
Environmental Drivers of the Divergence of Harveyi Clade Pathogens with Distinctive Virulence Gene Profiles


Abstract
1. Introduction
2. Materials and Methods
2.1. Field Sites and Sampling Events
2.2. Sample Collection and Processing
2.3. Measurement of Environmental Parameters
2.4. Genes Targeted and Generation of Standard Curves
2.5. Statistics
3. Results
3.1. The Presence and Seasonal Distribution of Virulence Genes
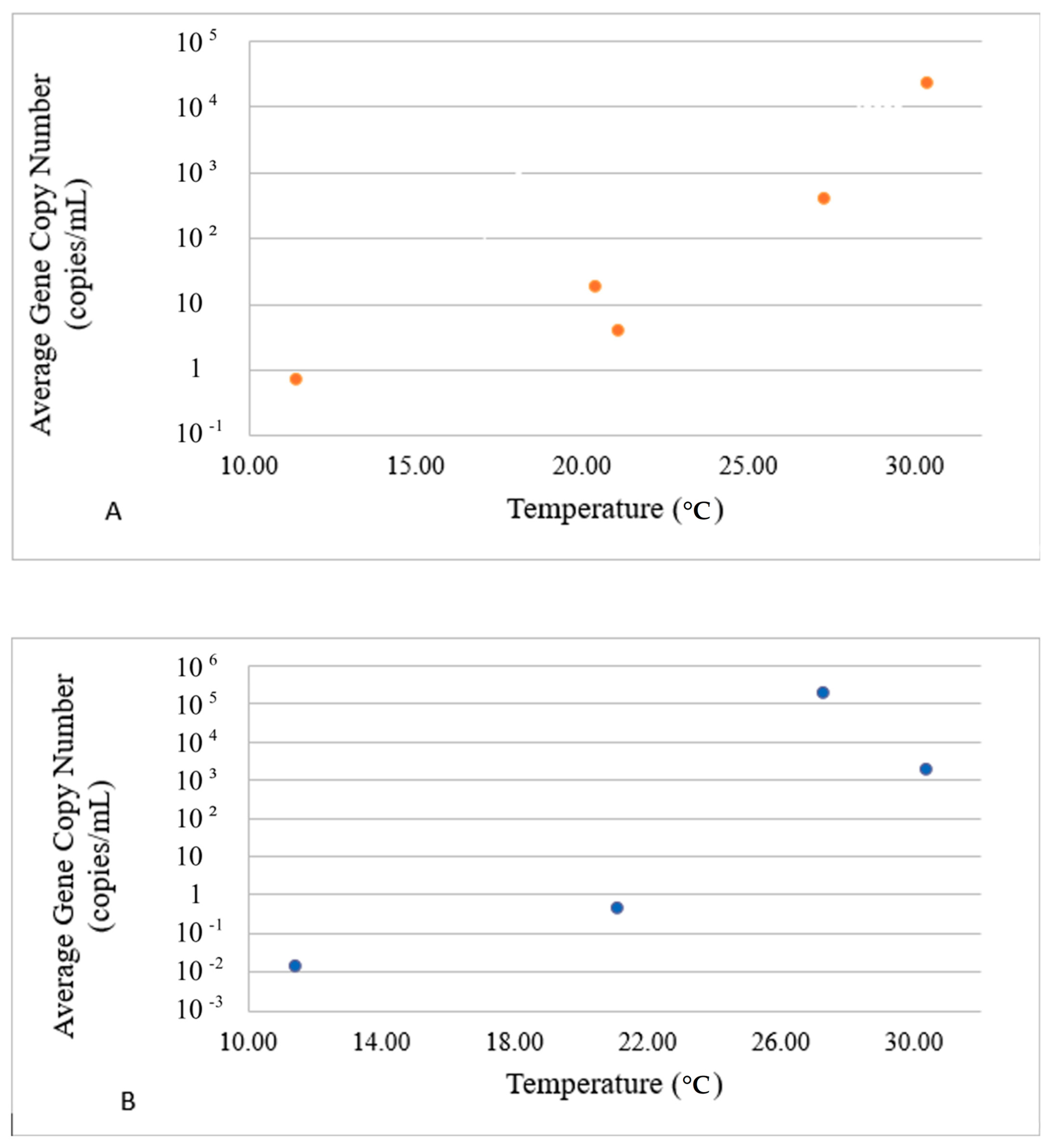
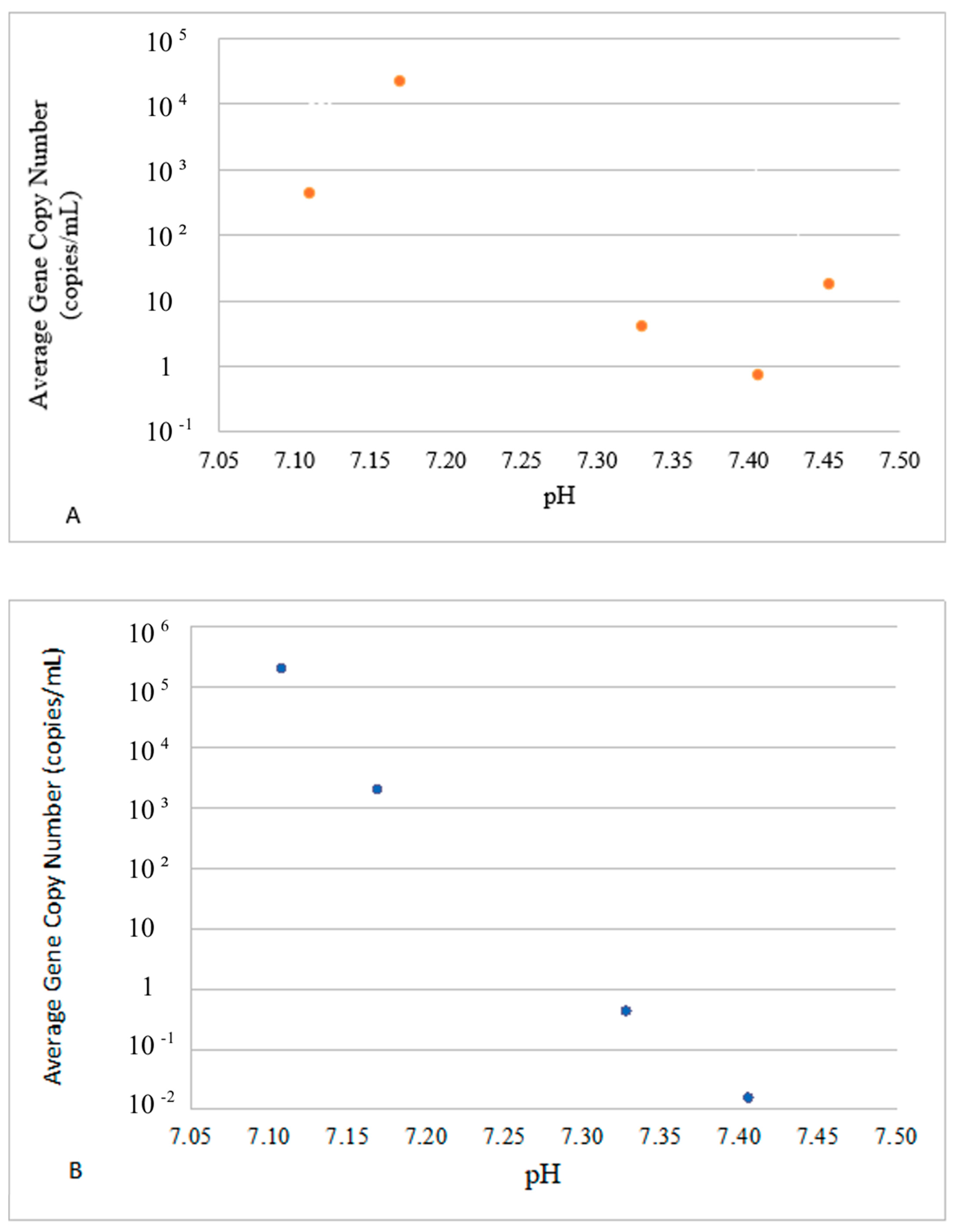
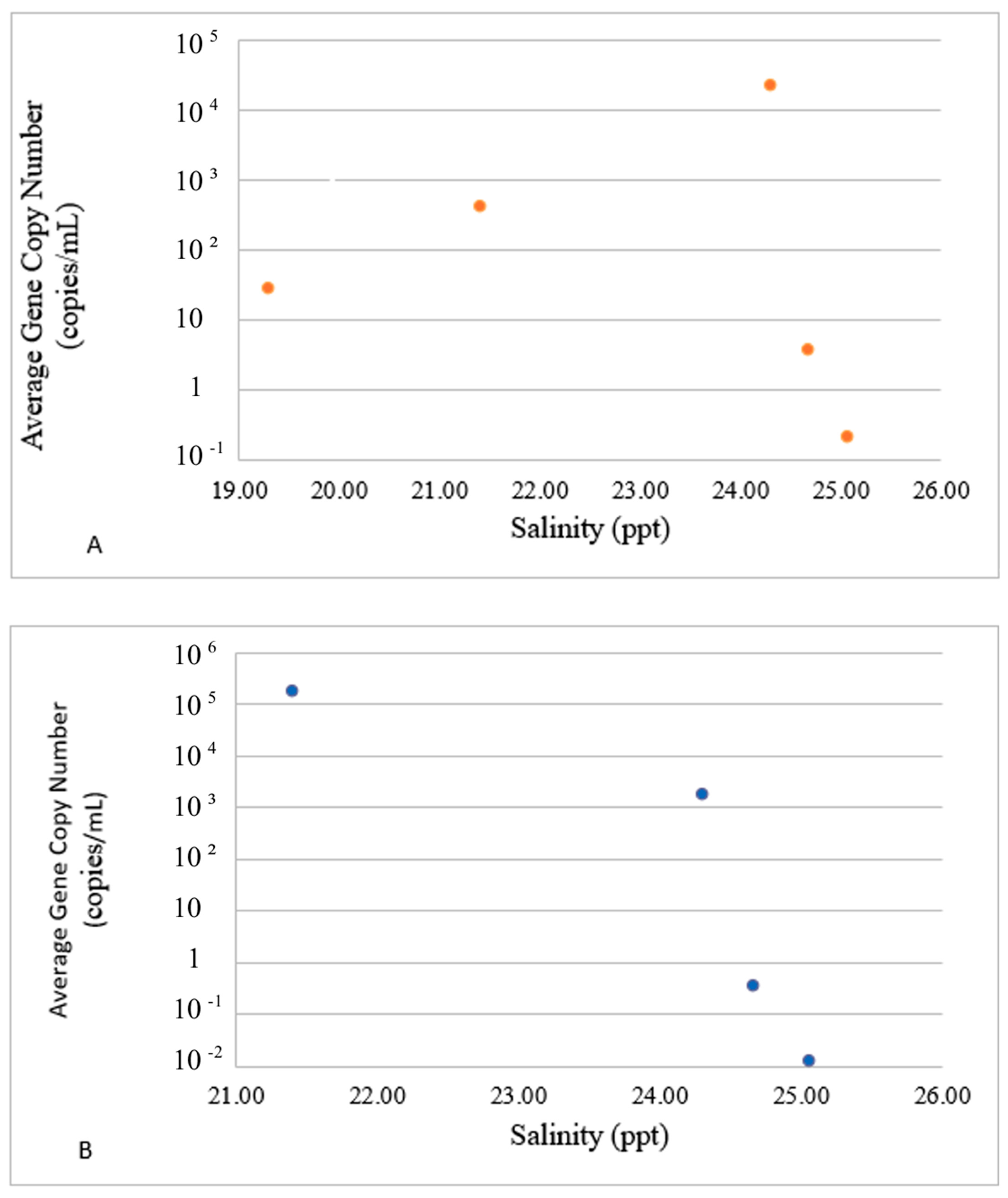
3.2. Divergence of VG Carriers in Response to Water Parameters
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, X.H.; He, X.; Austin, B. Vibrio harveyi: A serious pathogen of fish and invertebrates in mariculture. Mar. Life Sci. Technol. 2020, 2, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Culot, A.; Grosset, N.; Bruey, Q.; Auzou, M.; Giard, J.C.; Favard, B.; Wakatsuki, A.; Baron, S.; Frouel, S.; Teacher, C.; et al. Isolation of Harveyi clade Vibrio spp. collected in aquaculture farms: How can the identification be addressed? J. Microbiol. Methods 2021, 180, 106106. [Google Scholar] [CrossRef] [PubMed]
- Urbanczyk, H.; Ogura, Y.; Hayashi, T. Taxonomic revision of Harveyi clade bacteria (family Vibrionaceae) based on analysis of whole genome sequences. Int. J. Syst. Evol. Microbiol. 2013, 63 Pt 7, 2742–2751. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Trinanes, J.; Taylor, N.G.H.; Hartnell, R. Emerging Vibrio risk at high latitudes in response to ocean warming. Nat. Rep. Clim. Chang. 2012, 3, 73–77. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Trinanes, J.; Gonzalez-Escalona, N.; Martinez-Urtaza, J. Non-Cholera Vibrios: The microbial barometer of climate change. Trends Microbiol. 2017, 25, 76–84. [Google Scholar] [CrossRef]
- Vezzulli, L.; Colwell, R.R.; Pruzzo, C. Ocean warming and the spread of pathogenic Vibrios in the aquatic environment. Microbial. Ecol. 2013, 65, 817–825. [Google Scholar] [CrossRef]
- Brehm, T.T.; Berneking, L.; Rohde, H.; Chistner, M.; Schlickewei, C.; Sena Martins, M.; Schmiedel, S. Wound infection with Vibrio harveyi following a traumatic leg amputation after a motorboat propeller injury in Mallorca, Spain: A case report and review of literature. BMC Infect. Dis. 2020, 20, 104. [Google Scholar] [CrossRef]
- Vezzulli, L.; Grande, C.; Reid, P.C.; Helaouet, P.; Edwards, M.; Hofle, M.G.; Brettar, I.; Colwell, R.R.; Pruzzo, C. Climate influence on Vibrio and associated human diseases during the past half-century in the coastal North Atlantic. Proc. Natl. Acad. Sci. USA 2016, 113, 5062–5071. [Google Scholar] [CrossRef]
- Fleischmann, S.; Herrig, I.; Wesp, J.; Stiedl, J.; Reifferscheid, G.; Strauch, E.; Alter, T.; Brennholt, N. Prevalence and distribution of potentially human pathogenic Vibrio spp. on German North and Baltic Sea coasts. Front. Cell. Infect. Microbiol. 2022, 12, 846819. [Google Scholar] [CrossRef]
- Montánchez, I.; Kaberdin, V.R. Vibrio harveyi: A brief survey of general characteristics and recent epidemiological traits associated with climate change. Mar. Environ. Res. 2020, 154, 104850. [Google Scholar] [CrossRef]
- Johnson, C.N.; Flowers, A.R.; Noriea, N.F.; Zimmerman, A.M.; Bowers, J.C.; DePaola, A.; Grimes, D.J. Relationships between environmental factors and pathogenic vibrios in the northern Gulf of Mexico. Appl. Environ. Microb. 2010, 76, 7076–7084. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Cabanyero, C.; Sanjuan, E.; Fouz, B.; Pajuelo, D.; Vallejos-Vidal, E.; Reyes-Lopez, F.E.; Amaro, C. The effect of the environmental temperature on the adaptation to host in the zoonotic pathogen Vibrio vulnificus. Front. Microbiol. 2020, 11, 489. [Google Scholar] [CrossRef] [PubMed]
- Luna, G.M.; Bongiorni, L.; Gili, C.; Biavasco, F.; Danovaro, R. Vibrio harveyi as a causative agent of the white syndrome in tropical stony corals. Environ. Microbiol. Rep. 2010, 2, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Kloska, A.; Cech, G.M.; Nowicki, D.; Maciąg-Dorszyńska, M.; Bogucka, A.E.; Markert, S.; Becher, D.; Potrykus, K.; Czaplewska, P.; Szalewska-Pałasz, A. Three Microbial Musketeers of the Seas: Shewanella baltica, Aliivibrio fischeri and Vibrio harveyi, and Their Adaptation to Different Salinity Probed by a Proteomic Approach. Int. J. Mol. Sci. 2022, 23, 619. [Google Scholar] [CrossRef]
- Zhang, W.; Chen, K.; Zhang, L.; Zhang, X.; Zhu, B.; Lv, N.; Mi, K. The impact of global warming on the signature virulence gene, thermolabile hemolysin, of Vibrio parahaemolyticus. Microbiol. Spectr. 2023, 11, e01502–e01523. [Google Scholar] [CrossRef]
- Allen, C.; Finkel, S.E. Vibrio harveyi exhibits the growth advantage in stationary phase phenotype during long-term incubation. Microbiol. Spectr. 2022, 10, e02144-21. [Google Scholar] [CrossRef]
- Blackwell, K.D.; Oliver, J.D. The ecology of Vibrio vulnificus, Vibrio cholerae, and Vibrio parahaemolyticus in North Carolina estuaries. J. Microbiol. 2008, 46, 146–153. [Google Scholar] [CrossRef]
- Prescott, J.; Barkovskii, A.L. In situ dynamics of Vibrio parahaemolyticus and Vibrio vulnificus in water, sediment, and triploid Crassostrea virginica oysters cultivated in floating gear. J. Appl. Microbiol. 2021, 132, 3343–3354. [Google Scholar] [CrossRef]
- Prayitno, S.B.; Latchford, J.W. Experimental infections of crustaceans with luminous bacteria related to Photobacterium and Vibrio. Effect of salinity and pH on infectiosity. Aquaculture 1995, 132, 105–112. [Google Scholar] [CrossRef]
- Wang, Q.; Fu, S.; Yang, Q.; Hao, J.; Zhou, C.; Liu, Y. The impact of water intrusion on pathogenic Vibrio species to inland brackish waters of China. Int. J. Environ. Res. Public Health 2020, 17, 6781. [Google Scholar] [CrossRef]
- Gundogdu, K.; Iturriza, A.O.; Orruno, M.; Montanchez, I.; Eguiraun, H.; Martinez, I.; Arana, I.; Kaberdin, V.R. Addressing the joint impact of temperature and pH on Vibrio harveyi adaptation in the time of climate change. Microorganisms 2023, 11, 1075. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, W.B.; Parent, M.A.; Naughton, L.M.; Richards, G.P.; Blumerman, S.L.; Boyd, E.F. Modulation of responses of Vibrio parahaemolyticus O3: K6 to pH and temperature stresses by growth at different salt concentrations. Appl. Environ. Microbiol. 2010, 76, 4720–4729. [Google Scholar] [CrossRef] [PubMed]
- Nhu, N.T.Q.; Lee, J.S.; Wang, H.J.; Dufour, Y.S. Alkaline pH increases swimming speed and facilitates mucus penetration for Vibrio cholerae. J. Bacteriol. 2021, 203, 607–620. [Google Scholar] [CrossRef] [PubMed]
- Ke, H.M.; Liu, D.; Ogura, Y.; Hayashi, T.; Urbanczyk, H.; Tsai, I.J. Tracing genomic divergence of Vibrio bacteria in the Harveyi clade. J. Bacteriol. 2018, 200, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Orlić, K.; Kapetanović, D.; Kazazić, S.; Smrzlić, I.V.; Barac, F.; Nerlović, V.; Buha, T.; Bolotin, J.; Kožul, V.; Bobanović-Ćolić, S.; et al. Diversity, virulence and antibiotic resistance of Vibrio Harveyi clade species associated with bivalve aquaculture within marine protected areas. Aquaculture 2025, 594, 741392. [Google Scholar] [CrossRef]
- Li, L.; Lu, J.; Zhan, P.; Qiu, Q.; Chen, J.; Xiong, J. RNA-seq analysis unveils temperature and nutrient adaptation mechanisms relevant for pathogenicity in Vibrio parahaemolyticus. Aquaculture 2022, 558, 738397. [Google Scholar] [CrossRef]
- Gu, D.; Guo, M.; Yang, M.; Zhang, Y.; Zhou, X.; Wang, Q. A σE-mediated temperature gauge controls a switch from LuxR-mediated virulence gene expression to thermal stress adaptation in Vibrio alginolyticus. PLOS Pathog. 2016, 12, 1005645. [Google Scholar] [CrossRef]
- Ruwandeepika, H.A.D.; Defoirdt, T.; Bhowmick, P.P.; Karunasagar, I.; Bossier, P. In vitro and in vivo expression of virulence genes in Vibrio isolates belonging to the Harveyi clade in relation to their virulence towards gnotobiotic brine shrimp (Artemia franciscana). Environ. Microbiol. 2011, 12, 506–517. [Google Scholar] [CrossRef]
- Chandran, A.; Varghese, S.; Kandeler, E.; Thomas, A.; Hatha, M.; Mazumder, A. An assessment of potential public health risk associated with the extended survival of indicator and pathogenic bacteria in freshwater lake sediments. Int. J. Hyg. Environ. Health 2011, 214, 258–264. [Google Scholar] [CrossRef]
- Hassard, F.; Gwyther, C.L.; Farkas, K.; Andrews, A.; Jones, V.; Cox, B.; Malham, S.K. Abundance and distribution of enteric bacteria and viruses in coastal and estuarine sediments—A review. Front. Microbiol. 2016, 7, 1692. [Google Scholar] [CrossRef]
- Jones, C.E.; Barkovskii, A.L. Impact of roadside ditch dredging on bacterial communities and biological contamination of a tidal creek. Estuar. Coast. Shelf Sci. 2017, 188, 137–144. [Google Scholar] [CrossRef]
- Böer, S.I.; Heinemeyer, E.A.; Luden, K.; Erler, R.; Gerdts, G.; Janssen, F.; Brennholt, N. Temporal and spatial distribution patterns of potentially pathogenic Vibrio spp. at recreational beaches of the German North Sea. Microb. Ecol. 2013, 65, 1052–1067. [Google Scholar] [CrossRef] [PubMed]
- Di, D.Y.; Lee, A.; Jang, J.; Han, D.; Hur, H.G. Season-specific occurrence of potentially pathogenic Vibrio spp. on the southern coast of South Korea. Appl. Environ. Microbiol. 2017, 83, e02680-16. [Google Scholar] [CrossRef]
- Sheikh, H.I.; Najiah, M.; Fadhlina, A.; Laith, A.A.; Nor, M.M.; Jalal, K.C.A.; Kasan, N.A. Temperature upshift mostly but not always enhances the growth of Vibrio species: A systematic review. Front. Mar. Sci. 2022, 9, 959830. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. N.S. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef]
- Velez, K.C.; Leighton, R.E.; Decho, A.W.; Pinckney, J.L.; Norman, R.S. Modeling pH and temperature effects as climatic hazards in Vibrio vulnificus and Vibrio parahaemolyticus planktonic growth and biofilm formation. GeoHealth 2023, 7, e2022GH000769. [Google Scholar] [CrossRef]
- Kunkle, D.E.; Bina, X.R.; Bina, J.E. Vibrio cholerae OmpR contributes to virulence repression and fitness at alkaline pH. Infect. Immun. 2020, 88, 10–1128. [Google Scholar] [CrossRef]
- Huang, W.S.; Wong, H.C. Characterization of low salinity stress in Vibrio parahaemolyticus. J. Food Prot. 2012, 75, 231–237. [Google Scholar] [CrossRef]
- Joseph, A.; Philip, R. Immunocompetence of Penaeus monodon under acute salinity stress and pathogenicity of Vibrio harveyi with respect to ambient salinity. Fish Shellfish. Immunol. 2020, 106, 555–562. [Google Scholar] [CrossRef]
- Randa, M.A.; Polz, M.F.; Lim, E. Effects of temperature and salinity on Vibrio vulnificus population dynamics as assessed by quantitative PCR. Appl. Environ. Microbiol. 2004, 70, 5469–5476. [Google Scholar] [CrossRef]
- Parveen, S.; Hettiarachchi, K.A.; Bowers, J.C.; Jones, J.L.; Tamplin, M.L.; McKay, R.; Beatty, W.; Brohawn, K.; DaSilva, L.V.; DePaola, A. Seasonal distribution of total and pathogenic Vibrio parahaemolyticus in Chesapeake Bay oysters and waters. Int. J. Food Microbiol. 2008, 128, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Scro, A.K.; Westphalen, J.; Kite-Powell, H.L.; Brawley, J.W.; Smolowitz, R.M. The effect of off-bottom versus on-bottom oyster culture on total and pathogenic Vibrio spp. abundances in oyster tissue, water and sediment samples. Int. J. Food Microbiol. 2022, 379, 109870. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Amplicon Size (bp) | Reference |
|---|---|---|---|
| luxR | FW: TCAATTGCAAAGAGACCTCG RV: AGCAAACACTTCAAGAGCGA | 84 | [28] |
| toxR | FW: CGACAACCCAAAATACGGAA RV: AGAGCAATTTGCTGAAGCTA | 131 | [28] |
| srp | FW: TGCACGACCAGTTGCTTTAG RV: AAGTGGTCGTCAGCAAATCC | 232 | [28] |
| vhh | FW: TTCACGCTTGATGGCTACTG RV: GTCACCCAATGCTACGACCT | 234 | [28] |
| vhha | FW: GCGCTTGGTATCTTCTCTGA RV: CAGACAGCTCATCACGCATT | 226 | [28] |
| vhp | FW: CTGAACGACGCCCATTATTT RV: CGCTGACACATCAAGGCTAA | 201 | [28] |
| Site/Gene | toxR | luxR | srp | vhha | vhp | vhh |
|---|---|---|---|---|---|---|
| Site 1 | ||||||
| June Sediment | + | + | + | + | + | + |
| August Sediment | - | + | + | + | + | + |
| October Sediment | + | - | + | + | + | - |
| December Sediment | - | + | + | + | + | + |
| February Sediment | + | + | + | + | + | + |
| June Water | + | + | + | + | + | + |
| August Water | - | + | - | + | + | + |
| October Water | - | + | + | + | + | + |
| December Water | - | + | - | - | + | - |
| February Water | - | + | - | - | + | + |
| Site 2 | ||||||
| June Sediment | + | - | + | + | + | - |
| August Sediment | + | + | + | + | + | + |
| October Sediment | + | + | + | + | + | + |
| December Sediment | - | - | + | + | - | + |
| February Sediment | - | + | + | + | + | + |
| June Water | + | + | + | + | + | + |
| August Water | + | - | + | + | + | + |
| October Water | - | + | + | + | - | + |
| December Water | - | + | + | - | - | - |
| February Water | - | + | - | - | + | - |
| Site 3 | ||||||
| June Sediment | - | + | + | + | - | + |
| August Sediment | + | + | + | + | + | + |
| October Sediment | + | + | + | + | + | + |
| December Sediment | + | + | + | + | + | + |
| February Sediment | + | + | + | + | + | + |
| June Water | + | + | + | + | + | + |
| August Water | + | + | + | + | + | + |
| October Water | - | + | + | - | + | + |
| December Water | - | - | + | - | + | - |
| February Water | - | - | - | + | + | + |
| Detection Frequency | 15/30 | 24/30 | 25/30 | 24/30 | 26/30 | 24/30 |
| Sediment Total | 10/15 | 12/15 | 15/15 | 15/15 | 13/15 | 13/15 |
| Water Total | 5/15 | 12/15 | 10/15 | 9/15 | 13/15 | 11/15 |
| Water Parameters | pH | Temp. (°C) | Salinity (ppt) | Turbidity (NTU) | DO (mg/L) | Conduct. (mS/cm) | TDS (g/L) | Potential Water Density |
|---|---|---|---|---|---|---|---|---|
| June | ||||||||
| Site 1 | 7.10 | 27.63 | 18.3 | 23.6 | 1.24 | 31.5 | 18.1 | 10.2 |
| Site 2 | 7.23 | 26.33 | 22.4 | 21.1 | 1.18 | 35.7 | 22.9 | 11.4 |
| Site 3 | 7.00 | 28.05 | 23.5 | 22.5 | 1.26 | 38.3 | 23.4 | 13.6 |
| Average | 7.11 | 27.34 | 21.40 | 22.40 | 1.23 | 35.17 | 21.47 | 11.73 |
| August | ||||||||
| Site 1 | 7.11 | 29.42 | 23.8 | 15.7 | 1.50 | 38.8 | 24.4 | 13.8 |
| Site 2 | 7.10 | 31.05 | 24.2 | 18.4 | 1.43 | 39.4 | 25.1 | 13.9 |
| Site 3 | 7.30 | 30.80 | 24.9 | 17.3 | 0.87 | 40.0 | 24.4 | 14.1 |
| Average | 7.17 | 30.42 | 24.30 | 17.13 | 1.27 | 39.40 | 24.63 | 13.93 |
| October | ||||||||
| Site 1 | 7.28 | 20.65 | 24.3 | 18.9 | 2.54 | 39.5 | 25.3 | 15.8 |
| Site 2 | 7.33 | 20.95 | 24.6 | 20.2 | 2.30 | 39.6 | 25.6 | 17.3 |
| Site 3 | 7.37 | 21.85 | 25.1 | 20.3 | 2.90 | 39.0 | 26.1 | 16.2 |
| Average | 7.33 | 21.15 | 24.67 | 19.8 | 2.58 | 39.37 | 25.67 | 16.43 |
| December | ||||||||
| Site 1 | 7.43 | 11.53 | 23.8 | 20.4 | 3.6 | 43.8 | 26.7 | 20.2 |
| Site 2 | 7.43 | 11.52 | 26.6 | 25.2 | 3.51 | 43.5 | 26.9 | 20.0 |
| Site 3 | 7.36 | 11.32 | 24.8 | 21.7 | 3.22 | 40.4 | 24.4 | 18.1 |
| Average | 7.41 | 11.46 | 25.07 | 22.43 | 3.44 | 42.57 | 26.00 | 19.43 |
| February | ||||||||
| Site 1 | 7.52 | 21.29 | 19.5 | 17.6 | 1.5 | 33.0 | 20.1 | 12.8 |
| Site 2 | 7.37 | 20.38 | 21.4 | 11.2 | 0.9 | 35.6 | 21.7 | 14.5 |
| Site 3 | 7.47 | 19.54 | 17.0 | 7.0 | 1.98 | 29.6 | 18.4 | 11.3 |
| Average | 7.45 | 20.40 | 19.30 | 11.93 | 1.46 | 32.73 | 20.07 | 12.87 |
| Gene/ Parameter | pH | Temp. (°C) | Salinity (ppt) | Turbid. (NTU) | DO (g/L) | Cond. mS/cm | TDS (g/L) | Potent. Water Density |
|---|---|---|---|---|---|---|---|---|
| toxR | −0.697 (0.039) | 0.403 (0.136) | −0.344 (0.209) | 0.465 (0.081) | −0.443 (0.098) | −0.385 (0.156) | −0.440 (0.101) | −0.574 (0.025) |
| luxR | −0.555 (0.042) | 0.692 (0.043) | 0.271 (0.329) | −0.156 (0.579) | −0.474 (0.074) | 0.185 (0.509) | 0.180 (0.521) | −0.240 (0.389) |
| srp | −0.738 (0.002) | 0.450 (0.092) | −0.320 (0.245) | 0.467 (0.079) | −0.469 (0.078) | −0.371 (0.173) | −0.420 (0.119) | −0.588 (0.021) |
| vhha | −0.480 (0.070) | 0.644 (0.009) | 0.294 (0.288) | −0.200 (0.475) | −0.428 (0.111) | 0.213 (0.446) | 0.214 (0.444) | −0.186 (0.507) |
| vhp | −0.643 (0.097) | 0.747 (0.001) | 0.235 (0.399) | −0.101 (0.720) | −0.530 (0.042) | 0.142 (0.613) | 0.132 (0.639) | −0.310 (0.261) |
| vhh | −0.515 (0.049) | 0.668 (0.007) | 0.287 (0.299) | −0.179 (0.523) | −0.449 (0.093) | 0.203 (0.468) | 0.202 (0.470) | −0.210 (0.453) |
| Gene/ Parameter | pH | Temp. (°C) | Salinity (ppt) | Turbid. (NTU) | DO (g/L) | Cond. mS/cm) | TDS (g/L) | Potent. Water Density |
|---|---|---|---|---|---|---|---|---|
| toxR | −0.927 (0.0001) | 0.893 (0.0001) | 0.221 (0.429) | 0.227 (0.416) | −0.584 (0.226) | 0.029 (0.918) | 0.098 (0.728) | −0.472 (0.076) |
| luxR | −0.724 (0.023) | 0.791 (0.004) | 0.188 (0.502) | −0.045 (0.873) | −0.583 (0.226) | 0.092 (0.744) | 0.074 (0.793) | −0.380 (0.162) |
| srp | −0.897 (0.0001) | 0.666 (0.007) | −0.228 (0.414) | 0.390 (0.151) | −0.619 (0.014) | −0.302 (0.274) | −0.359 (0.189) | −0.654 (0.008) |
| vhha | −0.957 (0.0001) | 0.809 (0.0003) | −0.092 (0.744) | 0.278 (0.316) | −0.689 (0.004) | −0.185 (0.509) | −0.235 (0.399) | −0.635 (0.010) |
| vhp | −0.653 (0.083) | 0.772 (0.0007) | 0.292 (0.291) | −0.087 (0.758) | −0.510 (0.052) | 0.169 (0.547) | 0.193 (0.491) | −0.291 (0.293) |
| vhh | −0.852 (0.0006) | 0.600 (0.018) | −0.251 (0.367) | 0.440 (0.100) | −0.559 (0.030) | −0.335 (0.194) | −0.370 (0.175) | −0.636 (0.010) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barkovskii, A.L.; Brown, C. Environmental Drivers of the Divergence of Harveyi Clade Pathogens with Distinctive Virulence Gene Profiles. Microorganisms 2024, 12, 2234. https://doi.org/10.3390/microorganisms12112234
Barkovskii AL, Brown C. Environmental Drivers of the Divergence of Harveyi Clade Pathogens with Distinctive Virulence Gene Profiles. Microorganisms. 2024; 12(11):2234. https://doi.org/10.3390/microorganisms12112234
Chicago/Turabian StyleBarkovskii, Andrei L., and Cameron Brown. 2024. "Environmental Drivers of the Divergence of Harveyi Clade Pathogens with Distinctive Virulence Gene Profiles" Microorganisms 12, no. 11: 2234. https://doi.org/10.3390/microorganisms12112234
APA StyleBarkovskii, A. L., & Brown, C. (2024). Environmental Drivers of the Divergence of Harveyi Clade Pathogens with Distinctive Virulence Gene Profiles. Microorganisms, 12(11), 2234. https://doi.org/10.3390/microorganisms12112234