
Analysis of Airborne Fungal Communities on Pedestrian Bridges in Urban Environments

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Locations
2.2. Sampling Method
2.3. Enumeration of Fungi
2.4. Fungal Identification
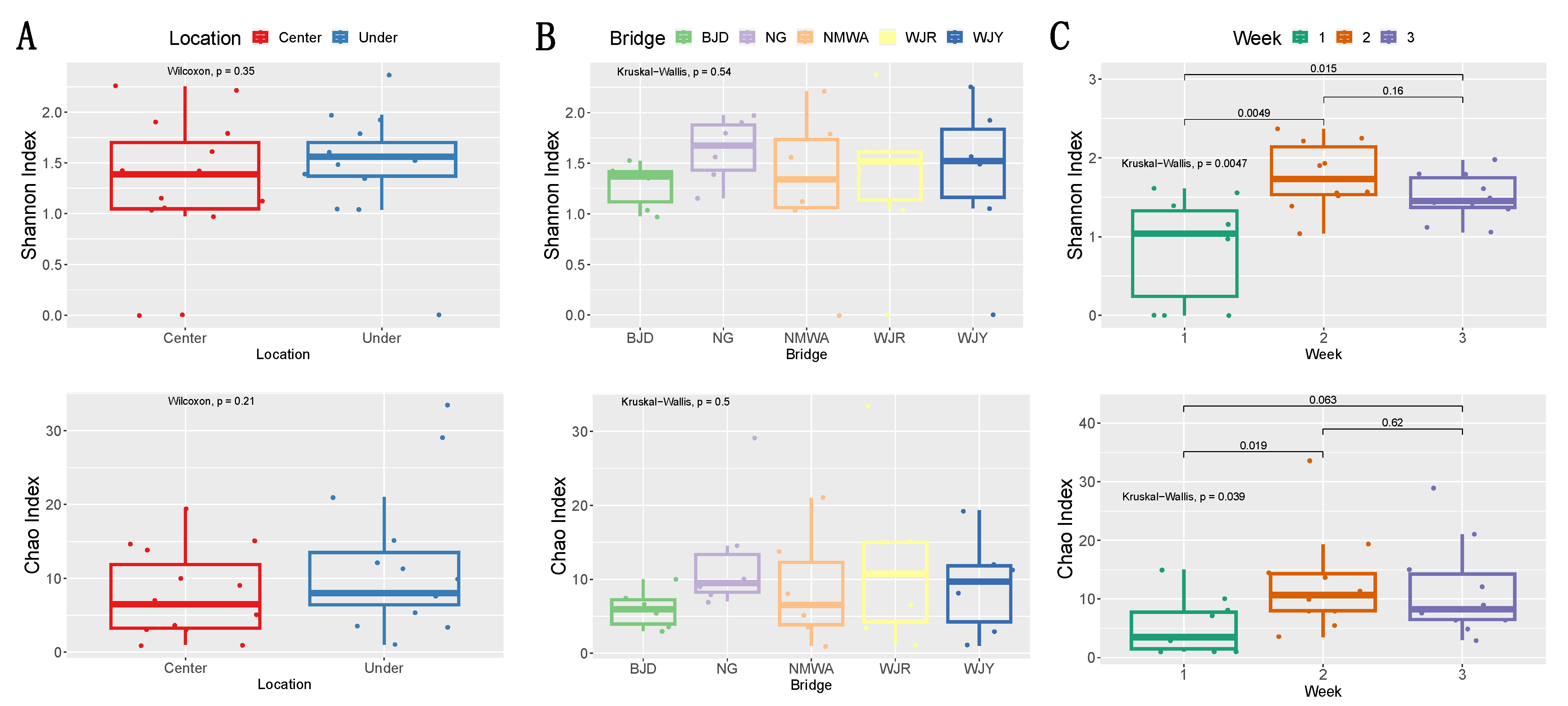
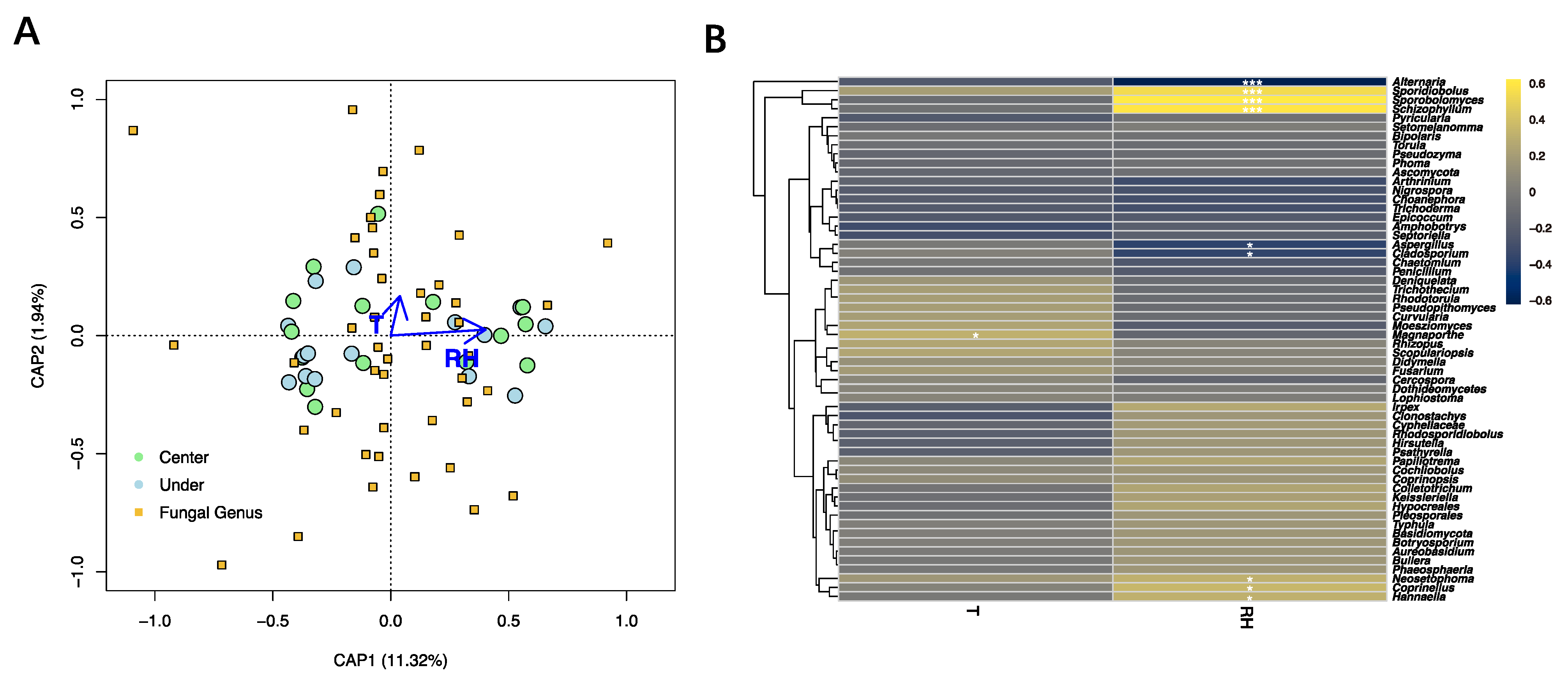
2.5. Statistical Analysis
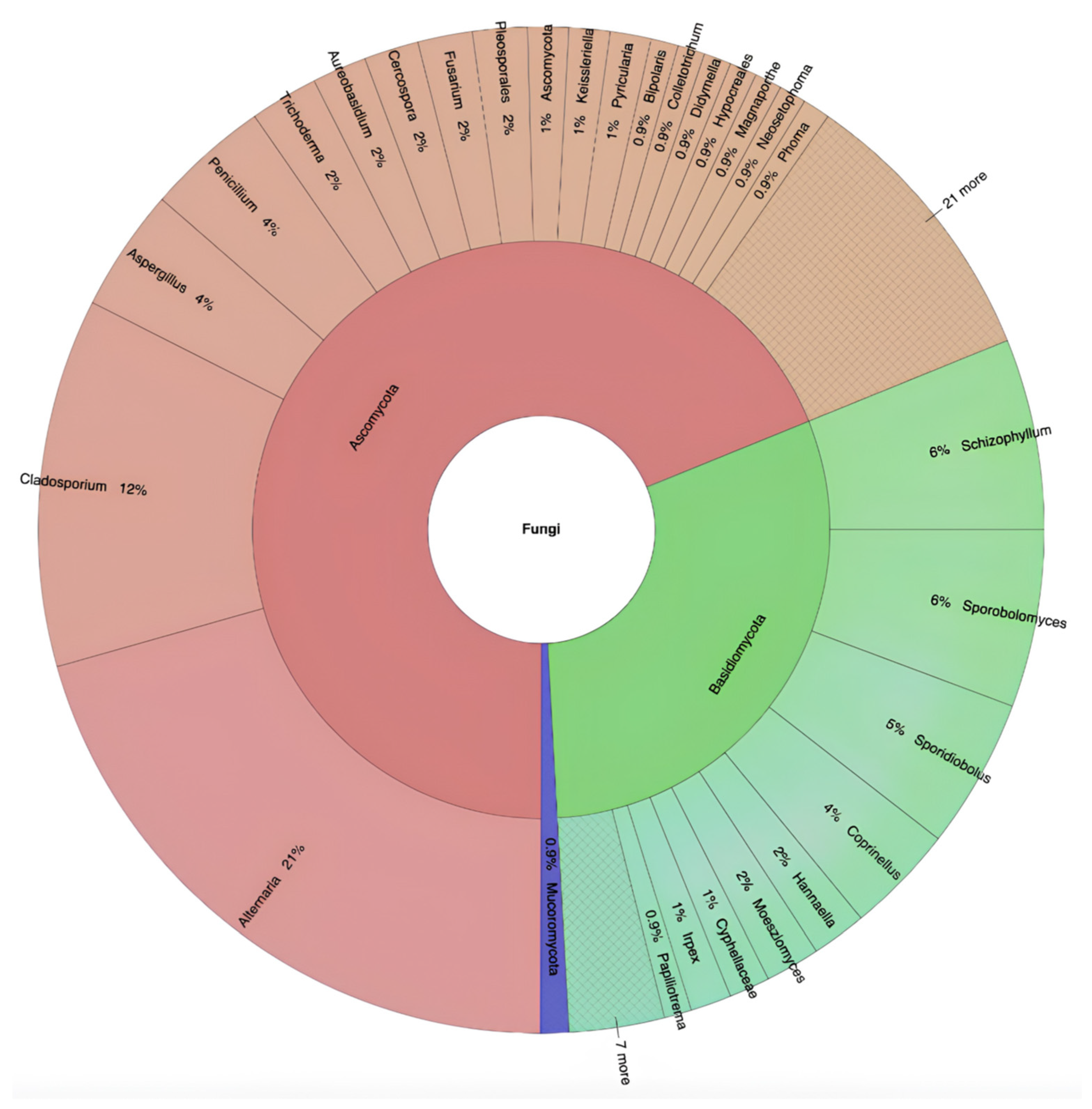
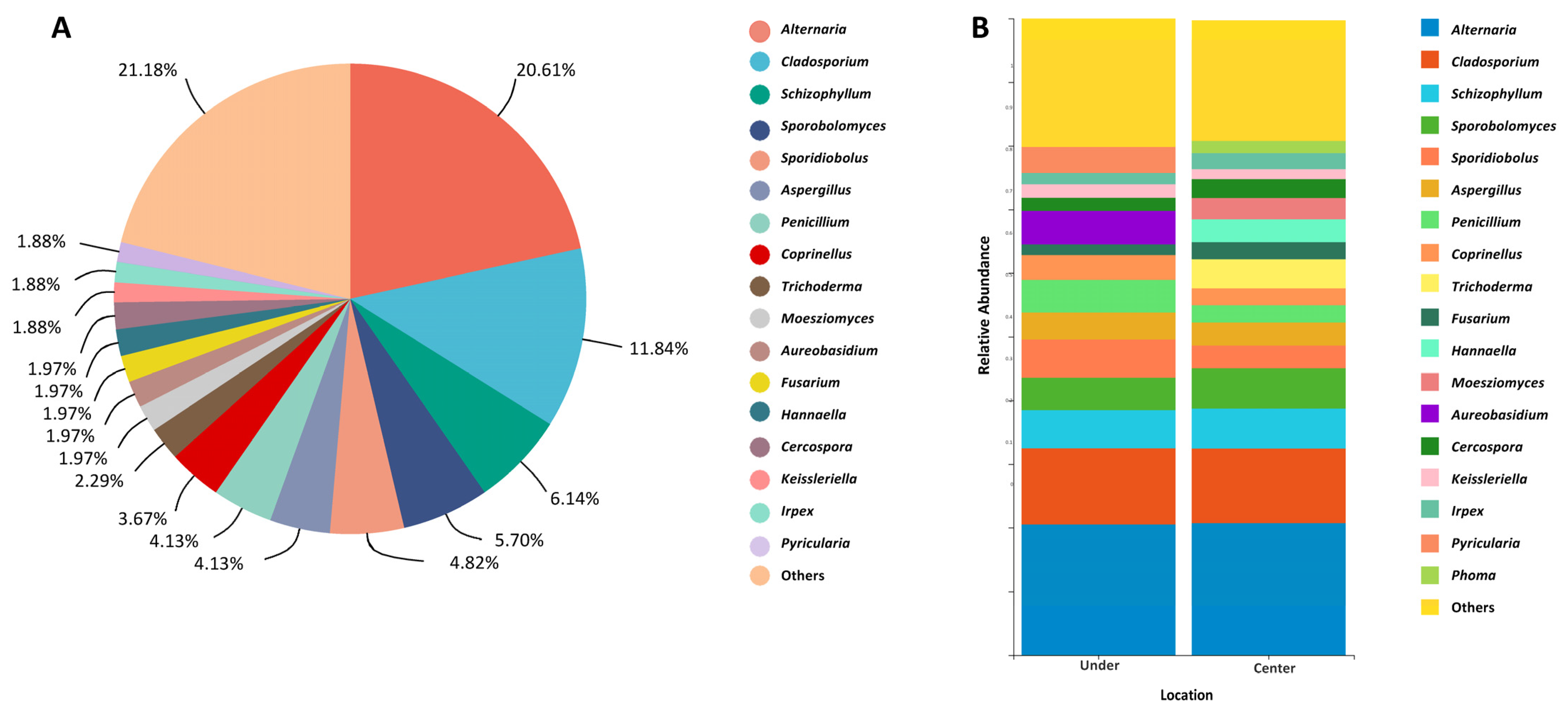
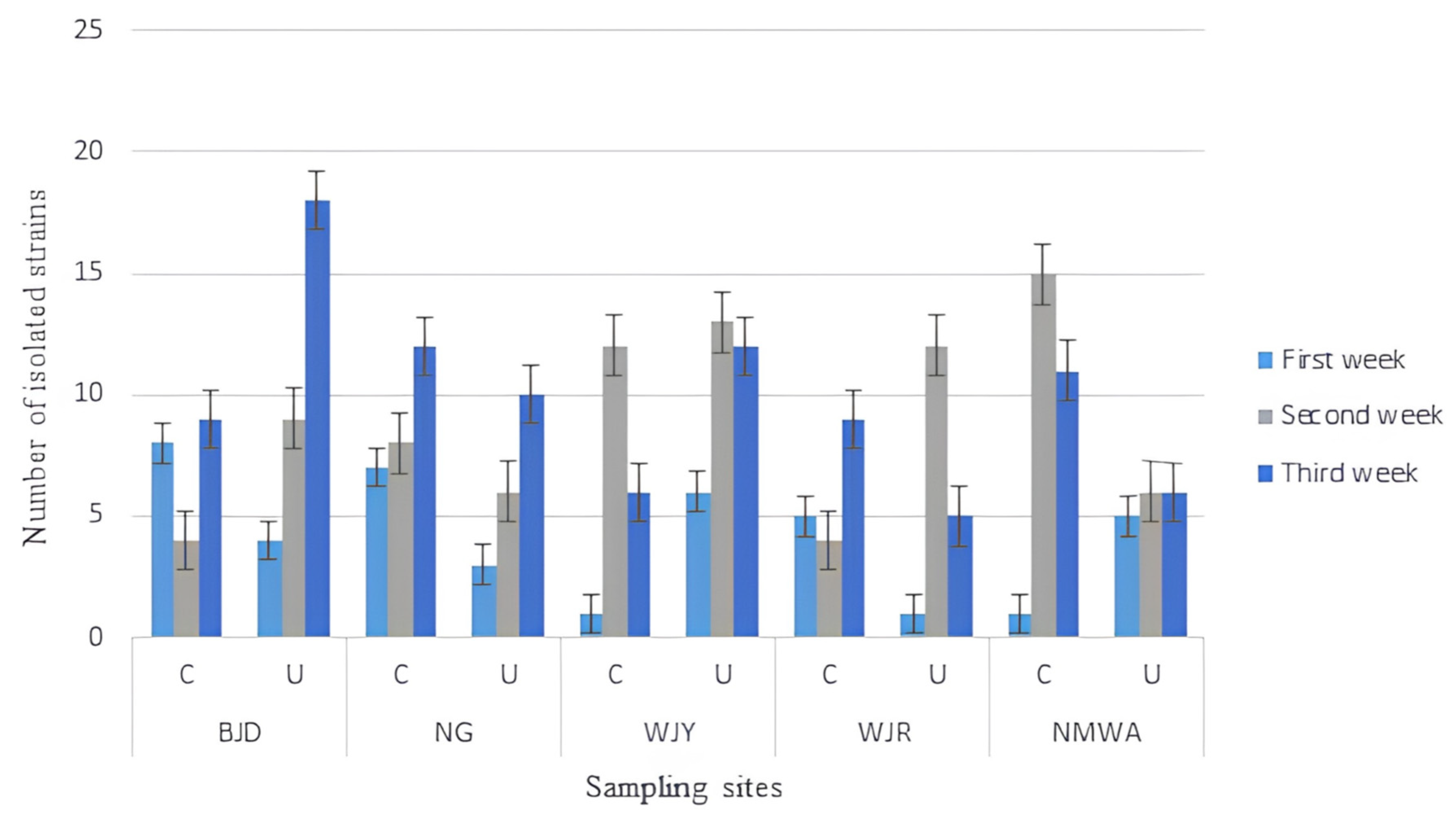
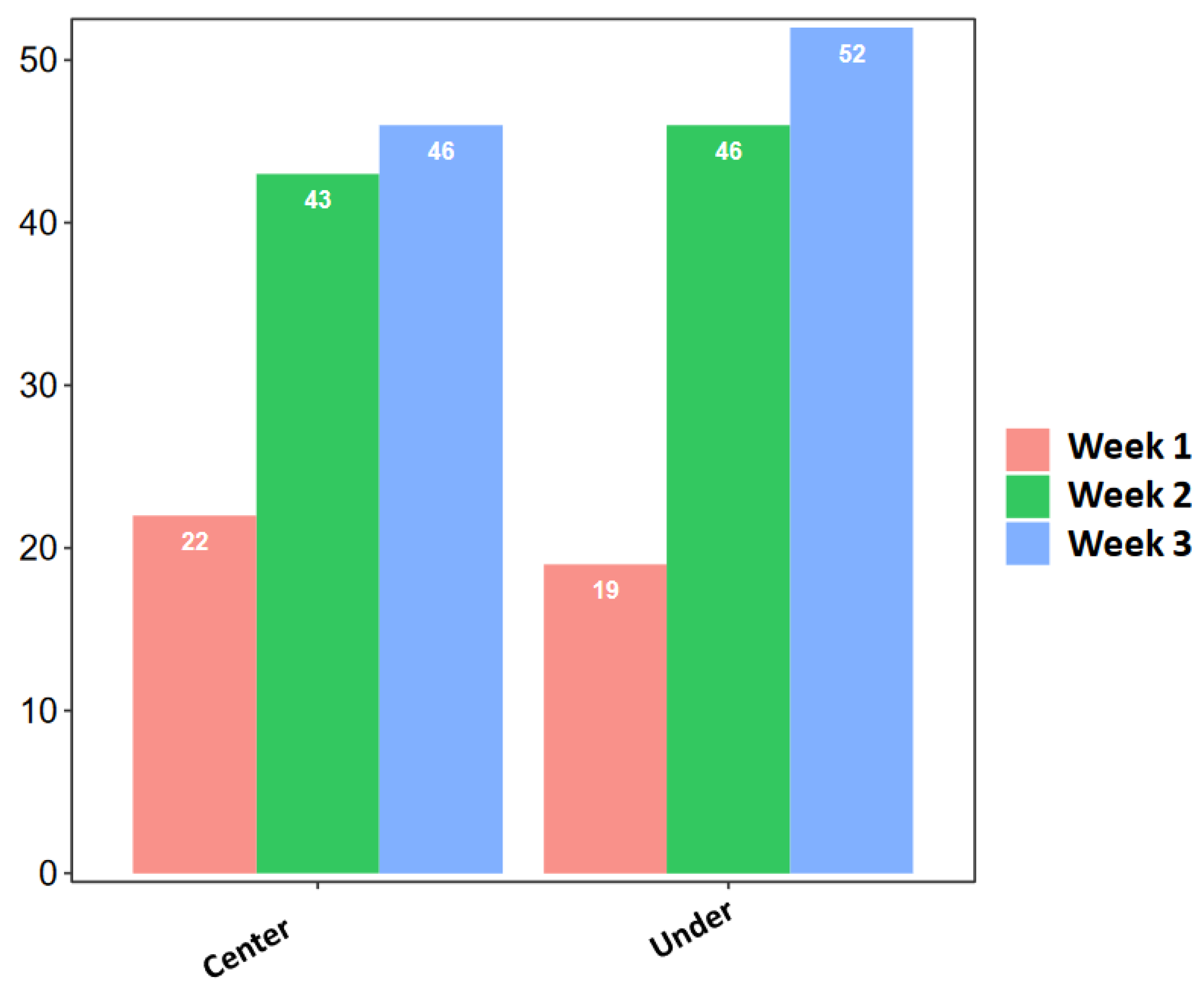
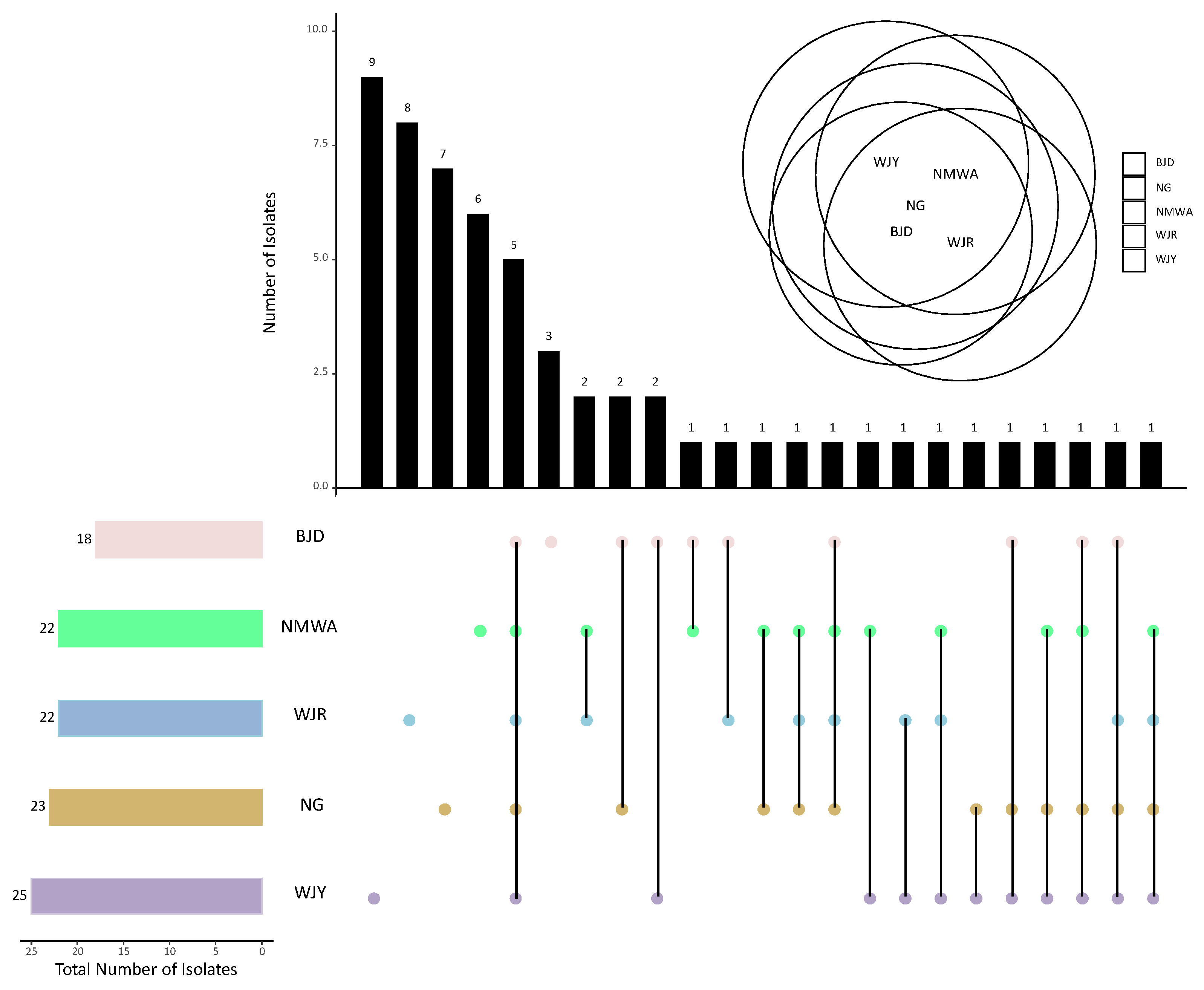
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bennett, J. Mycotechnology: The role of fungi in biotechnology. J. Biotechnol. 1998, 66, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, H.; Martinez, L.R. Relationship of environmental disturbances and the infectious potential of fungi. Microbiology 2018, 164, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.D. Fungi as contaminants in indoor air. Atmos. Environment. Part A Gen. Top. 1992, 26, 2163–2172. [Google Scholar] [CrossRef]
- Sorenson, W.G. Fungal spores: Hazardous to health? Environ. Health Perspect. 1999, 107, 469–472. [Google Scholar] [CrossRef]
- Fernstrom, A.; Goldblatt, M. Aerobiology and its role in the transmission of infectious diseases. J. Pathog. 2013, 2013, 493960. [Google Scholar] [CrossRef]
- Pei-Chih, W.; Huey-Jen, S.; Chia-Yin, L. Characteristics of indoor and outdoor airborne fungi at suburban and urban homes in two seasons. Sci. Total Environ. 2000, 253, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Horner, W.E.; Helbling, A.; Salvaggio, J.E.; Lehrer, S.B. Fungal allergens. Clin. Microbiol. Rev. 1995, 8, 161–179. [Google Scholar] [CrossRef]
- Beaumont, F. Clinical manifestations of pulmonary Aspergillus infections. Mycoses 1988, 31 (Suppl. S2), 15–20. [Google Scholar]
- Bush, R.K.; Portnoy, J.M. The role and abatement of fungal allergens in allergic diseases. J. Allergy Clin. Immunol. 2001, 107 (Suppl. S3), S430–S440. [Google Scholar] [CrossRef]
- Hargreaves, M.; Parappukkaran, S.; Morawska, L.; Hitchins, J.; He, C.; Gilbert, D. A pilot investigation into associations between indoor airborne fungal and non-biological particle concentrations in residential houses in Brisbane, Australia. Sci. Total Environ. 2003, 312, 89–101. [Google Scholar] [CrossRef]
- Saad-Hussein, A.; Ibrahim, K.S. Health Impact of Airborne Fungi. In Handbook of Healthcare in the Arab World; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1421–1435. [Google Scholar]
- Shelton, B.G.; Kirkland, K.H.; Flanders, W.D.; Morris, G.K. Profiles of airborne fungi in buildings and outdoor environments in the United States. Appl. Environ. Microbiol. 2002, 68, 1743–1753. [Google Scholar] [CrossRef] [PubMed]
- Pastuszka, J.S.; Paw, U.K.T.; Lis, D.O.; Wlazło, A.; Ulfig, K. Bacterial and fungal aerosol in indoor environment in Upper Silesia, Poland. Atmos. Environ. 2000, 34, 3833–3842. [Google Scholar] [CrossRef]
- Ahmed Adam, M.A.; Tabana, Y.M.; Musa, K.B.; Sandai, D.A. Effects of different mycotoxins on humans, cell genome and their involvement in cancer (Review). Oncol. Rep. 2017, 37, 1321–1336. [Google Scholar] [CrossRef] [PubMed]
- Dvorácková, I.; Stora, C.; Ayraud, N. Evidence of aflatoxin B1 in two cases of lung cancer in man. J. Cancer Res. Clin. Oncol. 1981, 100, 221–224. [Google Scholar] [CrossRef]
- Marchese, S.; Polo, A.; Ariano, A.; Velotto, S.; Costantini, S.; Severino, L. Aflatoxin B1 and M1: Biological Properties and Their Involvement in Cancer Development. Toxins 2018, 10, 214. [Google Scholar] [CrossRef]
- Olsen, J.H.; Dragsted, L.; Autrup, H. Cancer risk and occupational exposure to aflatoxins in Denmark. Br. J. Cancer 1988, 58, 392–396. [Google Scholar] [CrossRef]
- Adhikari, A.; Sen, M.M.; Gupta-Bhattacharya, S.; Chanda, S. Airborne viable, non-viable, and allergenic fungi in a rural agricultural area of India: A 2-year study at five outdoor sampling stations. Sci. Total Environ. 2004, 326, 123–141. [Google Scholar] [CrossRef]
- Kurup, V.P.; Shen, H.D.; Banerjee, B. Respiratory fungal allergy. Microbes Infect. 2000, 2, 1101–1110. [Google Scholar] [CrossRef]
- Douwes, J.; Thorne, P.; Pearce, N.; Heederik, D. Bioaerosol health effects and exposure assessment: Progress and prospects. Ann. Occup. Hyg. 2003, 47, 187–200. [Google Scholar]
- Eduard, W. Fungal spores: A critical review of the toxicological and epidemiological evidence as a basis for occupational exposure limit setting. Crit. Rev. Toxicol. 2009, 39, 799–864. [Google Scholar] [CrossRef]
- Pekkanen, J.; Hyvärinen, A.; Haverinen-Shaughnessy, U.; Korppi, M.; Putus, T.; Nevalainen, A. Moisture damage and childhood asthma: A population-based incident case-control study. Eur. Respir. J. 2007, 29, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Cui, Y.; Dong, J.G. Variation of airborne bacteria and fungi at Emperor Qin’s Terra-Cotta Museum, Xi’an, China, during the “Oct. 1” gold week period of 2006. Environ. Sci. Pollut. Res. Int. 2010, 17, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Fang, Z.; Zhang, J.; Guo, W.; Lou, X. Assemblages of culturable airborne fungi in a typical urban, tourism-driven center of southeast China. Aerosol Air Qual. Res. 2019, 19, 820–831. [Google Scholar] [CrossRef]
- Liu, Z.; Cheng, K.; Li, H.; Cao, G.; Wu, D.; Shi, Y. Exploring the potential relationship between indoor air quality and the concentration of airborne culturable fungi: A combined experimental and neural network modeling study. Environ. Sci. Pollut. Res. Int. 2018, 25, 3510–3517. [Google Scholar] [CrossRef] [PubMed]
- Nageen, Y.; Asemoloye, M.D.; Põlme, S.; Wang, X.; Xu, S.; Ramteke, P.W.; Pecoraro, L. Analysis of culturable airborne fungi in outdoor environments in Tianjin, China. BMC Microbiol. 2021, 21, 134. [Google Scholar] [CrossRef]
- Nageen, Y.; Wang, X.; Pecoraro, L. Seasonal variation of airborne fungal diversity and community structure in urban outdoor environments in Tianjin, China. Front. Microbiol. 2023, 13, 1043224. [Google Scholar] [CrossRef]
- Wu, D.; Zhang, Y.; Li, A.; Kong, Q.; Li, Y.; Geng, S.; Dong, X.; Liu, Y.; Chen, P. Indoor airborne fungal levels in selected comprehensive compartments of the urban utility tunnel in Nanjing, Southeast China. Sustain. Cities Soc. 2019, 51, 101723. [Google Scholar] [CrossRef]
- Fang, Z.; Ouyang, Z.; Hu, L.; Wang, X.; Zheng, H.; Lin, X. Culturable airborne fungi in outdoor environments in Beijing, China. Sci. Total Environ. 2005, 350, 47–58. [Google Scholar] [CrossRef]
- Wang, W.; Siu, K.W.M.; Wong, K.C.K. The pedestrian bridge as everyday place in high-density cities: An urban reference for necessity and sufficiency of placemaking. Urban Des. Int. 2016, 21, 236–253. [Google Scholar] [CrossRef]
- Duperrex, O.; Bunn, F.; Roberts, I. Safety education of pedestrians for injury prevention: A systematic review of randomised controlled trials. BMJ 2002, 324, 1129. [Google Scholar] [CrossRef]
- Räsänen, M.; Lajunen, T.; Alticafarbay, F.; Aydin, C. Pedestrian self-reports of factors influencing the use of pedestrian bridges. Accid. Anal. Prev. 2007, 39, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Castro, E.S.D.M.; Marcusso, R.M.N.; Barbosa, C.G.G.; Gonçalves, F.L.T.; Cardoso, M.R.A. Air pollution and its impact on the concentration of airborne fungi in the megacity of São Paulo, Brazil. Heliyon 2020, 6, e05065. [Google Scholar] [CrossRef] [PubMed]
- Grinn-Gofroń, A.; Strzelczak, A.; Wolski, T. The relationships between air pollutants, meteorological parameters and concentration of airborne fungal spores. Environ. Pollut. 2011, 159, 602–608. [Google Scholar] [CrossRef]
- Nriagu, J.O. Encyclopedia of Environmental Health; Elsevier: Amsterdam, The Netherlands, 2019; 4884p. [Google Scholar]
- Guo, Z.; Wang, Z.; Qian, L.; Zhao, Z.; Zhang, C.; Fu, Y.; Li, J.; Zhang, C.; Lu, B.; Qian, J. Biological and chemical compositions of atmospheric particulate matter during hazardous haze days in Beijing. Environ. Sci. Pollut. Res. Int. 2018, 25, 34540–34549. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Pecoraro, L.; Girlanda, M.; Kull, T.; Perini, C.; Perotto, S. Molecular identification of root fungal associates in Orchis pauciflora Tenore. Plant Biosyst. 2012, 146, 985–991. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Pecoraro, L.; Rasmussen, H.N.; Gomes, S.I.F.; Wang, X.; Merckx, V.S.F.T.; Cai, L.; Rasmussen, F.N. Fungal diversity driven by bark features affects phorophyte preference in epiphytic orchids from southern China. Sci. Rep. 2021, 11, 11287. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Pecoraro, L. Diversity and co-occurrence patterns of fungal and bacterial communities from alkaline sediments and water of Julong high-altitude Hot Springs at Tianchi volcano, Northeast China. Biology 2021, 10, 894. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 16 January 2023).
- McKight, P.E.; Najab, J. Kruskal-wallis test. In The Corsini Encyclopedia of Psychology; John Wiley & Sons: Hoboken, NJ, USA, 2010; p. 1. [Google Scholar]
- Wilcoxon, F. Individual comparisons by ranking methods. Biometrics 1945, 1, 80–83. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. _vegan: Community Ecology Package_.R Package Version 2.6-4. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 18 January 2023).
- Kolde, R. _Pheatmap: Pretty Heatmaps_.R Package Version 1.0.12. 2019. Available online: https://CRAN.R-project.org/package=pheatmap (accessed on 20 January 2023).
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 325–349. [Google Scholar] [CrossRef]
- Fang, Z.; Ouyang, Z.; Zheng, H.; Wang, X. Concentration and size distribution of culturable airborne microorganisms in outdoor environments in Beijing, China. Aerosol Sci. Technol. 2008, 42, 325–334. [Google Scholar] [CrossRef]
- Downs, S.H.; Mitakakis, T.Z.; Marks, G.B.; Car, N.G.; Belousova, E.G.; Leüppi, J.D.; Xuan, W.; Downie, S.R.; Tobias, A.; Peat, J.K. Clinical importance of Alternaria exposure in children. Am. J. Respir. Crit. Care Med. 2001, 164, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Didehdar, M.; Khoshbayan, A.; Vesal, S.; Darban-Sarokhalil, D.; Razavi, S.; Chegini, Z.; Shariati, A. An overview of possible pathogenesis mechanisms of Alternaria alternata in chronic rhinosinusitis and nasal polyposis. Microb. Pathog. 2021, 155, 104905. [Google Scholar] [CrossRef]
- Mokhtari Amirmajdi, M.; Mokhtari Amirmajdi, N.A.; Eftekharzadeh Mashhadi, I.; Jabari Azad, F.; Tavakol Afshari, J.; Shakeri, M.T. Alternaria in patients with allergic rhinitis. Iran J. Allergy Asthma Immunol. 2011, 10, 221–226. [Google Scholar]
- Yuan, C.; Wang, X.; Asemoloye, M.D.; Wang, Y.; Gargano, M.L.; Xue, H.-J.; Pecoraro, L. First record of Clonostachys rosea as entomopathogenic fungus of Coleoptera in China. Plant Biosyst. 2022, 155, 1240–1246. [Google Scholar] [CrossRef]
- Gravesen, S. Fungi as a cause of allergic disease. Allergy 1979, 34, 135–154. [Google Scholar] [CrossRef]
- Kasprzyk, I.; Kaszewski, B.M.; Weryszko-Chmielewska, E.; Nowak, M.; Sulborska, A.; Kaczmarek, J.; Szymanska, A.; Haratym, W.; Jedryczka, M. Warm and dry weather accelerates and elongates Cladosporium spore seasons in Poland. Aerobiologia 2016, 32, 109–126. [Google Scholar] [CrossRef]
- Maxwell, S. What to Know About Cladosporium Mold. Family Handyman. 2021. Available online: https://www.familyhandyman.com/article/cladosporium-mold/ (accessed on 11 January 2023).
- Gabriel, M.F.; Postigo, I.; Tomaz, C.T.; Martínez, J. Alternaria alternata allergens: Markers of exposure, phylogeny and risk of fungi-induced respiratory allergy. Environ. Int. 2016, 89–90, 71–80. [Google Scholar] [CrossRef]
- Bandoim, L. What Is Cladosporium and Is It Dangerous to Your Health? Healthline. 2020. Available online: https://www.healthline.com/health/cladosporium (accessed on 11 January 2023).
- Sellart-Altisent, M.; Torres-Rodríguez, J.M.; Gómez de Ana, S.; Alvarado-Ramírez, E. Nasal fungal microbiota in allergic and healthy subjects. Rev. Iberoam Micol. 2007, 24, 125–130. [Google Scholar] [CrossRef]
- Ma, X.; Gu, Y.; Liu, X.; Li, D.; Ling, S.; Hou, J.; Wang, C.; Cao, S.; Huang, X.; Wen, X.; et al. Phaeohyphomycotic dermatitis in a giant panda (Ailuropoda melanoleuca) caused by Cladosporium cladosporioides. Med. Mycol. Case Rep. 2013, 2, 119–121. [Google Scholar] [CrossRef] [PubMed]
- Niu, M.; Hu, W.; Cheng, B.; Wu, L.; Ren, L.; Deng, J.; Shen, F.; Fu, P. Influence of rainfall on fungal aerobiota in the urban atmosphere over Tianjin, China: A case study. Atmos. Environ. X 2021, 12, 100137. [Google Scholar] [CrossRef]
- Wessels, J.G.; De Vries, O.M.; Asgeirsdottir, S.A.; Schuren, F. Hydrophobin genes involved in formation of aerial hyphae and fruit bodies in Schizophyllum. Plant Cell 1991, 3, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Mu, Y.; Zhang, H.; Su, X.; Yang, C.; Shu, X.; Qing, D. First report on cutaneous infectious granuloma caused by Schizophyllum commune. BMC Infect. Dis. 2018, 18, 286. [Google Scholar] [CrossRef]
- Gargano, M.L.; Venturella, G.; Bosco, F.; Fabrini, M.G.; Gao, W.; Pecoraro, L. The Role of Medicinal Mushrooms in Brain Cancer Therapies: Review. Int. J. Med. Mushrooms 2021, 23, 13–20. [Google Scholar] [CrossRef]
- Unno, H.; Kamei, K.; Honda, A.; Nishimura, K.; Kuriyama, T. A murine model of pulmonary basidiomycosis by Schizophyllum commune. J. Infect. Chemother. 2005, 11, 136–140. [Google Scholar] [CrossRef]
- Chowdhary, A.; Randhawa, H.S.; Gaur, S.N.; Agarwal, K.; Kathuria, S.; Roy, P.; Klaassen, C.H.; Meis, J.F. Schizophyllum commune as an emerging fungal pathogen: A review and report of two cases. Mycoses 2013, 56, 1–10. [Google Scholar] [CrossRef]
- Ren, Q.; Wan, L.; Ju, Y.; Yuan-zhu, L. Animal experimental study of the pathogenicity of Schizophyllum commune. J. Clin. Dermatol. 2011, 40, 587–591. [Google Scholar]
- Kligman, A.M. A basidiomycete probably causing onychomycosis. J. Invest. Dermatol. 1950, 14, 67–70. [Google Scholar] [CrossRef]
- Vulinović, J.N.; Lolić, S.B.; Vujčić, S.B.; Matavulj, M.N. Schizophyllum commune—The dominant cause of trees decay in alleys and parks in the City of Novi Sad (Serbia). Biol. Serbica 2019, 40, 26–33. [Google Scholar]
- Larsen, L.S. A three-year-survey of microfungi in the air of Copenhagen 1977–79. Allergy 1981, 36, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Harrison, R.M. The effects of meteorological factors on atmospheric bioaerosol concentrations—A review. Sci. Total Environ. 2004, 326, 151–180. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, P.; Gupta-Bhattacharya, S.; Chowdhury, I.; Majumdar, M.R.; Chanda, S. Differences in concentrations of allergenic pollens and spores at different heights on an agricultural farm in West Bengal, India. Ann. Agric. Environ. Med. 2001, 8, 123–130. [Google Scholar] [PubMed]
- Lu, Y.; Wang, X.; Almeida, L.C.S.d.S.; Pecoraro, L. Environmental Factors Affecting Diversity, Structure, and Temporal Variation of Airborne Fungal Communities in a Research and Teaching Building of Tianjin University, China. J. Fungi 2022, 8, 431. [Google Scholar] [CrossRef]
- Moubasher, A.H.; Abdel-Fattah, H.M.; Swelim, M.A. Studies on air-borne fungi at Qena. I. Seasonal fluctuations. Z. Allg. Mikrobiol. 1981, 21, 247–253. [Google Scholar] [CrossRef]
- Boonen, J.; Malysheva, S.V.; Taevernier, L.; Di Mavungu, J.D.; De Saeger, S.; De Spiegeleer, B. Human skin penetration of selected model mycotoxins. Toxicology 2012, 301, 21–32. [Google Scholar] [CrossRef]
- Kemppainen, B.; Riley, R.; Pace, J. Skin absorption as a route of exposure for aflatoxin and trichothecenes. J. Toxicol. Toxin Rev. 1988, 7, 95–120. [Google Scholar] [CrossRef]
- Lewis, L.; Onsongo, M.; Njapau, H.; Schurz-Rogers, H.; Luber, G.; Kieszak, S.; Nyamongo, J.; Backer, L.; Dahiye, A.M.; Misore, A.; et al. Aflatoxin contamination of commercial maize products during an outbreak of acute aflatoxicosis in eastern and central Kenya. Environ. Health Perspect. 2005, 113, 1763–1767. [Google Scholar] [CrossRef]
- Viegas, S.; Viegas, C.; Oppliger, A. Occupational Exposure to Mycotoxins: Current Knowledge and Prospects. Ann. Work Expo Health 2018, 62, 923–941. [Google Scholar] [CrossRef]
- Yu, J.; Payne, G.A.; Nierman, W.C.; Machida, M.; Bennett, J.W.; Campbell, B.C.; Robens, J.F.; Bhatnagar, D.; Dean, R.A.; Cleveland, T.E. Aspergillus flavus genomics as a tool for studying the mechanism of aflatoxin formation. Food Addit. Contam. 2008, 25, 1152–1157. [Google Scholar]
- Githang’a, D.; Wangia, R.N.; Mureithi, M.W.; Wandiga, S.O.; Mutegi, C.; Ogutu, B.; Agweyu, A.; Wang, J.S.; Anzala, O. The effects of aflatoxin exposure on Hepatitis B-vaccine induced immunity in Kenyan children. Curr. Probl. Pediatr. Adolesc. Health Care 2019, 49, 117–130. [Google Scholar] [CrossRef] [PubMed]
- McCullough, A.K.; Lloyd, R.S. Mechanisms underlying aflatoxin-associated mutagenesis—Implications in carcinogenesis. DNA Repair 2019, 77, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Selim, M.I. Significance of aflatoxins in rural and global health: Concern for agricultural workers. North Carol. Med. J. 2010, 71, 438–441. [Google Scholar] [CrossRef]
- Alavanja, M.C.; Malker, H.; Hayes, R.B. Occupational cancer risk associated with the storage and bulk handling of agricultural foodstuff. J. Toxicol. Environ. Health 1987, 22, 247–254. [Google Scholar] [CrossRef]
- Bulpa, P.; Dive, A.; Sibille, Y. Invasive pulmonary aspergillosis in patients with chronic obstructive pulmonary disease. Eur. Respir. J. 2007, 30, 782–800. [Google Scholar] [CrossRef]
- Korzeniowska-Koseła, M.; Halweg, H.; Bestry, I.; Podsiadło, B.; Krakówka, P. Pulmonary aspergilloma caused by Aspergillus niger. Pneumonol. Pol. 1990, 58, 328–333. [Google Scholar] [PubMed]
- Person, A.K.; Chudgar, S.M.; Norton, B.L.; Tong, B.C.; Stout, J.E. Aspergillus niger: An unusual cause of invasive pulmonary aspergillosis. J. Med. Microbiol. 2010, 59, 834–838. [Google Scholar] [CrossRef]
- Araiza, J.; Canseco, P.; Bonifaz, A. Otomycosis: Clinical and mycological study of 97 cases. Rev. Laryngol. Otol. Rhinol. 2006, 127, 251–254. [Google Scholar]
- Loudon, K.W.; Coke, A.P.; Burnie, J.P.; Shaw, A.J.; Oppenheim, B.A.; Morris, C.Q. Kitchens as a source of Aspergillus niger infection. J. Hosp. Infect. 1996, 32, 191–198. [Google Scholar] [CrossRef]
- Ouf, S.A.; Basher, A.H.; Mohamed, A.A. Inhibitory effect of double atmospheric pressure argon cold plasma on spores and mycotoxin production of Aspergillus niger contaminating date palm fruits. J. Sci. Food Agric. 2015, 95, 3204–3210. [Google Scholar] [CrossRef]
- Jing, R.; Yang, W.H.; Xiao, M.; Li, Y.; Zou, G.L.; Wang, C.Y.; Li, X.W.; Xu, Y.C.; Hsueh, P.R. Species identification and antifungal susceptibility testing of Aspergillus strains isolated from patients with otomycosis in northern China. J. Microbiol. Immunol. Infect. 2021, 55, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Guiserix, J.; Ramdane, M.; Finielz, P.; Michault, A.; Rajaonarivelo, P. Trichoderma harzianum peritonitis in peritoneal dialysis. Nephron 1996, 74, 473–474. [Google Scholar] [CrossRef] [PubMed]
- Loeppky, C.B.; Sprouse, R.F.; Carlson, J.V.; Everett, E.D. Trichoderma viride peritonitis. South Med. J. 1983, 76, 798–799. [Google Scholar] [CrossRef]
- Ragnaud, J.; Marceau, C.; Roche-Bezian, M.; Wone, C. Infection peritoneale a Trichoderma koningii sur dialyse peritoneale continue ambulatoire. Méd. Mal. Infect. 1984, 14, 402–405. [Google Scholar] [CrossRef]
- Rota, S.; Marchesi, D.; Farina, C.; De Bièvre, C. Trichoderma pseudokoningii peritonitis in automated peritoneal dialysis patient successfully treated by early catheter removal. Perit. Dial Int. 2000, 20, 91–93. [Google Scholar] [CrossRef] [PubMed]
- Tanis, B.C.; Van der Pijl, H.; Van Ogtrop, M.L.; Kibbelaar, R.E.; Chang, P.C. Fatal fungal peritonitis by Trichoderma longibrachiatum complicating peritoneal dialysis. Nephrol. Dial Transpl. 1995, 10, 114–116. [Google Scholar]
- Xu, S.; Wang, X.; Nageen, Y.; Pecoraro, L. Analysis of gut-associated fungi from Chinese mitten crab Eriocheir sinensis. All Life 2021, 14, 610–621. [Google Scholar] [CrossRef]
- Alami, S.; Sekkal, I.; Aoufi, S.; Lyagoubi, M.; Benzekri, L.; Senouci, K. Subcutaneous phaeohyphomycosis caused by Aureobasidium Pullulans in an immunocompetent carpenter. Med. Mycol. Case Rep. 2022, 36, 1–4. [Google Scholar] [CrossRef]
- Abdel Hameed, A.A.; Khoder, M.I.; Yuosra, S.; Osman, A.M.; Ghanem, S. Diurnal distribution of airborne bacteria and fungi in the atmosphere of Helwan area, Egypt. Sci. Total Environ. 2009, 407, 6217–6222. [Google Scholar] [CrossRef]
- Hughes, K.M.; Price, D.; Torriero, A.A.J.; Symonds, M.R.E.; Suphioglu, C. Impact of Fungal Spores on Asthma Prevalence and Hospitalization. Int. J. Mol. Sci. 2022, 23, 4313. [Google Scholar] [CrossRef]
- Pashley, C.H.; Wardlaw, A.J. Allergic fungal airways disease (AFAD): An under-recognised asthma endotype. Mycopathologia 2021, 186, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Bundy, K.W.; Gent, J.F.; Beckett, W.; Bracken, M.B.; Belanger, K.; Triche, E.; Leaderer, B.P. Household airborne Penicillium associated with peak expiratory flow variability in asthmatic children. Ann. Allergy Asthma Immunol. 2009, 103, 26–30. [Google Scholar] [CrossRef]
- Knutsen, A.P.; Bush, R.K.; Demain, J.G.; Denning, D.W.; Dixit, A.; Fairs, A.; Greenberger, P.A.; Kariuki, B.; Kita, H.; Kurup, V.P.; et al. Fungi and allergic lower respiratory tract diseases. J. Allergy Clin. Immunol. 2012, 129, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Jacobsen, S.; Jørgensen, H.J.; Collinge, D.B.; Svensson, B.; Finnie, C. Fusarium graminearum and its interactions with cereal heads: Studies in the proteomics era. Front. Plant Sci. 2013, 4, 37. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Sites | Mean (CFU/m3) | SD | Min (CFU/m3) | Max (CFU/m3) | |
|---|---|---|---|---|---|
| BJD | C | 70.00 | 26.46 | 40 | 90 |
| U | 103.33 | 70.95 | 40 | 180 | |
| NG | C | 90.00 | 26.46 | 70 | 120 |
| U | 63.33 | 35.12 | 30 | 100 | |
| WJY | C | 63.33 | 55.08 | 10 | 120 |
| U | 103.33 | 37.86 | 60 | 130 | |
| WJR | C | 60.00 | 26.46 | 40 | 90 |
| U | 60.00 | 55.68 | 10 | 120 | |
| NMWA | C | 90.00 | 72.11 | 10 | 150 |
| U | 56.67 | 5.77 | 50 | 60 | |
| Sampling Locations | BJD | NG | WJY | WJR | NMWA | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| C | U | C | U | C | U | C | U | C | U | |
| First week | 0.79 | 1.04 | 1.15 | 1.1 | 0 | 1.56 | 1.61 | 0 | 0 | 1.33 |
| Second week | 1.38 | 1.52 | 1.91 | 1.56 | 2.14 | 1.93 | 1.04 | 2.37 | 2.21 | 1.56 |
| Third week | 1.43 | 1.35 | 1.79 | 1.97 | 1.33 | 1.31 | 1.43 | 1.61 | 1.12 | 1.79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Shaarani, A.A.Q.A.; Quach, Z.M.; Wang, X.; Muafa, M.H.M.; Nafis, M.M.H.; Pecoraro, L. Analysis of Airborne Fungal Communities on Pedestrian Bridges in Urban Environments. Microorganisms 2023, 11, 2097. https://doi.org/10.3390/microorganisms11082097
Al-Shaarani AAQA, Quach ZM, Wang X, Muafa MHM, Nafis MMH, Pecoraro L. Analysis of Airborne Fungal Communities on Pedestrian Bridges in Urban Environments. Microorganisms. 2023; 11(8):2097. https://doi.org/10.3390/microorganisms11082097
Chicago/Turabian StyleAl-Shaarani, Amran A. Q. A., Ziwei M. Quach, Xiao Wang, Mohammed H. M. Muafa, Md M. H. Nafis, and Lorenzo Pecoraro. 2023. "Analysis of Airborne Fungal Communities on Pedestrian Bridges in Urban Environments" Microorganisms 11, no. 8: 2097. https://doi.org/10.3390/microorganisms11082097
APA StyleAl-Shaarani, A. A. Q. A., Quach, Z. M., Wang, X., Muafa, M. H. M., Nafis, M. M. H., & Pecoraro, L. (2023). Analysis of Airborne Fungal Communities on Pedestrian Bridges in Urban Environments. Microorganisms, 11(8), 2097. https://doi.org/10.3390/microorganisms11082097