; Camson, A.; Uy, J.N.; Salameh, M.; Arizala, D.; et al. Genome-informed loop-mediated isothermal amplification assay for specific detection of Pectobacterium parmentieri in infected potato tissues and soil. Sci. Rep. 2021, 11, 21948. [Google Scholar] [CrossRef]
- Ge, T.; Ekbataniamiri, F.; Johnson, S.B.; Larkin, R.P.; Hao, J. Interaction between Dickeya dianthicola and Pectobacterium parmentieri in Potato Infection under Field Conditions. Microorganisms 2021, 9, 316. [Google Scholar] [CrossRef]
- Xie, P.; Huang, K.; Deng, A.; Mo, P.; Xiao, F.; Wu, F.; Xiao, D.; Wang, Y. The diversity and abundance of bacterial and fungal communities in the rhizosphere of Cathaya argyrophylla are affected by soil physicochemical properties. Front. Microbiol. 2023, 14, 1111087. [Google Scholar] [CrossRef]
- Nuppunen-Puputti, M.; Kietäväinen, R.; Purkamo, L.; Rajala, P.; Itävaara, M.; Kukkonen, I.; Bomberg, M. Rock Surface Fungi in Deep Continental Biosphere-Exploration of Microbial Community Formation with Subsurface In Situ Biofilm Trap. Microorganisms 2020, 9, 64. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, Y.; Hirano, S.-I.; Nagaoka, T.; Amano, Y. Genetic survey of indigenous microbial eukaryotic communities, mainly fungi, in sedimentary rock matrices of deep terrestrial subsurface. Ecol. Genet. Genom. 2019, 12, 100042. [Google Scholar] [CrossRef]
- Manici, L.M.; Caputo, F.; De Sabata, D.; Fornasier, F. The enzyme patterns of Ascomycota and Basidiomycota fungi reveal their different functions in soil. Appl. Soil Ecol. 2024, 196, 105323. [Google Scholar] [CrossRef]
- Damm, U.; Crous, P.W.; Fourie, P.H. A fissitunicate ascus mechanism in the Calosphaeriaceae, and novel species of Jattaea and Calosphaeria on Prunus wood. Persoonia 2008, 20, 39–52. [Google Scholar] [CrossRef]
- Barr, M.E. Notes on the Calosphaeriales. Mycologia 1985, 77, 549–565. [Google Scholar] [CrossRef]
- Mostert, L.; Groenewald, J.Z.; Summerbell, R.C.; Gams, W.; Crous, P.W. Taxonomy and pathology of Togninia (Diaporthales) and its Phaeoacremonium anamorphs. Stud. Mycol. 2006, 54, 1–113. [Google Scholar] [CrossRef]
- Baker, S.E.; Bennett, J.W. An overview of the genus Aspergillus. In The Aspergilli: Genomics, Medical Aspects, Biotechnology, and Research Methods; CRC Press: Boca Raton, FL, USA, 2007; pp. 3–13. [Google Scholar] [CrossRef]
- Krijgsheld, P.; Bleichrodt, R.; van Veluw, G.; Wang, F.; Müller, W.; Dijksterhuis, J.; Wösten, H. Development in aspergillus. Stud. Mycol. 2013, 74, 1–29. [Google Scholar] [CrossRef]
- Akhtyamova, N.; Sattarova, R.K. Endophytic yeast Rhodotorula rubra strain TG-1: Antagonistic and plant protection activities. Biochem. Physiol. 2013, 2, 104. [Google Scholar] [CrossRef]
- Mondo, S.J.; Jiménez, D.J.; Hector, R.E.; Lipzen, A.; Yan, M.; LaButti, K.; Barry, K.; van Elsas, J.D.; Grigoriev, I.V.; Nichols, N.N. Genome expansion by allopolyploidization in the fungal strain Coniochaeta 2T2. 1 and its exceptional lignocellulolytic machinery. Biotechnol. Biofuels 2019, 12, 1–18. [Google Scholar] [CrossRef]
- Jiménez, D.J.; Wang, Y.; de Mares, M.C.; Cortes-Tolalpa, L.; A Mertens, J.; E Hector, R.; Lin, J.; Johnson, J.; Lipzen, A.; Barry, K.; et al. Defining the eco-enzymological role of the fungal strain Coniochaeta sp. 2T2.1 in a tripartite lignocellulolytic microbial consortium. FEMS Microbiol. Ecol. 2020, 96, fiz186. [Google Scholar] [CrossRef]
- Firrincieli, A.; Otillar, R.; Salamov, A.; Schmutz, J.; Khan, Z.; Redman, R.S.; Fleck, N.D.; Lindquist, E.; Grigoriev, I.V.; Doty, S.L. Genome sequence of the plant growth promoting endophytic yeast Rhodotorula graminis WP1. Front. Microbiol. 2015, 6, 978. [Google Scholar] [CrossRef] [PubMed]
- Rosa, L.H.; Queiroz, S.C.N.; Moraes, R.M.; Wang, X.; Techen, N.; Pan, Z.; Cantrell, C.L.; Wedge, D.E. Coniochaeta ligniaria: Antifungal activity of the cryptic endophytic fungus associated with autotrophic tissue cultures of the medicinal plant Smallanthus sonchifolius (Asteraceae). Symbiosis 2013, 60, 133–142. [Google Scholar] [CrossRef]
- Bensch, K.; Groenewald, J.; Meijer, M.; Dijksterhuis, J.; Jurjević, Ž.; Andersen, B.; Houbraken, J.; Crous, P.; Samson, R. Cladosporium species in indoor environments. Stud. Mycol. 2018, 89, 177–301. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi; CBS Laboratory Manual Series; CBSKNAW Fungal Diversity Centre: Utrecht, The Netherlands, 2010; ISBN 978-90-70315-82-3. [Google Scholar]
- Ozimek, E.; Hanaka, A. Mortierella Species as the Plant Growth-Promoting Fungi Present in the Agricultural Soils. Agriculture 2021, 11, 7. [Google Scholar] [CrossRef]
- Lysøe, E.; Harris, L.J.; Walkowiak, S.; Subramaniam, R.; Divon, H.H.; Riiser, E.S.; Llorens, C.; Gabaldón, T.; Kistler, H.C.; Jonkers, W.; et al. The genome of the generalist plant pathogen Fusarium avenaceum is enriched with genes involved in redox, signaling and secondary metabolism. PLoS ONE 2014, 9, e112703. [Google Scholar] [CrossRef]
- Ma, L.-J.; Geiser, D.M.; Proctor, R.H.; Rooney, A.P.; O’Donnell, K.; Trail, F.; Gardiner, D.M.; Manners, J.M.; Kazan, K. Fusarium pathogenomics. Annu. Rev. Microbiol. 2013, 67, 399–416. [Google Scholar] [CrossRef]
- Zhang, T.; Jia, R.L.; Yu, L.Y. Diversity and distribution of soil fungal communities associated with biological soil crusts in the southeastern Tengger Desert (China) as revealed by 454 pyrosequencing. Fungal Ecol. 2016, 23, 156–163. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Culturable mycobiota from Karst caves in China, with descriptions of 20 new species. Persoonia 2017, 39, 1–31. [Google Scholar] [CrossRef]
- Ruisi, S.; Barreca, D.; Selbmann, L.; Zucconi, L.; Onofri, S. Fungi in Antarctica. Rev. Environ. Sci. Bio/Technol. 2007, 6, 127–141. [Google Scholar] [CrossRef]
- Connell, L.; Staudigel, H. Fungal diversity in a dark oligotrophic volcanic ecosystem (DOVE) on Mount Erebus, Antarctica. Biology 2013, 2, 798–809. [Google Scholar] [CrossRef]
- Zalar, P.; Gunde-Cimerman, N. Cold-adapted yeasts in Arctic habitats. In Cold-Adapted Yeasts: Biodiversity, Adaptation Strategies and Biotechnological Significance; Springer: Berlin/Heidelberg, Germany, 2014; pp. 49–74. [Google Scholar] [CrossRef]
- Li, W.; Long, Y.; Mo, F.; Shu, R.; Yin, X.; Wu, X.; Zhang, R.; Zhang, Z.; He, L.; Chen, T.; et al. Antifungal activity and biocontrol mechanism of Fusicolla violacea J-1 against soft rot in kiwifruit caused by Alternaria alternata. J. Fungi 2021, 7, 937. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Lopez, N.; Cano-Lira, J.F.; Guarro, J.; Sutton, D.A.; Wiederhold, N.; Crous, P.W.; Stchigel, A.M. Coelomycetous Dothideomycetes with emphasis on the families Cucurbitariaceae and Didymellaceae. Stud. Mycol. 2018, 90, 1–69. [Google Scholar] [CrossRef] [PubMed]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Van Kan, J.A.L. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Choiseul, J.W.; Carnegie, S.F. First Report of Botryotinia fuckeliana Causing Soft Rots in Potato in Scotland. Plant Dis. 2000, 84, 806. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, X.; Solairaj, D.; Lin, R.; Wang, K.; Zhang, H. TMT-Based Proteomic Analysis of Hannaella sinensis-Induced Apple Resistance-Related Proteins. Foods 2023, 12, 2637. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, E.L.M.; Ventorim, R.Z.; Ferreira, M.A.d.M.; da Silveira, W.B. Papiliotrema laurentii: General features and biotechnological applications. Appl. Microbiol. Biotechnol. 2022, 106, 6963–6976. [Google Scholar] [CrossRef] [PubMed]
- Philippot, L.; Piutti, S.; Martin-Laurent, F.; Hallet, S.; Germon, J.C. Molecular analysis of the nitrate-reducing community from unplanted and maize-planted soils. Appl. Environ. Microbiol. 2002, 68, 6121–6128. [Google Scholar] [CrossRef]
- Prade, K.; Trolldenier, G. Effect of wheat roots on denitrification at varying soil air-filled porosity and organic-carbon content. Biol Fertil. Soils 1988, 7, 1–6. [Google Scholar] [CrossRef]
- Warscheid, T.; Braams, J. Biodeterioration of stone: A review. Int. Biodeterior. Biodegrad. 2000, 46, 343–368. [Google Scholar] [CrossRef]
- Krumbein, W.E.; Pochon, J. Ecologie bacterienne des pierres alterees des monuments. Ann. Inst. Pasteur. 1964, 107, 724–732. [Google Scholar]
- Yuan, J.; Zhao, J.; Wen, T.; Zhao, M.; Li, R.; Goossens, P.; Huang, Q.; Bai, Y.; Vivanco, J.M.; Kowalchuk, G.A.; et al. Root exudates drive the soil-borne legacy of aboveground pathogen infection. Microbiome 2018, 6, 156. [Google Scholar] [CrossRef]
- Wang, Y.; Gong, J.; Li, J.; Xin, Y.; Hao, Z.; Chen, C.; Li, H.; Wang, B.; Ding, M.; Li, W.; et al. Insights into bacterial diversity in compost: Core microbiome and prevalence of potential pathogenic bacteria. Sci. Total Environ. 2020, 718, 137304. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Pan, Y.; Xiang, L.; Zhu, Z.; Fu, W.; Hao, G.; Geng, Z.; Chen, S.; Li, Y.; Han, D. Assembly of rhizosphere microbial communities in Artemisia annua: Recruitment of plant growth-promoting microorganisms and inter-kingdom interactions between bacteria and fungi. Plant Soil. 2022, 470, 127–139. [Google Scholar] [CrossRef]
- Ballhausen, M.B.; de Boer, W. The sapro-rhizosphere: Carbon flow from saprotrophic fungi into fungus-feeding bacteria. Soil Biol. Biochem. 2016, 102, 14–17. [Google Scholar] [CrossRef]
- Paterson, E.; Gebbing, T.; Abel, C.; Sim, A.; Telfer, G. Rhizodeposition shapes rhizosphere microbial community structure in organic soil. New Phytol. 2007, 173, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.H.; Anslan, S.; Bahram, M.; Wurzbacher, C.; Baldrian, P.; Tedersoo, L. Mycobiome diversity: High-throughput sequencing and identification of fungi. Nat. Rev. Microbiol. 2019, 17, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, Y.; Ren, X.; Chen, B.; Zhang, Y.; Shen, C.; Wang, F.; Wu, D. Long-Term Greenhouse Cucumber Production Alters Soil Bacterial Community Structure. J. Soil Sci. Plant Nutr. 2020, 20, 306–321. [Google Scholar] [CrossRef]
- Durán, P.; Thiergart, T.; Garrido-Oter, R.; Agler, M.; Kemen, E.; Schulze-Lefert, P.; Hacquard, S. Microbial Interkingdom Interactions in Roots Promote Arabidopsis Survival. Cell 2018, 175, 973–983.e14. [Google Scholar] [CrossRef]
- Geng, Y.; Pan, S.; Zhang, L.; Qiu, J.; He, K.; Gao, H.; Li, Z.; Tian, D. Phosphorus biogeochemistry regulated by carbonates in soil. Environ. Res. 2022, 214 Pt 2, 113894. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, K.; Ma, Y.; Bao, S.; Li, W.; Wang, Y.; Sun, C.; Lu, X.; Ran, J. Exploring the Impact of Coconut Peat and Vermiculite on the Rhizosphere Microbiome of Pre-Basic Seed Potatoes under Soilless Cultivation Conditions. Microorganisms 2024, 12, 584. https://doi.org/10.3390/microorganisms12030584
Yan K, Ma Y, Bao S, Li W, Wang Y, Sun C, Lu X, Ran J. Exploring the Impact of Coconut Peat and Vermiculite on the Rhizosphere Microbiome of Pre-Basic Seed Potatoes under Soilless Cultivation Conditions. Microorganisms. 2024; 12(3):584. https://doi.org/10.3390/microorganisms12030584
Chicago/Turabian StyleYan, Kan, Yanni Ma, Songming Bao, Wandi Li, Yunjiao Wang, Chao Sun, Xin Lu, and Juan Ran. 2024. "Exploring the Impact of Coconut Peat and Vermiculite on the Rhizosphere Microbiome of Pre-Basic Seed Potatoes under Soilless Cultivation Conditions" Microorganisms 12, no. 3: 584. https://doi.org/10.3390/microorganisms12030584
APA StyleYan, K., Ma, Y., Bao, S., Li, W., Wang, Y., Sun, C., Lu, X., & Ran, J. (2024). Exploring the Impact of Coconut Peat and Vermiculite on the Rhizosphere Microbiome of Pre-Basic Seed Potatoes under Soilless Cultivation Conditions. Microorganisms, 12(3), 584. https://doi.org/10.3390/microorganisms12030584