
Alginate–Bentonite Encapsulation of Extremophillic Bacterial Consortia Enhances Chenopodium quinoa Tolerance to Metal Stress

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganism
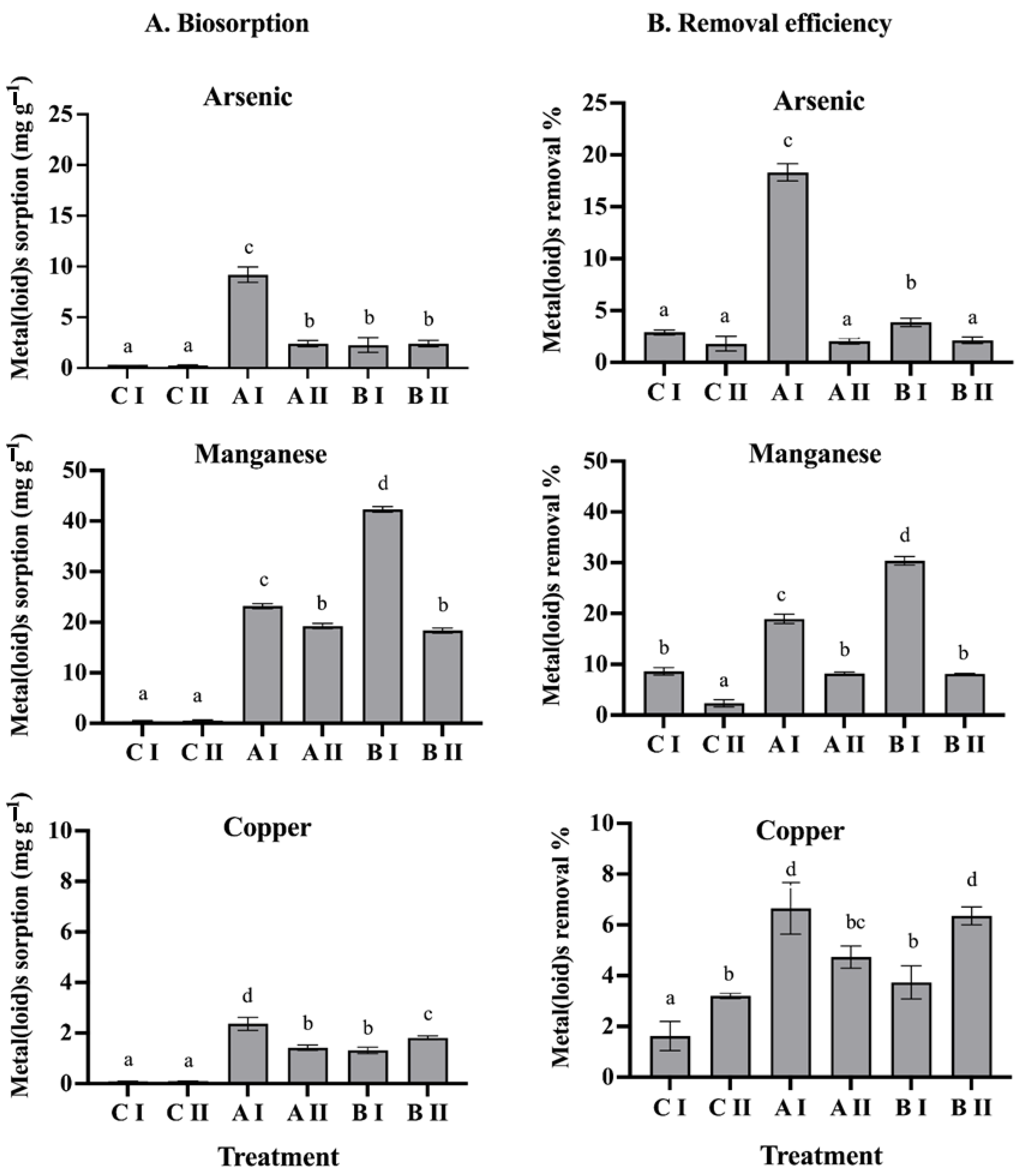
2.2. Biosorption of Metal(loid)s of Rhizosphere Bacterial Consortia
2.3. Encapsulation
2.3.1. Bacterial Encapsulation by Ionic Gelation
2.3.2. Analysis of Drying Temperature and Microbial Viability of the Beads
2.3.3. Pearl Encapsulation Efficiency (EE)
2.3.4. Size and Sphericity Factor
2.3.5. Analysis of Bead Swelling
2.4. Greenhouse Experiment
2.4.1. Soil Characteristics and Measurements
2.4.2. Experimental Design
2.4.3. Growth Conditions and Measurements
2.5. Statistical Analysis
3. Results
3.1. Biosorption of Metal(loid)s in the Bacterial Consortia
3.2. Encapsulation Matrices
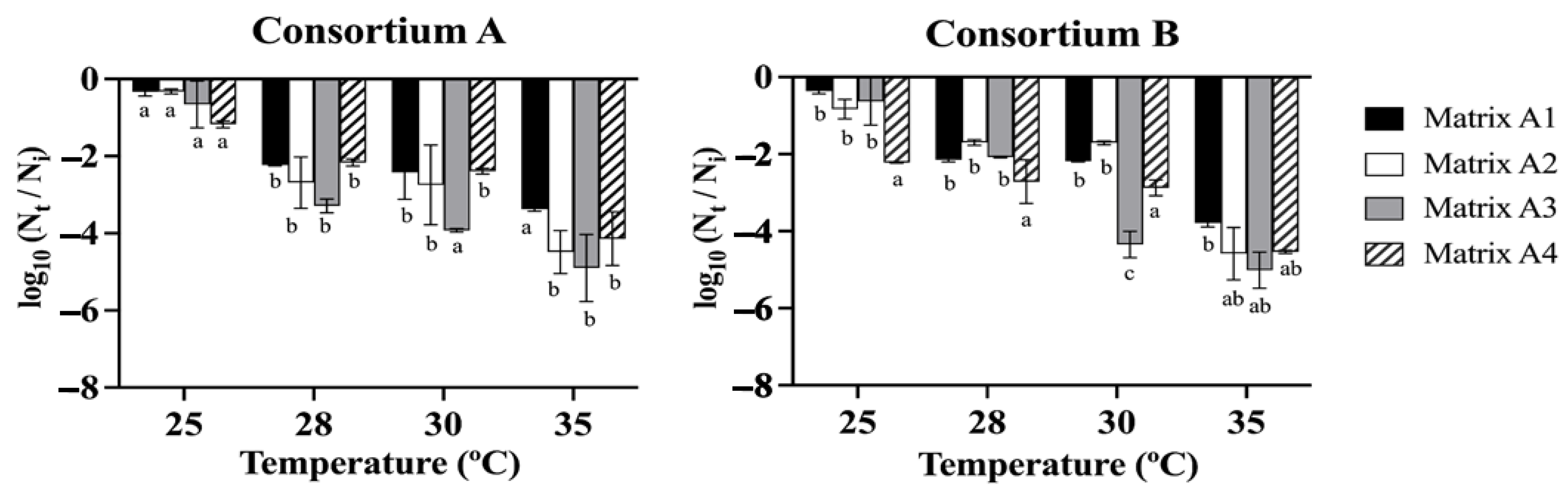
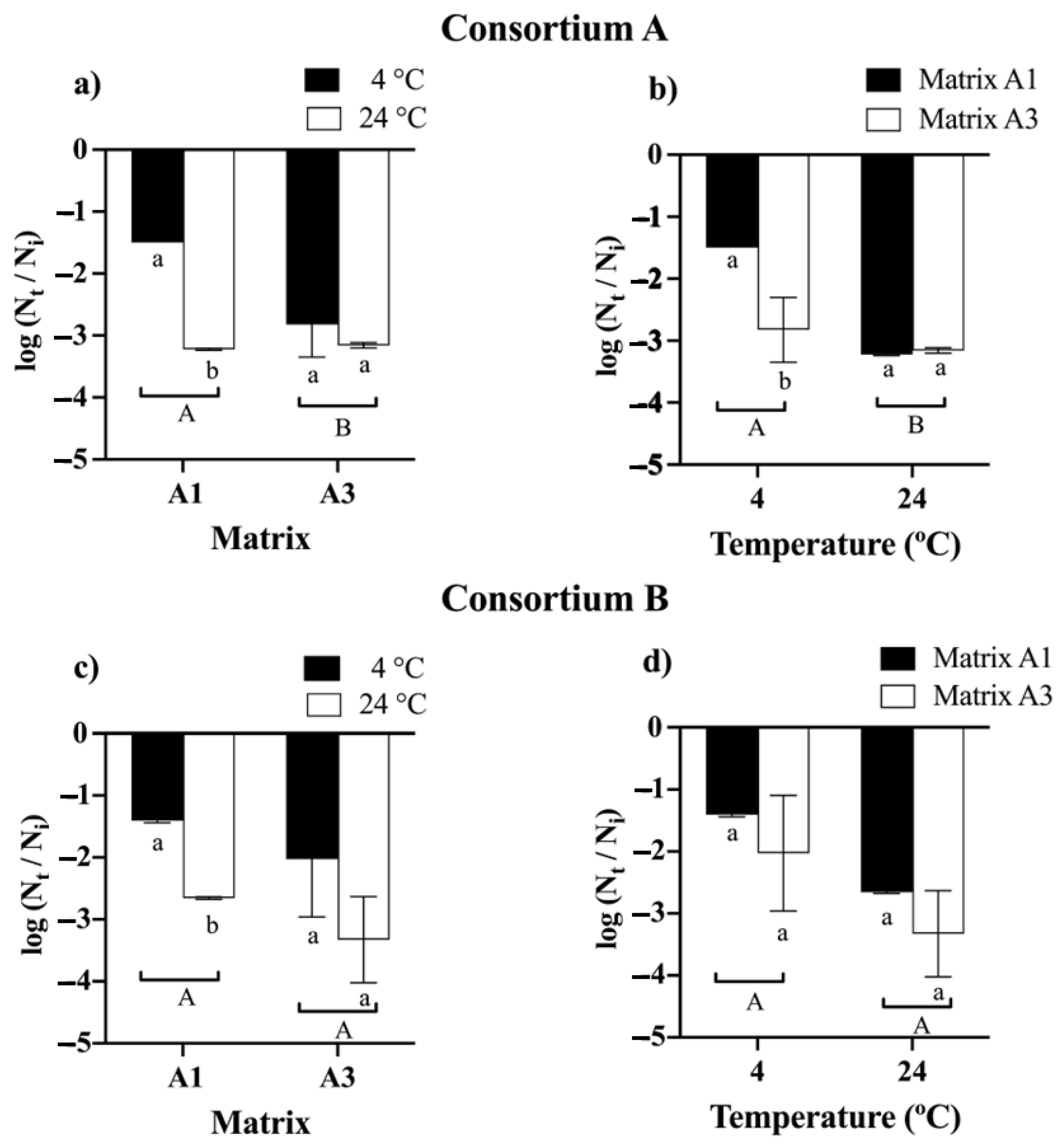
3.3. Analysis of Drying Temperature and Analysis of Microbiological Viability of the Beads
3.3.1. Drying Temperature Analysis
3.3.2. Feasibility Analysis in Bead
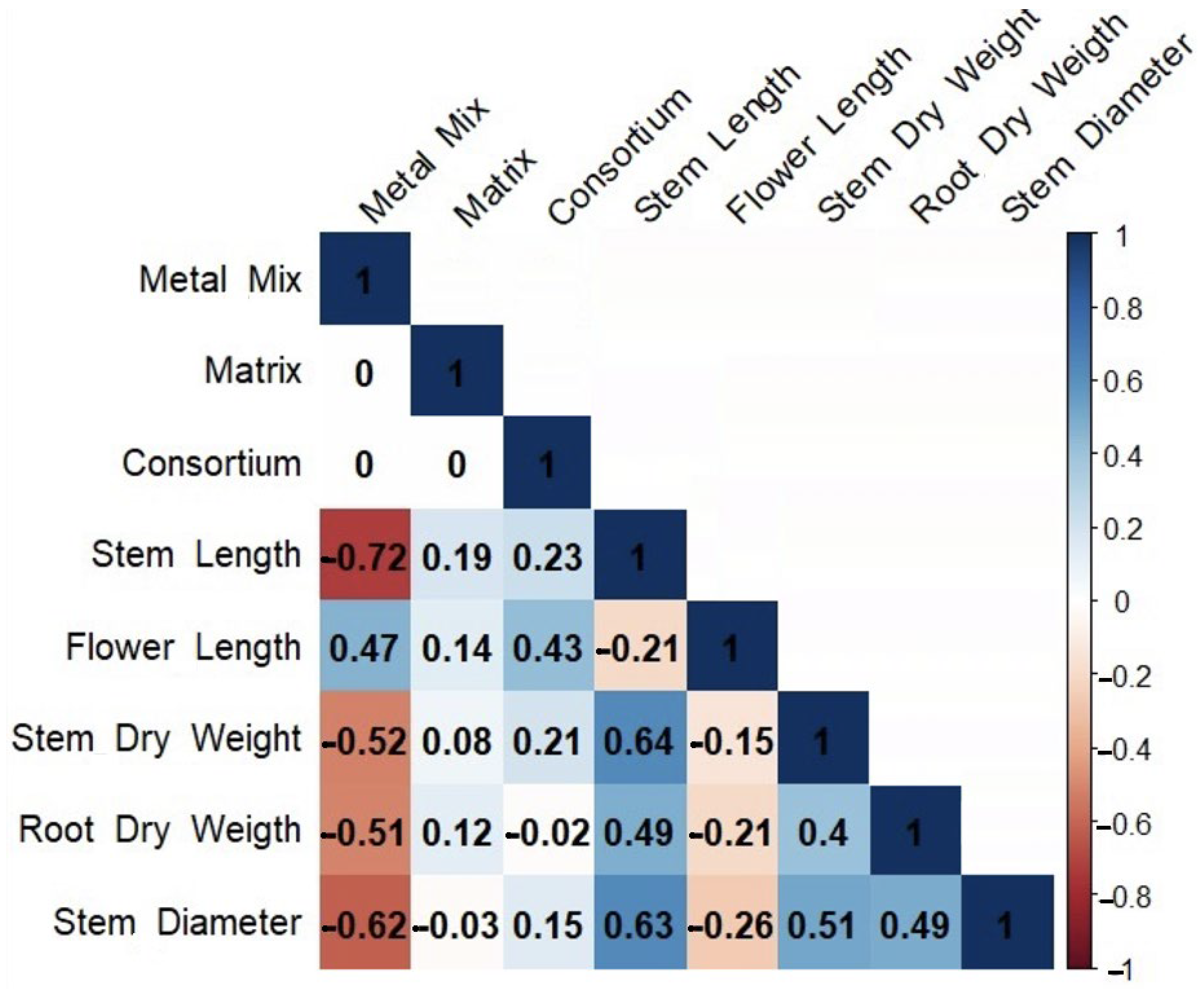
3.4. Greenhouse Experiment
3.4.1. Morphological Characteristics of Chenopodium quinoa
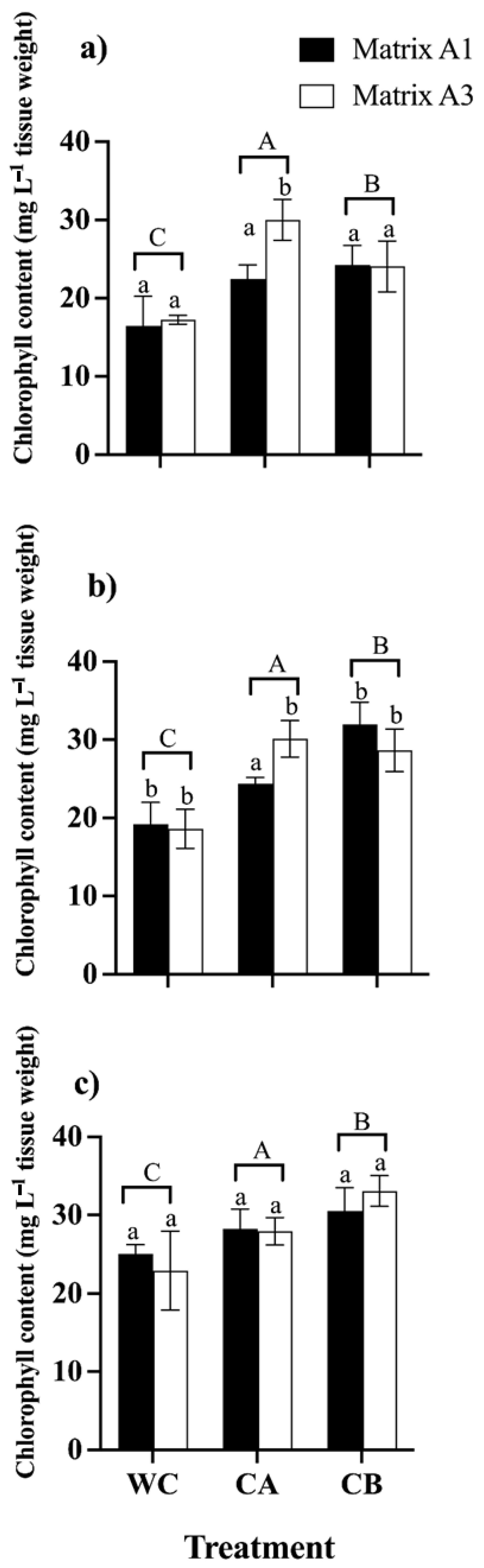
3.4.2. Photosynthetic Pigments
3.4.3. Lipid Peroxidation Assay
3.4.4. Chemical Analysis of Plant Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Angeli, V.; Silva, P.; Massuela, D.; Khan, M.W.; Hamar, A.; Khajehei, F.; Graeff-Hönninger, S.; Piatti, C. Quinoa (Chenopodium quinoa Willd.): An Overview of the Potentials of the “Golden Grain” and Socio-Economic and Environmental Aspects of Its Cultivation and Marketization. Foods 2020, 9, 216. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Tsao, R. Phytochemicals in quinoa and amaranth grains and their antioxidant, anti-inflammatory, and potential health beneficial effects: A review. Mol. Nourish Food Res. 2017, 61, 1600767. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Akhtar, S.; Amjad, M.; Iqbal, S.; Jacobsen, S.E. Growth and physiological responses of quinoa to drought and temperature stress. J. Agron. Crop Sci. 2016, 202, 445–453. [Google Scholar] [CrossRef]
- Bhargava, A.; Shukla, S.; Srivastava, J.; Singh, N.; Ohri, D. Chenopodium: A prospective plant for phytoextraction. Acta Physiol. Plants. 2008, 30, 111–120. [Google Scholar] [CrossRef]
- Hinojosa, L.; González, J.A.; Barrios-Masias, F.H.; Fuentes, F.; Murphy, K.M. Quinoa abiotic stress responses: A review. Plants 2018, 7, 106. [Google Scholar] [CrossRef]
- Thomas, E.; Lavkulich, L. Community considerations for quinoa production in the urban environment Dog. J. Plant Sci. 2015, 95, 397–404. [Google Scholar] [CrossRef]
- Clemens, S.; Ma, J. Toxic heavy metal and metalloid accumulation in crop plants and foods. Annu. Rev. Plant Biol. 2016, 67, 489–512. [Google Scholar] [CrossRef]
- Jain, S.; Muneer, S.; Guerriero, G.; Liu, S.; Vishwakarma, K.; Chauhan, D.K.; Dubey, N.K.; Tripathi, D.K.; Sharma, S. Tracing the role of plant proteins in the response to metal toxicity: A comprehensive review. Plant Signal. Behav. 2018, 13, e1507401. [Google Scholar] [CrossRef]
- Sharma, R.K.; Archana, G. Cadmium minimization in food crops by cadmium resistant plant growth promoting rhizobacteria. Appl. Soil Ecol. 2016, 107, 66–78. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic response of plants under different abiotic stresses: A review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Saleem, M.; Asghar, H.N.; Zahir, Z.A.; Shahid, M. Evaluation of lead tolerant plant growth promoting rhizobacteria for plant growth and phytoremediation in lead contamination. Rev. Int. Contam. 2019, 35, 999–1009. [Google Scholar] [CrossRef]
- Renu; Sarim, K.; Sahu, U.; Bhoyar, M.S.; Singh, D.P.; Singh, U.B.; Sahu, A.; Gupta, A.; Mandal, A.; Thakur, J.K.; et al. Augmentation of metal-tolerant bacteria elevates growth and reduces metal toxicity in spinach. Bioremediat. J. 2021, 25, 108–127. [Google Scholar] [CrossRef]
- Alvarado, R.; Fuentes, A.; Ortiz, J.; Herrera, H.; Arriagada, C. Metal (loid)-resistant bacterial consortia with antimycotic properties increase tolerance of Chenopodium quinoa Wild. to metal (loid) stress. Rhizosphere 2022, 23, 100569. [Google Scholar] [CrossRef]
- Gontia-Mishra, I.; Sapre, S.; Sikdar, S.; Tiwari, S. Belowground dialogue between plant roots and beneficial microbes. In Microbiome Stimulants for Crops; Elsevier: Amsterdam, The Netherlands, 2021; pp. 141–158. [Google Scholar] [CrossRef]
- Maestro, I.; Granado, S.; Redondo-Nieto, S.; Battaglia, A.; Poza-Viejo, L.; Matías, J.; Bolaños, L.; Reguera, M. Unveiling changes in rhizosphere-associated bacteria linked to the genotype and water stress in quinoa. Microb. Biotechnol. 2023, 16, 2326–2344. [Google Scholar] [CrossRef]
- Vidal, C.; González, F.; Santander, C.; Pérez, R.; Gallardo, V.; Santos, C.; Aponte, H.; Ruiz, A.; Cornejo, P. Management of rhizosphere microbiota and plant production under drought stress: A comprehensive review. Plants 2022, 11, 2437. [Google Scholar] [CrossRef] [PubMed]
- Aponte, H.; Sulbaran-Bracho, Y.; Mondaca, P.; Vidal, C.; Pérez, R.; Meier, S.; Cornejo, P.; Rojas, C. Biochemical, Catabolic, and PGP Activity of Microbial Communities and Bacterial Strains from the Root Zone of Baccharis linearis in a Mediterranean Mine Tailing. Microorganisms 2023, 11, 2639. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H.; Noori, F.; Ebadi, A.; Samani, R. Alleviation of stress-induced ethylene-mediated negative impact on crop plants by bacterial ACC deaminase: Perspectives and applications in stressed agriculture management. In Plant Microbiomes for Sustainable Agriculture; Springer: Singapore, 2020; pp. 287–315. [Google Scholar] [CrossRef]
- Merino, N.; Aronson, H.; Bojanova, D.; Feyhl-Buska, J.; Wong, M.L.; Zhang, S.; Giovannelli, D. Living at the extremes: Extremophiles and the limits of life in a planetary context. Front. Microbiol. 2019, 10, 780. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, J.; Soto, J.; Fuentes, A.; Herrera, H.; Meneses, C.; Arriagada, C. The Endophytic Fungus Chaetomium cupreum Regulates Expression of Genes Involved in the Tolerance to Metals and Plant Growth Promotion in Eucalyptus globulus Roots. Microorganisms 2019, 7, 490. [Google Scholar] [CrossRef]
- Harman, G.; Khadka, R.; Doni, F.; Uphoff, N. Benefits to plant health and productivity from enhancing plant microbial symbionts. Front. Plant Sci. 2021, 11, 610065. [Google Scholar] [CrossRef]
- Tabacchioni, S.; Passato, S.; Ambrosino, P.; Huang, L.; Caldara, M.; Cantale, C.; Hett, J.; Del Fiore, A.; Fiore, A.; Schlüter, A.; et al. Identification of beneficial microbial consortia and bioactive compounds with potential as plant biostimulants for a sustainable agriculture. Microorganisms 2021, 9, 426. [Google Scholar] [CrossRef]
- Khare, E.; Arora, N.K. Effects of soil environment on field efficacy of microbial inoculants. In Plant Microbes Symbiosis: Applied Facets; Springer: Berlin/Heidelberg, Germany, 2014; pp. 353–381. [Google Scholar] [CrossRef]
- Bradáčová, K.; Florea, A.S.; Bar-Tal, A.; Minz, D.; Yermiyahu, U.; Shawahna, R.; Kraut-Cohen, J.; Zolti, A.; Erel, R.; Dietel, K.; et al. Microbial consortia versus single-strain inoculants: An advantage in PGPM-assisted tomato production? Agronomy 2019, 9, 105. [Google Scholar] [CrossRef]
- Timofeeva, A.; Galyamova, M.; Sedykh, S. Plant Growth-Promoting Soil Bacteria: Nitrogen Fixation, Phosphate Solubilization, Siderophore Production, and Other Biological Activities. Plants 2023, 12, 4074. [Google Scholar] [CrossRef] [PubMed]
- Nunes, P.S.O.; Lacerda-Junior, G.V.; Mascarin, G.M.; Guimarães, R.A.; Medeiros, F.H.V.; Arthurs, S.; Bettiol, W. Microbial consortia of biological products: Do they have a future? Biol. Control 2024, 188, 105439. [Google Scholar] [CrossRef]
- Santoyo, G.; Guzman-Guzman, P.; Parra-Cota, F.; Santos-Villalobos, S.; Orozco-Mosqueda, M.; Glick, B. Plant growth stimulation by microbial consortia. Agronomy 2021, 2021, 219. [Google Scholar] [CrossRef]
- Emami, S.; Alikhani, H.; Pourbabaei, A.; Etesami, H.; Motashare Zadeh, B.; Sarmadian, F. Improved growth and nutrient acquisition of wheat genotypes in phosphorus deficient soils by plant growth-promoting rhizospheric and endophytic bacteria. Soil Sci. Plant Nutr. 2018, 64, 719–727. [Google Scholar] [CrossRef]
- Jha, C.; Saraf, M. Evaluation of Multispecies Plant-Growth-Promoting Consortia for the Growth Promotion of Jatropha curcas L. J. Plant Growth Regul. 2012, 31, 588–598. [Google Scholar] [CrossRef]
- Bashan, Y.; De-Bashan, L.E.; Prabhu, S. Superior polymeric formulations and emerging innovative products of bacterial inoculants for sustainable agriculture and the environment. In Agriculturally Important Microorganisms: Commercialization and Regulatory Requirements in Asia; Springer: Berlin/Heidelberg, Germany, 2016; pp. 15–46. [Google Scholar] [CrossRef]
- Martínez-Cano, B.; Mendoza-Meneses, C.J.; García-Trejo, J.F.; Macías-Bobadilla, G.; Aguirre-Becerra, H.; Soto-Zarazúa, G.M.; Feregrino-Pérez, A.A. Review and perspectives of the use of alginate as a polymer matrix for microorganisms applied in agro-industry. Molecules 2022, 27, 4248. [Google Scholar] [CrossRef]
- Pacheco-Aguirre, J.; Ruíz-Sánchez, E.; Ballina-Gómez, H.; Alvarado-López, C. Does polymer-based encapsulation enhance performance of plant growth promoting microorganisms? A meta-analysis view. Agrociencia 2017, 51, 173–187. [Google Scholar]
- Batista, D.; De Oliveira, I.; Ribeiro, A.; Fonseca, E.J.S.; Santos-Magalhães, N.S.; de Sena-Filho, J.G.; Teodoro, A.V.; Grillo, L.A.M.; de Almeida, R.S.; Dornelas, C.B. Encapsulation and release of Beauveria bassiana from alginate–bentonite nanocomposite. RSC Adv. 2017, 7, 26468–26477. [Google Scholar] [CrossRef]
- Fomina, M.; Skorochod, I. Microbial interaction with clay minerals and its environmental and biotechnological implications. Minerals 2020, 10, 861. [Google Scholar] [CrossRef]
- Khodaverdiloo, H.; Samadi, A. Batch equilibrium study on sorption, desorption, and immobilisation of cadmium in some semi-arid zone soils as affected by soil properties. Soil Res. 2011, 49, 444–454. [Google Scholar] [CrossRef]
- Whelehan, M.; Marison, I. Microencapsulation using vibrating technology. J. Microencapsul. 2011, 28, 669–688. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, M.; Abdallah, N.A.; El-Shafei, K.; Tawfik, N.F.; El-Sayed, H.S. Survivability of alginate-microencapsulated Lactobacillus plantarum during storage, simulated food processing and gastrointestinal conditions. Heliyon 2020, 6, e03541. [Google Scholar] [CrossRef] [PubMed]
- Lotfipour, F.; Mirzaeei, S.; Maghsoodi, M. Preparation and characterization of alginate and psyllium beads containing Lactobacillus acidophilus. Sci. World J. 2012, 2012, 680108. [Google Scholar] [CrossRef] [PubMed]
- Zazzali, I.; Calvo, T.R.A.; Ruíz-Henestrosa, V.M.P.; Santagapita, P.R.; Perullini, M. Effects of pH, extrusion tip size and storage protocol on the structural properties of Ca (II)-alginate beads. Carbohydr. Polym. 2019, 206, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Lang, Q.; Yildirimer, L.; Lin, Z.Y.; Cui, W.; Annabi, N.; Ng, K.W.; Dokmeci, M.R.; Ghaemmaghami, A.M.; Khademhosseini, A. Photocrosslinkable gelatin hydrogel for epidermal tissue engineering. Adv. Healthc. Mater. 2016, 5, 108–118. [Google Scholar] [CrossRef]
- Bech, J.; Tume, P.; Longan, L.; Reverter, F. Baseline concentrations of trace elements in surface soils of the Torrelles and Sant Climent Municipal Districts (Catalonia, Spain). Environ. Monit. Assess. 2005, 108, 309–322. [Google Scholar] [CrossRef]
- Soil Quality—Extraction of Trace Elements Soluble in Aqua Regia; International Organization of Standards: Geneva, Switzerland, 1995.
- Hiscox, J.; Israelstam, G. A method for the extraction of chlorophyll from leaf tissue without maceration Canada. J. Bot. 1979, 57, 1332–1334. [Google Scholar] [CrossRef]
- Du, Z.; Bramlage, W. Modified thiobarbituric acid assay for measuring lipid oxidation in sugar-rich plant tissue extracts. Agric. Food Chem. 1992, 40, 1566–1570. [Google Scholar] [CrossRef]
- Jiang, H.; Zhao, X.; Fang, J.; Xiao, Y. Physiological responses and metal uptake of Miscanthus under cadmium/arsenic stress. Environ. Sci. Pollut. Beef. 2018, 25, 28275–28284. [Google Scholar] [CrossRef]
- Khanna, K.; Jamwal, V.L.; Gandhi, S.G.; Ohri, P.; Bhardwaj, R. Metal resistant PGPR lowered Cd uptake and expression of metal transporter genes with improved growth and photosynthetic pigments in Lycopersicon esculentum under metal toxicity. Sci. Rep. 2019, 9, 5855. [Google Scholar] [CrossRef] [PubMed]
- Orellana, R.; Macaya, C.; Bravo, G.; Dorochesi, F.; Cumsille, A.; Valencia, R.; Rojas, C.; Seeger, M. Living at the frontiers of life: Extremophiles in Chile and their potential for bioremediation. Front. Microbiol. 2018, 9, 2309. [Google Scholar] [CrossRef] [PubMed]
- Kanamarlapudi, S.L.R.K.; Muddada, S. Structural Changes of Biomass on Biosorption of Iron (II) from Aqueous Solutions: Isotherm and Kinetic Studies. Pol. J. Microbiol. 2019, 68, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Priya, A.; Gnanasekaran, L.; Dutta, K.; Rajendran, S.; Balakrishnan, D.; Soto-Moscoso, M. Biosorption of heavy metals by microorganisms: Evaluation of different underlying mechanisms. Chemosphere 2022, 307, 135957. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, A.; Ahmed, B.; Zaidi, A.; Khan, M.S. Biosorption of heavy metals by dry biomass of metal tolerant bacterial biosorbents: An efficient metal clean-up strategy. Environ. Monit. Assess. 2020, 192, 801. [Google Scholar] [CrossRef]
- Lambrese, Y.S.; Illanes, C.O.; Ochoa, N.A. Advancing bacterial endophyte encapsulation in alginate for sustainable agriculture: Method comparisons, morphology and viability assessment. J. Clean. Prod. 2024, 457, 142473. [Google Scholar] [CrossRef]
- Meftah Kadmiri, I.; El Mernissi, N.; Azaroual, S.E.; Mekhzoum, M.E.M.; Qaiss, A.E.K.; Bouhfid, R. Bioformulation of Microbial Fertilizer Based on Clay and Alginate Encapsulation. Curr. Microbiol. 2020, 78, 86–94. [Google Scholar] [CrossRef]
- He, Y.; Wu, Z.; Tu, L.; Han, Y.; Zhang, G.; Li, C. Encapsulation and characterization of slow-release microbial fertilizer from the composites of bentonite and alginate. Appl. Clay Sci. 2015, 109, 68–75. [Google Scholar] [CrossRef]
- Kragović, M.; Pašalić, S.; Marković, M.; Petrović, M.; Nedeljković, B.; Momčilović, M.; Stojmenović, M. Natural and modified zeolite—Alginate composites. Application for removal of heavy metal cations from contaminated water solutions. Minerals 2018, 8, 11. [Google Scholar] [CrossRef]
- Zhou, J.; Sun, Q. Sodium Alginate/Modified Bentonite Composite Bead Adsorptive Removal of Norfloxacin: Static and Dynamic Adsorption. Polymers 2022, 14, 3984. [Google Scholar] [CrossRef]
- Zhang, H.; Shi, Y.; Xu, X.; Zhang, M.; Ma, L. Structure regulation of bentonite-alginate nanocomposites for controlled release of imidacloprid. ACS Omega 2020, 5, 10068–10076. [Google Scholar] [CrossRef] [PubMed]
- Ahmad Raus, R.; Wan Nawawi, W.M.F.; Nasaruddin, R.R. Alginate and alginate composites for biomedical applications. Asian J. Pharm. Sci. 2021, 16, 280–306. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Cybulska, J.; Frąc, M.; Zdunek, A. Application of polysaccharides for the encapsulation of beneficial microorganisms for agricultural purposes: A review. Int. J. Biol. Macromol. 2023, 244, 125366. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Sánchez, B.; Guzmán-Guzmán, P.; Morales-Cedeño, L.R.; Orozco-Mosqueda, M.d.C.; Saucedo-Martínez, B.C.; Sánchez-Yáñez, J.M.; Fadiji, A.E.; Babalola, O.O.; Glick, B.R.; Santoyo, G. Bioencapsulation of microbial inoculants: Mechanisms, formulation types and application techniques. Appl. Biosci. 2022, 1, 198–220. [Google Scholar] [CrossRef]
- Szopa, D.; Mielczarek, M.; Skrzypczak, D.; Izydorczyk, G.; Mikula, K.; Chojnacka, K.; Witek-Krowiak, A. Encapsulation efficiency and survival of plant growth-promoting microorganisms in an alginate-based matrix—A systematic review and protocol for a practical approach. Ind. Crops Prod. 2022, 181, 114846. [Google Scholar] [CrossRef]
- Piornos, J.A.; Burgos-Díaz, C.; Morales, E.; Rubilar, M.; Acevedo, F. Highly efficient encapsulation of linseed oil into alginate/lupin protein beads: Optimization of the emulsion formulation. Food Hydrocoll. 2017, 63, 139–148. [Google Scholar] [CrossRef]
- Purnomo, A.S.; Putra, S.R.; Putro, H.S.; Hamzah, A.; Rohma, N.A.; Rohmah, A.A.; Rizqi, H.D.; Asranudin; Tangahu, B.V.; Warmadewanthi, I.D.A.A.; et al. The application of biosurfactant-producing bacteria immobilized in PVA/SA/bentonite bio-composite for hydrocarbon-contaminated soil bioremediation. RSC Adv. 2023, 13, 21163–21170. [Google Scholar] [CrossRef]
- Schoebitz, M.; Simonin, H.; Poncelet, D. Starch filler and osmoprotectants improve the survival of rhizobacteria in dried alginate beads. J. Microencapsul. 2012, 29, 532–538. [Google Scholar] [CrossRef]
- Li, X.; Wu, Z.; He, Y.; Ye, B.-C.; Wang, J. Preparation and characterization of monodisperse microcapsules with alginate and bentonite via external gelation technique encapsulating Pseudomonas putida Rs-198. Int. J. Polym. Mater. 2017, 28, 1556–1571. [Google Scholar] [CrossRef]
- Saberi-Rise, R.; Moradi-Pour, M. The effect of Bacillus subtilis Vru1 encapsulated in alginate–bentonite coating enriched with titanium nanoparticles against Rhizoctonia solani on bean. Int. J. Biol. Macromol. 2020, 152, 1089–1097. [Google Scholar] [CrossRef]
- Dong, J.; He, Y.; Zhang, J.; Wu, Z. Tuning alginate-bentonite microcapsule size and structure for the regulated release of P. putida Rs-198. Chin. J. Chem. Eng. 2022, 48, 12–20. [Google Scholar] [CrossRef]
- Pop, O.L.; Diaconeasa, Z.; Thorsten, B.; Ciuzan, O.; Pamfil, D.; Vodnar, D.C.; Socaciu, C. Effect of glycerol, as cryoprotectant in the encapsulation and freeze drying of microspheres containing probiotic cells. Bulletin of University of Agricultural Sciences and Veterinary Medicine Cluj-Napoca. Food Sci. Technol. 2015, 72, 27–32. [Google Scholar] [CrossRef]
- Schoebitz, M.; López, M.D.; Roldán, A. Bioencapsulation of microbial inoculants for better soil–plant fertilization. A Review. Agron. Sustain. Dev. 2013, 33, 751–765. [Google Scholar] [CrossRef]
- Kebert, M.; Kostić, S.; Vuksanović, V.; Markić, A.G.; Kiprovski, B.; Zorić, M.; Orlović, S. Metal- and organ-specific response to heavy metal-induced stress mediated by antioxidant enzymes’ activities, polyamines, and plant hormones levels in Populus deltoides. Plants 2022, 11, 3246. [Google Scholar] [CrossRef] [PubMed]
- Haseeb, M.; Iqbal, S.; Hafeez, M.B.; Baloch, H.; Zahra, N.; Mumtaz, M.; Ahmad, G.; Fatima, E.M.; Jahanzaib; Raza, S.; et al. Characterizing of heavy metal accumulation, translocation and yield response traits of Chenopodium quinoa. J. Agric. Food Res. 2023, 14, 100741. [Google Scholar] [CrossRef]
- Muhammad, H.; Basra, S.M.; Irfan, A.; Abdul, W. Quinoa response to lead: Growth lead partitioning. Int. J. Agric. Biol. 2018, 20, 338–344. [Google Scholar] [CrossRef]
- Kumar, V.; Pandita, S.; Sidhu, G.P.S.; Sharma, A.; Khanna, K.; Kaur, P.; Bali, A.S.; Setia, R. Copper bioavailability, uptake, toxicity and tolerance in plants: A comprehensive review. Chemosphere 2021, 262, 127810. [Google Scholar] [CrossRef]
- Pizarro, I.; Gomez, M.M.; Cámara, C.; Palacios, M.A. Distribution of arsenic species in environmental samples collected in Northern Chile. Int. J. Environ. Anal. Chem. 2003, 83, 879–890. [Google Scholar] [CrossRef]
- Shabbir, A.; Abbas, G.; Asad, S.A.; Razzaq, H.; Anwar-Ul-Haq, M.; Amjad, M. Effects of arsenite on physiological, biochemical and grain yield attributes of quinoa (Chenopodium quinoa Willd.): Implications for phytoremediation and health risk assessment. Int. J. Phytoremediat. 2021, 23, 890–898. [Google Scholar] [CrossRef]
- Shilev, S.; Babrikova, I.; Babrikov, T. Consortium of plant growth-promoting bacteria improves spinach (Spinacea oleracea L.) growth under heavy metal stress conditions. J. Chem. Technol. Biotechnol. 2019, 95, 932–939. [Google Scholar] [CrossRef]
- Castro-Sowinski, S.; Herschkovitz, Y.; Okon, Y.; Jurkevitch, E. Effects of Inoculation with Plant Growth-Promoting Rhizobacteria on Resident Rhizosphere Microorganisms. FEMS Microbiol. Lett. 2007, 276, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Vannier, N.; Agler, M.; Hacquard, S. Microbiota-Mediated Disease Resistance in Plants. PLoS Pathog. 2019, 15, e1007740. [Google Scholar] [CrossRef] [PubMed]
- Mueller, U.G.; Sachs, J.L. Engineering Microbiomes to Improve Plant and Animal Health. Trends Microbiol. 2015, 23, 606–617. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Initial Concentrations | |
|---|---|
| Arsenic (As) mg Kg−1 | <0.01 |
| Copper (Cu) mg Kg−1 | 126 ± 2.99 |
| Manganese (Mn) mg Kg−1 | 2202 ± 36.37 |
| pH (H2O) | 5.58 |
| Matrix | Encapsulating Agent | Consortia | Initial Culture Total CFU | Encapsulation Efficiency (%) | Sphericity Factor | Size (mm) | Beads Swelling % |
|---|---|---|---|---|---|---|---|
| A1 | Alginate 3% Bentonite 2% | A | (9.2 ± 0.7) × 107 | 99 | 0.04 ± 0.01 | 0.7 ± 0.05 | 32.7 ± 6.4 |
| B | (9.8 ± 0.7) × 107 | 99 | 0.04 ± 0.03 | 0.7 ± 0.04 | 33.7 ± 4.4 | ||
| A2 | Alginate 3% Bentonite 2% Glycerol 3% | A | (1.6 ± 0.3) × 108 | 94 | 0.11 ± 0.02 | 1.4 ± 0.09 | 44.3 ± 6.3 |
| B | (2.6 ± 0.3) × 108 | 95 | 0.10 ± 0.02 | 1.6 ± 0.05 | 43.5 ± 2.2 | ||
| A3 | Alginate 3% Bentonite 2% LB medium 2.5% | A | (2.3 ± 0.7) × 108 | 89 | 0.04 ± 0.01 | 0.7 ± 0.05 | 38.6 ± 1.8 |
| B | (2.2 ± 0.7) × 108 | 90 | 0.04 ± 0.01 | 0.7 ± 0.05 | 35.3 ± 1.2 | ||
| A4 | Alginate 3% Molasses 3% | A | (4.6 ± 0.3) × 108 | 73 | 0.05 ± 0.01 | 0.9 ± 0.14 | 23.5 ± 1.0 |
| B | (4.8 ± 0.3) × 108 | 74 | 0.05 ± 0.02 | 0.9 ± 0.04 | 24.1 ± 0.4 |
| Metal(loid) Treatments | Formulation | Shoot Length (cm) | Flower Length (cm) | Root Dry Weight (g) | Stem Diameter (cm) | Shoot Dry Weight (g) |
|---|---|---|---|---|---|---|
| Control | Matrix A1 | 32.2 ± 0.6 a | 2.4 ± 0.7 a | 0.7 ± 0.1 a | 0.33 ± 0.2 a | 5.2 ± 0.1 a |
| Matrix A3 | 33.3 ± 0.7 a | 2.5 ± 0.4 a | 0.8 ± 0.1 a | 0.36 ± 0.4 a | 5.1 ± 0.1 a | |
| Consortium A in Matrix A1 | 37.7 ± 0.8 b | 3.4 ± 0.5 bc | 0.7 ± 0.1 a | 0.33 ± 0.2 a | 6.8 ± 0.4 b | |
| Consortium A in Matrix A3 | 39.6 ± 0.9 c | 3.9 ± 0.8 bc | 0.9 ± 0.1 a | 0.37 ± 0.4 a | 7.6 ± 0.6 c | |
| Consortium B in Matrix A1 | 37.1 ± 0.7 b | 3.2 ± 0.5 b | 0.8 ± 0.1 a | 0.34 ± 0.3 a | 5.7 ± 0.3 a | |
| Consortium B in Matrix A3 | 34.7 ± 0.8 ab | 4.2 ± 0.5 c | 0.6 ± 0.1 a | 0.31 ± 0.2 a | 5.2 ± 0.1 a | |
| Metal(loid) mix I | Matrix A1 | 30.7 ± 0.5 b | 2.3 ± 0.3 a | 0.3 ± 0.1 a | 0.26 ± 0.5 a | 5.7 ± 0.8 a |
| Matrix A3 | 28.5 ± 0.5 a | 3.5 ± 0.3 bc | 0.4 ± 0.1 ab | 0.22 ± 0.5 a | 5.4 ± 0.3 a | |
| Consortium A in Matrix A1 | 31.8 ± 0.6 b | 3.5 ± 0.2 bc | 0.5 ± 0.2 b | 0.35 ± 0.3 a | 6.8 ± 0.4 b | |
| Consortium A in Matrix A3 | 36.2 ± 0.7 d | 4.1 ± 0.6 cd | 0.5 ± 0.2 b | 0.35 ± 0.5 a | 7.5 ± 0.4 b | |
| Consortium B in Matrix A1 | 34.1 ± 0.7 c | 4.6 ± 0.5 d | 0.4 ± 0.2 ab | 0.30 ± 0.5 a | 5.9 ± 0.5 a | |
| Consortium B in Matrix A3 | 32.7 ± 0.7 b | 4.0 ± 0.2 cd | 0.6 ± 0.2 b | 0.31 ± 0.2 a | 5.9 ± 0.4 a | |
| Metal(loid) mix II | Matrix A1 | 17.3 ± 0.6 a | 4.1 ± 0.7 a | 0.4 ± 0.1 a | 0.19 ± 0.5 a | 2.0 ± 0.2 a |
| Matrix A3 | 17.1 ± 0.7 a | 3.8 ± 0.6 a | 0.6 ± 0.1 a | 0.23 ± 0.5 ab | 2.2 ± 0.5 a | |
| Consortium A in Matrix A1 | 26.9 ± 0.8 b | 4.7 ± 0.8 a | 0.4 ± 0.1 a | 0.28 ± 0.2 b | 2.7 ± 0.7 ab | |
| Consortium A in Matrix A3 | 30.1 ± 0.3 c | 4.9 ± 0.6 a | 0.8 ± 0.1 a | 0.25 ± 0.1 ab | 3.4 ± 0.8 bc | |
| Consortium B in Matrix A1 | 29.2 ± 0.7 c | 5.6 ± 0.1 a | 0.3 ± 0.0 a | 0.29 ± 0.5 b | 4.2 ± 0.7 d | |
| Consortium B in Matrix A3 | 28.9 ± 0.8 c | 4.9 ± 0.5 a | 0.4 ± 0.1 a | 0.27 ± 0.4 b | 3.2 ± 0.4 bc |
| Formulation | Metal Mixes mg L−1 | mg Cu kg−1 | mg As kg−1 | mg Mn kg−1 |
|---|---|---|---|---|
| Consortium A in Matrix A1 | Control | 7.75 | ND | 213.70 |
| Metal(loid) Mix I | 8.61 | ND | 354.57 | |
| Metal(loid) Mix II | 50.72 | ND | 430.71 | |
| Consortium A in Matrix A3 | Control | 7.89 | ND | 239.36 |
| Metal(loid) Mix I | 6.38 | ND | 294.16 | |
| Metal(loid) Mix II | 59.87 | ND | 305.33 | |
| Consortium B in Matrix A1 | Control | 6.57 | ND | 258.67 |
| Metal(loid) Mix I | 7.14 | ND | 339.44 | |
| Metal(loid) Mix II | 32.13 | ND | 322.53 | |
| Consortium B in Matrix A3 | Control | 6.11 | ND | 171.92 |
| Metal(loid) Mix I | 9.71 | ND | 331.97 | |
| Metal(loid) Mix II | 38.91 | ND | 357.73 | |
| Matrix A1 | Control | 9.65 | ND | 291.65 |
| Metal(loid) Mix I | 9.65 | ND | 282.13 | |
| Metal(loid) Mix II | 39.74 | ND | 335.62 | |
| Matrix A3 | Control | 7.63 | ND | 260.46 |
| Metal(loid) Mix I | 6.58 | ND | 375.45 | |
| Metal(loid) Mix II | 43.36 | ND | 336.52 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alvarado, R.; Arriagada-Escamilla, C.; Ortiz, J.; Campos-Vargas, R.; Cornejo, P. Alginate–Bentonite Encapsulation of Extremophillic Bacterial Consortia Enhances Chenopodium quinoa Tolerance to Metal Stress. Microorganisms 2024, 12, 2066. https://doi.org/10.3390/microorganisms12102066
Alvarado R, Arriagada-Escamilla C, Ortiz J, Campos-Vargas R, Cornejo P. Alginate–Bentonite Encapsulation of Extremophillic Bacterial Consortia Enhances Chenopodium quinoa Tolerance to Metal Stress. Microorganisms. 2024; 12(10):2066. https://doi.org/10.3390/microorganisms12102066
Chicago/Turabian StyleAlvarado, Roxana, Cesar Arriagada-Escamilla, Javier Ortiz, Reinaldo Campos-Vargas, and Pablo Cornejo. 2024. "Alginate–Bentonite Encapsulation of Extremophillic Bacterial Consortia Enhances Chenopodium quinoa Tolerance to Metal Stress" Microorganisms 12, no. 10: 2066. https://doi.org/10.3390/microorganisms12102066
APA StyleAlvarado, R., Arriagada-Escamilla, C., Ortiz, J., Campos-Vargas, R., & Cornejo, P. (2024). Alginate–Bentonite Encapsulation of Extremophillic Bacterial Consortia Enhances Chenopodium quinoa Tolerance to Metal Stress. Microorganisms, 12(10), 2066. https://doi.org/10.3390/microorganisms12102066