

From Viral Infection to Autoimmune Reaction: Exploring the Link between Human Herpesvirus 6 and Autoimmune Diseases

 , ,
, ,  and
and
Abstract
1. Introduction
- High prevalence and early primary infection. The global seroprevalence of HHV-6 ranges up to 95%, although the exact prevalence of each species is unclear [13,14,15]. Primary HHV-6 infection occurs in the first few years of life, with the earliest infection coinciding with the veining of maternal antibody titers [16,17]. It is generally believed that the majority of primary infections are caused by HHV-6B, in many cases associated with the exanthem subitum, an early febrile illness, which in rare cases can lead to encephalitis. HHV-6A is acquired later in life through asymptomatic infection [12].
- Establishment of latency, with the potential of reactivation and persistent infection. After the early primary infection, HHV-6, like other herpesviruses, can establish lifelong latency and reactivate. HHV-6 can cause a persistent infection, thus creating a prolonged inflammation due to viral antigens and the immune system’s response to them. Researchers believe HHV-6 mainly establishes latency by integrating into its hosts’ genome [18]. On rare occasions, the integration can be established in gametes, leading to a condition called inherited chromosomally integrated HHV-6 (iciHHV-6) [19]. The exact factors of HHV-6 reactivation are not definitively demonstrated, but reactivation has caused a severe disease burden. Neurological manifestations like encephalitis in immunocompromised individuals who are HIV-positive or have undergone solid organ or hematopoietic stem-cell transplantation have been associated with HHV-6 reactivation [20].
- Broad cell tropism. Even though HHV-6 was considered a strictly lymphotropic virus, its cell tropism has turned out to be much broader. Originally the cell entry receptor of both HHV-6 species was thought to be CD46 [21]. CD46 is present in all nucleated cells and is important in complement regulation [22]. Later, it was demonstrated that several strains of HHV-6B did not utilize this receptor and that soluble CD46 could not inhibit the infection. Studies determined that the cell entry receptor primarily used by HHV-B is CD134 [23,24]. Other widely expressed cell surface molecules have also been proposed to be important for HHV-6 entry. These include nectin cell adhesion molecule 2 (CD112) [25] and gp96 (glucose-regulated protein 94) [26]. HHV-6 has been found in cells and tissues with various origins, including immune, endothelial, epithelial, and neuronal, with HHV-6A showing an increased propensity to infect neuronal cells [12,27,28].
- Immune evasion and modulation mechanisms. Herpesviruses have long evolved and adapted to ensure a productive infection in their specific host. A large part of their genomes have been devoted to evading and modulating the host immune response. HHV-6 is no exception [29]. The “immunotropic” nature of HHV-6 in itself can be a mechanism of immunomodulation. The infection has been shown to alter immune cell functionality, for example, changing the profile of secreted immune mediators. HHV-6 infection can alter cell surface molecule expression, causing the downregulation of its cell entry receptor CD46, thus dysregulating complement activation, leading to tissue damage. Also, HHV-6 has been shown to increase cytokine levels and create an inflammatory environment [30,31]. Many herpesviruses encode chemokines and chemokine receptors as an immune modulation strategy. HHV-6 encodes a chemokine (U83) and two chemokine receptors (U12 and U51) [32]. Both of the chemokine receptors are quite poorly studied. However, studies have shown they bind a variety of human chemokines CCL2, 3, 4, 6, and others. In the case of the chemokine RANTES (CCL5), U51 has been shown to downregulate its expression and secretion [32,33]. A role in viral replication and possibly cell-to-cell transmission has also been demonstrated for U51 [34]. Additionally, U12 and U51 are putative G-protein couple-receptor (GPCR) homologs. They possess similarities with human CCRs not only in functionality, but on a molecular level as well. The ability to spread cell-to-cell is an immune evasion strategy since this mode of transmission could allow the virus to be practically invisible to the immune system and persist in solid tissues [35].
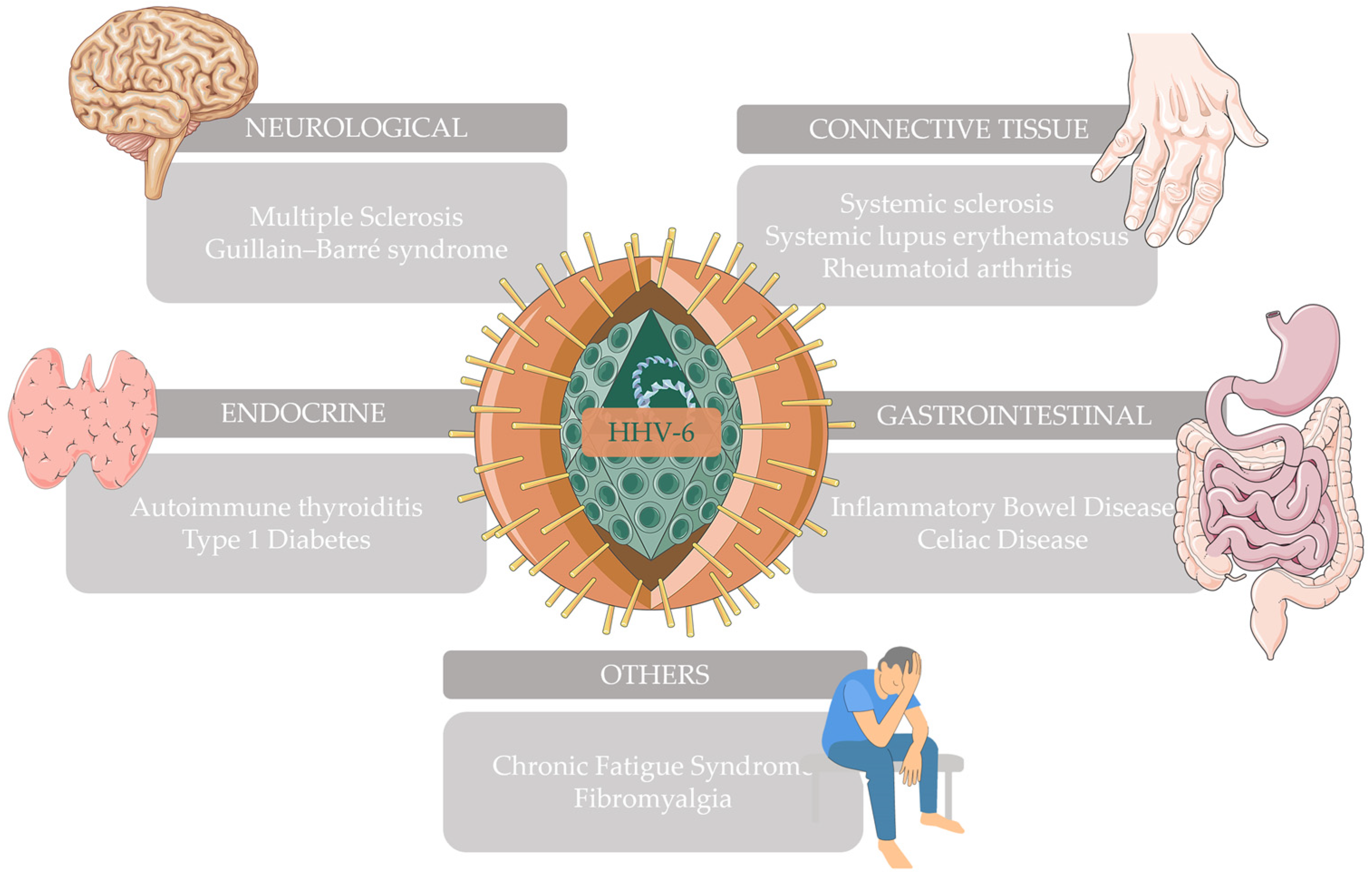
2. Autoimmune Neurological Diseases
3. Autoimmune Connective Tissue Diseases
4. Autoimmune Gastrointestinal Diseases
5. Autoimmune Endocrine Diseases
6. Other Autoimmune Diseases
7. DIHS/DRESS and Autoimmune Diseases
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ramos, P.S.; Shedlock, A.M.; Langefeld, C.D. Genetics of Autoimmune Diseases: Insights from Population Genetics. J. Hum. Genet. 2015, 60, 657–664. [Google Scholar] [CrossRef]
- Costenbader, K.H.; Gay, S.; Alarcón-Riquelme, M.E.; Iaccarino, L.; Doria, A. Genes, Epigenetic Regulation and Environmental Factors: Which Is the Most Relevant in Developing Autoimmune Diseases? Autoimmun. Rev. 2012, 11, 604–609. [Google Scholar] [CrossRef]
- Smatti, M.K.; Cyprian, F.S.; Nasrallah, G.K.; Al Thani, A.A.; Almishal, R.O.; Yassine, H.M. Viruses and Autoimmunity: A Review on the Potential Interaction and Molecular Mechanisms. Viruses 2019, 11, 762. [Google Scholar] [CrossRef]
- Fujinami, R.S.; von Herrath, M.G.; Christen, U.; Whitton, J.L. Molecular Mimicry, Bystander Activation, or Viral Persistence: Infections and Autoimmune Disease. Clin. Microbiol. Rev. 2006, 19, 80–94. [Google Scholar] [CrossRef]
- Salahuddin, S.Z.; Ablashi, D.V.; Markham, P.D.; Josephs, S.F.; Sturzenegger, S.; Kaplan, M.; Halligan, G.; Biberfeld, P.; Wong-Staal, F.; Kramarsky, B.; et al. Isolation of a New Virus, HBLV, in Patients with Lymphoproliferative Disorders. Science 1986, 234, 596–601. [Google Scholar] [CrossRef]
- Ablashi, D.; Agut, H.; Alvarez-Lafuente, R.; Clark, D.A.; Dewhurst, S.; DiLuca, D.; Flamand, L.; Frenkel, N.; Gallo, R.; Gompels, U.A.; et al. Classification of HHV-6A and HHV-6B as Distinct Viruses. Arch. Virol. 2014, 159, 863–870. [Google Scholar] [CrossRef]
- Caselli, E.; Zatelli, M.C.; Rizzo, R.; Benedetti, S.; Martorelli, D.; Trasforini, G.; Cassai, E.; degli Uberti, E.C.; Luca, D.D.; Dolcetti, R. Virologic and Immunologic Evidence Supporting an Association between HHV-6 and Hashimoto’s Thyroiditis. PLoS Pathog. 2012, 8, e1002951. [Google Scholar] [CrossRef] [PubMed]
- Sultanova, A.; Cistjakovs, M.; Gravelsina, S.; Chapenko, S.; Roga, S.; Cunskis, E.; Nora-Krukle, Z.; Groma, V.; Ventina, I.; Murovska, M. Association of Active Human Herpesvirus-6 (HHV-6) Infection with Autoimmune Thyroid Gland Diseases. Clin. Microbiol. Infect. 2017, 23, e1–e50. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Ma, Y.; Gong, F.; Hu, C.; Qian, L.; Huang, Q.; Yu, Q.; Zhang, J.; Chen, S.; Liu, Z.; et al. Cross-Reactivity of Autoreactive T Cells with MBP and Viral Antigens in Patients with MS. Front. Biosci. 2012, 17, 1648–1658. [Google Scholar] [CrossRef] [PubMed]
- Tejada-Simon, M.V.; Zang, Y.C.Q.; Hong, J.; Rivera, V.M.; Zhang, J.Z. Cross-Reactivity with Myelin Basic Protein and Human Herpesvirus-6 in Multiple Sclerosis. Ann. Neurol. 2003, 53, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Tsao, E.H.; Kellam, P.; Sin, C.S.Y.; Rasaiyaah, J.; Griffiths, P.D.; Clark, D.A. Microarray-Based Determination of the Lytic Cascade of Human Herpesvirus 6B. J. Gen. Virol. 2009, 90, 2581–2591. [Google Scholar] [CrossRef] [PubMed]
- Agut, H.; Bonnafous, P.; Gautheret-Dejean, A. Laboratory and Clinical Aspects of Human Herpesvirus 6 Infections. Clin. Microbiol. Rev. 2015, 28, 313–335. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.; Sumaya, C.; Liu, C.-R.; Ench, Y.; Kovacs, A.; Coronesi, M.; Kaplan, M. Fall in human herpesvirus 6 seropositivity with age. Lancet 1988, 332, 396. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.A.; Ferro, F.; Greenspan, D.; Lennette, E.T. Frequent Isolation of HHV-6 from Saliva and High Seroprevalence of the Virus in the Population. Lancet 1990, 335, 1047–1050. [Google Scholar] [CrossRef] [PubMed]
- Okuno, T.; Takahashi, K.; Balachandra, K.; Shiraki, K.; Yamanishi, K.; Takahashi, M.; Baba, K. Seroepidemiology of Human Herpesvirus 6 Infection in Normal Children and Adults. J. Clin. Microbiol. 1989, 27, 651–653. [Google Scholar] [CrossRef]
- Zerr, D.M.; Meier, A.S.; Selke, S.S.; Frenkel, L.M.; Huang, M.-L.; Wald, A.; Rhoads, M.P.; Nguy, L.; Bornemann, R.; Morrow, R.A.; et al. A Population-Based Study of Primary Human Herpesvirus 6 Infection. N. Engl. J. Med. 2005, 352, 768–776. [Google Scholar] [CrossRef]
- Hall, C.B.; Long, C.E.; Schnabel, K.C.; Caserta, M.T.; McIntyre, K.M.; Costanzo, M.A.; Knott, A.; Dewhurst, S.; Insel, R.A.; Epstein, L.G. Human Herpesvirus-6 Infection in Children—A Prospective Study of Complications and Reactivation. N. Engl. J. Med. 1994, 331, 432–438. [Google Scholar] [CrossRef]
- Pantry, S.N.; Medveczky, P.G. Latency, Integration, and Reactivation of Human Herpesvirus-6. Viruses 2017, 9, 194. [Google Scholar] [CrossRef]
- Flamand, L. Chromosomal Integration by Human Herpesviruses 6A and 6B. In Human Herpesviruses; Kawaguchi, Y., Mori, Y., Kimura, H., Eds.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2018; pp. 209–226. ISBN 978-981-10-7230-7. [Google Scholar]
- Agut, H.; Bonnafous, P.; Gautheret-Dejean, A. Human Herpesviruses 6A, 6B, and 7. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Santoro, F.; Kennedy, P.E.; Locatelli, G.; Malnati, M.S.; Berger, E.A.; Lusso, P. CD46 Is a Cellular Receptor for Human Herpesvirus 6. Cell 1999, 99, 817–827. [Google Scholar] [CrossRef]
- Yamamoto, H.; Fara, A.F.; Dasgupta, P.; Kemper, C. CD46: The ‘Multitasker’ of Complement Proteins. Int. J. Biochem. Cell Biol. 2013, 45, 2808–2820. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Serada, S.; Kawabata, A.; Ota, M.; Hayashi, E.; Naka, T.; Yamanishi, K.; Mori, Y. CD134 Is a Cellular Receptor Specific for Human Herpesvirus-6B Entry. Proc. Natl. Acad. Sci. USA 2013, 110, 9096–9099. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Mori, Y. Determinants of Human CD134 Essential for Entry of Human Herpesvirus 6B. J. Virol. 2015, 89, 10125–10129. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, H.; Fujikura, D.; Namba, H.; Yamashita, N.; Honda, T.; Yamada, M. Nectin-2 Acts as a Viral Entry Mediated Molecule That Binds to Human Herpesvirus 6B Glycoprotein B. Viruses 2022, 14, 160. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Jia, J.; Jiang, X.; Xu, M.; Guo, J.; Tang, T.; Xu, X.; Wu, Z.; Hu, B.; Yao, K.; et al. Gp96 Is Critical for Both Human Herpesvirus 6A (HHV-6A) and HHV-6B Infections. J. Virol. 2020, 94, e00311-20. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Hudnall, S.D. Anatomical Mapping of Human Herpesvirus Reservoirs of Infection. Mod. Pathol. 2006, 19, 726–737. [Google Scholar] [CrossRef]
- De Bolle, L.; Van Loon, J.; De Clercq, E.; Naesens, L. Quantitative Analysis of Human Herpesvirus 6 Cell Tropism. J. Med. Virol. 2005, 75, 76–85. [Google Scholar] [CrossRef]
- Cruz-Muñoz, M.E.; Fuentes-Pananá, E.M. Beta and Gamma Human Herpesviruses: Agonistic and Antagonistic Interactions with the Host Immune System. Front. Microbiol. 2018, 8, 2521. [Google Scholar] [CrossRef]
- Lusso, P. HHV-6 and the Immune System: Mechanisms of Immunomodulation and Viral Escape. J. Clin. Virol. 2006, 37 (Suppl. S1), S4–S10. [Google Scholar] [CrossRef]
- Dagna, L.; Pritchett, J.C.; Lusso, P. Immunomodulation and Immunosuppression by Human Herpesvirus 6A and 6B. Future Virol. 2013, 8, 273–287. [Google Scholar] [CrossRef]
- Pontejo, S.M.; Murphy, P.M.; Pease, J.E. Chemokine Subversion by Human Herpesviruses. J. Innate Immun. 2018, 10, 465–478. [Google Scholar] [CrossRef]
- Milne, R.S.; Mattick, C.; Nicholson, L.; Devaraj, P.; Alcami, A.; Gompels, U.A. RANTES Binding and Down-Regulation by a Novel Human Herpesvirus-6 Beta Chemokine Receptor. J. Immunol. 2000, 164, 2396–2404. [Google Scholar] [CrossRef]
- Zhen, Z.; Bradel-Tretheway, B.; Sumagin, S.; Bidlack, J.M.; Dewhurst, S. The Human Herpesvirus 6 G Protein-Coupled Receptor Homolog U51 Positively Regulates Virus Replication and Enhances Cell-Cell Fusion in Vitro. J. Virol. 2005, 79, 11914–11924. [Google Scholar] [CrossRef]
- Mothes, W.; Sherer, N.M.; Jin, J.; Zhong, P. Virus Cell-to-Cell Transmission. J. Virol. 2010, 84, 8360–8368. [Google Scholar] [CrossRef]
- Tarlinton, R.E.; Martynova, E.; Rizvanov, A.A.; Khaiboullina, S.; Verma, S. Role of Viruses in the Pathogenesis of Multiple Sclerosis. Viruses 2020, 12, 643. [Google Scholar] [CrossRef] [PubMed]
- Bjornevik, K.; Münz, C.; Cohen, J.I.; Ascherio, A. Epstein–Barr Virus as a Leading Cause of Multiple Sclerosis: Mechanisms and Implications. Nat. Rev. Neurol. 2023, 19, 160–171. [Google Scholar] [CrossRef]
- Komaroff, A.L.; Pellett, P.E.; Jacobson, S. Human Herpesviruses 6A and 6B in Brain Diseases: Association versus Causation. Clin. Microbiol. Rev. 2020, 34, e00143-20. [Google Scholar] [CrossRef] [PubMed]
- Challoner, P.B.; Smith, K.T.; Parker, J.D.; MacLeod, D.L.; Coulter, S.N.; Rose, T.M.; Schultz, E.R.; Bennett, J.L.; Garber, R.L.; Chang, M. Plaque-Associated Expression of Human Herpesvirus 6 in Multiple Sclerosis. Proc. Natl. Acad. Sci. USA 1995, 92, 7440–7444. [Google Scholar] [CrossRef] [PubMed]
- Gordon, L.; McQuaid, S.; Cosby, S.L. Detection of Herpes Simplex Virus (Types 1 and 2) and Human Herpesvirus 6 DNA in Human Brain Tissue by Polymerase Chain Reaction. Clin. Diagn. Virol. 1996, 6, 33–40. [Google Scholar] [CrossRef]
- Sanders, V.J.; Felisan, S.; Waddell, A.; Tourtellotte, W.W. Detection of Herpesviridae in Postmortem Multiple Sclerosis Brain Tissue and Controls by Polymerase Chain Reaction. J. Neurovirol. 1996, 2, 249–258. [Google Scholar] [CrossRef]
- Blumberg, B.M.; Mock, D.J.; Powers, J.M.; Ito, M.; Assouline, J.G.; Baker, J.V.; Chen, B.; Goodman, A.D. The HHV6 Paradox: Ubiquitous Commensal or Insidious Pathogen? A Two-Step in Situ PCR Approach. J. Clin. Virol. 2000, 16, 159–178. [Google Scholar] [CrossRef]
- Knox, K.K.; Brewer, J.H.; Henry, J.M.; Harrington, D.J.; Carrigan, D.R. Human Herpesvirus 6 and Multiple Sclerosis: Systemic Active Infections in Patients with Early Disease. Clin. Infect. Dis. 2000, 31, 894–903. [Google Scholar] [CrossRef]
- Cermelli, C.; Berti, R.; Soldan, S.S.; Mayne, M.; D’ambrosia, J.M.; Ludwin, S.K.; Jacobson, S. High Frequency of Human Herpesvirus 6 DNA in Multiple Sclerosis Plaques Isolated by Laser Microdissection. J. Infect. Dis. 2003, 187, 1377–1387. [Google Scholar] [CrossRef]
- Goodman, A.D.; Mock, D.J.; Powers, J.M.; Baker, J.V.; Blumberg, B.M. Human Herpesvirus 6 Genome and Antigen in Acute Multiple Sclerosis Lesions. J. Infect. Dis. 2003, 187, 1365–1376. [Google Scholar] [CrossRef]
- Opsahl, M.L. Early and Late HHV-6 Gene Transcripts in Multiple Sclerosis Lesions and Normal Appearing White Matter. Brain 2005, 128, 516–527. [Google Scholar] [CrossRef]
- Tuke, P.W.; Hawke, S.; Griffiths, P.D.; Clark, D.A. Distribution and Quantification of Human Herpesvirus 6 in Multiple Sclerosis and Control Brains. Mult. Scler. 2004, 10, 355–359. [Google Scholar] [CrossRef]
- Mameli, G.; Astone, V.; Arru, G.; Marconi, S.; Lovato, L.; Serra, C.; Sotgiu, S.; Bonetti, B.; Dolei, A. Brains and Peripheral Blood Mononuclear Cells of Multiple Sclerosis (MS) Patients Hyperexpress MS-Associated Retrovirus/HERV-W Endogenous Retrovirus, but Not Human Herpesvirus 6. J. Gen. Virol. 2007, 88, 264–274. [Google Scholar] [CrossRef]
- Goldberg, S.H.; Albright, A.V.; Lisak, R.P.; González-Scarano, F. Polymerase Chain Reaction Analysis of Human Herpesvirus-6 Sequences in the Sera and Cerebrospinal Fluid of Patients with Multiple Sclerosis. J. Neurovirol. 1999, 5, 134–139. [Google Scholar] [CrossRef]
- Tejada-Simon, M.V.; Zang, Y.C.Q.; Hong, J.; Rivera, V.M.; Killian, J.M.; Zhang, J.Z. Detection of Viral DNA and Immune Responses to the Human Herpesvirus 6 101-Kilodalton Virion Protein in Patients with Multiple Sclerosis and in Controls. J. Virol. 2002, 76, 6147–6154. [Google Scholar] [CrossRef]
- Álvarez-Lafuente, R.; Martín-Estefanía, C.; De Las Heras, V.; Castrillo, C.; Picazo, J.J.; Varela De Seijas, E.; González, R.A. Active Human Herpesvirus 6 Infection in Patients with Multiple Sclerosis. Arch. Neurol. 2002, 59, 929. [Google Scholar] [CrossRef]
- Álvarez-Lafuente, R.; De Las Heras, V.; Bartolomé, M.; Picazo, J.J.; Arroyo, R. Relapsing-Remitting Multiple Sclerosis and Human Herpesvirus 6 Active Infection. Arch. Neurol. 2004, 61, 1523. [Google Scholar] [CrossRef]
- Álvarez-Lafuente, R.; García-Montojo, M.; De Las Heras, V.; Domínguez-Mozo, M.; Bartolome, M.; Benito-Martin, M.; Arroyo, R. Herpesviruses and Human Endogenous Retroviral Sequences in the Cerebrospinal Fluid of Multiple Sclerosis Patients. Mult. Scler. 2008, 14, 595–601. [Google Scholar] [CrossRef]
- Ramroodi, N.; Sanadgol, N.; Ganjali, Z.; Niazi, A.A.; Sarabandi, V.; Moghtaderi, A. Monitoring of Active Human Herpes Virus 6 Infection in Iranian Patients with Different Subtypes of Multiple Sclerosis. J. Pathog. 2013, 2013, 194932. [Google Scholar] [CrossRef]
- Domínguez-Mozo, M.I.; Nieto-Guerrero, A.; Pérez-Pérez, S.; García-Martínez, M.Á.; Arroyo, R.; Álvarez-Lafuente, R. MicroRNAs of Human Herpesvirus 6A and 6B in Serum and Cerebrospinal Fluid of Multiple Sclerosis Patients. Front. Immunol. 2020, 11, 2142. [Google Scholar] [CrossRef]
- Berti, R.; Brennan, M.B.; Soldan, S.S.; Ohayon, J.M.; Casareto, L.; McFarland, H.F.; Jacobson, S. Increased Detection of Serum HHV-6 DNA Sequences during Multiple Sclerosis (MS) Exacerbations and Correlation with Parameters of MS Disease Progression. J. Neurovirol. 2002, 8, 250–256. [Google Scholar] [CrossRef]
- Chapenko, S.; Millers, A.; Nora, Z.; Logina, I.; Kukaine, R.; Murovska, M. Correlation between HHV-6 Reactivation and Multiple Sclerosis Disease Activity. J. Med. Virol. 2003, 69, 111–117. [Google Scholar] [CrossRef]
- Behzad-Behbahani, A.; Mikaeili, M.H.; Entezam, M.; Mojiri, A.; Pour, G.Y.; Arasteh, M.M.; Rahsaz, M.; Banihashemi, M.; Khadang, B.; Moaddeb, A.; et al. Human Herpesvirus-6 Viral Load and Antibody Titer in Serum Samples of Patients with Multiple Sclerosis. J. Microbiol. Immunol. Infect. 2011, 44, 247–251. [Google Scholar] [CrossRef]
- Nora-Krukle, Z.; Chapenko, S.; Logina, I.; Millers, A.; Platkajis, A.; Murovska, M. Human Herpesvirus 6 and 7 Reactivation and Disease Activity in Multiple Sclerosis. Medicina 2011, 47, 75. [Google Scholar] [CrossRef]
- Ablashi, D.V.; Eastman, H.B.; Owen, C.B.; Roman, M.M.; Friedman, J.; Zabriskie, J.B.; Peterson, D.L.; Pearson, G.R.; Whitman, J.E. Frequent HHV-6 Reactivation in Multiple Sclerosis (MS) and Chronic Fatigue Syndrome (CFS) Patients. J. Clin. Virol. 2000, 16, 179–191. [Google Scholar] [CrossRef]
- Villoslada, P.; Juste, C.; Tintore, M.; Llorenç, V.; Codina, G.; Pozo-Rosich, P.; Montalban, X. The Immune Response against Herpesvirus Is More Prominent in the Early Stages of MS. Neurology 2003, 60, 1944–1948. [Google Scholar] [CrossRef]
- Riverol, M.; Sepulcre, J.; Fernandez-Alonso, M.; Uccelli, A.; Brieva, L.; Rubio, M.; Rodriguez, A.; Fernandez-Diez, B.; Villoslada, P. Antibodies against Epstein–Barr Virus and Herpesvirus Type 6 Are Associated with the Early Phases of Multiple Sclerosis. J. Neuroimmunol. 2007, 192, 184–185. [Google Scholar] [CrossRef]
- Khaki, M.; Ghazavi, A.; Ghasami, K.; Rafiei, M.; Payani, M.-A.; Ghaznavi-Rad, E.; Mosayebi, G. Evaluation of Viral Antibodies in Iranian Multiple Sclerosis Patients. Neurosci. J. 2011, 16, 224–228. [Google Scholar]
- Ortega-Madueño, I.; Garcia-Montojo, M.; Dominguez-Mozo, M.I.; Garcia-Martinez, A.; Arias-Leal, A.M.; Casanova, I.; Arroyo, R.; Alvarez-Lafuente, R. Anti-Human Herpesvirus 6A/B IgG Correlates with Relapses and Progression in Multiple Sclerosis. PLoS ONE 2014, 9, e104836. [Google Scholar] [CrossRef]
- Engdahl, E.; Gustafsson, R.; Huang, J.; Biström, M.; Lima Bomfim, I.; Stridh, P.; Khademi, M.; Brenner, N.; Butt, J.; Michel, A.; et al. Increased Serological Response Against Human Herpesvirus 6A Is Associated with Risk for Multiple Sclerosis. Front. Immunol. 2019, 10, 2715. [Google Scholar] [CrossRef]
- Keyvani, H.; Zahednasab, H.; ali abraham Aljanabi, H.; Asadi, M.; Mirzaei, R.; Esghaei, M.; Karampoor, S. The Role of Human Herpesvirus-6 and Inflammatory Markers in the Pathogenesis of Multiple Sclerosis. J. Neuroimmunol. 2020, 346, 577313. [Google Scholar] [CrossRef]
- Flamand, L.; Stefanescu, I.; Ablashi, D.V.; Menezes, J. Activation of the Epstein-Barr Virus Replicative Cycle by Human Herpesvirus 6. J. Virol. 1993, 67, 6768–6777. [Google Scholar] [CrossRef]
- Fierz, W. Multiple Sclerosis: An Example of Pathogenic Viral Interaction? Virol. J. 2017, 14, 42. [Google Scholar] [CrossRef]
- Jakhmola, S.; Upadhyay, A.; Jain, K.; Mishra, A.; Jha, H.C. Herpesviruses and the Hidden Links to Multiple Sclerosis Neuropathology. J. Neuroimmunol. 2021, 358, 577636. [Google Scholar] [CrossRef]
- Sang, Y.; Zhang, R.; Scott, W.R.P.; Creagh, A.L.; Haynes, C.A.; Straus, S.K. U24 from Roseolovirus Interacts Strongly with Nedd4 WW Domains. Sci. Rep. 2017, 7, 39776. [Google Scholar] [CrossRef]
- Chandrashekhar, S.; Dimachkie, M. Guillain-Barré Syndrome in Adults: Pathogenesis, Clinical Features, and Diagnosis. Available online: https://www-uptodate-com.db.rsu.lv/contents/guillain-barre-syndrome-in-adults-pathogenesis-clinical-features-and-diagnosis?search=guillain%20barre&source=search_result&selectedTitle=1~150&usage_type=default&display_rank=1#H2546552464 (accessed on 23 November 2021).
- Gustafsson, R.; Reitsma, R.; Strålfors, A.; Lindholm, A.; Press, R.; Fogdell-Hahn, A. Incidence of Human Herpesvirus 6 in Clinical Samples from Swedish Patients with Demyelinating Diseases. J. Microbiol. Immunol. Infect. 2014, 47, 418–421. [Google Scholar] [CrossRef][Green Version]
- Pandey, U.; Greninger, A.L.; Levin, G.R.; Jerome, K.R.; Anand, V.C.; Dien Bard, J. Pathogen or Bystander: Clinical Significance of Detecting Human Herpesvirus 6 in Pediatric Cerebrospinal Fluid. J. Clin. Microbiol. 2020, 58, e00313-20. [Google Scholar] [CrossRef] [PubMed]
- Gaeta, A.; Verzaro, S.; Cristina, L.M.; Mancini, C.; Nazzari, C. Diagnosis of Neurological Herpesvirus Infections: Real Time PCR in Cerebral Spinal Fluid Analysis. New Microbiol. 2009, 32, 333–340. [Google Scholar] [PubMed]
- Galvan, M.; Rotola, A.; Govoni, V.; Granieri, E.; Cassai, E.; Di Luca, D. Simultaneous Guillain-Barrè Syndrome and Active Human Herpesvirus 6 Infection in the Central Nervous System. J. Clin. Virol. 2007, 38, 271–272. [Google Scholar] [CrossRef] [PubMed]
- Miyake, F.; Yoshikawa, T.; Suzuki, K.; Ohashi, M.; Suga, S.; Asano, Y. Guillain-Barré Syndrome after Exanthem Subitum. Pediatr. Infect. Dis. J. 2002, 21, 569–570. [Google Scholar] [CrossRef] [PubMed]
- Tomaszewska, A.; Nasilowska-Adamska, B.; Dzieciatkowski, T.; Marianska, B. Simultaneous Human Herpesvirus 6-Associated Encephalitis and Guillain-Barré Syndrome in a Patient after Matched Unrelated Donor Haematopoietic Stem Cell Transplantation. Arch. Med. Sci. 2010, 2, 288–290. [Google Scholar] [CrossRef] [PubMed]
- Piras, E.; Caocci, G.; Pisano, V.; Orrù, F.; Murgia, F.; Sanna, M.; Vacca, A.; La Nasa, G. Guillain–Barré Syndrome after Human Herpesvirus-6 Reactivation in Unrelated Hematopoietic Stem Cell Transplantation. Leuk. Lymphoma 2013, 54, 1332–1333. [Google Scholar] [CrossRef] [PubMed]
- Inazawa, N.; Hori, T.; Hatakeyama, N.; Yamamoto, M.; Yoto, Y.; Nojima, M.; Suzuki, N.; Shimizu, N.; Tsutsumi, H. Large-Scale Multiplex Polymerase Chain Reaction Assay for Diagnosis of Viral Reactivations after Allogeneic Hematopoietic Stem Cell Transplantation. J. Med. Virol. 2015, 87, 1427–1435. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.L.; Pritchett, J.C.; Leifer, C.; Zerr, D.M.; Koelle, D.M.; Di Luca, D.; Lusso, P. HHV-6B Infection, T-Cell Reconstitution, and Graft-vs-Host Disease after Hematopoietic Stem Cell Transplantation. Bone Marrow Transpl. 2018, 53, 1508–1517. [Google Scholar] [CrossRef]
- Broccolo, F.; Drago, F.; Paolino, S.; Cassina, G.; Gatto, F.; Fusetti, L.; Matteoli, B.; Zaccaria, E.; Parodi, A.; Lusso, P.; et al. Reactivation of Human Herpesvirus 6 (HHV-6) Infection in Patients with Connective Tissue Diseases. J. Clin. Virol. 2009, 46, 43–46. [Google Scholar] [CrossRef]
- Broccolo, F.; Drago, F.; Cassina, G.; Fava, A.; Fusetti, L.; Matteoli, B.; Ceccherini-Nelli, L.; Sabbadini, M.G.; Lusso, P.; Parodi, A.; et al. Selective Reactivation of Human Herpesvirus 6 in Patients with Autoimmune Connective Tissue Diseases. J. Med. Virol. 2013, 85, 1925–1934. [Google Scholar] [CrossRef]
- Ferri, C.; Arcangeletti, M.-C.; Caselli, E.; Zakrzewska, K.; Maccari, C.; Calderaro, A.; D’Accolti, M.; Soffritti, I.; Arvia, R.; Sighinolfi, G.; et al. Insights into the Knowledge of Complex Diseases: Environmental Infectious/Toxic Agents as Potential Etiopathogenetic Factors of Systemic Sclerosis. J. Autoimmun. 2021, 124, 102727. [Google Scholar] [CrossRef]
- Caruso, A.; Caselli, E.; Fiorentini, S.; Rotola, A.; Prandini, A.; Garrafa, E.; Saba, E.; Alessandri, G.; Cassai, E.; Di Luca, D. U94 of Human Herpesvirus 6 Inhibits in Vitro Angiogenesis and Lymphangiogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 20446–20451. [Google Scholar] [CrossRef]
- Rizzo, R.; D’Accolti, M.; Bortolotti, D.; Caccuri, F.; Caruso, A.; Di Luca, D.; Caselli, E. Human Herpesvirus 6A and 6B Inhibit in Vitro Angiogenesis by Induction of Human Leukocyte Antigen G. Sci. Rep. 2018, 8, 17683. [Google Scholar] [CrossRef]
- Caselli, E.; Soffritti, I.; D’Accolti, M.; Bortolotti, D.; Rizzo, R.; Sighinolfi, G.; Giuggioli, D.; Ferri, C. HHV-6A Infection and Systemic Sclerosis: Clues of a Possible Association. Microorganisms 2019, 8, 39. [Google Scholar] [CrossRef]
- Arcangeletti, M.-C.; D’Accolti, M.; Maccari, C.; Soffritti, I.; Conto, F.D.; Chezzi, C.; Calderaro, A.; Ferri, C.; Caselli, E. Impact of Human Cytomegalovirus and Human Herpesvirus 6 Infection on the Expression of Factors Associated with Cell Fibrosis and Apoptosis: Clues for Implication in Systemic Sclerosis Development. Int. J. Mol. Sci. 2020, 21, 6397. [Google Scholar] [CrossRef]
- Soffritti, I.; D’Accolti, M.; Ravegnini, G.; Arcangeletti, M.-C.; Maccari, C.; De Conto, F.; Calderaro, A.; Caselli, E. Modulation of microRNome by Human Cytomegalovirus and Human Herpesvirus 6 Infection in Human Dermal Fibroblasts: Possible Significance in the Induction of Fibrosis in Systemic Sclerosis. Cells 2021, 10, 1060. [Google Scholar] [CrossRef]
- Wallace, D.; Gladman, D. Clinical Manifestations and Diagnosis of Systemic Lupus Erythematosus in Adults—UpToDate. Available online: https://www-uptodate-com.db.rsu.lv/contents/clinical-manifestations-and-diagnosis-of-systemic-lupus-erythematosus-in-adults?search=lupus&source=search_result&selectedTitle=1~150&usage_type=default&display_rank=1 (accessed on 24 November 2021).
- Parks, C.G.; de Souza Espindola Santos, A.; Barbhaiya, M.; Costenbader, K.H. Understanding the Role of Environmental Factors in the Development of Systemic Lupus Erythematosus. Best. Pract. Res. Clin. Rheumatol. 2017, 31, 306–320. [Google Scholar] [CrossRef]
- Reis, A.D.; Mudinutti, C.; de Freitas Peigo, M.; Leon, L.L.; Costallat, L.T.L.; Rossi, C.L.; Costa, S.C.B.; Bonon, S.H.A. Active Human Herpesvirus Infections in Adults with Systemic Lupus Erythematosus and Correlation with the SLEDAI Score. Adv. Rheumatol. 2020, 60, 42. [Google Scholar] [CrossRef]
- Chen, X.; Li, H.; Wu, C.; Zhang, Y. Epstein–Barr Virus and Human Herpesvirus 6 Infection in Patients with Systemic Lupus Erythematosus. Virol. J. 2023, 20, 29. [Google Scholar] [CrossRef]
- Lino, K.; Alves, L.S.; Trizzotti, N.; Raposo, J.V.; Souza, C.F.; Silva, A.A.D.; Paula, V.S.D.; Almeida, J.R. Evaluation of Herpesvirus Members on Hospital Admission in Patients with Systemic Lupus Erythematous Shows Higher Frequency of Epstein-Barr Virus and Its Associated Renal Dysfunction. Braz. J. Nephrol. 2022, 44, 490–497. [Google Scholar] [CrossRef]
- Krueger, G.R.; Sander, C.; Hoffmann, A.; Barth, A.; Koch, B.; Braun, M. Isolation of Human Herpesvirus-6 (HHV-6) from Patients with Collagen Vascular Diseases. In Vivo 1991, 5, 217–225. [Google Scholar]
- Rasmussen, N.; Draborg, A.; Nielsen, C.; Jacobsen, S.; Houen, G. Antibodies to Early EBV, CMV, and HHV6 Antigens in Systemic Lupus Erythematosus Patients. Scand. J. Rheumatol. 2015, 44, 143–149. [Google Scholar] [CrossRef][Green Version]
- Scherer, H.U.; Häupl, T.; Burmester, G.R. The Etiology of Rheumatoid Arthritis. J. Autoimmun. 2020, 110, 102400. [Google Scholar] [CrossRef]
- Álvarez-Lafuente, R.; Fernández-Gutiérrez, B.; Miguel, S.d.; Jover, J.A.; Rollin, R.; Loza, E.; Clemente, D.; Lamas, J.R. Potential Relationship between Herpes Viruses and Rheumatoid Arthritis: Analysis with Quantitative Real Time Polymerase Chain Reaction. Ann. Rheum. Dis. 2005, 64, 1357–1359. [Google Scholar] [CrossRef]
- Kadiša, A.; Nora-Krūkle, Z.; Kozireva, S.; Svirskis, S.; Studers, P.; Groma, V.; Lejnieks, A.; Murovska, M. Effect of Human Herpesviruses 6 and 7 Infection on the Clinical Course of Rheumatoid Arthritis/Cilvēka Herpesvīrusa 6 Un 7 Infekcijas Ietekme Uz Reimatoīdā Artrīta Klīnisko Gaitu. Proc. Latv. Acad. Sci. Sect. B. Nat. Exact. Appl. Sci. 2016, 70, 165–174. [Google Scholar] [CrossRef]
- Burgos, R.; Ordoñez, G.; Vázquez-Mellado, J.; Pineda, B.; Sotelo, J. Occasional Presence of Herpes Viruses in Synovial Fluid and Blood from Patients with Rheumatoid Arthritis and Axial Spondyloarthritis. Clin. Rheumatol. 2015, 34, 1681–1686. [Google Scholar] [CrossRef]
- Wen, Z.; Fiocchi, C. Inflammatory Bowel Disease: Autoimmune or Immune-Mediated Pathogenesis? Clin. Dev. Immunol. 2004, 11, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide Incidence and Prevalence of Inflammatory Bowel Disease in the 21st Century: A Systematic Review of Population-Based Studies. Lancet 2017, 390, 2769–2778. [Google Scholar] [CrossRef] [PubMed]
- Tye-Din, J.A.; Galipeau, H.J.; Agardh, D. Celiac Disease: A Review of Current Concepts in Pathogenesis, Prevention, and Novel Therapies. Front. Pediatr. 2018, 6, 350. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Bernstein, C.N. Environmental Risk Factors for Inflammatory Bowel Disease. United Eur. Gastroenterol. J. 2022, 10, 1047–1053. [Google Scholar] [CrossRef] [PubMed]
- Hentrich, M.; Oruzio, D.; Jäger, G.; Schlemmer, M.; Schleuning, M.; Schiel, X.; Hiddemann, W.; Kolb, H.-J. Impact of Human Herpesvirus-6 after Haematopoietic Stem Cell Transplantation. Br. J. Haematol. 2005, 128, 66–72. [Google Scholar] [CrossRef]
- Axelrad, J.E.; Cadwell, K.H.; Colombel, J.-F.; Shah, S.C. The Role of Gastrointestinal Pathogens in Inflammatory Bowel Disease: A Systematic Review. Ther. Adv. Gastroenterol. 2021, 14, 175628482110044. [Google Scholar] [CrossRef] [PubMed]
- Sipponen, T.; Turunen, U.; Lautenschlager, I.; Nieminen, U.; Arola, J.; Halme, L. Human Herpesvirus 6 and Cytomegalovirus in Ileocolonic Mucosa in Inflammatory Bowel Disease. Scand. J. Gastroenterol. 2011, 46, 1324–1333. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Nagata, N.; Okahara, K.; Joya, A.; Hayashida, T.; Oka, S.; Sakurai, T.; Akiyama, J.; Uemura, N.; Gatanaga, H. PCR Detection of Human Herpesviruses in Colonic Mucosa of Individuals with Inflammatory Bowel Disease: Comparison with Individuals with Immunocompetency and HIV Infection. PLoS ONE 2017, 12, e0184699. [Google Scholar] [CrossRef] [PubMed]
- Hosomi, S.; Watanabe, K.; Nishida, Y.; Yamagami, H.; Yukawa, T.; Otani, K.; Nagami, Y.; Tanaka, F.; Taira, K.; Kamata, N.; et al. Combined Infection of Human Herpes Viruses: A Risk Factor for Subsequent Colectomy in Ulcerative Colitis. Inflamm. Bowel Dis. 2018, 24, 1307–1315. [Google Scholar] [CrossRef] [PubMed]
- Nahar, S. Evaluation of a Multiplex PCR Assay for Detection of Cytomegalovirus in Stool Samples from Patients with Ulcerative Colitis. World J. Gastroenterol. 2015, 21, 12667. [Google Scholar] [CrossRef] [PubMed]
- Kemppainen, K.M.; Lynch, K.F.; Liu, E.; Lönnrot, M.; Simell, V.; Briese, T.; Koletzko, S.; Hagopian, W.; Rewers, M.; She, J.-X.; et al. Factors That Increase Risk of Celiac Disease Autoimmunity After a Gastrointestinal Infection in Early Life. Clin. Gastroenterol. Hepatol. 2017, 15, 694–702.e5. [Google Scholar] [CrossRef] [PubMed]
- Calabretto, M.; Di Carlo, D.; Falasca, F.; Mazzuti, L.; Meacci, A.; Donato, G.; Greco, N.; Mezzatesta, L.; Morrone, A.; Turriziani, O.; et al. Analysis of Viral Nucleic Acids in Duodenal Biopsies from Adult Patients with Celiac Disease. Eur. J. Gastroenterol. Hepatol. 2022, 34, 1107–1110. [Google Scholar] [CrossRef] [PubMed]
- Garmendia Madariaga, A.; Santos Palacios, S.; Guillén-Grima, F.; Galofré, J.C. The Incidence and Prevalence of Thyroid Dysfunction in Europe: A Meta-Analysis. J. Clin. Endocrinol. Metab. 2014, 99, 923–931. [Google Scholar] [CrossRef]
- Antonelli, A.; Ferrari, S.M.; Corrado, A.; Di Domenicantonio, A.; Fallahi, P. Autoimmune Thyroid Disorders. Autoimmun. Rev. 2015, 14, 174–180. [Google Scholar] [CrossRef]
- Taylor, P.N.; Albrecht, D.; Scholz, A.; Gutierrez-Buey, G.; Lazarus, J.H.; Dayan, C.M.; Okosieme, O.E. Global Epidemiology of Hyperthyroidism and Hypothyroidism. Nat. Rev. Endocrinol. 2018, 14, 301–316. [Google Scholar] [CrossRef]
- Vargas-Uricoechea, H. Molecular Mechanisms in Autoimmune Thyroid Disease. Cells 2023, 12, 918. [Google Scholar] [CrossRef]
- Desailloud, R.; Hober, D. Viruses and Thyroiditis: An Update. Virol. J. 2009, 6, 5. [Google Scholar] [CrossRef]
- Cuan-Baltazar, Y.; Soto-Vega, E. Microorganisms Associated to Thyroid Autoimmunity. Autoimmun. Rev. 2020, 19, 102614. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.; Liakos, V.; Michou, V.; Kapranos, N.; Kaltsas, G.; Tsilivakos, V.; Tsatsoulis, A. Detection of Herpes Virus DNA in Post-Operative Thyroid Tissue Specimens of Patients with Autoimmune Thyroid Disease. Exp. Clin. Endocrinol. Diabetes 2008, 116, 35–39. [Google Scholar] [CrossRef] [PubMed]
- AL-Zarzour, N.; Monem, F. Are Human Herpes Viruses Associated with Autoimmune Thyroid Disease? J. Infect. Dev. Ctries. 2011, 5, 890–892. [Google Scholar] [CrossRef] [PubMed]
- Darvish Molla, Z.; Kalbasi, S.; Kalantari, S.; Bidari Zerehpoosh, F.; Shayestehpour, M.; Yazdani, S. Evaluation of the Association between Human Herpes Virus 6 (HHV-6) and Hashimoto’s Thyroiditis. Iran. J. Microbiol. 2022, 14, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Caselli, E.; D’Accolti, M.; Soffritti, I.; Zatelli, M.C.; Rossi, R.; Degli Uberti, E.; Di Luca, D. HHV-6A in Vitro Infection of Thyrocytes and T Cells Alters the Expression of miRNA Associated to Autoimmune Thyroiditis. Virol. J. 2017, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Seyyedi, N.; Dehbidi, G.R.; Karimi, M.; Asgari, A.; Esmaeili, B.; Zare, F.; Farhadi, A.; Dabbaghmanesh, M.H.; Saki, F.; Behzad-Behbahani, A. Human Herpesvirus 6A Active Infection in Patients with Autoimmune Hashimoto’s Thyroiditis. Braz. J. Infect. Dis. 2019, 23, 435–440. [Google Scholar] [CrossRef] [PubMed]
- Sultanova, A.; Cistjakovs, M.; Sokolovska, L.; Todorova, K.; Cunskis, E.; Murovska, M. HHV-6 Infection and Chemokine RANTES Signaling Pathway Disturbance in Patients with Autoimmune Thyroiditis. Viruses 2020, 12, 689. [Google Scholar] [CrossRef] [PubMed]
- Sultanova, A.; Cistjakovs, M.; Sokolovska, L.; Cunskis, E.; Murovska, M. Investigation of the Involvement of HHV-6 Encoded Viral Chemokine Receptors in Autoimmune Thyroiditis Development. Microbiol. Spectr. 2022, 10, e02369-21. [Google Scholar] [CrossRef] [PubMed]
- Ericsson, M.; Skog, O. Presence of Human Herpesvirus 6B in the Pancreas of Subjects with and without Type 1 Diabetes. Pancreas 2017, 46, 1341–1346. [Google Scholar] [CrossRef] [PubMed]
- Sabouri, S.; Benkahla, M.A.; Kiosses, W.B.; Rodriguez-Calvo, T.; Zapardiel-Gonzalo, J.; Castillo, E.; Von Herrath, M.G. Human Herpesvirus-6 Is Present at Higher Levels in the Pancreatic Tissues of Donors with Type 1 Diabetes. J. Autoimmun. 2020, 107, 102378. [Google Scholar] [CrossRef] [PubMed]
- Sotzny, F.; Blanco, J.; Capelli, E.; Castro-Marrero, J.; Steiner, S.; Murovska, M.; Scheibenbogen, C. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome—Evidence for an Autoimmune Disease. Autoimmun. Rev. 2018, 17, 601–609. [Google Scholar] [CrossRef] [PubMed]
- The European Network on ME/CFS (EUROMENE); Rasa, S.; Nora-Krukle, Z.; Henning, N.; Eliassen, E.; Shikova, E.; Harrer, T.; Scheibenbogen, C.; Murovska, M.; Prusty, B.K. Chronic Viral Infections in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS). J. Transl. Med. 2018, 16, 268. [Google Scholar] [CrossRef] [PubMed]
- Komaroff, A.L. Chronic Fatigue Syndromes: Relationship to Chronic Viral Infections. J. Virol. Methods 1988, 21, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Koelle, D.M.; Barcy, S.; Huang, M.; Ashley, R.L.; Corey, L.; Zeh, J.; Ashton, S.; Buchwald, D. Markers of Viral Infection in Monozygotic Twins Discordant for Chronic Fatigue Syndrome. Clin. Infect. Dis. 2002, 35, 518–525. [Google Scholar] [CrossRef]
- Cameron, B.; Flamand, L.; Juwana, H.; Middeldorp, J.; Naing, Z.; Rawlinson, W.; Ablashi, D.; Lloyd, A. Serological and Virological Investigation of the Role of the Herpesviruses EBV, CMV and HHV-6 in Post-infective Fatigue Syndrome. J. Med. Virol. 2010, 82, 1684–1688. [Google Scholar] [CrossRef]
- Oakes, B.; Hoagland-Henefield, M.; Komaroff, A.L.; Erickson, J.L.; Huber, B.T. Human Endogenous Retrovirus-K18 Superantigen Expression and Human Herpesvirus-6 and Human Herpesvirus-7 Viral Loads in Chronic Fatigue Patients. Clin. Infect. Dis. 2013, 56, 1394–1400. [Google Scholar] [CrossRef][Green Version]
- Watt, T.; Oberfoell, S.; Balise, R.; Lunn, M.R.; Kar, A.K.; Merrihew, L.; Bhangoo, M.S.; Montoya, J.G. Response to Valganciclovir in Chronic Fatigue Syndrome Patients with Human Herpesvirus 6 and Epstein–Barr Virus IgG Antibody Titers. J. Med. Virol. 2012, 84, 1967–1974. [Google Scholar] [CrossRef]
- Montoya, J.G.; Kogelnik, A.M.; Bhangoo, M.; Lunn, M.R.; Flamand, L.; Merrihew, L.E.; Watt, T.; Kubo, J.T.; Paik, J.; Desai, M. Randomized Clinical Trial to Evaluate the Efficacy and Safety of Valganciclovir in a Subset of Patients with Chronic Fatigue Syndrome. J. Med. Virol. 2013, 85, 2101–2109. [Google Scholar] [CrossRef]
- Chapenko, S.; Krumina, A.; Logina, I.; Rasa, S.; Chistjakovs, M.; Sultanova, A.; Viksna, L.; Murovska, M. Association of Active Human Herpesvirus-6, -7 and Parvovirus B19 Infection with Clinical Outcomes in Patients with Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Adv. Virol. 2012, 2012, 205085. [Google Scholar] [CrossRef]
- Gravelsina, S.; Vilmane, A.; Svirskis, S.; Rasa-Dzelzkaleja, S.; Nora-Krukle, Z.; Vecvagare, K.; Krumina, A.; Leineman, I.; Shoenfeld, Y.; Murovska, M. Biomarkers in the Diagnostic Algorithm of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Front. Immunol. 2022, 13, 928945. [Google Scholar] [CrossRef]
- Kasimir, F.; Toomey, D.; Liu, Z.; Kaiping, A.C.; Ariza, M.E.; Prusty, B.K. Tissue Specific Signature of HHV-6 Infection in ME/CFS. Front. Mol. Biosci. 2022, 9, 1044964. [Google Scholar] [CrossRef] [PubMed]
- Ariza, M.E.; Glaser, R.; Williams, M.V. Human Herpesviruses-Encoded dUTPases: A Family of Proteins That Modulate Dendritic Cell Function and Innate Immunity. Front. Microbiol. 2014, 5, 504. [Google Scholar] [CrossRef] [PubMed]
- Halpin, P.; Williams, M.V.; Klimas, N.G.; Fletcher, M.A.; Barnes, Z.; Ariza, M.E. Myalgic Encephalomyelitis/Chronic Fatigue Syndrome and Gulf War Illness Patients Exhibit Increased Humoral Responses to the Herpesviruses-encoded dUTPase: Implications in Disease Pathophysiology. J. Med. Virol. 2017, 89, 1636–1645. [Google Scholar] [CrossRef] [PubMed]
- Cox, B.S.; Alharshawi, K.; Mena-Palomo, I.; Lafuse, W.P.; Ariza, M.E. EBV/HHV-6A dUTPases Contribute to Myalgic Encephalomyelitis/Chronic Fatigue Syndrome Pathophysiology by Enhancing TFH Cell Differentiation and Extrafollicular Activities. JCI Insight 2022, 7, e158193. [Google Scholar] [CrossRef] [PubMed]
- Casanova, A.; Wevers, A.; Navarro-Ledesma, S.; Pruimboom, L. Mitochondria: It Is All about Energy. Front. Physiol. 2023, 14, 1114231. [Google Scholar] [CrossRef] [PubMed]
- Schreiner, P.; Harrer, T.; Scheibenbogen, C.; Lamer, S.; Schlosser, A.; Naviaux, R.K.; Prusty, B.K. Human Herpesvirus-6 Reactivation, Mitochondrial Fragmentation, and the Coordination of Antiviral and Metabolic Phenotypes in Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. ImmunoHorizons 2020, 4, 201–215. [Google Scholar] [CrossRef]
- Clauw, D.; Sarzi-Puttini, P.; Pellegrino, G.; Shoenfeld, Y. Is Fibromyalgia an Autoimmune Disorder? Autoimmun. Rev. 2023, 103424. [Google Scholar] [CrossRef]
- Krumina, A.; Chapenko, S.; Kenina, V.; Mihailova, M.; Logina, I.; Rasa, S.; Gintere, S.; Viksna, L.; Svirskis, S.; Murovska, M. The Role of HHV-6 and HHV-7 Infections in the Development of Fibromyalgia. J. Neurovirol. 2019, 25, 194–207. [Google Scholar] [CrossRef]
- Fialho, M.F.P.; Brum, E.S.; Oliveira, S.M. Could the Fibromyalgia Syndrome Be Triggered or Enhanced by COVID-19? Inflammopharmacology 2023, 31, 633–651. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.-T.; Yang, C.-W.; Chu, C.-Y. Drug Reaction with Eosinophilia and Systemic Symptoms (DRESS): An Interplay among Drugs, Viruses, and Immune System. Int. J. Mol. Sci. 2017, 18, 1243. [Google Scholar] [CrossRef] [PubMed]
- Hama, N.; Abe, R.; Gibson, A.; Phillips, E.J. Drug-Induced Hypersensitivity Syndrome (DIHS)/Drug Reaction with Eosinophilia and Systemic Symptoms (DRESS): Clinical Features and Pathogenesis. J. Allergy Clin. Immunol. Pract. 2022, 10, 1155–1167.e5. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Ren, V. Immunopathogenic Insights on Preferential Human Herpesvirus-6 Reactivation in Drug Rash with Eosinophilia and Systemic Symptoms: A Scoping Review. J. Cutan. Med. Surg. 2023, 27, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Pritchett, J.C.; Nanau, R.M.; Neuman, M.G. The Link between Hypersensitivity Syndrome Reaction Development and Human Herpes Virus-6 Reactivation. Int. J. Hepatol. 2012, 2012, e723062. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yang, F.; Chen, S.; Xiong, H.; Zhu, Q.; Gao, X.; Xing, Q.; Luo, X. Clinical, Viral and Genetic Characteristics of Drug Reaction with Eosinophilia and Systemic Symptoms (DRESS) in Shanghai, China. Acta Derm. Venereol. 2018, 98, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Funck-Brentano, E.; Duong, T.; Family, D.; Bouaziz, J.-D.; Ortonne, N.; Bagot, M.; Roujeau, J.-C.; Wolkenstein, P.; Valeyrie-Allanore, L. Auto-immune thyroiditis and drug reaction with eosinophilia and systemic symptoms (DRESS) associated with HHV-6 viral reactivation. Ann. Dermatol. Venereol. 2011, 138, 580–585. [Google Scholar] [CrossRef]
- Sato, M.; Mizuno, Y.; Matsuyama, K.; Shu, E.; Kanoh, H.; Suwa, T.; Seishima, M. Drug-Induced Hypersensitivity Syndrome Followed by Subacute Thyroiditis. Case Rep. Dermatol. 2015, 7, 161–165. [Google Scholar] [CrossRef]
- Descamps, V. Drug Reaction with Eosinophilia and Systemic Symptoms and Thyroiditis: Human Herpesvirus-6, the Possible Common Link. Br. J. Dermatol. 2013, 169, 952. [Google Scholar] [CrossRef]
- Descamps, V.; Ranger-Rogez, S. DRESS Syndrome. Jt. Bone Spine 2014, 81, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Ehrenfeld, M.; Tincani, A.; Andreoli, L.; Cattalini, M.; Greenbaum, A.; Kanduc, D.; Alijotas-Reig, J.; Zinserling, V.; Semenova, N.; Amital, H.; et al. COVID-19 and Autoimmunity. Autoimmun. Rev. 2020, 19, 102597. [Google Scholar] [CrossRef] [PubMed]
- Leveque, N.; Boulagnon, C.; Brasselet, C.; Lesaffre, F.; Boutolleau, D.; Metz, D.; Fornes, P.; Andreoletti, L. A Fatal Case of Human Herpesvirus 6 Chronic Myocarditis in an Immunocompetent Adult. J. Clin. Virol. 2011, 52, 142–145. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.; Renaud, C.; Kuypers, J.M.; Chiu, C.Y.; Huang, M.-L.; Samayoa, E.; Xie, H.; Yu, G.; Fisher, C.E.; Gooley, T.A.; et al. Idiopathic Pneumonia Syndrome after Hematopoietic Cell Transplantation: Evidence of Occult Infectious Etiologies. Blood 2015, 125, 3789–3797. [Google Scholar] [CrossRef] [PubMed]
- Skiba, M.A.; Kruse, A.C. Autoantibodies as Endogenous Modulators of GPCR Signaling. Trends Pharmacol. Sci. 2021, 42, 135–150. [Google Scholar] [CrossRef]
- Shafiee, A.; Teymouri Athar, M.M.; Amini, M.J.; Hajishah, H.; Siahvoshi, S.; Jalali, M.; Jahanbakhshi, B.; Mozhgani, S. Reactivation of Herpesviruses during COVID-19: A Systematic Review and Meta-Analysis. Rev. Med. Virol. 2023, 33, e2437. [Google Scholar] [CrossRef]
- Vojdani, A.; Vojdani, E.; Saidara, E.; Maes, M. Persistent SARS-CoV-2 Infection, EBV, HHV-6 and Other Factors May Contribute to Inflammation and Autoimmunity in Long COVID. Viruses 2023, 15, 400. [Google Scholar] [CrossRef]


{kind=link}
 | ↑ HHV-6 infection markers in plaques [39,40,41,42,43,44,45,46] |
| ↑ HHV-6 infection markers in CSF, sera, PBMC [49,50,51,52,53,54,55] | |
| ↑ HHV-6 load → Exacerbation, Relapse [56,57,58,59] | |
| ↑ HHV-6 IgM, IgG [54,60,61,62,63] | |
| ↓ HHV-6 IgG → ↓ Relapse episodes [64] | |
| ↑ HHV-6A IE1 IgG → ↑ MS risk [65] | |
| ↑ TNF-α, IFN-γ, IL-1β, IL-6, IL-12, CCL-5 in HHV-6 Ig (+) [59,66] | |
| Molecular mimicry HHV-6 U24 and MBP [9,10] | |
| ↑ Cross-reactive U24/MBP T cells and antibodies [9,10] | |
| U24 and Nedd4 → Demyelination [70] | |
| HHV-6 transactivates EBV [67,68,69] |
 | ↑ HHV-6A and B infection markers in thyroid, sera [7,8,122,123] |
| ↑ HHV-6A and B active infection in thyroid [7,8,123] | |
| Thyrocytes permissive to HHV-6 [7,121] | |
| ↑ HHV-6 markers in thyrocytes [7] | |
| HHV-6 in thyrocytes → ↑ HLA-II expression [7] | |
| HHV-6 in thyrocytes → ↑ NK-mediated killing [7] | |
| ↑ HHV-6 specific T cells [7] | |
| HHV-6 in thyrocytes → ↑ AITD miRNAs [121] | |
| ↓ RANTES (CCL-5) and HHV-6 vGPCRs [123] |
 | ↑ HHV-6 infection markers in sera, PBMCs [128,135,136] |
| HHV-6 active infection and Clinical symptoms [135] | |
| ↑ HHV-6 load in sera → ↑ Disease severity [136] | |
| ↑ HHV-6 load in sera → ↑ anti-M4 Abs [136] | |
| ↑ HHV-6 active infection in brain [137] | |
| ↑ HHV dUTPase antibodies [139] | |
| HHV dUTPase antibodies → ↑ activin A, IL-21 [140] | |
| HHV-6A reactivation → Mitochondrial fragmentation [141] | |
| HHV-6A reactivation → ↓ AA, FA, glucose metabolism [142] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sokolovska, L.; Cistjakovs, M.; Matroze, A.; Murovska, M.; Sultanova, A. From Viral Infection to Autoimmune Reaction: Exploring the Link between Human Herpesvirus 6 and Autoimmune Diseases. Microorganisms 2024, 12, 362. https://doi.org/10.3390/microorganisms12020362
Sokolovska L, Cistjakovs M, Matroze A, Murovska M, Sultanova A. From Viral Infection to Autoimmune Reaction: Exploring the Link between Human Herpesvirus 6 and Autoimmune Diseases. Microorganisms. 2024; 12(2):362. https://doi.org/10.3390/microorganisms12020362
Chicago/Turabian StyleSokolovska, Liba, Maksims Cistjakovs, Asnate Matroze, Modra Murovska, and Alina Sultanova. 2024. "From Viral Infection to Autoimmune Reaction: Exploring the Link between Human Herpesvirus 6 and Autoimmune Diseases" Microorganisms 12, no. 2: 362. https://doi.org/10.3390/microorganisms12020362
APA StyleSokolovska, L., Cistjakovs, M., Matroze, A., Murovska, M., & Sultanova, A. (2024). From Viral Infection to Autoimmune Reaction: Exploring the Link between Human Herpesvirus 6 and Autoimmune Diseases. Microorganisms, 12(2), 362. https://doi.org/10.3390/microorganisms12020362