
Determination of the Copy Number of Porcine Endogenous Retroviruses (PERV) in Auckland Island Pigs Repeatedly Used for Clinical Xenotransplantation and Elimination of PERV-C


Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Cells
2.2. DNA Isolation
2.3. Droplet Digital PCR
2.4. Spleen and Liver Tissues from Non-Auckland Island Pigs
2.5. Tests for PERV-C

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Location (Nucleotid Number) | Accession Number | Reference |
|---|---|---|---|---|
| PERV pol1-forward | CGACTGCCCCAAGGGTTCAA | 3568–3587 | HM159246 | Yang et al., 2015 [24] |
| PERV pol2-reverse | TCTCTCCTGCAAATCTGGGCC | 3803–3783 | ||
| PERV pol probe | /56FAM/CACGTACTGGAGGAGGGTCACCTG | 3678–3655 | ||
| Pig actin forward | TAACCGATCCTTTCAAGCATTT | Krüger et al., 2020 [20] | ||
| Pig actin reverse | TGGTTTCAAAGCTTGCATCATA | |||
| Pig actin probe | /5HEX/CGTGGGGATGCTTCCTGAGAAAG | |||
| Pig GAPDH forward Pig GAPDH reverse | CCGCGATCTAATGTTCTCTTTC TTCACTCCGACCTTCACCAT | 3951–3970 4022–4001 | NC_010447.5 (396823) | Krüger et al., 2020 [20] |
| Pig GAPDH probe | /5HEX/CAGCCGCGTCCCTGAGACAC | 3991–3972 | ||
| PCR1 * PERV-C forward PERV-C reverse | CTGACCTGGATTAGAACTGG ATGTTAGAGGATGGTCCTGG | 6606–6625 6867–6886 | AM229312 | Takeuchi et al. [22], Kaulitz et al. [23] |
| PCR6 * PERV-C 2 forward PERV-C 2 reverse | CCAGGACCACCAAATAATGG AAGTTTTGCCCCCATTTTAGT | 6435–6454 6924–6944 | Kaulitz et al. [23] | |
| Real-time PCR * PERV-C 3 forward PERV-C 3 reverse PERV-C probe | CCCCAACCCAAGGACCAG AAGTTTTGCCCCCATTTTAGT FAM-CTCTAACATAACTTCTGGATCAGACCC- BHQ1 | 6853–6870 6924–6944 6878–6904 |
2.6. Tests for Additional Porcine Viruses
3. Results
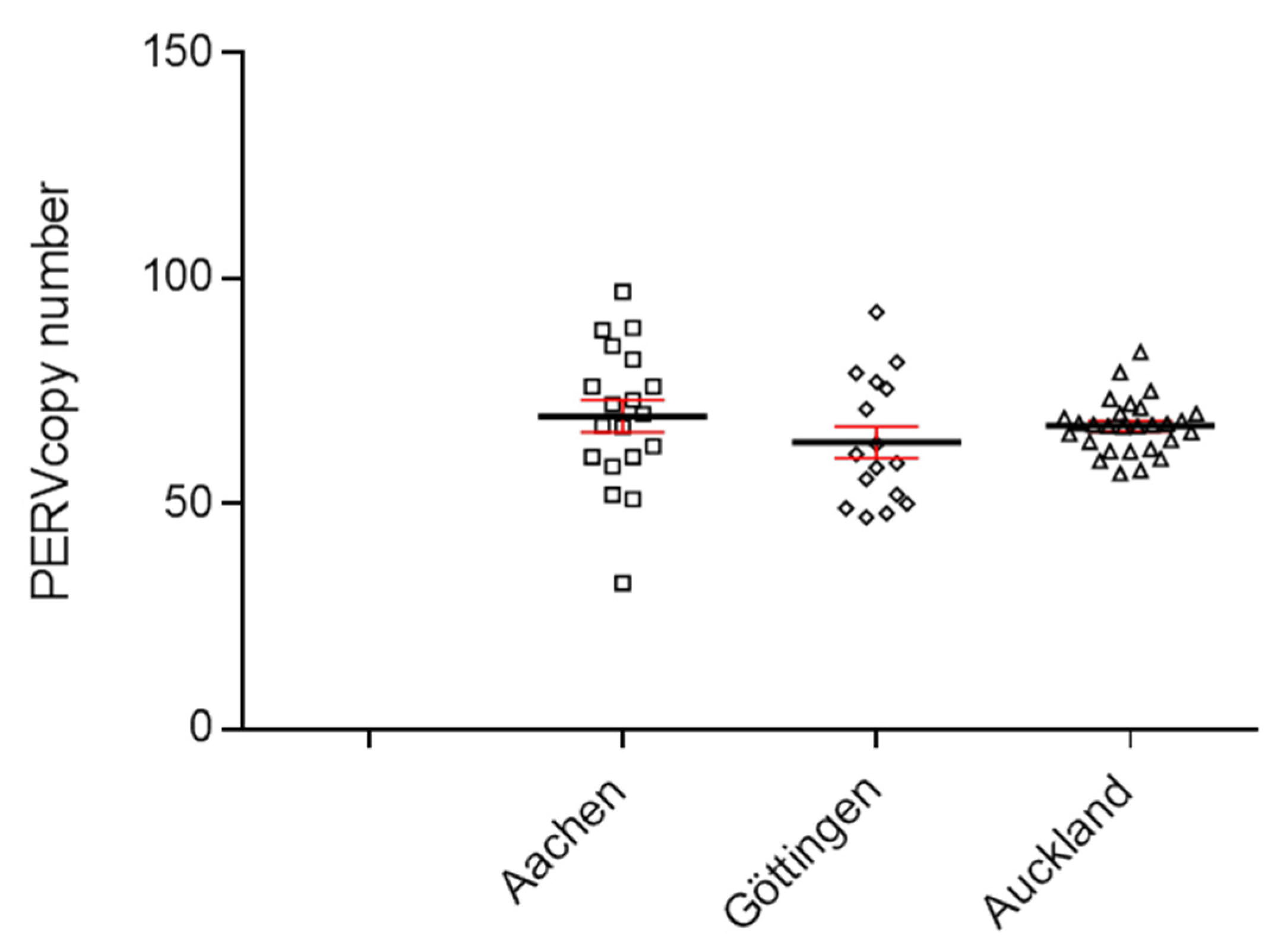
3.1. PERV Copy Number in the Genome of Adult Auckland Island Pigs
3.2. Selection of PERV-C-Negative Animals
3.3. PERV Copy Number in the Genome of Cell Lines from Very Young Piglets
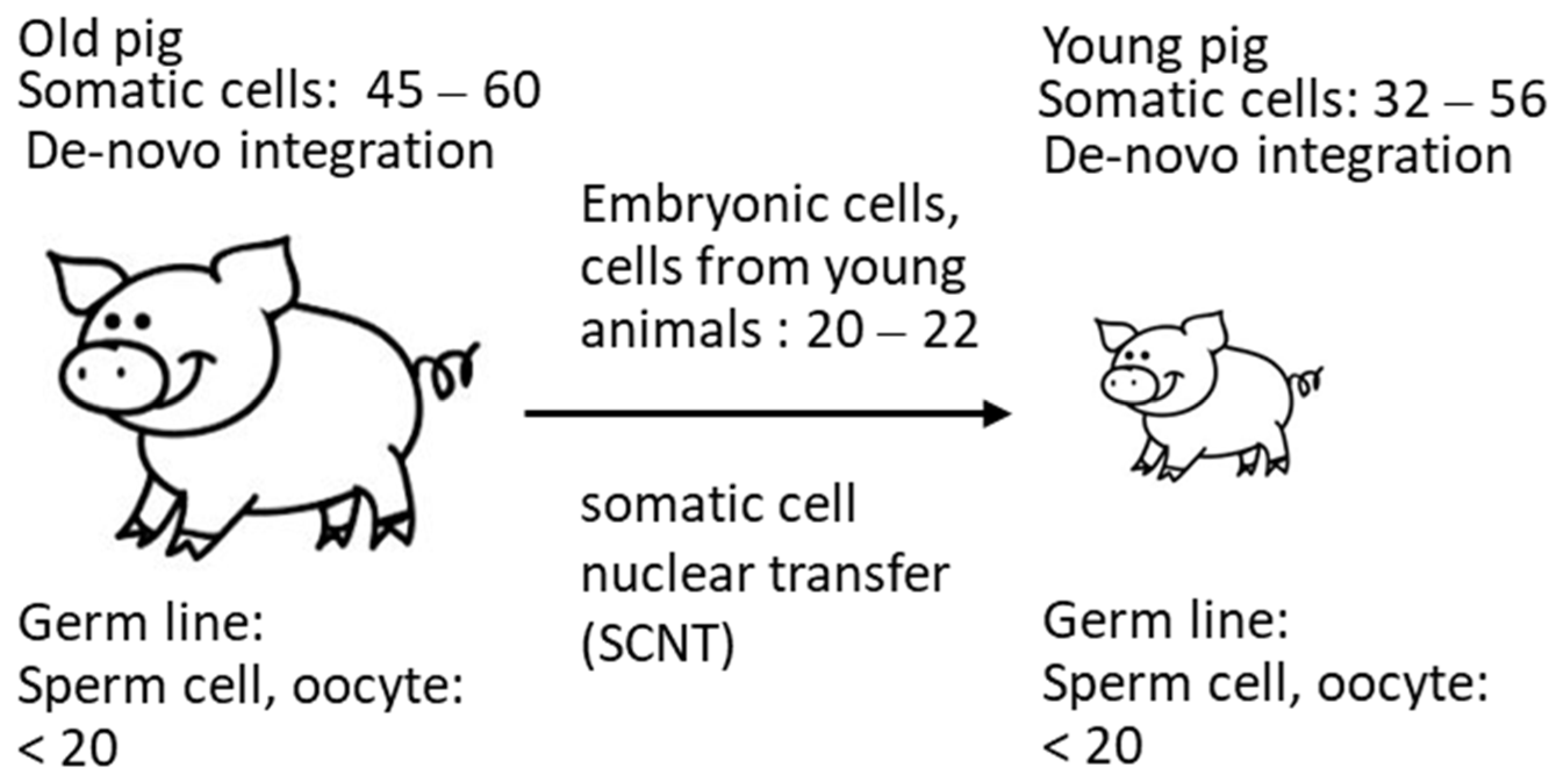
3.4. PERV Copy Number in the Genome of Auckland Island Pigs Obtained by SCNT
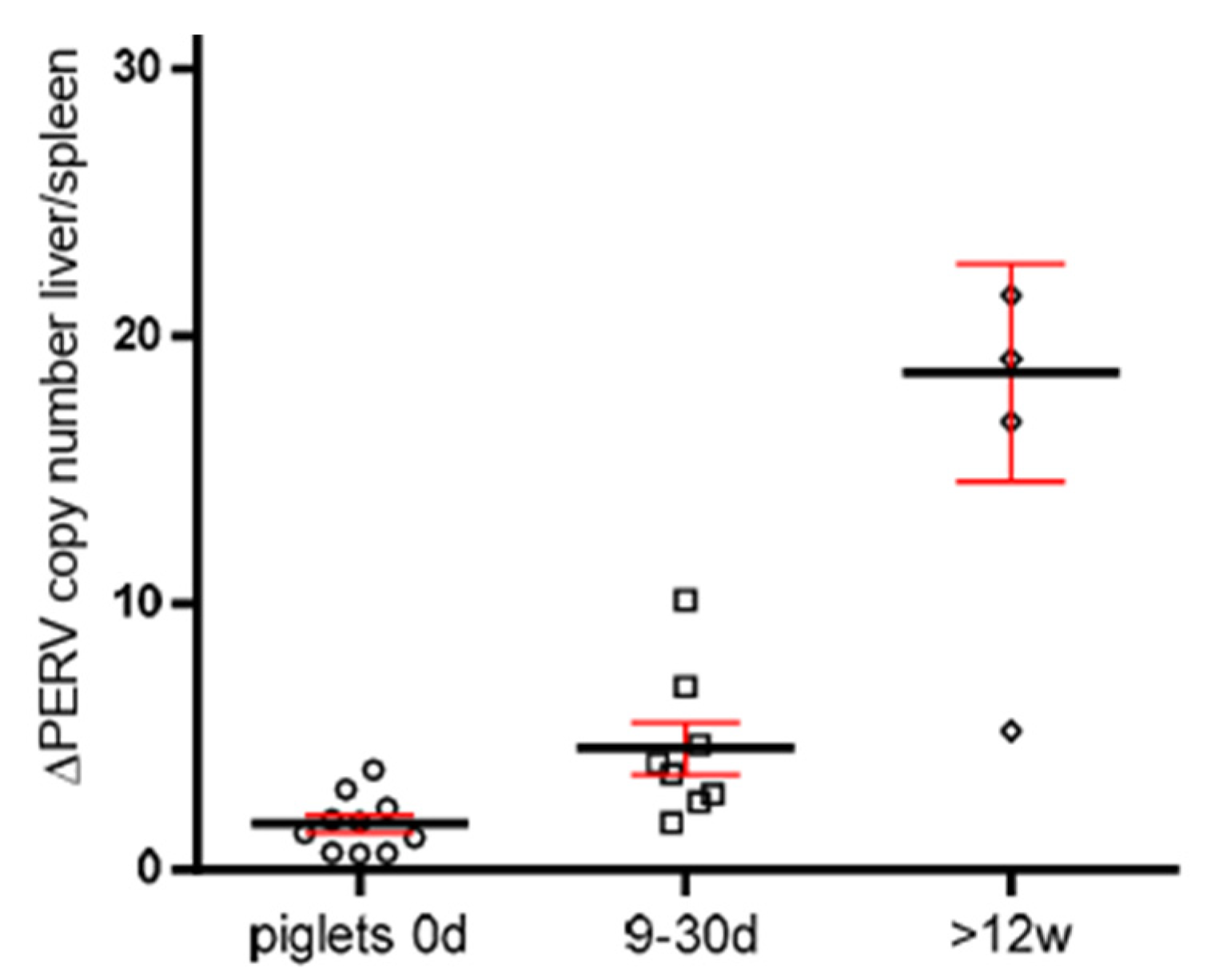
3.5. Increase in the PERV Copy Number with Age in Non-Auckland Island Pigs
3.6. Further Virological Characterization of the Auckland Island Pigs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gongora, J.; Garkavenko, O.; Moran, C. Origins of Kune Kune and Auckland Island pigs in New Zealand. In Proceedings of the 7th World Congress on Genetic Applied to Livestock Production, Montpellier, France, 19–23 August 2002. [Google Scholar]
- Robins, J.H.; Matisoo-Smith, E.; Ross, H.A. The origins of the feral pigs on the Auckland Islands. J. R. Soc. N. Z. 2003, 33, 561–569. [Google Scholar] [CrossRef]
- Fan, B.; Gongora, J.; Chen, Y.; Garkavenko, O.; Li Moran, C. Population genetic variability and origin of Auckland Island feral pigs. J. R. Soc. N. Z. 2005, 35, 279–285. [Google Scholar] [CrossRef]
- Garkavenko, O.; Muzina, M.; Muzina, Z.; Powels, K.; Elliott, R.B.; Croxson, M.C. Monitoring for potentially xenozoonotic viruses in New Zealand pigs. J. Med. Virol. 2004, 72, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Garkavenko, O.; Wynyard, S.; Nathu, D.; Simond, D.; Muzina, M.; Muzina, Z.; Scobie, L.; Hector, R.D.; Croxson, M.C.; Tan, P.; et al. Porcine endogenous retrovirus (PERV) and its transmission characteristics: A study of the New Zealand designated pathogen-free herd. Cell Transplant. 2008, 17, 1381–1388. [Google Scholar] [CrossRef] [PubMed]
- Garkavenko, O.; Dieckhoff, B.; Wynyard, S.; Denner, J.; Elliott, R.B.; Tan, P.L.; Croxson, M.C. Absence of transmission of potentially xenotic viruses in a prospective pig to primate islet xenotransplantation study. J. Med. Virol. 2008, 80, 2046–2052. [Google Scholar] [CrossRef] [PubMed]
- Wynyard, S.; Nathu, D.; Garkavenko, O.; Denner, J.; Elliott, R. Microbiological safety of the first clinical pig islet xenotransplantation trial in New Zealand. Xenotransplantation 2014, 21, 309–323. [Google Scholar] [CrossRef] [PubMed]
- Morozov, V.A.; Wynyard, S.; Matsumoto, S.; Abalovich, A.; Denner, J.; Elliott, R. No PERV transmission during a clinical trial of pig islet cell transplantation. Virus Res. 2017, 227, 34–40. [Google Scholar] [CrossRef]
- Denner, J.; Längin, M.; Reichart, B.; Krüger, L.; Fiebig, U.; Mokelke, M.; Radan, J.; Mayr, T.; Milusev, A.; Luther, F.; et al. Impact of porcine cytomegalovirus on long-term orthotopic cardiac xenotransplant survival. Sci. Rep. 2020, 10, 17531. [Google Scholar] [CrossRef]
- Yamada, K.; Tasaki, M.; Sekijima, M.; Wilkinson, R.A.; Villani, V.; Moran, S.G.; Cormack, T.A.; Hanekamp, I.M.; Hawley, R.J.; Arn, J.S.; et al. Porcine cytomegalovirus infection is associated with early rejection of kidney grafts in a pig to baboon xenotransplantation model. Transplantation 2014, 98, 411–418. [Google Scholar] [CrossRef]
- Sekijima, M.; Waki, S.; Sahara, H.; Tasaki, M.; Wilkinson, R.A.; Villani, V.; Shimatsu, Y.; Nakano, K.; Matsunari, H.; Nagashima, H.; et al. Results of life-supporting galactosyltransferase knockout kidneys in cynomolgus monkeys using two different sources of galactosyltransferase knockout Swine. Transplantation 2014, 98, 419–426. [Google Scholar] [CrossRef]
- Griffith, B.P.; Goerlich, C.E.; Singh, A.K.; Rothblatt, M.; Lau, C.L.; Shah, A.; Lorber, M.; Grazioli, A.; Saharia, K.K.; Hong, S.N.; et al. Genetically Modified Porcine-to-Human Cardiac Xenotransplantation. N. Engl. J. Med. 2022, 387, 35–44. [Google Scholar] [CrossRef]
- Mohiuddin, M.M.; Singh, A.K.; Scobie, L.; Goerlich, C.E.; Grazioli, A.; Saharia, K.; Crossan, C.; Burke, A.; Drachenberg, C.; Oguz, C.; et al. Graft dysfunction in compassionate use of genetically engineered pig-to-human cardiac xenotransplantation: A case report. Lancet 2023, 402, 397–410. [Google Scholar] [CrossRef] [PubMed]
- Denner, J.; Specke, V.; Thiesen, U.; Karlas, A.; Kurth, R. Genetic alterations of the long terminal repeat of an ecotropic porcine endogenous retrovirus during passage in human cells. Virology 2003, 314, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Harrison, I.; Takeuchi, Y.; Bartosch, B.; Stoye, J.P. Determinants of high titer in recombinant porcine endogenous retroviruses. J. Virol. 2004, 78, 13871–13879. [Google Scholar] [CrossRef]
- Wilson, C.A.; Wong, S.; Muller, J.; Davidson, C.E.; Rose, T.M.; Burd, P. Type C retrovirus released from porcine primary peripheral blood mononuclear cells infects human cells. J. Virol. 1998, 72, 3082–3087. [Google Scholar] [CrossRef] [PubMed]
- Krüger, L.; Kristiansen, Y.; Reuber, E.; Möller, L.; Laue, M.; Reimer, C.; Denner, J. A comprehensive strategy for screening for xenotransplantation-relevant viruses in a second isolated population of Göttingen Minipigs. Viruses 2019, 12, 38. [Google Scholar] [CrossRef]
- Halecker, S.; Krabben, L.; Kristiansen, Y.; Krüger, L.; Möller, L.; Becher, D.; Laue, M.; Kaufer, B.; Reimer, C.; Denner, J. Rare isolation of human-tropic recombinant porcine endogenous retroviruses PERV-A/C from Göttingen minipigs. Virol. J. 2022, 19, 30. [Google Scholar] [CrossRef]
- Pal, N.; Baker, R.; Schalk, S.; Scobie, L.; Tucker, A.W.; Opriessnig, T. Detection of porcine endogenous retrovirus (PERV) viremia in diseased versus healthy US pigs by qualitative and quantitative real-time RT-PCR. Transbound Emerg. Dis. 2011, 58, 344–351. [Google Scholar] [CrossRef]
- Krüger, L.; Stillfried, M.; Prinz, C.; Schröder, V.; Neubert, L.K.; Denner, J. Copy Number and Prevalence of Porcine Endogenous Retroviruses (PERVs) in German Wild Boars. Viruses 2020, 12, 419. [Google Scholar] [CrossRef]
- Fiebig, U.; Fischer, K.; Bähr, A.; Runge, C.; Schnieke, A.; Wolf, E.; Denner, J. Porcine endogenous retroviruses: Quantification of the copy number in cell lines, pig breeds, and organs. Xenotransplantation 2018, 25, e12445. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Patience, C.; Magre, S.; Weiss, R.A.; Banerjee, P.T.; Le Tissier, P.; Stoye, J.P. Host range and interference studies of three classes of pig endogenous retrovirus. J. Virol. 1998, 72, 9986–9991. [Google Scholar] [CrossRef] [PubMed]
- Kaulitz, D.; Mihica, D.; Adlhoch, C.; Semaan, M.; Denner, J. Improved pig donor screening including newly identified variants of porcine endogenous retrovirus-C (PERV-C). Arch. Virol. 2013, 158, 341–348. [Google Scholar] [CrossRef]
- Yang, L.; Güell, M.; Niu, D.; George, H.; Lesha, E.; Grishin, D.; Aach, J.; Shrock, E.; Xu, W.; Poci, J.; et al. Genome-wide inactivation of porcine endogenous retroviruses (PERVs). Science 2015, 350, 1101–1104. [Google Scholar] [CrossRef] [PubMed]
- Morozov, V.A.; Morozov, A.V.; Denner, J. New PCR diagnostic systems for the detection and quantification of porcine cytomegalovirus (PCMV). Arch. Virol. 2016, 161, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Heinze, J.; Plotzki, E.; Denner, J. Virus Safety of Xenotransplantation: Prevalence of Porcine Cicrovirus 2 (PCV2) in Pigs. Ann. Virol. Res. 2016, 2, 1023. [Google Scholar]
- Prinz, C.; Stillfried, M.; Neubert, L.K.; Denner, J. Detection of PCV3 in German wild boars. Virol. J. 2019, 16, 25. [Google Scholar] [CrossRef] [PubMed]
- Morozov, V.A.; Morozov, A.V.; Rotem, A.; Barkai, U.; Bornstein, S.; Denner, J. Extended Microbiological Characterization of Göttingen Minipigs in the Context of Xenotransplantation: Detection and Vertical Transmission of Hepatitis E Virus. PLoS ONE 2015, 10, e0139893. [Google Scholar] [CrossRef]
- Denner, J. How Active Are Porcine Endogenous Retroviruses (PERVs)? Viruses 2016, 8, 215. [Google Scholar] [CrossRef]
- Denner, J. Porcine Lymphotropic Herpesviruses (PLHVs) and Xenotransplantation. Viruses 2021, 13, 1072. [Google Scholar] [CrossRef]
- Wynyard, S.; Garkavenko, O.; Elliot, R. Multiplex high resolution melting assay for estimation of Porcine Endogenous Retrovirus (PERV) relative gene dosage in pigs and detection of PERV infection in xenograft recipients. J. Virol. Methods 2011, 175, 95–100. [Google Scholar] [CrossRef]
- Mourad, N.I.; Crossan, C.; Cruikshank, V.; Scobie, L.; Gianello, P. Characterization of porcine endogenous retrovirus expression in neonatal and adult pig pancreatic islets. Xenotransplantation 2017, 24, e12311. [Google Scholar] [CrossRef] [PubMed]
- Bartosch, B.; Stefanidis, D.; Myers, R.; Weiss, R.; Patience, C.; Takeuchi, Y. Evidence and consequence of porcine endogenous retrovirus recombination. J. Virol. 2004, 78, 13880–13890. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Q.; Zhang, M.P.; Tong, X.K.; Li, J.Q.; Zhang, Z.; Huang, F.; Du, H.P.; Zhou, M.; Ai, H.S.; Huang, L.S. Scan of the endogenous retrovirus sequences across the swine genome and survey of their copy number variation and sequence diversity among various Chinese and Western pig breeds. Zool Res. 2022, 43, 423–441. [Google Scholar] [CrossRef] [PubMed]
- Le Tissier, P.; Stoye, J.P.; Takeuchi, Y.; Patience, C.; Weiss, R.A. Two sets of human-tropic pig retrovirus. Nature 1997, 389, 681–682. [Google Scholar] [CrossRef] [PubMed]
- Patience, C.; Takeuchi, Y.; Weiss, R.A. Infection of human cells by an endogenous retrovirus of pigs. Nat. Med. 1997, 3, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Patience, C.; Switzer, W.M.; Takeuchi, Y.; Griffiths, D.J.; Goward, M.E.; Heneine, W.; Stoye, J.P.; Weiss, R.A. Multiple groups of novel retroviral genomes in pigs and related species. J. Virol. 2001, 75, 2771–2775. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Li, Z.; Pan, M.; Ge, M.; Wang, Y.; Gao, Y. Genetic prevalence of porcine endogenous retrovirus in Chinese experimental miniature pigs. Transplant. Proc. 2011, 43, 2762–2769. [Google Scholar] [CrossRef]
- Lee, D.; Lee, J.; Yoon, J.K.; Kim, N.Y.; Kim, G.W.; Park, C.; Oh, Y.K.; Kim, Y.B. Rapid determination of PERV copy number from porcine genomic DNA by real-time polymerase chain reaction. Anim. Biotechnol. 2011, 22, 175–180. [Google Scholar] [CrossRef]
- Yoon, J.K.; Choi, J.; Lee, H.J.; Cho, Y.; Gwon, Y.D.; Jang, Y.; Kim, S.; Choi, H.; Lee, J.H.; Kim, Y.B. Distribution of Porcine Endogenous Retrovirus in Different Organs of the Hybrid of a Landrace and a Jeju Domestic Pig in Korea. Transplant. Proc. 2015, 47, 2067–2071. [Google Scholar] [CrossRef]
- Zhang, P.; Yu, P.; Wang, W.; Zhang, L.; Li, S.; Bu, H. An effective method for the quantitative detection of porcine endogenous retrovirus in pig tissues. In Vitro Cell. Dev. Biol. Anim. 2010, 46, 408–410. [Google Scholar] [CrossRef]
- Quereda, J.J.; Herrero-Medrano, J.M.; Abellaneda, J.M.; García-Nicolás, O.; Martínez-Alarcón, L.; Pallarés, F.J.; Ramírez, P.; Muñoz, A.; Ramis, G. Porcine endogenous retrovirus copy number in different pig breeds is not related to genetic diversity. Zoonoses Public Health 2012, 59, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Mang, R.; Maas, J.; Chen, X.; Goudsmit, J.; van der Kuyl, A.C. Identification of a novel type C porcine endogenous retrovirus: Evidence that copy number of endogenous retroviruses increases during host inbreeding. J. Gen. Virol. 2001, 82, 1829–1834. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Webb, G.C.; Allen, R.D.; Moran, C. Characterizing and mapping porcine endogenous retrovirusesin Westran pigs. J. Virol. 2002, 76, 5548–5556. [Google Scholar] [CrossRef] [PubMed]
- Groenen, M.A.; Archibald, A.L.; Uenishi, H.; Tuggle, C.K.; Takeuchi, Y.; Rothschild, M.F.; Rogel-Gaillard, C.; Park, C.; Milan, D.; Megens, H.J.; et al. Analyses of pig genomes provide insight into porcine demography and evolution. Nature 2012, 491, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, L.B.; Coleman, V.A.; Hindson, C.M.; Herrmann, J.; Hindson, B.J.; Bhat, S.; Emslie, K.R. Evaluation of a droplet digital polymerase chain reaction format for DNA copy number quantification. Anal. Chem. 2012, 84, 1003–1011. [Google Scholar] [CrossRef] [PubMed]
- Denner, J. What does the PERV copy number tell us? Xenotransplantation 2022, 2, e12732. [Google Scholar] [CrossRef]
- Subramanian, R.P.; Wildschutte, J.H.; Russo, C.; Coffin, J.M. Identification, characterization, and comparative genomic distribution of the HERV-K (HML-2) group of human endogenous retroviruses. Retrovirology 2011, 8, 90. [Google Scholar] [CrossRef]
- Contreras-Galindo, R.; Kaplan, M.H.; He, S.; Contreras-Galindo, A.C.; Gonzalez-Hernandez, M.J.; Kappes, F.; Dube, D.; Chan, S.M.; Robinson, D.; Meng, F.; et al. HIV infection reveals widespread expansion of novel centromeric human endogenous retroviruses. Genome Res. 2013, 23, 1505–1513. [Google Scholar] [CrossRef]
- Dieckhoff, B.; Puhlmann, J.; Büscher, K.; Hafner-Marx, A.; Herbach, N.; Bannert, N.; Büttner, M.; Wanke, R.; Kurth, R.; Denner, J. Expression of porcine endogenous retroviruses (PERVs) in melanomas of Munich miniature swine (MMS) Troll. Vet. Microbiol. 2007, 123, 53–68. [Google Scholar] [CrossRef]
- Bittmann, I.; Mihica, D.; Plesker, R.; Denner, J. Expression of porcine endogenous retroviruses (PERV) in different organs of a pig. Virology 2012, 433, 329–336. [Google Scholar] [CrossRef]
- Scobie, L.; Taylor, S.; Wood, J.C.; Suling, K.M.; Quinn, G.; Meikle, S.; Patience, C.; Schuurman, H.J.; Onions, D.E. Absence of replication-competent human-tropic porcine endogenous retroviruses in the germ line DNA of inbred miniature Swine. J. Virol. 2004, 78, 2502–2509. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.C.; Quinn, G.; Suling, K.M.; Oldmixon, B.A.; Van Tine, B.A.; Cina, R.; Arn, S.; Huang, C.A.; Scobie, L.; Onions, D.E.; et al. Identification of exogenous forms of human-tropic porcine endogenous retrovirus in miniature Swine. J. Virol. 2004, 78, 2494–2501. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.I.; Wilkinson, R.; Fishman, J.A. Genomic presence of recombinant porcine endogenous retrovirus in transmitting miniature swine. Virol. J. 2006, 3, 91. [Google Scholar] [CrossRef] [PubMed]
- Hinrichs, A.; Riedel, E.O.; Klymiuk, N.; Blutke, A.; Kemter, E.; Längin, M.; Dahlhoff, M.; Keßler, B.; Kurome, M.; Zakhartchenko, V.; et al. Growth hormone receptor knockout to reduce the size of donor pigs for preclinical xenotransplantation studies. Xenotransplantation 2021, 28, e12664. [Google Scholar] [CrossRef]
- Goerlich, C.E.; Griffith, B.; Hanna, P.; Hong, S.N.; Ayares, D.; Singh, A.K.; Mohiuddin, M.M. The growth of xenotransplanted hearts can be reduced with growth hormone receptor knockout pig donors. J. Thorac. Cardiovasc. Surg. 2023, 165, e69–e81. [Google Scholar] [CrossRef]
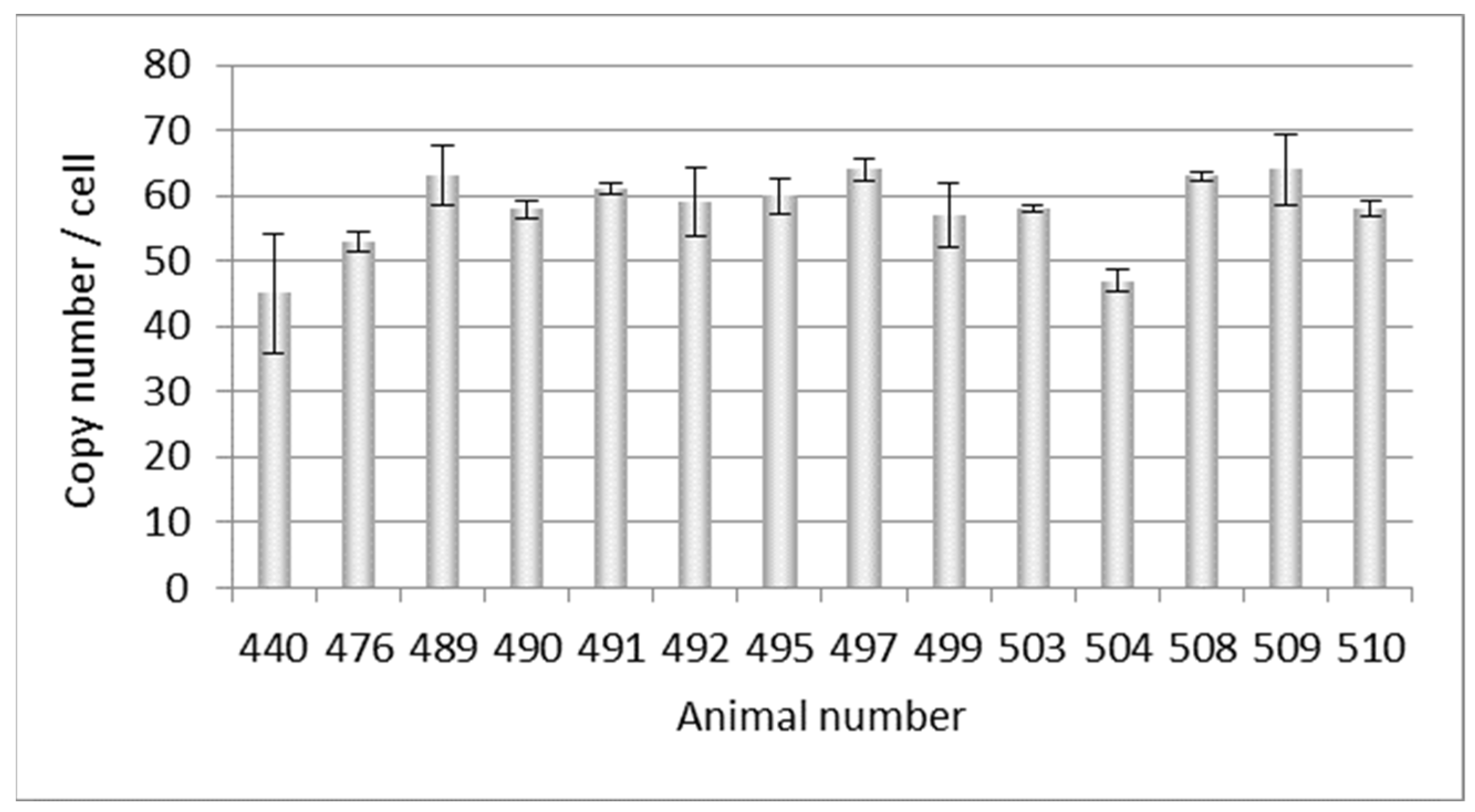

| Pig | Viruses * | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| PERV Copy Number | PERV-C | PCMV/PRV | HEV | HEV | HEV | PCV1/2 | PCV3 | PLHV-1/2 | PLHV-3 | |
| PCR | PCR | PCR | WB | ELISA | PCR | PCR | PCR | PCR | ||
| 440 | 45 | − | − | − | − | − | − | − | − | − |
| 476 | 53 | − | − | − | − | n.t. | − | − | − | + |
| 489 | 63 | + | − | − | − | − | − | − | − | − |
| 490 | 58 | + | − | − | − | n.t. | − | − | − | − |
| 491 | 61 | + | − | − | − | − | − | − | − | − |
| 492 | 59 | + | − | − | − | n.t. | − | − | − | − |
| 494 | n.t. | + | − | − | − | − | − | − | − | − |
| 495 | 60 | − | − | − | − | − | − | − | − | − |
| 497 | 64 | − | − | − | − | − | − | − | − | − |
| 499 | 57 | − | − | − | − | − | − | − | − | − |
| 503 | 58 | − | − | − | − | − | − | − | − | − |
| 504 | 47 | − | − | − | − | n.t. | − | − | − | − |
| 508 | 63 | + | − | − | − | n.t. | − | − | − | − |
| 509 | 64 | + | − | − | − | n.t. | − | − | − | − |
| 510 | 58 | + | − | − | − | n.t. | − | − | − | |
| f1 | 21 | − | − | n.t. | n.a. | n.a. | n.t. | − | n.t. | n.t. |
| f2 | 20 | − | − | n.t. | n.a. | n.a. | n.t. | − | n.t. | n.t. |
| m1 | 20 | − | − | n.t. | n.a. | n.a. | n.t. | − | n.t. | n.t. |
| m2 | 22 | − | − | n.t. | n.a. | n.a. | n.t. | − | n.t. | n.t. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fiebig, U.; Krüger, L.; Denner, J. Determination of the Copy Number of Porcine Endogenous Retroviruses (PERV) in Auckland Island Pigs Repeatedly Used for Clinical Xenotransplantation and Elimination of PERV-C. Microorganisms 2024, 12, 98. https://doi.org/10.3390/microorganisms12010098
Fiebig U, Krüger L, Denner J. Determination of the Copy Number of Porcine Endogenous Retroviruses (PERV) in Auckland Island Pigs Repeatedly Used for Clinical Xenotransplantation and Elimination of PERV-C. Microorganisms. 2024; 12(1):98. https://doi.org/10.3390/microorganisms12010098
Chicago/Turabian StyleFiebig, Uwe, Luise Krüger, and Joachim Denner. 2024. "Determination of the Copy Number of Porcine Endogenous Retroviruses (PERV) in Auckland Island Pigs Repeatedly Used for Clinical Xenotransplantation and Elimination of PERV-C" Microorganisms 12, no. 1: 98. https://doi.org/10.3390/microorganisms12010098
APA StyleFiebig, U., Krüger, L., & Denner, J. (2024). Determination of the Copy Number of Porcine Endogenous Retroviruses (PERV) in Auckland Island Pigs Repeatedly Used for Clinical Xenotransplantation and Elimination of PERV-C. Microorganisms, 12(1), 98. https://doi.org/10.3390/microorganisms12010098