
Three-Year Monitoring of Microorganisms’ Composition and Concentration in Atmospheric Aerosols of Novosibirsk City and Suburbs

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
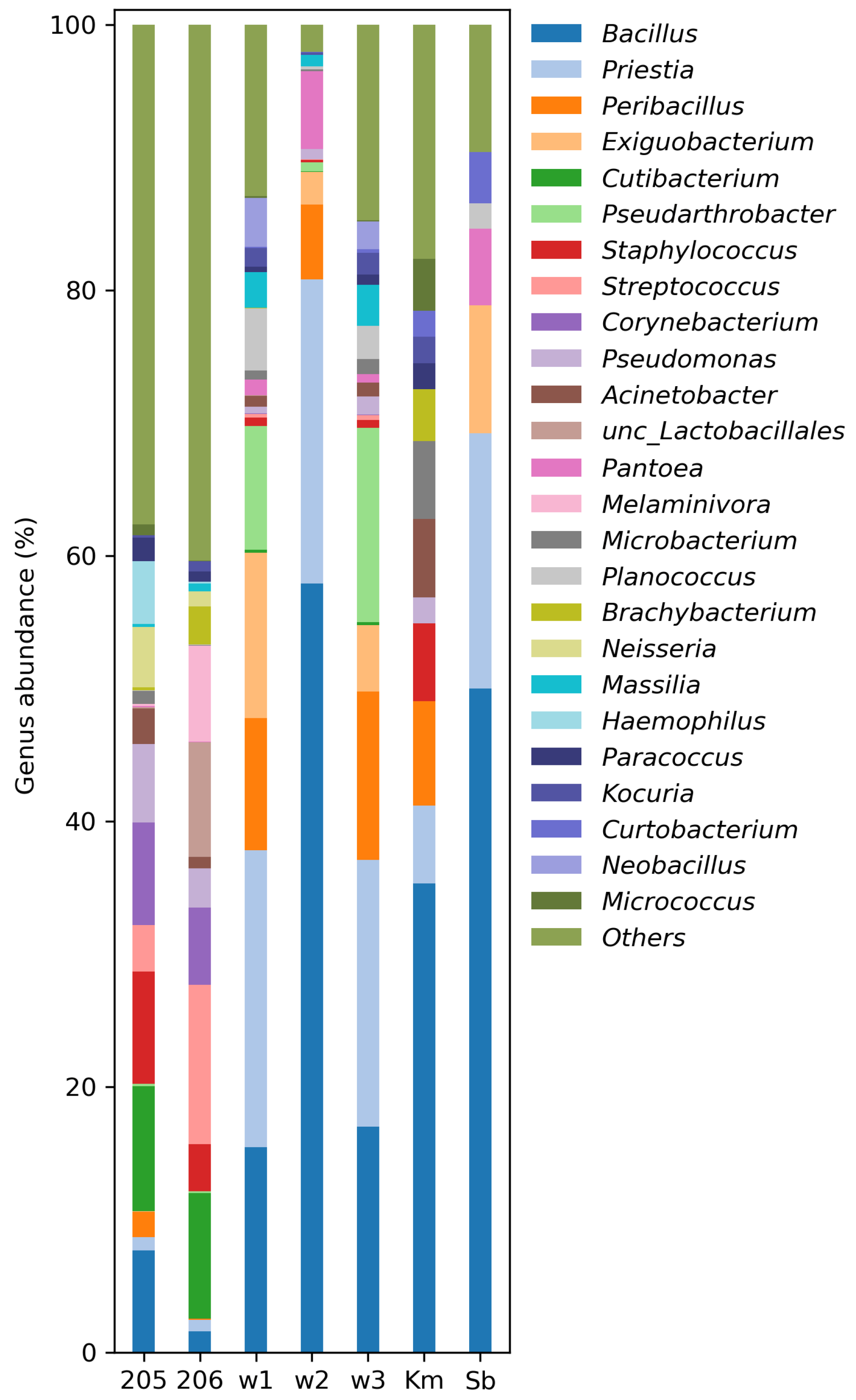
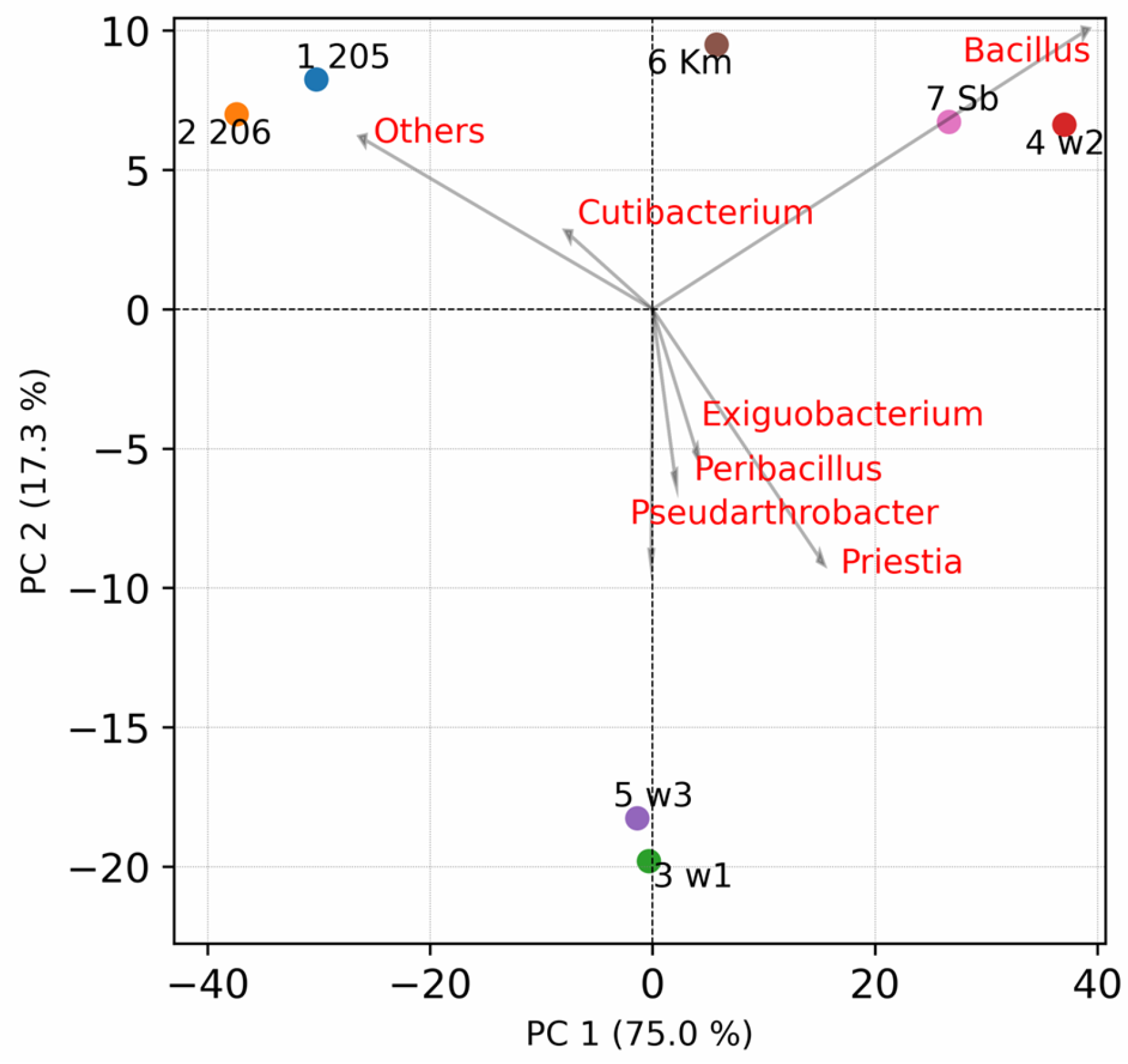
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- On the State and Protection of the Novosibirsk Region Environment in 2022; Ministry of Natural Resources and Ecology of the Novosibirsk Region: Novosibirsk, Russia, 2023. (In Russian)
- Archer, S.; Lee, K.; Caruso, T.; Leung, M.; Tong, X.; Salter, S.J.; Hinchliffe, G.; Maki, T.; Santl-Temkiv, T.; Warren-Rhodes, K.; et al. Diverse recruitment to a taxonomically structured global atmospheric microbiota. Res. Sq. 2021. Online ahead of print. [Google Scholar] [CrossRef]
- Poschl, U. Atmospheric aerosols: Composition, transformation, climate and health effects. Angew. Chem. Int. Edit. 2005, 44, 7520–7540. [Google Scholar] [CrossRef] [PubMed]
- Franchitti, E.; Caredda, C.; Anedda, E.; Traversi, D. Urban aerobiome and effects on human health: A systematic review and missing evidence. Atmosphere 2022, 13, 1148. [Google Scholar] [CrossRef]
- Lee, Y.G.; Lee, P.H.; Choi, S.M.; An, M.H.; Jang, A.S. Effects of Air Pollutants on Airway Diseases. Int. J. Environ. Res. Public Health 2021, 18, 9905. [Google Scholar] [CrossRef]
- Golikov, R.A.; Surzhikov, V.D.; Kislitsyna, V.V.; Shtaiger, V.A. Influence of environmental pollution to the health of the population (review of literature). Sci. Rev. Med. Sci. 2017, 5, 20–31. (In Russian) [Google Scholar]
- Chezganova, E.A.; Efimova, O.S.; Sakharova, V.M.; Efimova, A.R.; Sozinov, S.A.; Ismagilov, Z.R.; Brusina, E.B. Particulate matter as a possible reservoir of multidrug-resistant microorganisms in surgical healthcare settings. Fundam. Clin. Med. 2020, 5, 15–25. [Google Scholar] [CrossRef]
- Balyan, P.; Ghosh, C.; Das, S.; Banerjee, B.D. Spatio-temporal characterisation of bioaerosols at diverse outdoor land-use sites in an urban environment. Aerobiologia 2020, 36, 77–81. [Google Scholar] [CrossRef]
- Yang, L.; Shen, Z.; Wang, D.; Wie, J.; Wang, X.; Sun, J.; Xu, H.; Cao, J. Diurnal variations of size-resolved bioaerosols during autumn and winter over a semi-arid megacity in Northwest China. GeoHealth 2021, 5, e2021GH000411. [Google Scholar] [CrossRef]
- Chmiel, M.; Lenart-Boron, A. Morning–evening variation and determinants of bacterial and fungal aerosol concentration in historical objects in Kraków. Aerobiologia 2021, 37, 253–270. [Google Scholar] [CrossRef]
- Lysak, L.V.; Shoba, S.A.; Prokof’eva, T.V.; Glushakova, A.M.; Goncharov, N.V.; Belov, A.A. Abundance and diversity of prokaryotic communities of dust aerosol and urban soils in the territory of Moscow. Euras. Soil Sci. 2023, 56, 663–671. [Google Scholar] [CrossRef]
- Fröhlich-Nowoisky, J.; Kampf, C.J.; Weber, B.; Huffman, J.A.; Pöhlker, C.; Andreae, M.O.; Lang-Yona, N.; Burrows, S.M.; Gunthe, S.S.; Elbert, W.; et al. Bioaerosols in the Earth system: Climate, health, and ecosystem interactions. Atmos. Res. 2016, 182, 346–376. [Google Scholar] [CrossRef]
- Chen, X.; Kumari, D.; Achal, V. A Review on airborne microbes: The characteristics of sources, pathogenicity and geography. Atmosphere 2020, 11, 919. [Google Scholar] [CrossRef]
- Kim, K.-H.; Kabir, E.; Jahan, S.A. Airborne bioaerosols and their impact on human health. J. Environ. Sci. 2018, 67, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Mack, S.M.; Madl, A.K.; Pinkerton, K.E. Respiratory health effects of exposure to ambient particulate matter and bioaerosols. Compr. Physiol. 2020, 10, 1–20. [Google Scholar] [CrossRef]
- O’Gorman, C.M.; Fuller, H.T. Prevalence of culturable airborne spores of selected allergenic and pathogenic fungi in outdoor air. Atmos. Environ. 2008, 42, 4355–4368. [Google Scholar] [CrossRef]
- Zhang, Z.; Reponen, T.; Hershey, G.K.K. Fungal exposure and asthma: IgE and non-IgE-mediated mechanisms. Curr. Allergy Asthma Rep. 2016, 16, 86. [Google Scholar] [CrossRef]
- Gusareva, E.S.; Gaultier, N.P.E.; Premkrishnan, B.N.V.; Kee, C.; Lim, S.B.Y.; Heinle, C.E.; Purbojati, R.W.; Nee, A.P.; Lohar, S.R.; Yanqing, K.; et al. Taxonomic composition and seasonal dynamics of the air microbiome in West Siberia. Sci. Rep. 2020, 10, 21515. [Google Scholar] [CrossRef]
- Andreeva, I.S.; Safatov, A.S.; Morozova, V.V.; Solovyanova, N.A.; Puchkova, L.I.; Buryak, G.A.; Olkin, S.E.; Reznikova, I.K.; Emelyanova, E.K.; Okhlopkova, O.V.; et al. Composition and concentration of the biogenic components of the aerosols collected over Vasyugan marshes and Karakan pine forest at altitudes from 500 to 7000 m. Atmosphere 2023, 14, 301. [Google Scholar] [CrossRef]
- Belan, B.D.; Ancellet, G.; Andreeva, I.S.; Antokhin, P.N.; Arshinova, V.G.; Arshinov, M.Y.; Balin, Y.S.; Barsuk, V.E.; Belan, S.B.; Chernov, D.G.; et al. Integrated airborne investigation of the air composition over the Russian Sector of the Arctic. Atmos. Meas. Technol. 2022, 15, 3941–3967. [Google Scholar] [CrossRef]
- Safatov, A.; Andreeva, I.; Buryak, G.; Olkin, S.; Reznikova, I.; Belan, B.; Panchenko, M.; Simonenkov, D. Long-term studies of biological components of atmospheric aerosol: Trends and variability. Atmosphere 2022, 13, 651. [Google Scholar] [CrossRef]
- Andreeva, I.S.; Safatov, A.S.; Puchkova, L.I.; Solovyanova, N.A.; Ohlopkova, O.V.; Rebus, M.E.; Buryak, G.A.; Belan, B.D.; Simonenkov, D.V. Culturable microorganisms of aerosols sampled during aircraft sounding of the atmosphere over the Russian Arctic seas. Atmosphere 2024, 15, 365. [Google Scholar] [CrossRef]
- Wang, Y.; Qian, P.-Y. Conservative fragments in bacterial 16S rRNA genes and primer design for 16S ribosomal DNA amplicons in metagenomic studies. PLoS ONE 2009, 10, e7401. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Cardenas, J.A.; Manzano-Agugliaro, F. The metagenomics worldwide research. Curr. Genet. 2017, 63, 819–829. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Cadenasso, M.L.; Grove, J.M.; Boone, C.G.; Groffman, P.M.; Irwin, E.; Kaushal, S.S.; Marshall, V.; McGrath, B.P.; Nilon, C.H.; et al. Urban ecological systems: Scientific foundations and a decade of progress. J. Environ. Manag. 2010, 92, 331–362. [Google Scholar] [CrossRef]
- Lykov, I.N.; Gulordava, N.T.; Kovaleva, E.O. Medical and ecological aspects of the presence of microorganisms in the air. Ecol. Urban Areas 2020, 4, 20–25. (In Russian) [Google Scholar] [CrossRef]
- Griffin, D.W. Atmospheric movement of microorganisms in clouds of desert dust and implications for human health. Clin. Microbiol. Rev. 2007, 20, 459–477. [Google Scholar] [CrossRef]
- Ikit, M.; Durdu, M. Tinea pedis: The etiology and global epidemiology of a common fungal infection. Crit. Rev. Microbiol. 2015, 41, 374–388. [Google Scholar] [CrossRef]
- Andreeva, I.S.; Baturina, O.A.; Safatov, A.S.; Solovyanova, N.A.; Alikina, T.Y.; Puchkova, L.I.; Rebus, M.E.; Buryak, G.A.; Olkin, S.E.; Kozlov, A.S.; et al. Concentration and composition of cultured microorganisms in atmospheric air aerosols in Novosibirsk depending on the season. Atmos. Ocean. Opt. 2022, 35, 667–672. [Google Scholar] [CrossRef]
- Saggie, J. Methods of Soil Microbiology; Kolos Publishers: Moscow, Russia, 1983. (In Russian) [Google Scholar]
- Gerhardt, F.; Murray, R.G.E.; Wood, W.A.; Krieg, N.R. Methods of General Bacteriology, 2nd ed.; American Society for Microbiology: Washington, DC, USA, 1994. [Google Scholar]
- Labinskaya, A.C.; Volina, E.G. (Eds.) Guide to Medical Microbiology. General and Sanitary Microbiology. Book I; Binom: Moscow, Russia, 2020. (In Russian) [Google Scholar]
- Safatov, A.; Andreeva, I.; Buryak, G.; Ohlopkova, O.; Olkin, S.; Puchkova, L.; Reznikova, I.; Solovyanova, N.; Belan, B.; Panchenko, M.; et al. How Has the Hazard to Humans of Microorganisms Found in Atmospheric Aerosol in the South of Western Siberia Changed over 10 Years? Int. J. Environ. Res. Public Health 2020, 17, 1651. [Google Scholar] [CrossRef]
- Fadrosh, D.W.; Ma, B.; Gajer, P.; Sengamalay, N.; Ott, S.; Brotman, R.M.; Ravel, J. An improved dual-indexing approach for multiplexed 16S rRNA gene sequencing on the Illumina MiSeq platform. Microbiome 2014, 2, 6. [Google Scholar] [CrossRef]
- Naumova, N.; Alikina, T.; Tupikin, A.; Kalmykova, A.; Soldatova, G.; Vlassov, V.; Kabilov, M. Human gut microbiome response to short-term Bifidobacterium-based probiotic treatment. Indian. J. Microbiol. 2020, 60, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. SINTAX: A simple non-Bayesian taxonomy classifier for 16S and ITS sequences. bioRxiv 2016. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar] [CrossRef]
- Shin, S.K.; Kim, J.; Ha, S.M.; Oh, H.S.; Chun, J.; Sohn, J.; Yi, H. Metagenomic insights into the bioaerosols in the indoor and outdoor environments of childcare facilities. PLoS ONE 2015, 10, e0126960. [Google Scholar] [CrossRef]
- Naumova, N.B.; Kabilov, M.R. About the biodiversity of the air microbiome. Acta Naturae 2022, 14, 50–56. [Google Scholar] [CrossRef]
- Tringe, S.G.; Zhang, T.; Liu, X.; Yu, Y.; Lee, W.H.; Yap, J.; Yao, F.; Suan, S.T.; Ing, S.K.; Haynes, M.; et al. The airborne metagenome in an indoor urban environment. PLoS ONE 2008, 3, e1862. [Google Scholar] [CrossRef]
- Tong, Y.; Lighthart, B. The annual bacterial particle concentration and size distribution in the ambient atmosphere in a rural area of the Willamette Valley, Oregon. Aerosol Sci. Technol. 2000, 32, 393–403. [Google Scholar] [CrossRef]
- Huang, Z.; Yu, X.; Liu, Q.; Maki, T.; Alam, K.; Wang, Y.; Xue, F.; Tang, S.; Du, P.; Dong, Q.; et al. Bioaerosols in the atmosphere: A comprehensive review on detection methods, concentration and influencing factors. Sci. Total Environ. 2024, 912, 168818. [Google Scholar] [CrossRef]
- Els, N.; Larose, C.; Baumann-Stanzer, K.; Tignat-Perrier, R.; Keuschnig, C.; Vogel, T.M.; Sattler, B. Microbial composition in seasonal time series of free tropospheric air and precipitation reveals community separation. Aerobiologia 2019, 35, 671–701. [Google Scholar] [CrossRef]
- Harrison, R.M.; Jones, A.M.; Biggins, P.; Pomeroy, N.; Cox, C.S.; Kidd, S.P.; Hobman, J.L.; Brown, N.L.; Beswick, A. Climate factors influencing bacterial count in background air samples. Int. J. Biometeorol. 2005, 49, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Bowers, R.M.; Clements, N.; Emerson, J.B.; Wiedinmyer, C.; Hannigan, M.P.; Fierer, N. Seasonal variability in bacterial and fungal diversity of the near-surface atmosphere. Environ. Sci. Technol. 2013, 47, 12097–12106. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Ouyang, Z.; Zheng, H.; Wang, X.; Hu, L. Culturable airborne bacteria in outdoor environments in Beijing, China. Microb. Ecol. 2007, 54, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Harrison, R.M. The effects of meteorological factors on atmospheric bioaerosol concentrations—A review. Sci. Total Environ. 2004, 326, 151–180. [Google Scholar] [CrossRef]
- Jensen, L.Z.; Glasius, M.; Gryning, S.-E.; Massling, A.; Finster, K.; Šantl-Temkiv, T. Seasonal variation of the atmospheric bacterial community in the Greenlandic high Arctic is influenced by weather events and local and distant sources. Front. Microbiol. 2022, 13, 909980. [Google Scholar] [CrossRef]
- Brown, J.K.M.; Hovmøller, M.S. Aerial dispersal of pathogens on the global and continental scales and its impact on plant disease. Science 2002, 297, 537–541. [Google Scholar] [CrossRef]
- Kellogg, C.A.; Griffin, D.W. Aerobiology and global transport of desert dust. Trends Ecol. Evol. 2006, 21, 638–644. [Google Scholar] [CrossRef]
- Andreeva, I.S.; Safatov, A.S.; Puchkova, L.I.; Emelyanova, E.K.; Buryak, G.A.; Ternovoi, V.A. Spore forming bacteria isolated from atmospheric aerosols in Southwestern Siberia during atmospheric transfer of air masses. Bull. Nizhnevartovsk State Univ. 2018, 3, 123–130. (In Russian) [Google Scholar]
- Gritsenko, L.Z.; Kolokolova, Y.V.; Kolesnikova, A.G.; Mishin, V.V.; Ananyeva, M.N. Role of acinetobacter in the occurrence of problematic infections. Med. Soc. Probl. Fam. 2014, 19, 122–127. (In Russian) [Google Scholar]
- Moreira, J.S.; Riccetto, A.G.; da Silva, M.T.N.; dos Santos Vilela, M.M. Endocarditis by Kocuria rosea in an immunocompetent child. Braz. J. Infect. Dis. 2015, 19, 82–84. [Google Scholar] [CrossRef] [PubMed]
- von Hertzen, L.; Hanski, I.; Haahtela, T. Natural immunity. Biodiversity loss and inflammatory diseases are two global megatrends that might be related. EMBO Rep. 2011, 12, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- Ege, M.J.; Mayer, M.; Normand, A.-C.; Genuneit, J.; Cookson, W.O.C.M.; Braun-Fahrländer, C.; Heederik, D.; Piarroux, R.; von Mutius, E. Exposure to environmental microorganisms and childhood asthma. N. Engl. J. Med. 2011, 364, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Vojnovic, S.; Aleksic, I.; Ilic-Tomic, T.; Stevanovic, M.; Nikodinovic-Runic, J. Bacillus and Streptomyces spp. as hosts for production of industrially relevant enzymes. Appl. Microbiol. Biotechnol. 2024, 108, 185. [Google Scholar] [CrossRef]
- Yang, M.; Hutchinson, N.; Ye, N.; Yin, J.; Guan, M.; Wang, Z.; Chen, P.; Yang, S.; Crane, J.D.; Zhang, K.; et al. Engineered Bacillus subtilis as oral probiotics to enhance clearance of blood lactate. bioRxiv 2023. [Google Scholar] [CrossRef]
- Lal, S.; Tabacchioni, S. Ecology and biotechnological potential of Paenibacillus polymyxa: A minireview. Indian J. Microbiol. 2009, 49, 2–10. [Google Scholar] [CrossRef]
- Biedendieck, R.; Knuuti, T.; Moore, S.J.; Jahn, D. The “beauty in the beast”—The multiple uses of Priestia megaterium in biotechnology. Appl. Microbiol. Biotechnol. 2021, 105, 5719–5737. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample, Sampling Time | Medium of Isolation/Number of Microorganisms Isolated (CFU/mL) | ||||||
|---|---|---|---|---|---|---|---|
| LA | LA (1:10) | SAA | SA | Tg | Sabouraud Medium | Total Quantity | |
| V1-1 8 September 2021 (day) | 58 f 9 b, 12 n 4 akt, 6 k | 44 f 38 b, 7 k, 28 n, 6 akt | 25 f 2 b 6 akt 7 n | >1000 mch + akt | 22 f 2 n 1 b | 51 f 20 b | >1283 |
| V2-1 8–9 September 2021 (night) | 65 f 39 b, 13 k 7 n | 58 f 60 b, 38 n | >100 fungi, bacteria count is impossible | >1000 mch + akt | 12 f 5 n 3 d | 50 f 30 b | >1450 |
| Sample, Sampling Time | Medium of Isolation/Number of Microorganisms Isolated (CFU/mL) | ||||||
|---|---|---|---|---|---|---|---|
| LA | LA (1:10) | SAA | SA | Tg | Sabouraud Medium | Total Quantity | |
| V1 8 September 2021 (day) | 60 f 5 b 5 n | 30 f 10 k | 90 k 25 n 15 b | 25 f | 10 f 5 k | 16 f | 296 |
| V2 8–9 September 2021 (night) | 65 f 10 n | 60 f 5 b | 5 f | 60 f | 35 f 5 n | 34 f 1 n | 275 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andreeva, I.; Safatov, A.; Totmenina, O.; Olkin, S.; Rebus, M.; Buryak, G.; Alikina, T.; Baturina, O.; Kabilov, M. Three-Year Monitoring of Microorganisms’ Composition and Concentration in Atmospheric Aerosols of Novosibirsk City and Suburbs. Microorganisms 2024, 12, 2068. https://doi.org/10.3390/microorganisms12102068
Andreeva I, Safatov A, Totmenina O, Olkin S, Rebus M, Buryak G, Alikina T, Baturina O, Kabilov M. Three-Year Monitoring of Microorganisms’ Composition and Concentration in Atmospheric Aerosols of Novosibirsk City and Suburbs. Microorganisms. 2024; 12(10):2068. https://doi.org/10.3390/microorganisms12102068
Chicago/Turabian StyleAndreeva, Irina, Aleksandr Safatov, Olga Totmenina, Sergei Olkin, Maxim Rebus, Galina Buryak, Tatiana Alikina, Olga Baturina, and Marsel Kabilov. 2024. "Three-Year Monitoring of Microorganisms’ Composition and Concentration in Atmospheric Aerosols of Novosibirsk City and Suburbs" Microorganisms 12, no. 10: 2068. https://doi.org/10.3390/microorganisms12102068
APA StyleAndreeva, I., Safatov, A., Totmenina, O., Olkin, S., Rebus, M., Buryak, G., Alikina, T., Baturina, O., & Kabilov, M. (2024). Three-Year Monitoring of Microorganisms’ Composition and Concentration in Atmospheric Aerosols of Novosibirsk City and Suburbs. Microorganisms, 12(10), 2068. https://doi.org/10.3390/microorganisms12102068