

Visceral Leishmaniasis in Immunocompetent Hosts in Brescia: A Case Series and Analysis of Cytokine Cascade

 , ,
, ,  and
and
Abstract
1. Introduction
1.1. Epidemiology
1.2. Physiopathology
2. Case Series
3. Aim of the Study
4. Materials and Methods
5. Results
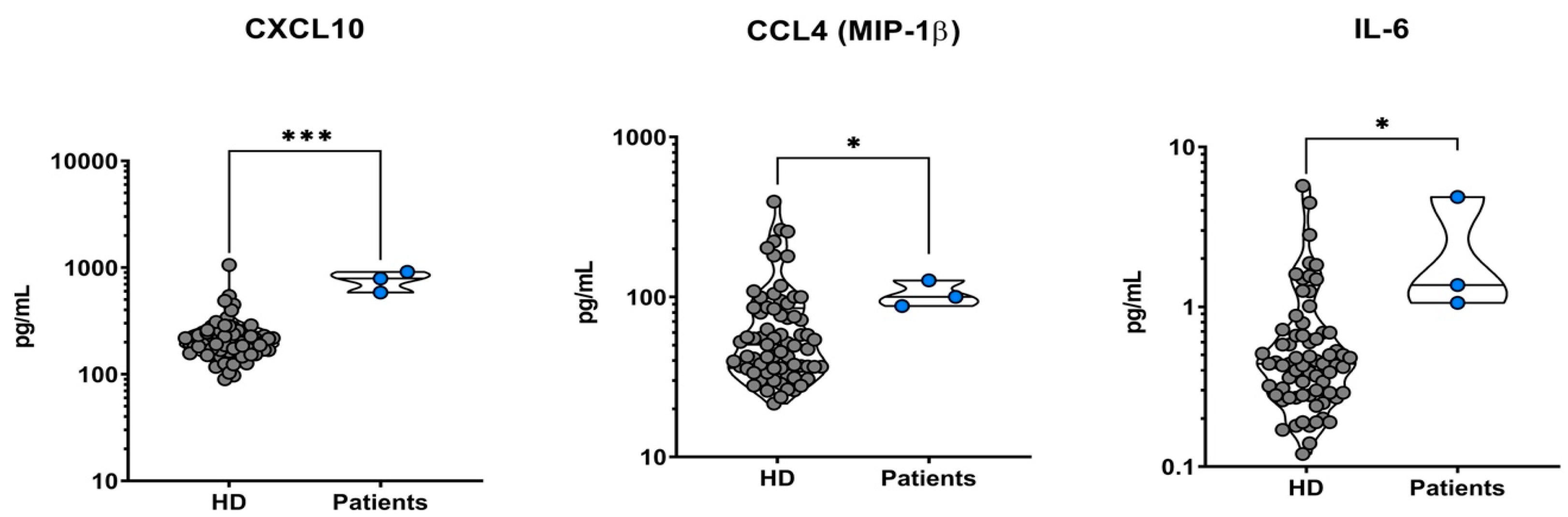
5.1. Analysis of All Patients’ Sera versus Healthy Controls
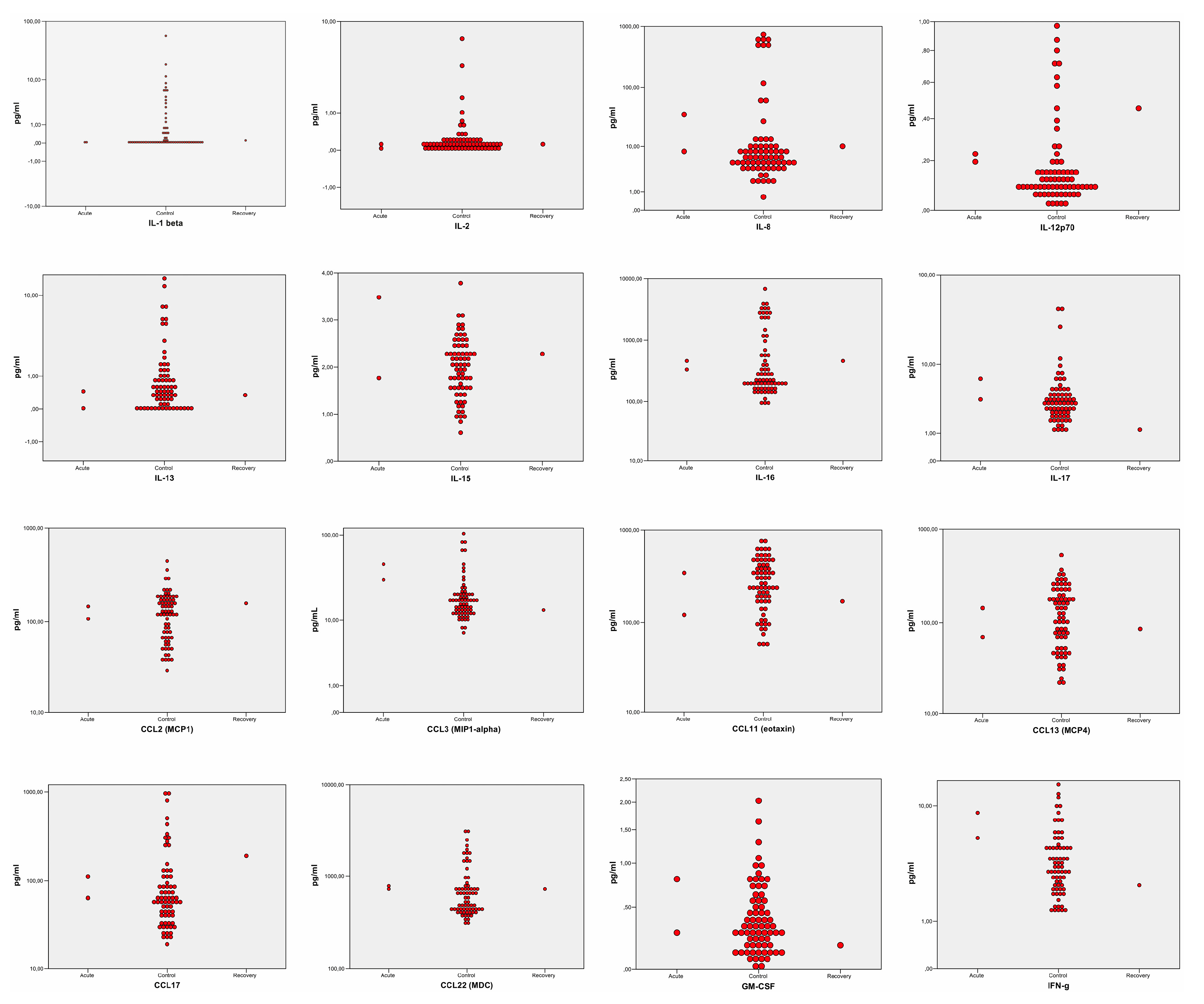
5.2. Sub-Analysis Stratified by Disease Stages: The Recovery Phase
5.3. Sub-Analysis Stratified by Disease Stages: The Acute Phase
5.4. Interferon-γ(IFN-γ): The Missing Elevation
6. Discussion
6.1. COVID-19 Pandemic
6.2. Epidemiology in Brescia
6.3. Cytokine Cascade
6.4. Limitations of the Study
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A


References
- Fichoux, Y.L.E.; Quaranta, O.I.S.; Aufeuvre, J.; Lelievre, A.; Marty, P.; Suffia, I.; Rousseau, D.; Kubar, J. Occurrence of Leishmania Infantum Parasitemia in Asymptomatic Blood Donors Living in an Area of Endemicity in Southern France. J. Clin. Microbiol. 1999, 37, 1953–1957. [Google Scholar] [CrossRef]
- Saporito, L.; Giammanco, G.M.; De Grazia, S.; Colomba, C. Visceral Leishmaniasis: Host-Parasite Interactions and Clinical Presentation in the Immunocompetent and in the Immunocompromised Host. Int. J. Infect. Dis. 2013, 17, e572–e576. [Google Scholar] [CrossRef]
- Cruz, I.; Morales, M.A.; Noguer, I.; Alvar, J. Leishmania in Discarded Syringes from Intravenous Drug Users HMG CoA Reductase Inhibitor (Statin) and Aortic Valve Calcium. Lancet 2002, 359, 1124–1125. [Google Scholar] [CrossRef] [PubMed]
- Patrícia, E.A.F.; Beitune, E.; Queiroz, G.T.; Somensi, R.S.; Morais, N.O.; Elizabeth, M.; Maria, S.; Geraldo, Q. Visceral Leishmaniasis and Pregnancy: Analysis of Cases Reported in a Central-Western Region of Brazil. Arch. Gynecol. Obstet. 2008, 278, 13–16. [Google Scholar] [CrossRef]
- Butenko, A.; Kostygov, A.Y.; Sádlová, J.; Kleschenko, Y.; Bečvář, T.; Podešvová, L.; MacEdo, D.H.; Žihala, D.; Lukeš, J.; Bates, P.A.; et al. Comparative Genomics of Leishmania (Mundinia). BMC Genomics 2019, 20, 726. [Google Scholar] [CrossRef]
- Haouas, N.; Garrab, S.; Gorcii, M.; Khorchani, H.; Chargui, N. Development of a Polymerase Chain Reaction-Restriction Fragment Length Polymorphism Assay for Leishmania Major / Leishmania Killicki / Leishmania Infantum Discrimination from Clinical Samples, Application in a Tunisian Focus. Diagn. Microbiol. Infect. Dis. 2010, 68, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Hailu, A.; Van Baarle, D.; Knol, G.J.; Berhe, N.; Miedema, F.; Kager, P.A. T Cell Subset and Cytokine Profiles in Human Visceral Leishmaniasis during Active and Asymptomatic or Sub-Clinical Infection with Leishmania Donovani. Clin. Immunol. 2005, 117, 182–191. [Google Scholar] [CrossRef]
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, Regional, and National Incidence, Prevalence, and Years Lived with Disability for 354 Diseases and Injuries for 195 Countries and Territories, 1990-2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef]
- Hotez, P.J.; Alvarado, M.; Basáñez, M.G.; Bolliger, I.; Bourne, R.; Boussinesq, M.; Brooker, S.J.; Brown, A.S.; Buckle, G.; Budke, C.M.; et al. The Global Burden of Disease Study 2010: Interpretation and Implications for the Neglected Tropical Diseases. PLoS Negl. Trop. Dis. 2014, 8, e2865. [Google Scholar] [CrossRef]
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; Boer, M. den; Team, the W.H.O.L.C. Leishmaniasis Worldwide and Global Estimates of Its Incidence. PLoS ONE 2012, 7, e35671. [Google Scholar] [CrossRef] [PubMed]
- Elmahallawy, E.K.; Alkhaldi, A.A.M.; Saleh, A.A. Host Immune Response against Leishmaniasis and Parasite Persistence Strategies: A Review and Assessment of Recent Research. Biomed. Pharmacother. 2021, 139, 111671. [Google Scholar] [CrossRef] [PubMed]
- mondiale de la Santé, O.; World Health Organization. WHO TEAM Control of Neglected Tropical Diseases Global Leishmaniasis Suveillance: 2019–2020, a Baseline for the 2030 Roadmap. Wkly. Epidemiol. Rec. 2021, 35, 401–419. [Google Scholar]
- Wamai, R.G.; Kahn, J.; McGloin, J.; Ziaggi, G. Visceral Leishmaniasis: A Global Overview. J. Glob. Health Sci. 2020, 2, e3. [Google Scholar] [CrossRef]
- Van der Auwera, G.; Davidsson, L.; Buffet, P.; Ruf, M.T.; Gramiccia, M.; Varani, S.; Chicharro, C.; Bart, A.; Harms, G.; Chiodini, P.L.; et al. Surveillance of Leishmaniasis Cases from 15 European Centres, 2014 to 2019: A Retrospective Analysis. Eurosurveillance 2022, 27, 2002028. [Google Scholar] [CrossRef] [PubMed]
- Ready, P.D. Leishmaniasis Emergence in Europe. Eurosurveillance 2010, 15, 29–39. [Google Scholar] [CrossRef]
- Bettini, S.; Pampiglione, S.; Maroli, M. Studies on Mediterranean Leishmaniasis: V. a Preliminary Epidemiological Survey of Human Leishmaniasis in Tuscany. Trans. R. Soc. Trop. Med. Hyg. 1977, 71, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Abdalmaula, G.H.; Barbadoro, P.; Marigliano, A.; Illuminati, D.; Di Stanislao, F.; D’Errico, M.M.; Prospero, E. Human Visceral Leishmaniasis: A Picture from Italy. J. Infect. Public Health 2013, 6, 465–472. [Google Scholar] [CrossRef][Green Version]
- Varani, S.; Cagarelli, R.; Melchionda, F.; Attard, L.; Salvadori, C.; Finarelli, A.C.; Gentilomi, G.A.; Tigani, R.; Rangoni, R.; Todeschini, R.; et al. Ongoing Outbreak of Visceral Leishmaniasis in Bologna Province, Italy, November 2012 to May 2013. Eurosurveillance 2013, 18, 20530. [Google Scholar] [CrossRef]
- Ortalli, M.; De Pascali, A.M.; Longo, S.; Pascarelli, N.; Porcellini, A.; Ruggeri, D.; Randi, V.; Procopio, A.; Re, M.C.; Varani, S. Asymptomatic Leishmania Infantum Infection in Blood Donors Living in an Endemic Area, Northeastern Italy. J. Infect. 2020, 80, 116–120. [Google Scholar] [CrossRef]
- Taddei, R.; Bregoli, A.; Galletti, G.; Carra, E.; Fiorentini, L.; Fontana, M.C.; Frasnelli, M.; Musto, C.; Pupillo, G.; Reggiani, A.; et al. Wildlife Hosts of Leishmania Infantum in a Re-Emerging Focus of Human Leishmaniasis, in Emilia-Romagna, Northeast Italy. Pathogens 2022, 11, 1308. [Google Scholar] [CrossRef] [PubMed]
- Abbate, J.M.; Arfuso, F.; Napoli, E.; Gaglio, G.; Giannetto, S.; Latrofa, M.S.; Otranto, D.; Brianti, E. Leishmania Infantum in Wild Animals in Endemic Areas of Southern Italy. Comp. Immunol. Microbiol. Infect. Dis. 2019, 67, 101374. [Google Scholar] [CrossRef]
- Moreno, I.; Álvarez, J.; García, N.; de la Fuente, S.; Martínez, I.; Marino, E.; Toraño, A.; Goyache, J.; Vilas, F.; Domínguez, L.; et al. Detection of Anti-Leishmania Infantum Antibodies in Sylvatic Lagomorphs from an Epidemic Area of Madrid Using the Indirect Immunofluorescence Antibody Test. Vet. Parasitol. 2014, 199, 264–267. [Google Scholar] [CrossRef]
- Gradoni, L.; Ferroglio, E.; Zanet, S.; Mignone, W.; Venco, L.; Bongiorno, G.; Fiorentino, E.; Cassini, R.; Grillini, M.; Simonato, G.; et al. Monitoring and Detection of New Endemic Foci of Canine Leishmaniosis in Northern Continental Italy: An Update from a Study Involving Five Regions (2018–2019). Vet. Parasitol. Reg. Stud. Reports 2022, 27, 100676. [Google Scholar] [CrossRef]
- Peters, N.C.; Egen, J.G.; Secundino, N.; Debrabant, A.; Kamhawi, S.; Lawyer, P.; Fay, M.P.; Germain, R.N.; Sacks, D. In Vivo Imaging Reveals an Essential Role for Neutrophils in Leishmaniasis Transmitted by Sand Flies. NIH Public Access 2009, 321, 970–974. [Google Scholar] [CrossRef]
- Martínez-lópez, M.; Soto, M.; Iborra, S.; Sancho, D. Leishmania Hijacks Myeloid Cells for Immune Escape. Front. Microbiol. 2018, 9, 883. [Google Scholar] [CrossRef]
- Bhor, R.; Rafati, S.; Pai, K. Cytokine Saga in Visceral Leishmaniasis. Cytokine 2021, 147, 155322. [Google Scholar] [CrossRef]
- Samant, M.; Sahu, U.; Pandey, S.C.; Khare, P. Role of Cytokines in Experimental and Human Visceral Leishmaniasis. Front. Cell. Infect. Microbiol. 2021, 11, 624009. [Google Scholar] [CrossRef] [PubMed]
- Kupani, M.; Pandey, R.K.; Mehrotra, S. Neutrophils and Visceral Leishmaniasis: Impact on Innate Immune Response and Cross-Talks with Macrophages and Dendritic Cells. J. Cell. Physiol. 2021, 236, 2255–2267. [Google Scholar] [CrossRef] [PubMed]
- McCall, L.I.; Zhang, W.W.; Matlashewski, G. Determinants for the Development of Visceral Leishmaniasis Disease. PLoS Pathog. 2013, 9, e1003053. [Google Scholar] [CrossRef] [PubMed]
- Scharton, B.T.M.; Scott, P. Natural Killer Cells Are a Source o f Interferon 3 ’ That Drives Differentiation o f CD4 + T Cell Subsets and Induces Early Resistance to Leishmania Major in Mice. J. Exp. Med. 1993, 178, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Kantzanou, M.; Karalexi, M.A.; Theodoridou, K.; Kostares, E.; Kostare, G.; Loka, T.; Vrioni, G.; Tsakris, A. Prevalence of Visceral Leishmaniasis among People with HIV: A Systematic Review and Meta-Analysis. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 42, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Das, V.N.R.; Bimal, S.; Siddiqui, N.A.; Kumar, A.; Pandey, K.; Sinha, S.K.; Topno, R.K.; Mahentesh, V.; Singh, A.K.; Lal, C.S.; et al. Conversion of Asymptomatic Infection to Symptomatic Visceral Leishmaniasis: A Study of Possible Immunological Markers. PLoS Negl. Trop. Dis. 2020, 14, e0008272. [Google Scholar] [CrossRef] [PubMed]
- Pikoulas, A.; Piperaki, E.T.; Spanakos, G.; Kallianos, A.; Mparmparousi, D.; Rentziou, G.; Trakada, G. Visceral Leishmaniasis and COVID-19 Coinfection—A Case Report. IDCases 2022, 27, e01358. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; Singh, S. Visceral Leishmaniasis in the COVID-19 Pandemic Era. Trans. R. Soc. Trop. Med. Hyg. 2022, 117, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Miotti, A.M.; Patacca, A.; Grosso, C.; Cristini, F. COVID-19 in a Patient with Visceral Leishmaniasis. J. Infect. Dis. Ther. 2020, 8, 4–5. [Google Scholar]
- Colomba, C.; Guccione, C.; Rubino, R.; Scalisi, M.; Condemi, A.; Bagarello, S.; Giordano, S.; Cascio, A. Third Case of Visceral Leishmaniasis in COVID-19: Mini Review Article. Pathogens 2022, 11, 913. [Google Scholar] [CrossRef] [PubMed]
- Coomes, E.A.; Haghbayan, H. Interleukin-6 in Covid-19: A Systematic Review and Meta-Analysis. Rev. Med. Virol. 2020, 30, e2141. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.; McAuley, D.F.; Brown, M.; Sanchez, E.; Tattersall, R.S.; Manson, J.J. COVID-19: Consider Cytokine Storm Syndromes and Immunosuppression. Lancet 2020, 395, 1033–1034. [Google Scholar] [CrossRef]
- Costa, A.S.A.; Costa, G.C.; de Aquino, D.M.C.; de Mendonça, V.R.R.; Barral, A.; Barral-Netto, M.; Caldas, A. de J.M. Cytokines and Visceral Leishmaniasis: A Comparison of Plasma Cytokine Profiles between the Clinical Forms of Visceral Leishmaniasis. Mem. Inst. Oswaldo Cruz 2012, 107, 735–739. [Google Scholar] [CrossRef]
- Caldas, A.; Favali, C.; Aquino, D.; Vinhas, V.; van Weyenbergh, J.; Brodskyn, C.; Costa, J.; Barral-Netto, M.; Barral, A. Balance of IL-10 and Interferon-γ Plasma Levels in Human Visceral Leishmaniasis: Implications in the Pathogenesis. BMC Infect. Dis. 2005, 5, 113. [Google Scholar] [CrossRef]
- Dutra, B.M.; Rodrigues, N.L.d.C.; Fonseca, F.R.M.; de Moura, T.R.; de Almeida, R.P.; de Jesus, A.R.; Abreu, T.M.; Pompeu, M.M.d.L.; Teixeira, C.R.; Teixeira, M.J. CXCL10 Immunomodulatory Effect against Infection Caused by an Antimony Refractory Isolate of Leishmania Braziliensis in Mice. Parasite Immunol. 2021, 43, e12805. [Google Scholar] [CrossRef]
- Khoshdel, A.; Alborzi, A.; Rosouli, M.; Taheri, E.; Kiany, S.; Javadian, M.H. Increased Levels of IL-10, IL-12, and IFN-γ in Patients with Visceral Leishmaniasis. Brazilian J. Infect. Dis. 2009, 13, 44–46. [Google Scholar] [CrossRef]
- Kumari, S.; Shivam, P.; Kumar, S.; Jamal, F.; Singh, M.K.; Bimal, S.; Narayan, S.; Pandey, K.; Das, V.N.R.; Das, P.; et al. Leishmania Donovani Mediated Higher Expression of CCL4 Induces Differential Accumulation of CD4+CD56+NKT and CD8+CD56+NKT Cells at Infection Site. Cytokine 2018, 110, 306–315. [Google Scholar] [CrossRef]
- Scott, B.Y.P.; Natovitz, P.; Coffman, R.L.; Pearce, E.; Sher, A. Immunoregulation of Cutaneous Leishmaniasis. T Cell Lines That Transfer Protective Immunity or Exacerbation Belong to Different T Helper Subsets and Respond to Distinct Parasite Antigens. J. Exp. Med. 1988, 168, 1675–1684. [Google Scholar] [CrossRef]
- Kropf, P.; Fuentes, J.M.; Fähnrich, E.; Arpa, L.; Herath, S.; Weber, V.; Soler, G.; Celada, A.; Modolell, M.; Müller, I. Arginase and Polyamine Synthesis Are Key Factors in the Regulation of Experimental Leishmaniasis In Vivo. FASEB J. 2005, 19, 1000–1002. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef]
- Zandbergen, G. Van Anti-Tumor Necrosis Factor α Therapeutics Differentially Affect Leishmania Infection of Human Macrophages. Front. Immunol. 2018, 9, 1772. [Google Scholar] [CrossRef]
- Gessner, A.; Vieth, M.; Will, A.; Schröppel, K.R.M. Interleukin-7 Enhances Antimicrobial Activity against Leishmania Major in Murine Macrophages. Infect Immun. 1993, 61, 4008–4012. [Google Scholar] [CrossRef] [PubMed]
- Gessner, A.; Will, A.; Vieth, M.; Schröppel, K.; Röllinghoff, M. Stimulation of B-Cell Lymphopoiesis by Interleukin-7 Leads to Aggravation of Murine Leishmaniasis. Immunology 1995, 85, 416–422. [Google Scholar]
- Hoerauf, A.; Solbach, W.; Rollinghoff, M.; Gessner, A. Effect of IL-7 Treatment on Leishmania Major-Infected BALB.Xid Mice: Enhanced Lymphopoiesis with Sustained Lack of B1 Cells and Clinical Aggravation of Disease. Int. Immunol. 1995, 7, 1879–1884. [Google Scholar] [PubMed]
- Quiros-Roldan, E.; Sottini, A.; Signorini, S.G.; Serana, F.; Tiecco, G.; Imberti, L. Autoantibodies to Interferons in Infectious Diseases. Viruses 2023, 15, 1215. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, J. Mendelian Susceptibility to Mycobacterial Disease: Recent Discoveries. Hum. Genet. 2020, 139, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Jouanguy, E.; Lamhamedi-Cherradi, S.; Lammas, D.; Dorman, S.E.; Fondanèche, M.C.; Dupuis, S.; Döffinger, R.; Altare, F.; Girdlestone, J.; Emile, J.F.; et al. A Human IFNGR1 Small Deletion Hotspot Associated with Dominant Susceptibility to Mycobacterial Infection. Nat. Genet. 1999, 21, 370–378. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Risk Factors | Patient 1 | Patient 2 | Patient 3 |
|---|---|---|---|
| Leishmania previous infection | No | No | No |
| Last travel in endemic area | Greece (October 2019) | Albania (August 2021) | Greece (August 2021) |
| Previous imported infections | None | None | None |
| Pets or animal exposure | Domestic dog | None | Domestic dog |
| Previous opportunistic infections | VZV, hHSV-1 | None | VZV |
| HIV-antibodies | Negative | Negative | Negative |
| Autoimmunity markers | RF + | RF +, dCT +, ANA +, ↓ C3-C4 | RF +, dCT +, ANA + |
| Cytokines | Healthy Controls | Patient 1 | Patient 2 | Patient 3 | p-Value |
|---|---|---|---|---|---|
| CXCL-10 | 201, 168–248 | 584 | 914 | 792 | <0.001 |
| CCL-4 | 52, 36–87 | 88 | 100 | 127 | 0.044 |
| IL-6 | 0.40, 0.28–0.60 | 1.06 | 4.90 | 1.40 | 0.003 |
| Cytokines | Healthy Controls | Patient 1 | Patient 2 | Patient 3 | p-Value |
|---|---|---|---|---|---|
| TNF-α | 2.40, 1.27–7.82 | 9.01 | 1.80 | 3.21 | 0.022 |
| IL-7 | 5.51, 3.05–7.51 | 21.43 | 6.04 | 6.41 | <0.001 |
| Cytokines | Healthy Controls | Patient 1 | Patient 2 | Patient 3 | p-Value |
|---|---|---|---|---|---|
| IL-1α | 0.15, 0.04–0.3 | 0.3 | 1.8 | 6.5 | 0.020 |
| IL-4 | 0.01, 0.01–0.03 | 0 | 0.03 | 0.24 | 0.057 |
| IL-5 | 0.45, 0.20–0.80 | 0.58 | 0.86 | 1.86 | 0.040 |
| IL-10 | 0.26, 0.2–0.3 | 0.3 | 3.5 | 2.5 | <0.001 |
| IL-12 | 150, 103–216 | 134 | 201 | 599 | 0.009 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mulè, A.; Crosato, V.; Kuhns, D.B.; Lorenzi, L.; Chirico, C.; Maifredi, G.; Notarangelo, L.D.; Castelli, F.; Tomasoni, L.R. Visceral Leishmaniasis in Immunocompetent Hosts in Brescia: A Case Series and Analysis of Cytokine Cascade. Microorganisms 2024, 12, 394. https://doi.org/10.3390/microorganisms12020394
Mulè A, Crosato V, Kuhns DB, Lorenzi L, Chirico C, Maifredi G, Notarangelo LD, Castelli F, Tomasoni LR. Visceral Leishmaniasis in Immunocompetent Hosts in Brescia: A Case Series and Analysis of Cytokine Cascade. Microorganisms. 2024; 12(2):394. https://doi.org/10.3390/microorganisms12020394
Chicago/Turabian StyleMulè, Alice, Verena Crosato, Douglas Byron Kuhns, Luisa Lorenzi, Claudia Chirico, Giovanni Maifredi, Luigi D. Notarangelo, Francesco Castelli, and Lina R. Tomasoni. 2024. "Visceral Leishmaniasis in Immunocompetent Hosts in Brescia: A Case Series and Analysis of Cytokine Cascade" Microorganisms 12, no. 2: 394. https://doi.org/10.3390/microorganisms12020394
APA StyleMulè, A., Crosato, V., Kuhns, D. B., Lorenzi, L., Chirico, C., Maifredi, G., Notarangelo, L. D., Castelli, F., & Tomasoni, L. R. (2024). Visceral Leishmaniasis in Immunocompetent Hosts in Brescia: A Case Series and Analysis of Cytokine Cascade. Microorganisms, 12(2), 394. https://doi.org/10.3390/microorganisms12020394