
Comparative Study of Mycobacterium bovis and Mycobacterium avium subsp. paratuberculosis In Vitro Infection in Bovine Bone Marrow Derived Macrophages: Preliminary Results
 ,
,  ,
,  , ,
, ,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of TB and Paratuberculosis Officially Free Farms and Selection of the Animals for the Experiment
2.2. Bone Marrow Collection and Macrophagic Culture
2.3. In-Vitro Infection with Field Strains of M. bovis and MAP
2.4. qRT-PCR
2.5. Cytokines Measurement
2.6. Statistical Analysis
3. Results
3.1. qRT-PCR
3.2. Time Course RNA Studies for IL-1β and IL-6
3.2.1. IL-1β
3.2.2. IL-6
3.3. Plasma Cytokines Measurement
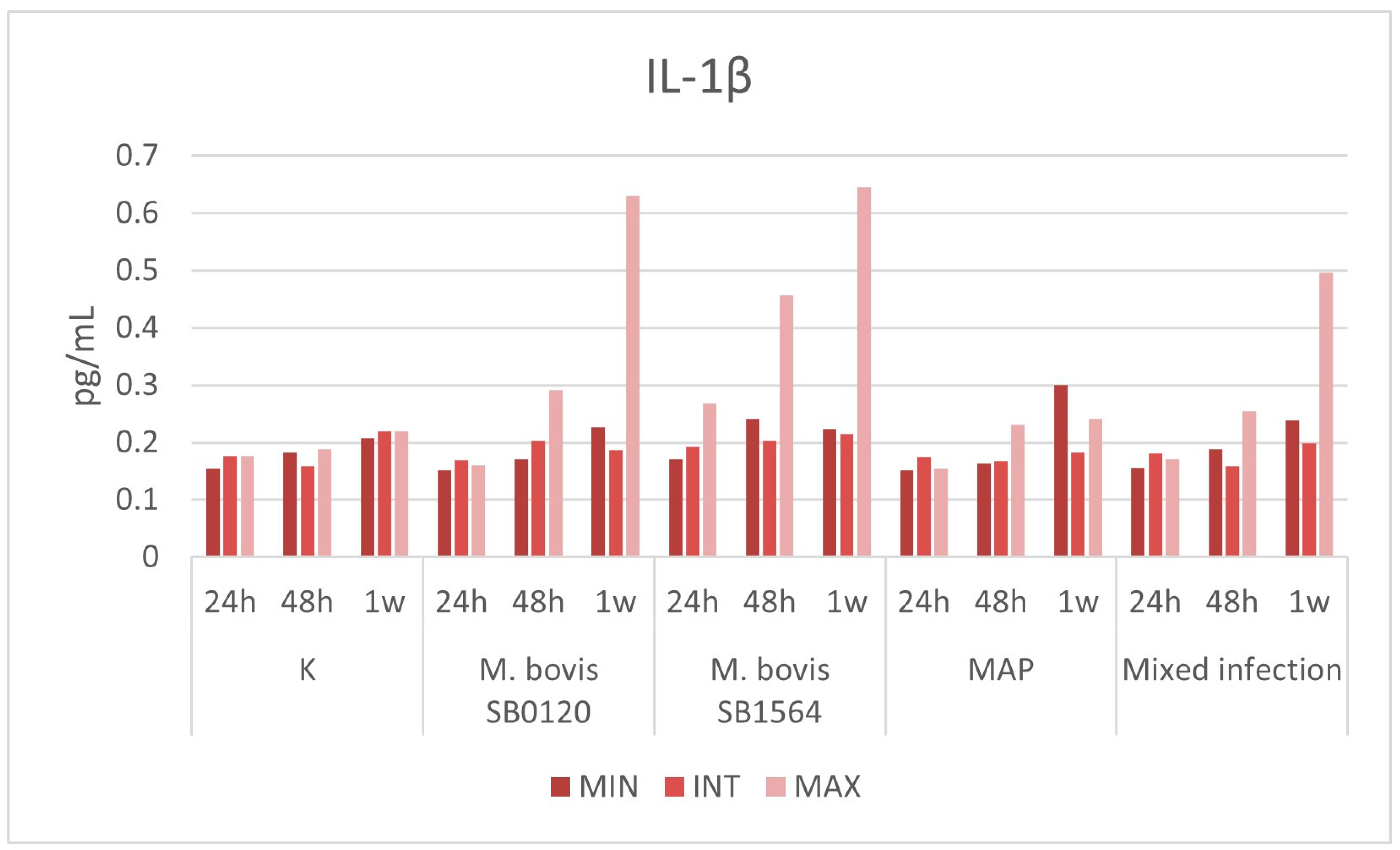
3.3.1. IL-1β
3.3.2. IL-6
4. Discussion
4.1. Activation of the Immune Response in Relation to the Pathogenicity of Mycobacteria: The Modulating Effect
4.2. IL-6
4.3. IL-1β
4.4. IL-2
4.5. TNFα
4.6. IL-10
4.7. IL-12
4.8. IFN-γ
5. Conclusions
- -
- The variability of the macrophage response not only linked to different bacterial species M. bovis/M. paratuberculosis (interspecific) but also intraspecific (two different spoligotypes) which could also be correlated to factors such as strain pathogenicity and virulence. In particular, the comparison in gene expression and cytokines measurements among spoligotypes, revealed a stronger immune stimulation given by SB1564 when compared with both SB0841 and SB0120, respectively. The differences found corroborated the hypothesis that immune response to SB1564 might be more effective, thus partly explaining the reason why this spoligotype is less widespread compared with others.
- -
- The variability of the immune response is associated with the bacterial concentration especially for Mycobacterium bovis. Exposure to high levels of bacterial concentration may enhance the immune response and facilitate the identification of the infection through the diagnostic tests whereas low levels of infection may not be detected by the official tests and persist within the infected area for a longer time allowing the spreading of the mycobacteria. Some M. bovis strains appear to produce a reduced/delayed inflammatory reaction similar to what observed with MAP potentially showing the same ability to “camouflage” within the host cell.
- -
- The identification of IL-6 as an early marker of bovine tuberculosis in the bovine species. The different production noticed in the MAP infection may suggest that this bacterium stimulates the host cell to induce an inflammatory pattern only after a longer period of incubation so even one week was not sufficient to induce a clear cytokine production. This hypothesis is supported by the detection of the transcript by qRT-PCR analysis. Another interesting point is related to the pathogenic aspect. Its massive production, as it can happen in chronic inflammatory processes, induces an imbalance and an unfavourable outcome (e.g., autoimmune diseases such as multiple sclerosis and Chron’s disease). This suggests how mycobacteria can influence and determine the onset of these diseases by stimulating this type of response. It also stimulates the production of B-lymphocytes notoriously ineffective against the tuberculous infections.
- -
- The possible phenomenon of inhibition or modulatory effect observed and induced by the copresence of the two microbial species (MAP and M. bovis) may lead to important implications in the application of the eradication plans. It could be considered as evidence supporting the hypothesis that the coinfection in the same animal interferes with the response to the official diagnostic tests and therefore with the outcome of the test, contributing to the maintenance of M. bovis infection in the farms.
- -
- The presence of a primary transcript attributable to IL-2 in macrophages in both infections.
- -
- The lack of correlation between gene expression and cytokine production (ELISA test). It can be partly explained by the macrophage cell and its post-transcriptional modifications. In fact, there is no direct correlation between the RNA produced and protein synthesis. Possible explanations are related to species characteristics, the complex mycobacterium/cell interaction that influences the production and/or inhibition of some cytokines and other aspects such as survival mechanisms and/or proliferation within the host cell, or finally to possible regulatory and inhibitory feedback mechanisms [54].
- -
- The inhibitory action on the TNFα transcript induced by both mycobacteria regardless of the strain can potentially be interpreted as a survival mechanism.
- -
- The inhibition of the gene expression of IL-10 during the early stages of the disease whilst notoriously increased in the most advanced stages of tuberculous infection may be linked to the need of the mycobacteria to recruit more macrophages as host cells where they can hide and multiply.
- -
- Inhibition of IL-12 production as a possible further attempt made by MAP to delay the activation of the immune response.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Riojas, M.A.; McGough, K.J.; Rider-Riojas, C.J.; Rastogi, N.; Hazbón, M.H. Phylogenomic Analysis of the Species of the Mycobacterium Tuberculosis Complex Demonstrates That Mycobacterium Africanum, Mycobacterium Bovis, Mycobacterium Caprae, Mycobacterium Microti and Mycobacterium Pinnipedii Are Later Heterotypic Synonyms of Mycobacterium Tuberculosis. Int. J. Syst. Evol. Microbiol. 2018, 68, 324–332. [Google Scholar] [CrossRef]
- Sharma, S.K.; Upadhyay, V. Epidemiology, Diagnosis & Treatment of Non-Tuberculous Mycobacterial Diseases. Indian J. Med. Res. 2020, 152, 185–226. [Google Scholar] [CrossRef] [PubMed]
- Rindi, L.; Garzelli, C. Genetic Diversity and Phylogeny of Mycobacterium Avium. Infect. Genet. Evol. 2014, 21, 375–383. [Google Scholar] [CrossRef]
- Harris, N.B.; Barletta, R.G. Mycobacterium Avium Subsp. Paratuberculosis in Veterinary Medicine. Clin. Microbiol. Rev. 2001, 14, 489–512. [Google Scholar] [CrossRef]
- Bates, A.; O’Brien, R.; Liggett, S.; Griffin, F. Control of Mycobacterium Avium Subsp. Paratuberculosis Infection on a New Zealand Pastoral Dairy Farm. BMC Vet. Res. 2019, 15, 266. [Google Scholar] [CrossRef] [PubMed]
- Amato, B.; Di Marco Lo Presti, V.; Gerace, E.; Capucchio, M.T.; Vitale, M.; Zanghì, P.; Pacciarini, M.L.; Marianelli, C.; Boniotti, M.B. Molecular Epidemiology of Mycobacterium Tuberculosis Complex Strains Isolated from Livestock and Wild Animals in Italy Suggests the Need for a Different Eradication Strategy for Bovine Tuberculosis. Transbound. Emerg. Dis. 2018, 65, e416–e424. [Google Scholar] [CrossRef] [PubMed]
- Marianelli, C.; Amato, B.; Boniotti, M.B.; Vitale, M.; Pruiti Ciarello, F.; Pacciarini, M.L.; Di Marco Lo Presti, V. Genotype Diversity and Distribution of Mycobacterium Bovis from Livestock in a Small, High-Risk Area in Northeastern Sicily, Italy. PLoS Negl. Trop. Dis. 2019, 13, e0007546. [Google Scholar] [CrossRef]
- Di Marco, V.; Mazzone, P.; Capucchio, M.T.; Boniotti, M.B.; Aronica, V.; Russo, M.; Fiasconaro, M.; Cifani, N.; Corneli, S.; Biasibetti, E.; et al. Epidemiological Significance of the Domestic Black Pig (Sus Scrofa) in Maintenance of Bovine Tuberculosis in Sicily. J. Clin. Microbiol. 2012, 50, 1209–1218. [Google Scholar] [CrossRef]
- Marianelli, C.; Verrubbi, V.; Pruiti Ciarello, F.; Ippolito, D.; Pacciarini, M.L.; Di Marco Lo Presti, V. Geo-Epidemiology of Animal Tuberculosis and Mycobacterium Bovis Genotypes in Livestock in a Small, High-Incidence Area in Sicily, Italy. Front. Microbiol. 2023, 14, 1107396. [Google Scholar] [CrossRef]
- Sundaramurthy, V.; Pieters, J. Interactions of Pathogenic Mycobacteria with Host Macrophages. Microbes Infect. 2007, 9, 1671–1679. [Google Scholar] [CrossRef]
- Rastogi, N.; David, H.L. Mechanisms of Pathogenicity in Mycobacteria. Biochimie 1988, 70, 1101–1120. [Google Scholar] [CrossRef]
- Momotani, E.; Romona, N.M.; Yoshihara, K.; Momotani, Y.; Hori, M.; Ozaki, H.; Eda, S.; Ikegami, M. Molecular Pathogenesis of Bovine Paratuberculosis and Human Inflammatory Bowel Diseases. Vet. Immunol. Immunopathol. 2012, 148, 55–68. [Google Scholar] [CrossRef]
- Scanu, A.M.; Bull, T.J.; Cannas, S.; Sanderson, J.D.; Sechi, L.A.; Dettori, G.; Zanetti, S.; Hermon-Taylor, J. Mycobacterium Avium Subspecies Paratuberculosis Infection in Cases of Irritable Bowel Syndrome and Comparison with Crohn’s Disease and Johne’s Disease: Common Neural and Immune Pathogenicities. J. Clin. Microbiol. 2007, 45, 3883–3890. [Google Scholar] [CrossRef]
- Cossu, D.; Yokoyama, K.; Nobutaka, N.; Sechi, L.A. From Sardinia to Japan: Update on the Role of MAP in Multiple Sclerosis. Future Microbiol. 2019, 14, 643–646. [Google Scholar] [CrossRef] [PubMed]
- Raffo, E.; Steuer, P.; Monti, G.; Salgado, M. Effect of Mycobacterium Avium Subsp. Paratuberculosis (MAP) Infection on the Diagnostic Accuracy for Mycobacterium Bovis (M. Bovis) Infection under Field Conditions in Cattle Belonging to Low M. Bovis Prevalence Herds. Trop. Anim. Health Prod. 2017, 49, 771–775. [Google Scholar] [CrossRef] [PubMed]
- Marais, B.J.; Buddle, B.M.; de Klerk-Lorist, L.-M.; Nguipdop-Djomo, P.; Quinn, F.; Greenblatt, C. BCG Vaccination for Bovine Tuberculosis; Conclusions from the Jerusalem One Health Workshop. Transbound. Emerg. Dis. 2019, 66, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Serrano, M.; Elguezabal, N.; Sevilla, I.A.; Geijo, M.V.; Molina, E.; Arrazuria, R.; Urkitza, A.; Jones, G.J.; Vordermeier, M.; Garrido, J.M.; et al. Tuberculosis Detection in Paratuberculosis Vaccinated Calves: New Alternatives against Interference. PLoS ONE 2017, 12, e0169735. [Google Scholar] [CrossRef] [PubMed]
- de la Rua-Domenech, R.; Goodchild, A.T.; Vordermeier, H.M.; Hewinson, R.G.; Christiansen, K.H.; Clifton-Hadley, R.S. Ante Mortem Diagnosis of Tuberculosis in Cattle: A Review of the Tuberculin Tests, γ-Interferon Assay and Other Ancillary Diagnostic Techniques. Res. Vet. Sci. 2006, 81, 190–210. [Google Scholar] [CrossRef] [PubMed]
- Seva, J.; Sanes, J.M.; Ramis, G.; Mas, A.; Quereda, J.J.; Villarreal-Ramos, B.; Villar, D.; Pallares, F.J. Evaluation of the Single Cervical Skin Test and Interferon Gamma Responses to Detect Mycobacterium Bovis Infected Cattle in a Herd Co-Infected with Mycobacterium Avium Subsp. Paratuberculosis. Vet. Microbiol. 2014, 171, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, J.; De Juan, L.; Bezos, J.; Romero, B.; Sáez, J.L.; Marqués, S.; Domínguez, C.; Mínguez, O.; Fernández-Mardomingo, B.; Mateos, A.; et al. Effect of Paratuberculosis on the Diagnosis of Bovine Tuberculosis in a Cattle Herd with a Mixed Infection Using Interferon-Gamma Detection Assay. Vet. Microbiol. 2009, 135, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Blanco, F.C.; Schierloh, P.; Bianco, M.V.; Caimi, K.; Meikle, V.; Alito, A.E.; Cataldi, A.A.; Sasiain, M.D.C.; Bigi, F. Study of the Immunological Profile towards Mycobacterium Bovis Antigens in Naturally Infected Cattle. Microbiol. Immunol. 2009, 53, 460–467. [Google Scholar] [CrossRef]
- Klepp, L.I.; Eirin, M.E.; Garbaccio, S.; Soria, M.; Bigi, F.; Blanco, F.C. Identification of Bovine Tuberculosis Biomarkers to Detect Tuberculin Skin Test and IFNγ Release Assay False Negative Cattle. Res. Vet. Sci. 2019, 122, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.; De Silva, K.; Plain, K.M.; Purdie, A.C.; Blair, T.A.; Duggin, I.G.; Britton, W.J.; Oehlers, S.H. Mycobacterial Infection-Induced miR-206 Inhibits Protective Neutrophil Recruitment via the CXCL12/CXCR4 Signalling Axis. PLoS Pathog. 2021, 17, e1009186. [Google Scholar] [CrossRef]
- Rhodes, S.G.; Palmer, N.; Graham, S.P.; Bianco, A.E.; Hewinson, R.G.; Vordermeier, H.M. Distinct Response Kinetics of Gamma Interferon and Interleukin-4 in Bovine Tuberculosis. Infect. Immun. 2000, 68, 5393–5400. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Singh, P.P.; Goyal, A. Interleukin-6: A Potent Biomarker of Mycobacterial Infection. SpringerPlus 2013, 2, 686. [Google Scholar] [CrossRef]
- Coad, M.; Clifford, D.; Rhodes, S.G.; Hewinson, R.G.; Vordermeier, H.M.; Whelan, A.O. Repeat Tuberculin Skin Testing Leads to Desensitisation in Naturally Infected Tuberculous Cattle Which Is Associated with Elevated Interleukin-10 and Decreased Interleukin-1 Beta Responses. Vet. Res. 2010, 41, 14. [Google Scholar] [CrossRef]
- Stabel, J.R.; Bannantine, J.P.; Hostetter, J.M. Comparison of Sheep, Goats, and Calves as Infection Models for Mycobacterium Avium Subsp. Paratuberculosis. Vet. Immunol. Immunopathol. 2020, 225, 110060. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, A.E.; Lambert, N.; Duan, X.; Xie, J. Interleukin-10 Family and Tuberculosis: An Old Story Renewed. Int. J. Biol. Sci. 2016, 12, 710–717. [Google Scholar] [CrossRef]
- Trouplin, V.; Boucherit, N.; Gorvel, L.; Conti, F.; Mottola, G.; Ghigo, E. Bone Marrow-Derived Macrophage Production. J Vis Exp. 2013, 81, e50966. [Google Scholar] [CrossRef]
- Giardina, A.; Alduina, R.; Gottardi, E.; Di Caro, V.; Süssmuth, R.D.; Puglia, A.M. Two Heterologously Expressed Planobispora Rosea Proteins Cooperatively Induce Streptomyces Lividans Thiostrepton Uptake and Storage from the Extracellular Medium. Microb. Cell Factories 2010, 9, 44. [Google Scholar] [CrossRef]
- Giardina, A.; Alduina, R.; Gallo, G.; Monciardini, P.; Sosio, M.; Puglia, A.M. Inorganic phosphate is a trigger factor for Microbispora sp. ATCC-PTA-5024 growth and NAI-107 production. Microb. Cell Fact. 2014, 13, 133. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.L.; Sutherland, I.A.; Sutherland, J. Validation of candidate bovine reference genes for use with real-time PCR. Vet. Immunol. Immunopathol. 2007, 115, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Marino, R.; Capoferri, R.; Panelli, S.; Minozzi, G.; Strozzi, F.; Trevisi, E.; Snel, G.G.M.; Ajmone-Marsan, P.; Williams, J.L. Johne’s Disease in Cattle: An in Vitro Model to Study Early Response to Infection of Mycobacterium Avium Subsp. Paratuberculosis Using RNA-Seq. Mol. Immunol. 2017, 91, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Coussens, P.M.; Colvin, C.J.; Wiersma, K.; Abouzied, A.; Sipkovsky, S. Gene Expression Profiling of Peripheral Blood Mononuclear Cells from Cattle Infected with Mycobacterium Paratuberculosis. Infect. Immun. 2002, 70, 5494–5502. [Google Scholar] [CrossRef] [PubMed]
- Alfonseca-Silva, E.; Hernández-Pando, R.; Gutiérrez-Pabello, J.A. Mycobacterium Bovis-Infected Macrophages from Resistant and Susceptible Cattle Exhibited a Differential pro-Inflammatory Gene Expression Profile Depending on Strain Virulence. Vet. Immunol. Immunopathol. 2016, 176, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, P.C.; Behrmann, I.; Haan, S.; Hermanns, H.M.; Müller-Newen, G.; Schaper, F. Principles of Interleukin (IL)-6-Type Cytokine Signalling and Its Regulation. Biochem. J. 2003, 374, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Kimura, A.; Kishimoto, T. IL-6: Regulator of Treg/Th17 Balance. Eur. J. Immunol. 2010, 40, 1830–1835. [Google Scholar] [CrossRef] [PubMed]
- Kumari, N.; Dwarakanath, B.S.; Das, A.; Bhatt, A.N. Role of Interleukin-6 in Cancer Progression and Therapeutic Resistance. Tumor Biol. 2016, 37, 11553–11572. [Google Scholar] [CrossRef]
- Pedersen, B.K.; Febbraio, M.A. Muscle as an Endocrine Organ: Focus on Muscle-Derived Interleukin-6. Physiol. Rev. 2008, 88, 1379–1406. [Google Scholar] [CrossRef]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The Pro- and Anti-Inflammatory Properties of the Cytokine Interleukin-6. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2011, 1813, 878–888. [Google Scholar] [CrossRef]
- Ghavidel, M.; Mansury, D.; Nourian, K.; Ghazvini, K. The Most Common Spoligotype of Mycobacterium Bovis Isolated in the World and the Recommended Loci for VNTR Typing; A Systematic Review. Microb. Pathog. 2018, 118, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.L.; Czuprynski, C.J. Ex Vivo Induction of TNF-α and IL-6 mRNA in Bovine Whole Blood by Mycobacterium Paratuberculosis and Mycobacterial Cell Wall Components. Microb. Pathog. 1995, 19, 19–29. [Google Scholar] [CrossRef]
- Everman, J.L.; Eckstein, T.M.; Roussey, J.; Coussens, P.; Bannantine, J.P.; Bermudez, L.E. Characterization of the Inflammatory Phenotype of Mycobacterium Avium Subspecies Paratuberculosis Using a Novel Cell Culture Passage Model. Microbiology 2015, 161, 1420–1434. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lopez-Castejon, G.; Brough, D. Understanding the Mechanism of IL-1β Secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Simon, A.; Van De Veerdonk, F.; Kullberg, B.-J.; Van Der Meer, J.W.M.; Joosten, L.A.B. IL-1β Processing in Host Defense: Beyond the Inflammasomes. PLoS Pathog. 2010, 6, e1000661. [Google Scholar] [CrossRef]
- Bourigault, M.; Segueni, N.; Rose, S.; Court, N.; Vacher, R.; Vasseur, V.; Erard, F.; Le Bert, M.; Garcia, I.; Iwakura, Y.; et al. Relative Contribution of IL-1α, IL-1β and TNF to the Host Response to Mycobacterium Tuberculosis and Attenuated M. Bovis BCG. Immun. Inflamm. Dis. 2013, 1, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Capobianco, M.P.; Cassiano, G.C.; Da Cruz Furini, A.A.; Storti De Melo, L.M.; Bonini Domingos, C.R.; Dantas Machado, R.L. Human Interleukin 2 (IL-2) Promotion of Immune Regulation and ClinicalOutcomes: A Review. J. Cytokine Biol. 2016, 1, 14–17. [Google Scholar] [CrossRef]
- Ahmad, S. Pathogenesis, Immunology, and Diagnosis of Latent Mycobacterium Tuberculosis Infection. Clinical and Developmental Immunology 2011, 2011, 814943. [Google Scholar] [CrossRef]
- Gupta, R.; Pandey, M.; Pandey, A.K.; Tiwari, P.K.; Amrathlal, R.S. Novel Genetic Polymorphisms Identified in the Clinical Isolates of Mycobacterium Tuberculosis PE_PGRS33 Gene Modulate Cytokines Expression and Promotes Survival in Macrophages. J. Infect. Public Health 2022, 15, 245–254. [Google Scholar] [CrossRef]
- Ouyang, W.; Rutz, S.; Crellin, N.K.; Valdez, P.A.; Hymowitz, S.G. Regulation and Functions of the IL-10 Family of Cytokines in Inflammation and Disease. Annu. Rev. Immunol. 2011, 29, 71–109. [Google Scholar] [CrossRef]
- Trinchieri, G. Interleukin-12: A Proinflammatory Cytokine with Immunoregulatory Functions That Bridge Innate Resistance and Antigen-Specific Adaptive Immunity. Annu. Rev. Immunol. 1995, 13, 251–276. [Google Scholar] [CrossRef] [PubMed]
- Gessani, S.; Belardelli, F. IFN-γ Expression in Macrophages and Its Possible Biological Significance. Cytokine Growth Factor Rev. 1998, 9, 117–123. [Google Scholar] [CrossRef]
- Arsenault, R.J.; Li, Y.; Bell, K.; Doig, K.; Potter, A.; Griebel, P.J.; Kusalik, A.; Napper, S. Mycobacterium Avium Subsp. Paratuberculosis Inhibits Gamma Interferon-Induced Signaling in Bovine Monocytes: Insights into the Cellular Mechanisms of Johne’s Disease. Infect. Immun. 2012, 80, 3039–3048. [Google Scholar] [CrossRef] [PubMed]
- Vogel, C.; Marcotte, E.M. Insights into the Regulation of Protein Abundance from Proteomic and Transcriptomic Analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | Forward Primer 5′–3′ | Reverse Primer 5′–3′ |
|---|---|---|---|
| IL-1β | NM_174093.1 | CTAGCCCATGTGTGCTGAAG | CCACTTCTCGGTTCATTTCC |
| IL-2 | NM_180997.2 | ACTCCTGCCACAATGTACAAG | TGTTCCCCGTAGAGCTTGAA |
| IL-6 | NM_173923.2 | TAACACCATCAAGGACCTGT | TGCCCAGGAACTACCACAAT |
| IL-10 | M_174088.1 | ACTCTGTTGCCTGGTCTTCC | GACAGGGTGCTCGCATCT |
| IL12p40 | EU276076.1 | TTGCTCTCAGCAGAGAAGGTC | CTGCCCTCCTGACACTCC |
| IFN γ | NM_174086.1 | GGCATGTCAGACAGCACTTG | TGAAGCGCCAGGTATAAGGT |
| TNFα | AF348421.1 | GTGTGAAGCTGGAAGACAACC | CCCTGAAGAGGACCTGTGAG |
| GAPDH | NM_001034034.2 | AGATGGTGAAGGTCGGAGTG | GACGATGTCCACTTTGCCAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amato, B.; Ippolito, D.; Vitale, M.; Alduina, R.; Galluzzo, P.; Gerace, E.; Pruiti Ciarello, F.; Fiasconaro, M.; Cannella, V.; Di Marco Lo Presti, V. Comparative Study of Mycobacterium bovis and Mycobacterium avium subsp. paratuberculosis In Vitro Infection in Bovine Bone Marrow Derived Macrophages: Preliminary Results. Microorganisms 2024, 12, 407. https://doi.org/10.3390/microorganisms12020407
Amato B, Ippolito D, Vitale M, Alduina R, Galluzzo P, Gerace E, Pruiti Ciarello F, Fiasconaro M, Cannella V, Di Marco Lo Presti V. Comparative Study of Mycobacterium bovis and Mycobacterium avium subsp. paratuberculosis In Vitro Infection in Bovine Bone Marrow Derived Macrophages: Preliminary Results. Microorganisms. 2024; 12(2):407. https://doi.org/10.3390/microorganisms12020407
Chicago/Turabian StyleAmato, Benedetta, Dorotea Ippolito, Maria Vitale, Rosa Alduina, Paola Galluzzo, Elisabetta Gerace, Flavia Pruiti Ciarello, Michele Fiasconaro, Vincenza Cannella, and Vincenzo Di Marco Lo Presti. 2024. "Comparative Study of Mycobacterium bovis and Mycobacterium avium subsp. paratuberculosis In Vitro Infection in Bovine Bone Marrow Derived Macrophages: Preliminary Results" Microorganisms 12, no. 2: 407. https://doi.org/10.3390/microorganisms12020407
APA StyleAmato, B., Ippolito, D., Vitale, M., Alduina, R., Galluzzo, P., Gerace, E., Pruiti Ciarello, F., Fiasconaro, M., Cannella, V., & Di Marco Lo Presti, V. (2024). Comparative Study of Mycobacterium bovis and Mycobacterium avium subsp. paratuberculosis In Vitro Infection in Bovine Bone Marrow Derived Macrophages: Preliminary Results. Microorganisms, 12(2), 407. https://doi.org/10.3390/microorganisms12020407