
Current Understanding of Potential Linkages between Biocide Tolerance and Antibiotic Cross-Resistance


Abstract
1. Introduction
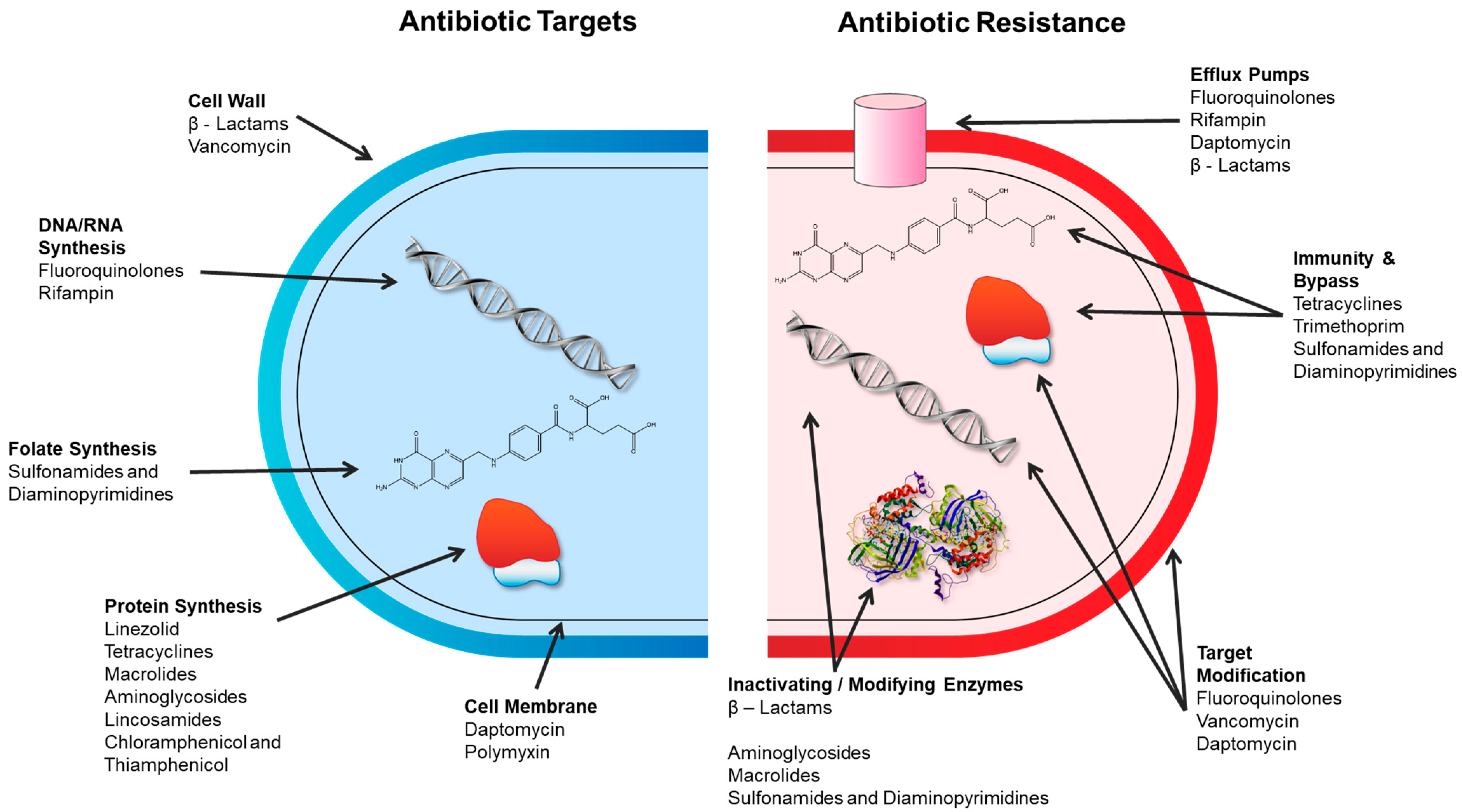
2. Antibiotics—Major Drug Classes, Chemistries, Modes of Action, and Resistance Mechanisms
- Alteration of the target thereby preventing the drug from binding;
- Enzymatic modification of the drug to degrade or modify it;
- Decrease in the accumulation of the antibiotic by the alteration of porins (reducing access) or by the overexpression of efflux transporters (increasing removal);
- Overproduction of the target to overwhelm the drug.
2.1. Antibiotics That Target Cell Wall Biosynthesis
2.2. Antibiotics That Target Protein Synthesis
2.3. Antibiotics That Affect Nucleic Acids
2.4. Antimetabolite Antibiotics
2.5. Antibiotics That Target the Membrane
2.6. Antibiotics Summary
3. Biocides—Major Classes, Chemistries, Modes of Action, and Resistance Mechanisms
3.1. Biocides That Inactivate through Ionic Interactions
3.1.1. Quaternary Ammonium Compounds (QACs)
3.1.2. Bisbiguanides
3.2. Biocides That Inactivate through the Disruption of Hydrogen Bonds
3.2.1. Phenolics
3.2.2. Alcohols
3.3. Biocides That Inactivate through Chemical Reactions
3.3.1. Metals
3.3.2. Chlorine-Releasing Agents
3.3.3. Fixatives (Aldehydes)
3.3.4. Peroxygens
3.3.5. Iodine
3.3.6. Bronopol
3.3.7. Ethylene Oxide
3.3.8. Isothiazolinone
3.4. Biocides Summary
4. Summary of Studies Investigating the Potential for Antibiotic Cross-Resistance
4.1. Quaternary Ammonium Compounds (QACs)
4.2. Bisbiguanides
4.3. Phenolics
4.4. Metals
4.5. Chlorine-Releasing Agents
4.6. Fixatives
4.7. Peroxygens
4.8. Alcohols
4.9. Iodine
5. Discussion
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- European Chemicals Agency (ECHA). Biocidal Products Regulation (BPR) Annex V Product Types. Available online: https://echa.europa.eu/regulations/biocidal-products-regulation/product-types (accessed on 19 August 2022).
- United States Environmental Protection Agency (EPA). What Are Antimicrobial Pesticides? Available online: https://www.epa.gov/pesticide-registration/what-are-antimicrobial-pesticides#types (accessed on 19 August 2022).
- Jones, I.A.; Joshi, L.T. Biocide Use in the Antimicrobial Era: A Review. Molecules 2021, 26, 2276. [Google Scholar] [CrossRef] [PubMed]
- Russell, A.D. Introduction of biocides into clinical practice and the impact on antibiotic-resistant bacteria. J. Appl. Microbiol. 2002, 92, 121S–135S. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, present and future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Center for Disease Control and Prevention (CDC). Antibiotic Resistance Threats in the United States; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2019.
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Center for Disease Control and Prevention (CDC). Glossary of Terms Related to Antibiotic Resistance. Available online: https://www.cdc.gov/narms/resources/glossary.html#:~:text=Multidrug%2Dresistant%20(MDR)%20isolate,to%20one%20or%20more%20antibiotics (accessed on 16 July 2023).
- Reddy, G.K.K.; Padmavathi, A.R.; Nancharaiah, Y.V. Fungal infections: Pathogenesis, antifungals and alternate treatment approaches. Curr. Res. Microb. Sci. 2022, 3, 100137. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, E.; Li, G. Approved Antiviral Drugs over the Past 50 Years. Clin. Microbiol. Rev. 2016, 29, 695–747. [Google Scholar] [CrossRef] [PubMed]
- Kausar, S.; Said Khan, F.; Ishaq Mujeeb Ur Rehman, M.; Akram, M.; Riaz, M.; Rasool, G.; Hamid Khan, A.; Saleem, I.; Shamim, S.; Malik, A. A review: Mechanism of action of antiviral drugs. Int. J. Immunopathol. Pharmacol. 2021, 35, 20587384211002621. [Google Scholar] [CrossRef]
- Maillard, J.Y. Resistance of Bacteria to Biocides. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Curiao, T.; Marchi, E.; Viti, C.; Oggioni, M.R.; Baquero, F.; Martinez, J.L.; Coque, T.M. Polymorphic variation in susceptibility and metabolism of triclosan-resistant mutants of Escherichia coli and Klebsiella pneumoniae clinical strains obtained after exposure to biocides and antibiotics. Antimicrob. Agents Chemother. 2015, 59, 3413–3423. [Google Scholar] [CrossRef]
- Zeng, W.; Xu, W.; Xu, Y.; Liao, W.; Zhao, Y.; Zheng, X.; Xu, C.; Zhou, T.; Cao, J. The prevalence and mechanism of triclosan resistance in Escherichia coli isolated from urine samples in Wenzhou, China. Antimicrob. Resist. Infect. Control 2020, 9, 161. [Google Scholar] [CrossRef]
- Martin, D.J.; Denyer, S.P.; McDonnell, G.; Maillard, J.Y. Resistance and cross-resistance to oxidising agents of bacterial isolates from endoscope washer disinfectors. J. Hosp. Infect. 2008, 69, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Sasatsu, M.; Shimizu, K.; Noguchi, N.; Kono, M. Triclosan-resistant Staphylococcus aureus. Lancet 1993, 341, 756. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.J.; Yu, Y.T.; Shapiro, M.A.; Olson, E.; Rock, C.O. Broad spectrum antimicrobial biocides target the FabI component of fatty acid synthesis. J. Biol. Chem. 1998, 273, 30316–30320. [Google Scholar] [CrossRef]
- Chapman, J.S. Disinfectant resistance mechanisms, cross-resistance, and co-resistance. Int. Biodeterior. Biodegrad. 2003, 51, 271–276. [Google Scholar] [CrossRef]
- Manzoor, S.E.; Lambert, P.A.; Griffiths, P.A.; Gill, M.J.; Fraise, A.P. Reduced glutaraldehyde susceptibility in Mycobacterium chelonae associated with altered cell wall polysaccharides. J. Antimicrob. Chemother. 1999, 43, 759–765. [Google Scholar] [CrossRef]
- Winder, C.L.; Al-Adham, I.S.; Abdel Malek, S.M.; Buultjens, T.E.; Horrocks, A.J.; Collier, P.J. Outer membrane protein shifts in biocide-resistant Pseudomonas aeruginosa PAO1. J. Appl. Microbiol. 2000, 89, 289–295. [Google Scholar] [CrossRef] [PubMed]
- O’Rourke, E.; Runyan, D.; O’Leary, J.; Stern, J. Contaminated iodophor in the operating room. Am. J. Infect. Control 2003, 31, 255–256. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, J.T.; Monach, P.; Chou, J.H.; Josephy, P.D.; Demple, B. Positive control of a global antioxidant defense regulon activated by superoxide-generating agents in Escherichia coli. Proc. Natl. Acad. Sci. USA 1990, 87, 6181–6185. [Google Scholar] [CrossRef]
- Dukan, S.; Touati, D. Hypochlorous acid stress in Escherichia coli: Resistance, DNA damage, and comparison with hydrogen peroxide stress. J. Bacteriol. 1996, 178, 6145–6150. [Google Scholar] [CrossRef]
- Walsh, S.E.; Maillard, J.Y.; Russell, A.D.; Catrenich, C.E.; Charbonneau, D.L.; Bartolo, R.G. Development of bacterial resistance to several biocides and effects on antibiotic susceptibility. J. Hosp. Infect. 2003, 55, 98–107. [Google Scholar] [CrossRef]
- Tattawasart, U.; Maillard, J.Y.; Furr, J.R.; Russell, A.D. Development of resistance to chlorhexidine diacetate and cetylpyridinium chloride in Pseudomonas stutzeri and changes in antibiotic susceptibility. J. Hosp. Infect. 1999, 42, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Araújo, P.A.; Lemos, M.; Mergulhão, F.J.; Melo, L.F.; Simões, M. Antimicrobial resistance to disinfectants in biofilms. In Science Against Microbial Pathogens: Communicating Current Research and Technological Advances; Mendez-Vilas, A., Ed.; Microbiology Series; Formatex Research Center: Badajoz, Spain, 2011; Volume 1, pp. 826–834. [Google Scholar]
- Candido, E.S.; de Barros, E.; Cardoso, M.H.; Franco, O.L. Bacterial cross-resistance to anti-infective compounds. Is it a real problem? Curr. Opin. Pharmacol. 2019, 48, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, P.; Allison, D.G.; McBain, A.J. Biofilms in vitro and in vivo: Do singular mechanisms imply cross-resistance? Symp. Ser. Soc. Appl. Microbiol. 2002, 92, 98S–110S. [Google Scholar] [CrossRef]
- van Bambeke, F.; Mingeot-Leclercq, M.-P.; Glupczynski, Y.; Tulkens, P.M. Mechanisms of Action. In Infectious Diseases, 4th ed.; Cohen, J., Powderly, W.G., Opal, S.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1162–1180.e1161. [Google Scholar]
- Purssell, E. Antimicrobials. In Understanding Pharmacology in Nursing Practice; Hood, P., Khan, E., Eds.; Springer: Cham, Switzerland, 2020; pp. 147–165. [Google Scholar]
- Kapoor, G.; Saigal, S.; Elongavan, A. Action and resistance mechanisms of antibiotics: A guide for clinicians. J. Anaesthesiol. Clin. Pharmacol. 2017, 33, 300–305. [Google Scholar] [CrossRef]
- Wright, G.D. Q&A: Antibiotic resistance: Where does it come from and what can we do about it? BMC Biol. 2010, 8, 123. [Google Scholar] [CrossRef]
- Alfei, S.; Schito, A.M. Beta-Lactam Antibiotics and beta-Lactamase Enzymes Inhibitors, Part 2: Our Limited Resources. Pharmaceuticals 2022, 15, 476. [Google Scholar] [CrossRef] [PubMed]
- Krause, K.M.; Serio, A.W.; Kane, T.R.; Connolly, L.E. Aminoglycosides: An Overview. Cold Spring Harb. Perspect. Med. 2016, 6, a027029. [Google Scholar] [CrossRef]
- Wilson, D.N. On the specificity of antibiotics targeting the large ribosomal subunit. Ann. N. Y. Acad. Sci. 2011, 1241, 1–16. [Google Scholar] [CrossRef]
- Bozdogan, B.; Appelbaum, P.C. Oxazolidinones: Activity, mode of action, and mechanism of resistance. Int. J. Antimicrob. Agents 2004, 23, 113–119. [Google Scholar] [CrossRef]
- Straus, S.K.; Hancock, R.E. Mode of action of the new antibiotic for Gram-positive pathogens daptomycin: Comparison with cationic antimicrobial peptides and lipopeptides. Biochim. Biophys. Acta 2006, 1758, 1215–1223. [Google Scholar] [CrossRef]
- Velkov, T.; Roberts, K.D.; Nation, R.L.; Thompson, P.E.; Li, J. Pharmacology of polymyxins: New insights into an ‘old’ class of antibiotics. Future Microbiol. 2013, 8, 711–724. [Google Scholar] [CrossRef] [PubMed]
- Gaibani, P.; Giani, T.; Bovo, F.; Lombardo, D.; Amadesi, S.; Lazzarotto, T.; Coppi, M.; Rossolini, G.M.; Ambretti, S. Resistance to Ceftazidime/Avibactam, Meropenem/Vaborbactam and Imipenem/Relebactam in Gram-Negative MDR Bacilli: Molecular Mechanisms and Susceptibility Testing. Antibiotics 2022, 11, 628. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Wu, C.; Gao, H.; Xu, C.; Dai, M.; Huang, L.; Hao, H.; Wang, X.; Cheng, G. Bacterial Multidrug Efflux Pumps at the Frontline of Antimicrobial Resistance: An Overview. Antibiotics 2022, 11, 520. [Google Scholar] [CrossRef] [PubMed]
- Mingeot-Leclercq, M.P.; Glupczynski, Y.; Tulkens, P.M. Aminoglycosides: Activity and resistance. Antimicrob. Agents Chemother. 1999, 43, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Kehrenberg, C.; Doublet, B.; Cloeckaert, A. Molecular basis of bacterial resistance to chloramphenicol and florfenicol. FEMS Microbiol. Rev. 2004, 28, 519–542. [Google Scholar] [CrossRef]
- Van Bambeke, F.; Michot, J.M.; Van Eldere, J.; Tulkens, P.M. Quinolones in 2005: An update. Clin. Microbiol. Infect. 2005, 11, 256–280. [Google Scholar] [CrossRef]
- Azargun, R.; Gholizadeh, P.; Sadeghi, V.; Hosainzadegan, H.; Tarhriz, V.; Memar, M.Y.; Pormohammad, A.; Eyvazi, S. Molecular mechanisms associated with quinolone resistance in Enterobacteriaceae: Review and update. Trans. R. Soc. Trop. Med. Hyg. 2020, 114, 770–781. [Google Scholar] [CrossRef]
- Robicsek, A.; Jacoby, G.A.; Hooper, D.C. The worldwide emergence of plasmid-mediated quinolone resistance. Lancet Infect. Dis. 2006, 6, 629–640. [Google Scholar] [CrossRef]
- Xu, G.; Liu, H.; Jia, X.; Wang, X.; Xu, P. Mechanisms and detection methods of Mycobacterium tuberculosis rifampicin resistance: The phenomenon of drug resistance is complex. Tuberculosis 2021, 128, 102083. [Google Scholar] [CrossRef]
- Peng, Z.; Jin, D.; Kim, H.B.; Stratton, C.W.; Wu, B.; Tang, Y.W.; Sun, X. Update on Antimicrobial Resistance in Clostridium difficile: Resistance Mechanisms and Antimicrobial Susceptibility Testing. J. Clin. Microbiol. 2017, 55, 1998–2008. [Google Scholar] [CrossRef]
- Leeds, J.A.; Sachdeva, M.; Mullin, S.; Barnes, S.W.; Ruzin, A. In vitro selection, via serial passage, of Clostridium difficile mutants with reduced susceptibility to fidaxomicin or vancomycin. J. Antimicrob. Chemother. 2014, 69, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Kardan-Yamchi, J.; Kazemian, H.; Haeili, M.; Harati, A.A.; Amini, S.; Feizabadi, M.M. Expression analysis of 10 efflux pump genes in multidrug-resistant and extensively drug-resistant Mycobacterium tuberculosis clinical isolates. J. Glob. Antimicrob. Resist. 2019, 17, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Estrada, A.; Wright, D.L.; Anderson, A.C. Antibacterial Antifolates: From Development through Resistance to the Next Generation. Cold Spring Harb. Perspect. Med. 2016, 6, a028324. [Google Scholar] [CrossRef]
- Center for Disease Control and Prevention (CDC). Chemical Disinfectants—Guideline for Disinfection and Sterilization in Healthcare Facilities. Available online: https://www.cdc.gov/infectioncontrol/guidelines/disinfection/disinfection-methods/chemical.html (accessed on 18 July 2023).
- Ortega Morente, E.; Fernandez-Fuentes, M.A.; Grande Burgos, M.J.; Abriouel, H.; Perez Pulido, R.; Galvez, A. Biocide tolerance in bacteria. Int. J. Food Microbiol. 2013, 162, 13–25. [Google Scholar] [CrossRef]
- McDonnell, G.; Russell, A.D. Antiseptics and disinfectants: Activity, action, and resistance. Clin. Microbiol. Rev. 1999, 12, 147–179. [Google Scholar] [CrossRef] [PubMed]
- Maillard, J.Y. Bacterial target sites for biocide action. J. Appl. Microbiol. 2002, 92, 16S–27S. [Google Scholar] [CrossRef]
- Williams, T.M. The Mechanism of Action of Isothiazolone Biocides. In Proceedings of the NACE Conference CORROSION/2006, San Diego, CA, USA, 12–16 March 2006. [Google Scholar]
- Capriotti, K.; Capriotti, J.A. Topical iodophor preparations: Chemistry, microbiology, and clinical utility. Dermatol. Online J. 2012, 18, 1. [Google Scholar] [CrossRef]
- Saleh, S.; Haddadin, R.N.; Baillie, S.; Collier, P.J. Triclosan—An update. Lett. Appl. Microbiol. 2011, 52, 87–95. [Google Scholar] [CrossRef]
- Poole, K. Mechanisms of bacterial biocide and antibiotic resistance. J. Appl. Microbiol. 2002, 92, 55S–64S. [Google Scholar] [CrossRef]
- SCENIHR (Scientific Committee on Emerging and Newly Identified Health Risks). Assessment of the Antibiotic Resistance Effects of Biocides; European Commission, 2009.
- Cloete, T.E. Resistance mechanisms of bacteria to antimicrobial compounds. Int. Biodeterior. Biodegrad. 2003, 51, 277–282. [Google Scholar] [CrossRef]
- Vereshchagin, A.N.; Frolov, N.A.; Egorova, K.S.; Seitkalieva, M.M.; Ananikov, V.P. Quaternary Ammonium Compounds (QACs) and Ionic Liquids (ILs) as Biocides: From Simple Antiseptics to Tunable Antimicrobials. Int. J. Mol. Sci. 2021, 22, 6793. [Google Scholar] [CrossRef]
- Denyer, S.P.; Stewart, G.S.A.B. Mechanisms of action of disinfectants. Int. Biodeterior. Biodegrad. 1998, 41, 261–268. [Google Scholar] [CrossRef]
- Bouarab-Chibane, L.; Forquet, V.; Lanteri, P.; Clement, Y.; Leonard-Akkari, L.; Oulahal, N.; Degraeve, P.; Bordes, C. Antibacterial Properties of Polyphenols: Characterization and QSAR (Quantitative Structure-Activity Relationship) Models. Front. Microbiol. 2019, 10, 829. [Google Scholar] [CrossRef] [PubMed]
- The Norwegian Scientific Committee for Food and Environment (VKM). Antimicrobial Resistance Due to the Use of Biocides and Heavy Metals: A Literature Review; The Norwegian Scientific Committee for Food and Environment: Oslo, Norway, 2016. [Google Scholar]
- Lepelletier, D.; Maillard, J.Y.; Pozzetto, B.; Simon, A. Povidone Iodine: Properties, Mechanisms of Action, and Role in Infection Control and Staphylococcus aureus Decolonization. Antimicrob. Agents Chemother. 2020, 64, 10–1128. [Google Scholar] [CrossRef]
- Shepherd, J.A.; Waigh, R.D.; Gilbert, P. Antibacterial action of 2-bromo-2-nitropropane-1,3-diol (bronopol). Antimicrob. Agents Chemother. 1988, 32, 1693–1698. [Google Scholar] [CrossRef] [PubMed]
- Shintani, H. Ethylene Oxide Gas Sterilization of Medical Devices. Biocontrol Sci. 2017, 22, 1–16. [Google Scholar] [CrossRef]
- Gilbert, P.; Moore, L.E. Cationic antiseptics: Diversity of action under a common epithet. J. Appl. Microbiol. 2005, 99, 703–715. [Google Scholar] [CrossRef]
- Chuanchuen, R.; Beinlich, K.; Hoang, T.T.; Becher, A.; Karkhoff-Schweizer, R.R.; Schweizer, H.P. Cross-resistance between triclosan and antibiotics in Pseudomonas aeruginosa is mediated by multidrug efflux pumps: Exposure of a susceptible mutant strain to triclosan selects nfxB mutants overexpressing MexCD-OprJ. Antimicrob. Agents Chemother. 2001, 45, 428–432. [Google Scholar] [CrossRef]
- Davin-Regli, A.; Pages, J.M. Cross-resistance between biocides and antimicrobials: An emerging question. Rev. Sci. Tech. 2012, 31, 89–104. [Google Scholar]
- Hughes, L.; Roberts, W.; Johnson, D. The impact of DNA adenine methyltransferase knockout on the development of triclosan resistance and antibiotic cross-resistance in Escherichia coli. Access Microbiol. 2021, 3, acmi000178. [Google Scholar] [CrossRef]
- Sonbol, F.I.; El-Banna, T.E.; Abd El-Aziz, A.A.; El-Ekhnawy, E. Impact of triclosan adaptation on membrane properties, efflux and antimicrobial resistance of Escherichia coli clinical isolates. J. Appl. Microbiol. 2019, 126, 730–739. [Google Scholar] [CrossRef] [PubMed]
- Fraise, A.P. Susceptibility of antibiotic-resistant cocci to biocides. J. Appl. Microbiol. 2002, 92, 158S–162S. [Google Scholar] [CrossRef]
- Henly, E.L.; Dowling, J.A.R.; Maingay, J.B.; Lacey, M.M.; Smith, T.J.; Forbes, S. Biocide Exposure Induces Changes in Susceptibility, Pathogenicity, and Biofilm Formation in Uropathogenic Escherichia coli. Antimicrob. Agents Chemother. 2019, 63, e01892-18. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Z.; Nikaido, H.; Williams, K.E. Silver-resistant mutants of Escherichia coli display active efflux of Ag+ and are deficient in porins. J. Bacteriol. 1997, 179, 6127–6132. [Google Scholar] [CrossRef]
- Percival, S.L.; Salisbury, A.M.; Chen, R. Silver, biofilms and wounds: Resistance revisited. Crit. Rev. Microbiol. 2019, 45, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Miloud, S.B.; Dziri, O.; Ferjani, S.; Ali, M.M.; Mysara, M.; Boutiba, I.; Houdt, R.V.; Chouchani, C. First Description of Various Bacteria Resistant to Heavy Metals and Antibiotics Isolated from Polluted Sites in Tunisia. Pol. J. Microbiol. 2021, 70, 161–174. [Google Scholar] [CrossRef]
- Timkova, I.; Lachka, M.; Kiskova, J.; Malinicova, L.; Nosalova, L.; Pristas, P.; Sedlakova-Kadukova, J. High frequency of antibiotic tolerance in deep subsurface heterotrophic cultivable bacteria from the Rozalia Gold Mine, Slovakia. Environ. Sci. Pollut. Res. Int. 2020, 27, 44036–44044. [Google Scholar] [CrossRef]
- Chandrangsu, P.; Rensing, C.; Helmann, J.D. Metal homeostasis and resistance in bacteria. Nat. Rev. Microbiol. 2017, 15, 338–350. [Google Scholar] [CrossRef]
- Russell, A.D. Glutaraldehyde: Current status and uses. Infect. Control Hosp. Epidemiol. 1994, 15, 724–733. [Google Scholar] [CrossRef]
- Dorsey, C.W.; Actis, L.A. Analysis of pVU3695, a plasmid encoding glutathione-dependent formaldehyde dehydrogenase activity and formaldehyde resistance in the Escherichia coli VU3695 clinical strain. Plasmid 2004, 51, 116–126. [Google Scholar] [CrossRef]
- Svetlikova, Z.; Skovierova, H.; Niederweis, M.; Gaillard, J.L.; McDonnell, G.; Jackson, M. Role of porins in the susceptibility of Mycobacterium smegmatis and Mycobacterium chelonae to aldehyde-based disinfectants and drugs. Antimicrob. Agents Chemother. 2009, 53, 4015–4018. [Google Scholar] [CrossRef] [PubMed]
- European Chemicals Agency (ECHA). Investigation Report Formaldehyde and Formaldehyde Releasers. Available online: https://echa.europa.eu/documents/10162/13641/annex_xv_report_formaldehyde_en.pdf/58be2f0a-7ca7-264d-a594-da5051a1c74b (accessed on 18 July 2023).
- Lachapelle, J.-M.; Castel, O.; Fueyo Casado, A.; Leroy, B.; Micali, G.; Tennstedt, D.; Lambert, J. Antiseptics in the era of bacterial resistance: A focus on povidone iodine. Clin. Pract. 2013, 10, 579–592. [Google Scholar] [CrossRef]
- Sutton, S.; Jimenez, L. A Review of Reported Recalls Involving Microbiological Control 2004-2011 with Emphasis on FDA Considerations of “Objectionable Organisms”. Am. Pharm. Rev. 2012, 15, 42–57. [Google Scholar]
- MacLehose, H.G.; Gilbert, P.; Allison, D.G. Biofilms, homoserine lactones and biocide susceptibility. J. Antimicrob. Chemother. 2004, 53, 180–184. [Google Scholar] [CrossRef]
- Escalada, M.G.; Russell, A.D.; Maillard, J.Y.; Ochs, D. Triclosan-bacteria interactions: Single or multiple target sites? Lett. Appl. Microbiol. 2005, 41, 476–481. [Google Scholar] [CrossRef]
- Cerf, O.; Carpentier, B.; Sanders, P. Tests for determining in-use concentrations of antibiotics and disinfectants are based on entirely different concepts: “resistance” has different meanings. Int. J. Food Microbiol. 2010, 136, 247–254. [Google Scholar] [CrossRef] [PubMed]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Clinical Breakpoints and Dosing of Antibiotics. Available online: https://www.eucast.org/clinical_breakpoints/ (accessed on 30 August 2022).
- Clinical and Laboratory Standards Institute (CLSI). Rationale for Using CLSI Breakpoints. Available online: https://clsi.org/meetings/rationale-for-using-clsi-breakpoints/ (accessed on 30 August 2022).
- ASTM E1054-21e1; Standard Practices for Evaluation of Inactivators of Antimicrobial Agents. ASTM: West Conshohocken, PA, USA, 2021.
- EN 14885:2018; Chemical Disinfectants and Antiseptics—Application of European Standards for Chemical Disinfectants and Antiseptics. CEN: Brussels, Belgium, 2018.
- Romanowski, E.G.; Yates, K.A.; Shanks, R.M.Q.; Kowalski, R.P. Benzalkonium Chloride Demonstrates Concentration-Dependent Antiviral Activity Against Adenovirus In Vitro. J. Ocul. Pharmacol. Ther. 2019, 35, 311–314. [Google Scholar] [CrossRef]
- Knapp, L.; Amezquita, A.; McClure, P.; Stewart, S.; Maillard, J.Y. Development of a protocol for predicting bacterial resistance to microbicides. Appl. Environ. Microbiol. 2015, 81, 2652–2659. [Google Scholar] [CrossRef][Green Version]
- Chen, B.; Han, J.; Dai, H.; Jia, P. Biocide-tolerance and antibiotic-resistance in community environments and risk of direct transfers to humans: Unintended consequences of community-wide surface disinfecting during COVID-19? Environ. Pollut. 2021, 283, 117074. [Google Scholar] [CrossRef]
- Gadea, R.; Fernandez Fuentes, M.A.; Perez Pulido, R.; Galvez, A.; Ortega, E. Effects of exposure to quaternary-ammonium-based biocides on antimicrobial susceptibility and tolerance to physical stresses in bacteria from organic foods. Food Microbiol. 2017, 63, 58–71. [Google Scholar] [CrossRef]
- Kampf, G. Biocidal Agents Used for Disinfection Can Enhance Antibiotic Resistance in Gram-Negative Species. Antibiotics 2018, 7, 110. [Google Scholar] [CrossRef] [PubMed]
- Kampf, G. Antibiotic ResistanceCan Be Enhanced in Gram-Positive Species by Some Biocidal Agents Used for Disinfection. Antibiotics 2019, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Braoudaki, M.; Hilton, A.C. Adaptive resistance to biocides in Salmonella enterica and Escherichia coli O157 and cross-resistance to antimicrobial agents. J. Clin. Microbiol. 2004, 42, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Forbes, S.; Knight, C.G.; Cowley, N.L.; Amezquita, A.; McClure, P.; Humphreys, G.; McBain, A.J. Variable Effects of Exposure to Formulated Microbicides on Antibiotic Susceptibility in Firmicutes and Proteobacteria. Appl. Environ. Microbiol. 2016, 82, 3591–3598. [Google Scholar] [CrossRef]
- Han, Y.; Zhou, Z.C.; Zhu, L.; Wei, Y.Y.; Feng, W.Q.; Xu, L.; Liu, Y.; Lin, Z.J.; Shuai, X.Y.; Zhang, Z.J.; et al. The impact and mechanism of quaternary ammonium compounds on the transmission of antibiotic resistance genes. Environ. Sci. Pollut. Res. Int. 2019, 26, 28352–28360. [Google Scholar] [CrossRef]
- Karatzas, K.A.; Webber, M.A.; Jorgensen, F.; Woodward, M.J.; Piddock, L.J.; Humphrey, T.J. Prolonged treatment of Salmonella enterica serovar Typhimurium with commercial disinfectants selects for multiple antibiotic resistance, increased efflux and reduced invasiveness. J. Antimicrob. Chemother. 2007, 60, 947–955. [Google Scholar] [CrossRef]
- Langsrud, S.; Sundheim, G.; Holck, A.L. Cross-resistance to antibiotics of Escherichia coli adapted to benzalkonium chloride or exposed to stress-inducers. J. Appl. Microbiol. 2004, 96, 201–208. [Google Scholar] [CrossRef]
- Merchel Piovesan Pereira, B.; Wang, X.; Tagkopoulos, I. Biocide-Induced Emergence of Antibiotic Resistance in Escherichia coli. Front. Microbiol. 2021, 12, 640923. [Google Scholar] [CrossRef]
- Nhung, N.T.; Thuy, C.T.; Trung, N.V.; Campbell, J.; Baker, S.; Thwaites, G.; Hoa, N.T.; Carrique-Mas, J. Induction of Antimicrobial Resistance in Escherichia coli and Non-Typhoidal Salmonella Strains after Adaptation to Disinfectant Commonly Used on Farms in Vietnam. Antibiotics 2015, 4, 480–494. [Google Scholar] [CrossRef]
- Yu, T.; Jiang, X.; Zhang, Y.; Ji, S.; Gao, W.; Shi, L. Effect of Benzalkonium Chloride Adaptation on Sensitivity to Antimicrobial Agents and Tolerance to Environmental Stresses in Listeria monocytogenes. Front. Microbiol. 2018, 9, 2906. [Google Scholar] [CrossRef]
- Kim, M.; Weigand, M.R.; Oh, S.; Hatt, J.K.; Krishnan, R.; Tezel, U.; Pavlostathis, S.G.; Konstantinidis, K.T. Widely Used Benzalkonium Chloride Disinfectants Can Promote Antibiotic Resistance. Appl. Environ. Microbiol. 2018, 84, e01201-18. [Google Scholar] [CrossRef] [PubMed]
- Kampf, G. Adaptive microbial response to low-level benzalkonium chloride exposure. J. Hosp. Infect. 2018, 100, e1–e22. [Google Scholar] [CrossRef] [PubMed]
- Short, F.L.; Lee, V.; Mamun, R.; Malmberg, R.; Li, L.; Espinosa, M.I.; Venkatesan, K.; Paulsen, I.T. Benzalkonium chloride antagonises aminoglycoside antibiotics and promotes evolution of resistance. EBioMedicine 2021, 73, 103653. [Google Scholar] [CrossRef]
- Mavri, A.; Smole Mozina, S. Involvement of efflux mechanisms in biocide resistance of Campylobacter jejuni and Campylobacter coli. J. Med. Microbiol. 2012, 61, 800–808. [Google Scholar] [CrossRef]
- Roedel, A.; Vincze, S.; Projahn, M.; Roesler, U.; Robe, C.; Hammerl, J.A.; Noll, M.; Al Dahouk, S.; Dieckmann, R. Genetic but No Phenotypic Associations between Biocide Tolerance and Antibiotic Resistance in Escherichia coli from German Broiler Fattening Farms. Microorganisms 2021, 9, 651. [Google Scholar] [CrossRef]
- Roedel, A.; Dieckmann, R.; Brendebach, H.; Hammerl, J.A.; Kleta, S.; Noll, M.; Al Dahouk, S.; Vincze, S. Biocide-Tolerant Listeria monocytogenes Isolates from German Food Production Plants Do Not Show Cross-Resistance to Clinically Relevant Antibiotics. Appl. Environ. Microbiol. 2019, 85, e01253-19. [Google Scholar] [CrossRef] [PubMed]
- Soumet, C.; Ragimbeau, C.; Maris, P. Screening of benzalkonium chloride resistance in Listeria monocytogenes strains isolated during cold smoked fish production. Lett. Appl. Microbiol. 2005, 41, 291–296. [Google Scholar] [CrossRef]
- Morrissey, I.; Oggioni, M.R.; Knight, D.; Curiao, T.; Coque, T.; Kalkanci, A.; Martinez, J.L.; Consortium, B. Evaluation of epidemiological cut-off values indicates that biocide resistant subpopulations are uncommon in natural isolates of clinically-relevant microorganisms. PLoS ONE 2014, 9, e86669. [Google Scholar] [CrossRef]
- Oggioni, M.R.; Coelho, J.R.; Furi, L.; Knight, D.R.; Viti, C.; Orefici, G.; Martinez, J.L.; Freitas, A.T.; Coque, T.M.; Morrissey, I.; et al. Significant Differences Characterise the Correlation Coefficients between Biocide and Antibiotic Susceptibility Profiles in Staphylococcus aureus. Curr. Pharm. Des. 2015, 21, 2054–2057. [Google Scholar] [CrossRef]
- Bischofberger, A.M.; Baumgartner, M.; Pfrunder-Cardozo, K.R.; Allen, R.C.; Hall, A.R. Associations between sensitivity to antibiotics, disinfectants and heavy metals in natural, clinical and laboratory isolates of Escherichia coli. Environ. Microbiol. 2020, 22, 2664–2679. [Google Scholar] [CrossRef]
- Amsalu, A.; Sapula, S.A.; De Barros Lopes, M.; Hart, B.J.; Nguyen, A.H.; Drigo, B.; Turnidge, J.; Leong, L.E.; Venter, H. Efflux Pump-Driven Antibiotic and Biocide Cross-Resistance in Pseudomonas aeruginosa Isolated from Different Ecological Niches: A Case Study in the Development of Multidrug Resistance in Environmental Hotspots. Microorganisms 2020, 8, 1647. [Google Scholar] [CrossRef] [PubMed]
- Romero, J.L.; Grande Burgos, M.J.; Perez-Pulido, R.; Galvez, A.; Lucas, R. Resistance to Antibiotics, Biocides, Preservatives and Metals in Bacteria Isolated from Seafoods: Co-Selection of Strains Resistant or Tolerant to Different Classes of Compounds. Front. Microbiol. 2017, 8, 1650. [Google Scholar] [CrossRef] [PubMed]
- Condell, O.; Iversen, C.; Cooney, S.; Power, K.A.; Walsh, C.; Burgess, C.; Fanning, S. Efficacy of biocides used in the modern food industry to control Salmonella enterica, and links between biocide tolerance and resistance to clinically relevant antimicrobial compounds. Appl. Environ. Microbiol. 2012, 78, 3087–3097. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, P.; Hans, A.; Ruikar, K.; Guan, Z.; Palmer, K.L. Reduced Chlorhexidine and Daptomycin Susceptibility in Vancomycin-Resistant Enterococcus faecium after Serial Chlorhexidine Exposure. Antimicrob. Agents Chemother. 2018, 62, 10–1128. [Google Scholar] [CrossRef]
- Wu, D.; Lu, R.; Chen, Y.; Qiu, J.; Deng, C.; Tan, Q. Study of cross-resistance mediated by antibiotics, chlorhexidine and Rhizoma coptidis in Staphylococcus aureus. J. Glob. Antimicrob. Resist. 2016, 7, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Coelho, J.R.; Carrico, J.A.; Knight, D.; Martinez, J.L.; Morrissey, I.; Oggioni, M.R.; Freitas, A.T. The use of machine learning methodologies to analyse antibiotic and biocide susceptibility in Staphylococcus aureus. PLoS ONE 2013, 8, e55582. [Google Scholar] [CrossRef] [PubMed]
- Department of Health and Human Services—Food and Drug Administration. Safety and Effectiveness of Consumer Antiseptics; Topical Antimicrobial Drug Products for Over-the-Counter Human Use; Department of Health and Human Services—Food and Drug Administration: Silver Spring, MD, USA, 2016.
- Eaves, D.J.; Ricci, V.; Piddock, L.J. Expression of acrB, acrF, acrD, marA, and soxS in Salmonella enterica serovar Typhimurium: Role in multiple antibiotic resistance. Antimicrob. Agents Chemother. 2004, 48, 1145–1150. [Google Scholar] [CrossRef]
- Aiello, A.E.; Larson, E.L.; Levy, S.B. Consumer antibacterial soaps: Effective or just risky? Clin. Infect. Dis. 2007, 45 (Suppl. S2), S137–S147. [Google Scholar] [CrossRef]
- Pycke, B.F.; Crabbe, A.; Verstraete, W.; Leys, N. Characterization of triclosan-resistant mutants reveals multiple antimicrobial resistance mechanisms in Rhodospirillum rubrum S1H. Appl. Environ. Microbiol. 2010, 76, 3116–3123. [Google Scholar] [CrossRef]
- Pi, B.; Yu, D.; Hua, X.; Ruan, Z.; Yu, Y. Genomic and transcriptome analysis of triclosan response of a multidrug-resistant Acinetobacter baumannii strain, MDR-ZJ06. Arch. Microbiol. 2017, 199, 223–230. [Google Scholar] [CrossRef]
- Rojo-Bezares, B.; Azcona-Gutierrez, J.M.; Martin, C.; Jareno, M.S.; Torres, C.; Saenz, Y. Streptococcus agalactiae from pregnant women: Antibiotic and heavy-metal resistance mechanisms and molecular typing. Epidemiol. Infect. 2016, 144, 3205–3214. [Google Scholar] [CrossRef]
- Lin, H.; Ye, C.; Chen, S.; Zhang, S.; Yu, X. Viable but non-culturable E. coli induced by low level chlorination have higher persistence to antibiotics than their culturable counterparts. Environ. Pollut. 2017, 230, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Romeu, M.J.; Rodrigues, D.; Azeredo, J. Effect of sub-lethal chemical disinfection on the biofilm forming ability, resistance to antibiotics and expression of virulence genes of Salmonella enteritidis biofilm-surviving cells. Biofouling 2020, 36, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Wesgate, R.; Grasha, P.; Maillard, J.Y. Use of a predictive protocol to measure the antimicrobial resistance risks associated with biocidal product usage. Am. J. Infect. Control 2016, 44, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Shan, K.; Li, J.; Yao, W.; Li, J.; He, X.; Li, F.; Wen, W.; Liu, Q.; Guo, W. Evaluation of Resistance by Clinically Pathogenic Bacteria to Antimicrobials and Common Disinfectants in Beijing, China. J. Nippon Med. Sch. 2018, 85, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Bigliardi, P.; Langer, S.; Cruz, J.J.; Kim, S.W.; Nair, H.; Srisawasdi, G. An Asian Perspective on Povidone Iodine in Wound Healing. Dermatology 2017, 233, 223–233. [Google Scholar] [CrossRef]
- Reimer, K.; Wichelhaus, T.A.; Schafer, V.; Rudolph, P.; Kramer, A.; Wutzler, P.; Ganzer, D.; Fleischer, W. Antimicrobial effectiveness of povidone-iodine and consequences for new application areas. Dermatology 2002, 204 (Suppl. S1), 114–120. [Google Scholar] [CrossRef]
- Weber, D.J.; Rutala, W.A. Use of germicides in the home and the healthcare setting: Is there a relationship between germicide use and antibiotic resistance? Infect. Control Hosp. Epidemiol. 2006, 27, 1107–1119. [Google Scholar] [CrossRef]
- Weber, D.J.; Rutala, W.A.; Sickbert-Bennett, E.E. Use of germicides in health care settings-is there a relationship between germicide use and antimicrobial resistance: A concise review. Am. J. Infect. Control 2019, 47S, A106–A109. [Google Scholar] [CrossRef]
- Tezel, U.; Pavlostathis, S.G. Quaternary ammonium disinfectants: Microbial adaptation, degradation and ecology. Curr. Opin. Biotechnol. 2015, 33, 296–304. [Google Scholar] [CrossRef]
- Poole, K. Efflux-mediated antimicrobial resistance. J. Antimicrob. Chemother. 2005, 56, 20–51. [Google Scholar] [CrossRef] [PubMed]
- Putman, M.; van Veen, H.W.; Konings, W.N. Molecular properties of bacterial multidrug transporters. Microbiol. Mol. Biol. Rev. 2000, 64, 672–693. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.M.; Kobayashi, N.; Uehara, N.; Watanabe, N. Analysis on distribution and genomic diversity of high-level antiseptic resistance genes qacA and qacB in human clinical isolates of Staphylococcus aureus. Microb. Drug Resist. 2003, 9, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Shafaati, M.; Boroumand, M.; Nowroozi, J.; Amiri, P.; Kazemian, H. Correlation Between qacE and qacE∆1 Efflux Pump Genes, Antibiotic and Disinfectant Resistant Among Clinical Isolates of E. coli. Recent Pat. Antiinfect. Drug Discov. 2016, 11, 189–195. [Google Scholar] [CrossRef]
- Poole, K. Efflux pumps as antimicrobial resistance mechanisms. Ann. Med. 2007, 39, 162–176. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Antibiotic Class | Representative Chemical Structure | Mode of Action | |
|---|---|---|---|
| Drugs that Target Cell Wall Biosynthesis | β-Lactams: Penicillin Cephalosporins Carbapenems Monobactams |  | Inhibits the synthesis of the peptidoglycan layer of bacterial cell walls by binding to the active site of transpeptidases, known as penicillin-binding proteins (PBPs) [33] |
| Glycopeptides and Lipoglycopeptides |  Vancomycin | Inhibits late stages of cell wall peptidoglycan synthesis by binding to precursors within the cell wall, preventing addition of new units to the peptidoglycan [29] | |
| Drugs that Target Protein Synthesis | Aminoglycosides |  Streptomycin | Inhibits protein synthesis through high-affinity binding to the A-site of the 16S ribosomal RNA of the 30S ribosome [34] |
| Tetracyclines and Alkylaminocyclines |  Tetracycline | Interferes with initiation step of protein synthesis by binding to the ribosomal 30S subunit thereby inhibiting binding of aminoacyl tRNA [29] | |
| Macrolides |  Erythromycin | Inhibits protein synthesis by binding to the peptidyl transferase center at the 50S surface, which causes multiple alterations of the 50S subunit functions [29] | |
| Lincosamides |  Clindamycin | Similar to macrolides [29,35] | |
| Chloramphenicol and Thiaphenicol |  Chloramphenicol | Competitive inhibition for the binding of tRNA to the 50S peptidyltransferase domain. This triggers a conformational change in the ribosome that slows or inhibits aminoacyl tRNA incorporation [29] | |
| Oxazolidinones |  Linezolid | Inhibits protein synthesis by interfering with assembly of the initiation ternary complex of the 30S and 50S ribosomal subunits [29,36] | |
| Drugs that Affect Nucleic Acids | Fluoroquinolones |  Ciprofloxacin | Inhibits the activity of topoisomerases [29] |
| Ansamycins and Lipiarmycins |  Rifampicin | Inhibits the initiation of DNA transcription by binding to the RNA polymerase or the DNA-RNA complex [29] | |
| Antimetabolites | Sulfonamides and Diaminopyrimidines |  Sulfamethoxazole | Inhibits the folate pathway [29] |
| Drugs that Target the Membrane | Lipopeptides |  Daptomycin | Forms micelles (oligomeric assemblies) that interact with the membrane to cause a leakage of cytosolic contents [29,37] |
| Cyclic Polypeptides (Polymyxins/Colistins) |  Polymyxin B | Acts as detergents and alters the permeability of the membrane [29,38] |
| Biocide | Representative Chemical Structure(s) | Mode of Action | ||
|---|---|---|---|---|
| Ionic Interactions | Quaternary Ammonium Compounds (QACs) |  General QAC Structures  Benzalkonium chloride | Acts as a cationic detergent with electrostatic interactions with phospholipids [26,51,52,53,61,62] | |
| Bisbiguanides |  Chlorhexidine | Electrostatic interaction with phospholipids [26,52,53,62] | ||
| Hydrogen bond disruptors | Phenolics |  Triclosan | Not fully understood, but proposed to induce changes in membrane permeability and intracellular functions through hydrogen bonding [51,52,63] At low concentrations, triclosan acts as a site-specific inhibitor of enoyl-acyl carrier protein reductase [57] | |
| Alcohols |  | Solubilizes phospholipids and denatures proteins through disruption of hydrogen bonding [26,51,62] | ||
| Ethanol | Isopropanol | |||
| Chemical reactions | Metals | Ag | Interacts with thiol groups [26,53,54,62] | |
| Chlorine-releasing agents |  Sodium Hypochlorite | Halogenation of amino groups in proteins; oxidation of thiol groups [51,59] | ||
| Fixatives |  Glutaraldehyde  Formaldehyde | Alkylation of biomolecules with amino, imino, amide, carboxyl, and thiol groups (nucleophilic) [51,59] | ||
| Peroxygens |  |  | Oxidizing agents that produces hydroxyl free radicals that attack cell components, e.g., enzyme and protein thiols [26,51,53,54,62,64] | |
| Hydrogen Peroxide | Peracetic Acid | |||
| Iodine |  Povidone−−iodine | Oxidization of thiol groups on proteins, as well as oxidation of nucleotides and fatty acids [53,54,56,64,65] | ||
| Bronopol |  | Oxidizes thiolcontaining materials and produces active oxygen species such as superoxide and peroxide [54,66] | ||
| Ethylene oxide |  | Alkylation of amino and thiol groups in proteins, as well as DNA and RNA [26,67] | ||
| Isothiazolinone |  | Acts as an electrophilic agent reacting with critical enzymes, reacting with thiols on proteins, and producing free radicals [26,55] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coombs, K.; Rodriguez-Quijada, C.; Clevenger, J.O.; Sauer-Budge, A.F. Current Understanding of Potential Linkages between Biocide Tolerance and Antibiotic Cross-Resistance. Microorganisms 2023, 11, 2000. https://doi.org/10.3390/microorganisms11082000
Coombs K, Rodriguez-Quijada C, Clevenger JO, Sauer-Budge AF. Current Understanding of Potential Linkages between Biocide Tolerance and Antibiotic Cross-Resistance. Microorganisms. 2023; 11(8):2000. https://doi.org/10.3390/microorganisms11082000
Chicago/Turabian StyleCoombs, Kent, Cristina Rodriguez-Quijada, Jason O. Clevenger, and Alexis F. Sauer-Budge. 2023. "Current Understanding of Potential Linkages between Biocide Tolerance and Antibiotic Cross-Resistance" Microorganisms 11, no. 8: 2000. https://doi.org/10.3390/microorganisms11082000
APA StyleCoombs, K., Rodriguez-Quijada, C., Clevenger, J. O., & Sauer-Budge, A. F. (2023). Current Understanding of Potential Linkages between Biocide Tolerance and Antibiotic Cross-Resistance. Microorganisms, 11(8), 2000. https://doi.org/10.3390/microorganisms11082000