Abstract
The Buryatian horse is an ancient breed and, as an indigenous breed, they have unique adaptive abilities to use scarce pastures, graze in winter, and survive in harsh conditions with minimal human care. In this study, fecal microbiota of Buryatian horses grazing in the warm and cold seasons were investigated using NGS technology on the Illumina MiSeq platform. We hypothesized that the composition of microbial communities in the feces of horses maintained on pasture would change in the different seasons, depending on the grass availability and different plant diets. We conducted microhistological fecal studies of horse diet composition on steppe pasture. The alpha diversity analysis showed horses had a more abundant and diverse gut microbiota in summer. There were significant effects on the beta diversity of microbial families, which were clustered by the warm and cold season in a principal coordinate analysis (PCoA), with 45% of the variation explained by two principal coordinates. This clustering by season was further confirmed by the significant differences observed in the relative abundances of microbial families and genera. The obtained results can serve as an experimental substantiation for further study of the impact of pasture grasses, which have a pharmacological effect, on the diversity of the gut microbiome and horse health.
1. Introduction
The horse gut microbial community plays a decisive role in maintaining intestinal and organism health [1,2,3]. Several research groups have studied the effects of different factors such as age, breed, environmental conditions (such as season, diet, pasture access, fasting, transportation, and exercise), and the use of pre- and probiotics treatment on the structure of these microbial communities [2,3,4,5,6,7]. It is thought that the ancient horse microbiota was richer and more resilient, conferring protection against environmental challenges, but modern veterinary practices and lifestyles may have changed it [8]. Thus, the comparison of the fecal microbiome from indigenous horses with an ancestral-like lifestyle from different geographical locations enables the characterization of the core horse microbiome and an understanding of the impact of local diets and conditions [1].
The conservation of native domestic animal breeds is among the most important indicators of sustainable agricultural development [9]. Indigenous breeds contribute greatly to the genetic diversity of the world’s domesticated animal populations and have high significance for socio-economic and cultural heritage [10,11]. Indigenous breeds of horses have unique adaptive qualities and are able to use scarce pastures, graze in winter, and survive in harsh conditions with minimal human care [12]. Local environmental conditions, nature, and the nomadic form of husbandry were the prime factors determining the evolution of aboriginal horse breeds [13].
The Buryatian horse is one of the most ancient breeds that are actively used in agriculture today [13]. The Buryatian breed of horse is common in Eastern Siberia and the Far East of Russia, in particular Buryatia and the Trans-Baikal Territory. They are perfectly adapted for breeding using the herd method for keeping on steppe pastures, regardless of the season. The Buryatian native horse is a historically integral part of the local landscape, without which rational nature management and farming would be impossible.
The Buryatian horse comes from the Mongolian breed, with which it has much in common in exterior and biological features. It also has a high level of similarity with the Trans-Baikal breed, which has also showed a high level of genetic diversity [14]. The number of breeding queens of the Buryat breed is estimated at 1000 heads, and its risk status is vulnerable [12].
The existence of the Buryatian horse almost entirely depends on natural and climatic conditions, since it is in the open air on pasture all year round. Based on the zone characteristics of herd horse breeding, the habitat of the Buryatian horses was assessed as sharply continental, cold, and moderately humid [15]. A feature of the temperature regime is large annual amplitudes that reach 70–75 degrees.
The ecology, structure, and diversity of the microbial community of the equine gastrointestinal tract have been the subject of research in recent years [1,2,3,4,5,6,7,8]. The equine hindgut is an anaerobic fermentation ecosystem comprised of thousands of microorganisms that play a critical role in health and energy requirements [4]. Equids are hind-gut fermenters and, unlike ruminants, they can compensate for a low-quality diet by increasing consumption of lower-quality forage or bulk [16].
Differences in the digestive system, extreme environments, and eating habits of horses can lead to distinctive gut microbiota. Changes in the fecal microbiota represent an adaptation to changes in food supply and the environment.
The diversity and community structure of the fecal microbiota of pastured horses varies over a 12-month period, and this variability reflects changes in pasture nutrient composition, which in turn is influenced by climatic conditions [2]. Hindgut microbial adaptation to winter diet occurs to a greater extent in indigenous horses [17].
The bacterial microbiota of the gastrointestinal tract and feces of the Mongolian horse, as the closest relative of the Buryatian horse, was studied, and some interesting findings on the relationship between horse breeds and gut microbiota were revealed [4,5]. It was shown that the relative abundance of 31.25% (5/16) phyla and 40% (30/75) genera was significantly different between Mongolian and Thoroughbred horses living in Inner Mongolia (China) [4]. Studies on the composition of the fecal microbiota of Mongolian horses showed that, although the composition of the gut microbial community was similar to the microbiota of Thoroughbred horses at the phylum level (with the phyla Firmicutes and Bacteroidetes dominating), differences were found at the genus level, explained by environment influences [4,5]. There have been very few studies using metagenomic techniques to examine the dynamics of microbial communities in the feces of horses maintained on pasture [2,18].
The diversity of the fecal microbiota of the Buryatian horse has not yet been studied. We assumed that the composition of microbial communities in the feces of horses maintained on pasture would change in different seasons, depending on grass availability and different plant diets. In our study, we investigated the seasonal fluctuations in the diet composition of steppe pasture and the changes in the structure and composition of fecal microbiota in horses grazing on steppe grass pastures in the warm and cold season. The composition of the fecal microbiota of Buryatian horses was studied using the NGS method for the first time.
2. Materials and Methods
2.1. Sample Collection
Fresh fecal samples were collected from healthy Buryatian horses grazing warm- (July, summer) and cold season (November, meteorological winter) grass pastures. The total number of horses in each group was 14 and consisted of 11 ♂ and 3 ♀, with a mean age of 5.45 years (range, 0.8 to 13 years). All horses were kept on two different farms in the Republic of Buryatia (Russia) for at least 12 months, were grazed on the steppe, and had not experienced any recent changes in housing conditions. The climatic conditions of the region are characterized by a low moisture supply, which consists of 200–350 mm of annual precipitation. The average annual air temperature minimum ranges from −40 to −45 °C, and the absolute minimum reaches −55 °C. The period with temperatures above 0 °C lasts 70–120 days, the sum of temperatures above 10 °C is 1600–1800°. The summer is warm, the average monthly temperature in July is 19.2 °C, and the highest temperatures are 36–38 °C. The mountainous relief to a large extent redistributes the incoming heat and moisture, as a result of which, even in small spaces, habitats with sharply different microclimates are formed [19]. The ambient air temperature at the time of sampling was −7 °C in November and +27 °C in July. The freshly excreted feces were collected in sterilized plastic sealing bags, stored in an icebox, and returned to the laboratory. Samples were collected directly after defecation from the subsurface fecal layer at three points. For DNA extraction, 1 mL of the freshly excreted feces were collected aseptically into fecal collection tubes for safe sample storage and transportation, using a DNA/RNA Shield (Zymo Research, Irvine, CA, USA). For microhistological fecal analyses, 20 mg of the freshly excreted feces was collected in sterilized plastic sealing bags. The samples were transported to the laboratory and were kept in a refrigerator.
2.2. Ethical Considerations
No procedures had to be performed on the animals included in this study. Therefore, ethical approval was not required for this research.
2.3. Microhistological Fecal Analysis
This technique involves preparing a small amount of fecal sample on a slide and then determining the occurrence of individual plant species across up to 100 fields on the slide. Plant species are identified by their histological characteristics, such as cell size and shape and cuticle structure [20,21]. Microhistology is known to provide good precision for diet and is a commonly used tool for many wildlife species [21]. However, it can have a low accuracy, due to differential digestion of plant species, particularly tending to underestimate the abundance of forbs, which can be more completely digested [22]. Microhistology is also labor intensive, requiring extensive training for accurate determination of plant species.
2.4. DNA Extraction, Amplicon Library Construction, Sequencing, and Data Analysis
Total genomic DNA from horse feces was extracted using a Quick-DNA™ Fecal/Soil Microbe Miniprep Kit (Catalog #6010) (Zymo Research, Irvine, California, USA) according to the instructions provided by the manufacturer and stored at −20 °C. The quality and concentration of DNA were assessed spectrophotometrically with a NanoDrop™8000 instrument (Thermo Fischer Scientific, Waltham, MA, USA).
16S rRNA gene amplification and MiSeq sequencing were carried as a part of the project “Large-scale search, study of microorganisms and microbial communities, farm animals and animal products” (https://microbiomeatlas.ru/, accessed on 15 May 2023), according to protocols [23,24]. The protocols showed a high convergence of results. The 16S rRNA gene fragments were amplified using PCR with the primers 341F and 806R [25]. PCR fragments were barcoded using a Nextera XT Index Kit v.2 (Illumina, San Diego, CA, USA) and sequenced on a Illumina MiSeq (2 × 300 nt paired-end reads). Overlapping reads were merged using FLASH v.1.2.11 [26]. Low-quality reads were excluded, and the remaining sequences were clustered into operational taxonomic units (OTUs) at 97% identity using the USEARCH v.11 program [27]. Chimeric sequences were removed during clustering with the USEARCH algorithm. To calculate the relative abundances of OTU, all 16S rRNA gene sequences were mapped to OTU sequences at a 97% global identity threshold using Usearch [28]. OTUs comprising only a single read were discarded. The taxonomic identification of OTUs was performed using searches against the SILVA v.138 rRNA sequence database using the VSEARCH v. 2.14.1 algorithm [28]. 16S rRNA gene fragments were amplified from DNA extracted from other parts of the samples prepared, using a two-stage PCR strategy [29] with the primers 515F [30] and Pro-mod-805R [31]. PCR of every DNA sample was carried out in duplicate; the detailed amplification protocol was described previously [23]. The libraries were checked using agarose gel and pooled equimolarly. The final pool was purified with a QIAquick Gel Extraction Kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol. Sequencing was performed with a MiSeq™ Personal Sequencing System (Illumina, San Diego, CA, USA) using 156-bp paired-end reads.
High quality read pairs were used as an input for the DADA2 pipeline [32], with the SILVA v.138 rRNA sequence database using the VSEARCH v. 2.14.1 algorithm [28]. The taxonomy of the amplified sequence variants was assigned using a naive Bayesian classifier using the Silva138 16S rRNA gene database [33]. The obtained ASV reference sequences, sample metadata, abundance, and taxonomy tables were imported into the phyloseq package [34], and all further operations were performed with the phyloseq object. Decontamination of amplicon data was performed with the decontam R package using the “prevalence” contaminant identification method with default parameters [35]. Rarefaction curve analysis was performed with the iNEXT package [36].
Alpha diversity metrics, including Simpson’s index of diversity, Shannon index of entropy, and the Chao1 index for the richness of bacterial genera were calculated using the phyloseq [34], vegan [37], and microbiome [38] R packages and the MatLab11 package (MatWorks, Inc., Beltsville, MD, USA). Pearson correlations were used to assess the associations between the microbial alpha diversity and seasonal changes. A value of p less than 0.03 was considered to be statistically significant. Relative abundances of microbial taxa were summarized at phylum, class, family, and genus level and included only those microbial taxa that represented >1% of the total community in at least one sample. Microbial phyla, classes, families, and genera with relative abundances <1% in all samples were grouped as “other phyla”, “other class”, “other family”, and “other genera”, respectively. Differences between microbial communities were determined using Bray–Curtis dissimilarity, which takes into account the presence or absence of a species and the relative abundance. A principal coordinate analysis (PCoA) was performed in the MatLab11 package, with the clustering of samples based on the first two principal coordinates. Preliminary processing for data standardization was carried out according to the recommendations in [39].
3. Results
3.1. Microhistology
A common feature of the flora of the studied steppes is the high phytocenotic significance of dense sod grasses, which are indicators of the dry steppes of Southern Siberia (Poa botryoides, Agropyron cristatum, Koeleria cristata) [40]. The role of pasture-resistant plants (Potentilla acaulis, Artemisia frigida, Carex duriuscula) is also significant [19]. The main diversity of grass vegetation in the forest–steppe landscapes of Transbaikalia is made up of mesoxerophyte grass communities, which grow in drier conditions than mesophytes but wetter than xerophytes, mainly meadow steppes with dominant Aster alpinus, Bupleurum scorzonerifolium, Carex pediformis, Potentilla longifolia, Schizonepeta multifida, and others [19]. The most species-rich families are characteristic of the steppe flora of Southern Siberia. The steppe flora of Buryatia contains a number of endemic plants, especially in more southern regions [40]. However, endemics are not distributed everywhere (including in the studied areas) and, moreover, do not play a significant phytocenotic role in the steppe communities, having, as a rule, a very small abundance [19]. No endemic plants were identified in our study. Out of 47 and 23 species found in the summer and winter fecal samples, accordingly (Table 1), there were an average of 18.9 per sample in summer and 10.6 per sample in winter. In winter samples, the diet was mostly made up of forbs (93%, including 51% Schizonepeta multifida), with 7% graminoids and 0.43% shrubs (Table 1). Other grass species found at high abundance in winter diet were Aster alpinus (9.4%), Artemisia sieversiana (8.7%), Heteropappus altaicus (6.8%), and Potentilla acaulis (6.1%). The most abundant graminoids were Poa botryoides (2.9%) and Carex pediformis (2.4%). The diet composition of horse fecal samples in summer using microhistological analysis showed a greater abundance of graminoids (41.9%), including Poa botryoides (14.3%), Agropyron cristatum (11.1%), and Carex pediformis, which was found in all samples and made up 8.9% of the diet. In summer, horse diets were composed of lower proportions of forbs (57.7%) and were mostly made up of Lappula myosotis (8.8%), Silene repens (8.5%), and Aster alpinus (7.8%). Our results showed the diet composition significantly varied for the graminoids Poa botryoides, Agropyron cristatum, and Achnatherum splendens, and the forbs Schizonepeta multifida, Artemisia sieversiana, Lappula myosotis, and Cymbaria dahurica in the warm and cold season (p ≤ 0.003 after Bonferroni correction for multiple comparisons).

Table 1.
Diet composition of Buryatian horse fecal samples from microhistological analyses.
3.2. Analysis of the Fecal Microbiota (Microbial Communities) Using 16S rRNA Profiling
Seasonal-Specific Differences in Diversity of the Fecal Microbial Community
Buryatian horse fecal microbial communities showed a high diversity, both in the warm and cold season (Table S1, Figures S1–S3). The Shannon diversity index of Buryatian horses in the cold season was 5.51, which was significantly lower than that in the warm season (5.71, p < 0.01). This was reflected in the trend of differences in the richness of bacterial genera (Chao1) between the two seasons (Table 2, Figure S1). We found that the Simpson’s indexes were higher in horses grazing in the cold season than in horses in the warm season. However, the level of significance after Bonferroni correction for multiple comparisons revealed non-significant differences among the seasons.
Table 2.
Analysis of alpha diversity at 97% similarity.
The microbial community of both seasons comprised 28 phyla, 14 of which were present at ≥1% abundance of the total community in at least one sample, while the remaining 14 phyla (other phyla) had abundances <1%. Overall, 14 taxa were chosen for comparison at the phylum level. The microbial community was dominated by two phyla, Firmicutes (62.9% in winter; 49.9% in summer) and Bacteroidetes (25.3% in winter; 26.7% in summer), which together accounted for >80% of the overall microbial abundance (Figure 1, Table 2 and Table S1). The fecal archaeal community was the most diverse and abundant in horses grazing warm season grass pasture.
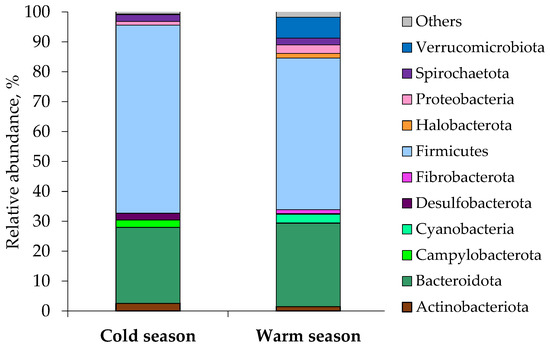
Figure 1.
Mean relative abundance of microbial phyla identified in the fecal samples of Buryatian horses grazing cold and warm season grass pastures. “Others” included Euryarchaeota, Deferribacterota and Synergistota.
In the summer fecal samples, Euryarchaeota and Halobacterota were present at ≥1% abundance in 4 and 5 horses, accordingly. In the winter fecal samples, Euryarchaeota were detected in 3 horses only, but their relative abundance was less than 1%. Other archaeal phyla were not found. Horses grazing warm season grass pasture appeared to have a higher abundance of Cyanobacteria, Fibrobacterota, and Verrucomicrobiota and a lower abundance of Campylobacterota and Desulfobacterota than horses grazing cold season grass pasture (Figure 1, Table 3). Among the top 14 phyla in relative abundance, Firmicutes, Campylobacterota, Desulfobacterota, and Deferribacterota were significantly enriched in the gut of horses in the cold season (Table S2). In contrast, the relative abundance of Verrucomicrobiota, Euryarchaeota, Fibrobacterota, Cyanobacteria, and Synergistota in Buryatian horses was significantly higher in the warm season (Table S2).
Table 3.
Phyla, classes, and families with relative abundance of >1% in the Buryatian horse fecal microbiota.
At a more detailed level, several taxa were significantly less abundant in samples collected in the summer than samples collected in the winter. These were assigned to the bacterial families Lactobacillaceae, Muribaculaceae, Erysipelotrichaceae, Helicobacteraceae, Desulfovibrionaceae, Peptostreptococcaceae, and Bacteroidaceae (Table 3). Other taxa were more abundant in samples collected in the summer than in samples collected in the winter, belonging to the bacterial families Acidaminococcaceae, Rikenellaceae, UCG-010, p-251-o5, F082, WCHB1-41, and unidentified taxa of Order_Gastranaerophilales.
Tannerellaceae, Helicobacteraceae, and Bifidobacteriaceae were not detected in horse guts in summer. Horses grazing in winter lacked WCHB1-41, p-251-o5, and some Bacteroidales families, including BS11_gut_group and UCG-001. Among the top 30 families, the relative abundances of Acidaminococcaceae, Anaerovoracaceae, Bacteroidaceae, Bacteroidales RF16 group, Butyricicoccaceae, Desulfovibrionaceae, Erysipelotrichaceae, F082, Fibrobacteraceae, Lactobacillaceae, Methanobacteriaceae, Muribaculaceae, Clostridia UCG-014, Gastranaerophilales, RF39, Peptostreptococcaceae, Prevotellaceae, Rikenellaceae, Ruminococcaceae, Saccharimonadaceae, Selenomonadaceae, and UCG-010 were significantly different between the two groups (p ≤ 0.003) (Table S3).
There were significant effects of season on the beta diversity of microbial families, which clustered by warm and cold season in the principal coordinate analysis (PCoA), with 45% of the variation explained by two principal coordinates (34.9% on PCs 1 and 10.1% on PCs 2) (Figure 2). This clustering by season was further reinforced by the significant differences observed in the relative abundances of microbial families and microbial genera (Tables S3 and S4).
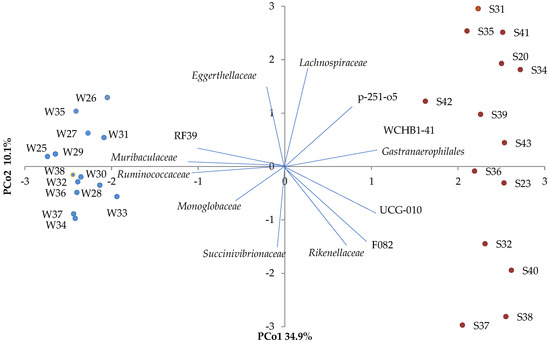
Figure 2.
PCoA of the beta diversity of fecal microbial community in horses illustrated by season, showing the principal coordinate analysis (PCoA) of the beta diversity in the microbial community; with each fecal sample represented as a dot (i.e., each dot represents a horse within a season: W—cold season, S—warm season). The principal coordinates explain 34.9% (PCo1) and 10.1% (PCo2) of the variation.
Among the top 14 genera (Figure 3), the relative abundances of Christensenellaceae_R-7_group, Clostridia_UCG-014, Lachnospiraceae_AC2044_group, Ligilactobacillus, NK4A214_group, Rikenellaceae_RC9_gut_group, UCG-002, and Ruminococcus were significantly different between the two groups (p ≤ 0.003). The relative abundances of Clostridia_UCG-014, Ligilactobacillus, and Ruminococcus in the gut of Buryatian horses were significantly higher in the cold season, while the relative abundances of Christensenellaceae_R-7_group, NK4A214_group, UCG-002, Lachnospiraceae_AC2044_group, and Rikenellaceae_RC9_gut_ group were significantly higher in the warm season (Table S4).
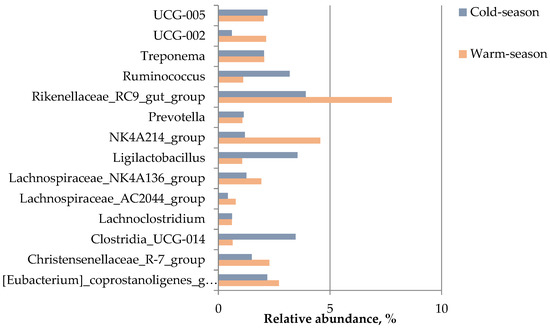
Figure 3.
Top 14 genera in abundance.
4. Discussion
The Buryatian horse is an important breed in Buryatia and the Trans-Baikal Territory (Russia) and is actively used in agriculture today [13]. As an indigenous breed, they have a rich genomic diversity [14] and a variety of excellent traits, such as adaptability, cold resistance, roughage resistance, disease resistance, and good stamina. The harsh climate of this herd horse breeding zone has led to the formation of a distinctive vegetation cover of pastures, which is represented by the presence of sedges, and meadow and subalpine forbs. To the south, closer to lakes and floodplains, forest–tundra vegetation is replaced by forest–steppe and steppe. On the pastures used by the Buryatian horses, 86 species of herbs and several types of shrubs have been identified [15].
Most studies of feral horse diets have used microhistological analyses of fecal matter [21]. We conducted a quantitative synthesis of microhistological fecal studies of the year-round grazing Buryatian horses. Diet composition significantly varied for graminoids and forbs in the warm and cold season (p ≤ 0.03). For wild horses on the western rangelands of North America, the seasonal diet composition consisted of graminoids (77–89%), forbs (4–15%), and browse (3–10%) [21]. Unlike these data, our results indicated that Buryatian horse diets are primarily composed of forbs (93% in the cold season and 57.7% in the warm season). There are many reasons for this difference between the two studies, including regional and climate habitat features, specific shifts in the grass community, and the response to winter snow that limited graminoid accessibility, leading to higher forbs composition. Our results showed the diet composition significantly varied for graminoids and forbs in the warm and cold seasons. Steppe graminoids, Poa botryoides, Agropyron cristatum, and different Carex sp., make up a significant part of the horses’ diet in the warm season. All of these species are resistant to trampling [19]. While animals prefer to eat certain species at the beginning of summer, when the leaves are tender, others can also be used in winter. Agropyron cristatum is a perennial grass that is considered a good fattening feed and provides valuable food from early spring to late autumn. Forbs play an important role throughout the year. The importance of wormwood Artemisia sieversiana increases in horse nutrition in winter. Wormwood has a bitterness and becomes suitable for forage in late autumn after frost. In winter, Potentilla acaulis, which hibernates with a green rosette of leaves and well-formed buds, is maintained in the diet of horses and considered a good fattening food. The importance of Schizonepeta multifida and Heteropappus altaicus, with strong frost resistance, significantly increases in the winter diet. Schizonepeta multifida is both a medicinal and pasture plant [41]. Schizonepeta multifida (or tenuifolia), also known in China as Jing Jie, belongs to the family Lamiaceae and is a perennial herbaceous plant and an herbal medicine that has been widely used for thousands of years in China, Japan, and Korea [42]. As well as the wormwood Artemisia sieversiana, Schizonepeta multifida has clear antimicrobial, antioxidant, immunomodulatory, and anti-inflammatory effects [41,42,43] and is used as medicinal plants in the complex therapy of endogenous infections [44]. Diet is the main factor that contributes to gut microbiome modification in horses [45]. Therefore, the abundance of Schizonepeta multifida and Artemisia sieversiana with high pharmacological effects in the winter diet of the Buryatian horse may be an important factor influencing gut microbiome composition and horse health. The variation in diet composition and the gut microbiota changes observed in the current study may have implications for grazing management and the preparation of conserved forages for horses susceptible to perturbations in hindgut microbial community.
Changes in the composition and diversity of the horse gut microbial communities can reflect feeding patterns and seasonal variations in diet composition, especially in year-round grazing animals [5]. Variation sin pasture composition and fluctuations in the diversity of fecal microbiota were observed for mature adult horses maintained on pasture in New Zealand [2]. Firmicutes and Bacteroidetes were considered important when monitoring changes in microbial populations, because major shifts in bacterial diversity and abundance were found in these two phyla [2]. Firmicutes dominated the gut community when the pasture was growing, whereas when the pasture was drought-stressed, the abundance of Bacteroidetes increased. Similarly, Firmicutes (62.9% in winter; 49.9% in summer) and Bacteroidetes (25.3% in winter; 26.7% in summer) were the main dominant phyla in our study. Actinobacteriota, Campylobacterota, Desulfobacterota, Spirochaetota, and Proteobacteria were other dominant phyla in winter. Verrucomicrobiota, Proteobacteria, Cyanobacteria, Spirochaetota, Halobacterota, Actinobacteriota, and Fibrobacterota had a high relative abundance in summer.
The bacterial microbiota of the gastrointestinal tract and feces of the Mongolian horse, as the closest relative of the Buryatian horse, have been thoroughly explored in studying the gut microbiota differences between Thoroughbred and Mongolian horses [4,5]. Studies on the composition of the fecal microbiota of Mongolian horses grazed on the prairie in September showed Firmicutes, Bacteroidetes, Spirochaetes, Proteobacteria, Verrucomicrobia, and Fibrobacteres were the predominant phyla [4]. The same dominant phyla, accounting for 97.6%, with the addition of Kiritimatiellaeota as another dominant were detected by Wen et al. [5]. Although the composition of the community was similar at the phylum level, the proportion of each phylum was different, except for Firmicutes (56%), which was the most dominant phylum in the gut of Mongolian horses in both studies. The proportion of Bacteroidetes varied from 16% to 33% in Wen’s and Zhao’s studies, accordingly. The proportion of Proteobacteria was 19% for Mongolian horses in Wen’s study, while it was 1% in Zhao’s study.
At the family level, horses grazing cold season grass pasture had larger numbers of two lactic acid bacteria: Lachnospiraceae and Lactobacillaceae, as well as Prevotellaceae. Previous findings showed that increased relative abundance of lactic acid bacteria and excessive lactate production are associated with large intestinal illness in horses [3]. In the current study, we investigated the gut microbiota composition in healthy Buryatian horses, but an increased relative abundance of the lactic acid bacteria Lachnospiraceae was revealed in horses, both in the warm and cold seasons. It is not clear whether this is related to the breed of Buryatian horses, which are perfectly adapted for breeding using the herd method and for keeping on steppe pastures, regardless of the season. However, at the same time, the relative abundance of Escherichia and Streptococcus, which are other markers of horse intestinal diseases [3], were 0.2 and 0.1%, accordingly, in both seasons. Prevotellaceae has enzymes capable of fermenting and utilizing complex polysaccharides [46]. Members of the underexplored family Muribaculaceae were significantly enriched in the horses’ gut in winter but were barely identified in the warm season. Muribaculaceae, as well as members of the Lachnospiraceae, Rikenellaceae, and Bacteroidaceae families, were commensals, which use mucin-derived sugars and impede pathogen colonization [47]. It would be of interest to determine if these taxonomic characteristics have any impact on improving the efficiency of energy and nitrogen utilization in horses, to survive the nutritional stress in severe cold season conditions.
The fecal archaeal community was the most diverse and abundant in horses grazing warm season grass pasture. The dominant archaeal taxa in feces were related to g. Methanocorpusculum and g. Methanobrevibacter. As demonstrated in other studies [48,49,50], methanogenic archaea are often abundant in healthy equine colons. A comparative analysis of the archaeal diversity in horse gut showed that methanogens affiliated to Methanobacteriales and Methanomicrobiales were predominant [50]. The abundance of Methanobrevibacter (Methanobacteriales) is considered a biomarker for monitoring horse intestinal health, because Methanobrevibacter decreases significantly in horses with large and small intestinal colic compared to healthy horses [3]. In the current study, the dominant archaeal taxa of feces were related to Methanocorpusculum in the warm season, while Methanobrevibacter were found in samples from both seasons, but its abundance was reduced in winter.
5. Conclusions
The first attempt to assess the diet composition and taxonomic diversity of fecal microbial communities in healthy Buryatian horses revealed significant differences in microbial community composition among the warm and cold seasons. These findings can serve as an experimental substantiation for further study of the impact of the pasture grasses Schizonepeta multifida and Artemisia sieversiana, which have antimicrobial, antioxidant, immunomodulatory, and anti-inflammatory effects, on the diversity of the gut microbiome and health of horses. The obtained results can improve management treatment protocols for horses suffering from health issues associated with nutrition or feeding.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms11081947/s1, Figure S1: Boxplots of differences in alpha diversity between the warm and cold seasons; Figure S2. Alpha rarefaction curves of studied samples. Figure S3. Heatmap of species abundance clustering at genus level. Table S1: Statistical reads characteristics; Table S2: Comparison of mean relative abundance at phylum level between cold and warm season; Table S3: Comparison of mean relative abundance at family level between cold and warm season; Table S4: Comparison of mean relative abundance at genus level between cold and warm season.
Author Contributions
Conceptualization, S.Z. and D.D.B.; Data curation, A.K. (Anastasia Krylova) and D.D.B.; Formal analysis, O.D., A.K. (Aleksandra Kozlova), A.I., A.K. (Anastasia Krylova), S.N., S.B. and E.P.; Investigation, A.R., A.K. (Aleksandra Kozlova), A.I., A.K. (Anastasia Krylova), S.N., S.B., E.P. and D.D.B.; Methodology, O.D. and A.R.; Software, S.Z. and S.B.; Validation, S.Z. and O.D.; Writing—original draft, S.Z.; Writing—review and editing, O.D. and D.D.B. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Ministry of Science and Higher Education of the Russian Federation in the framework of the Federal scientific-technical program of the genetic technologies development for 2019–2027 (Agreement № 075-15-2021-1401, 3 November 2021).
Data Availability Statement
FASTQ sequences of this metagenomic sample have been deposited in the NCBI Short Read Archive under BioProject PRJNA987555.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ang, L.; Vinderola, G.; Endo, A.; Kantanen, J.; Jingfeng, C.; Binetti, A.; Burns, P.; Qingmiao, S.; Suying, D.; Zujiang, Y.; et al. Gut Microbiome Characteristics in feral and domesticated horses from different geographic locations. Commun. Biol. 2022, 5, 172. [Google Scholar] [CrossRef]
- Fernandes, K.A.; Gee, E.K.; Rogers, C.W.; Kittelmann, S.; Biggs, P.J.; Bermingham, E.N.; Bolwell, C.F.; Thomas, D.G. Seasonal Variation in the Faecal Microbiota of Mature Adult Horses Maintained on Pasture in New Zealand. Animals 2021, 11, 2300. [Google Scholar] [CrossRef]
- Park, T.; Cheong, H.; Yoon, J.; Kim, A.; Yun, Y.; Unno, T. Comparison of the Fecal Microbiota of Horses with Intestinal Disease and Their Healthy Counterparts. Vet. Sci. 2021, 8, 113. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, B.; Bai, D.; Huang, J.; Shiraigo, W.; Yang, L.; Zhao, Q.; Ren, X.; Wu, J.; Bao, W.; et al. Comparison of fecal microbiota of mongolian and thoroughbred horses by high-throughput sequencing of the V4 region of the 16s rRNA gene. Asian-Australas J. Anim. Sci. 2016, 29, 1345–1352. [Google Scholar] [CrossRef]
- Wen, X.; Luo, S.; Lv, D.; Jia, C.; Zhou, X.; Zhai, Q.; Xi, L.; Yang, C. Variations in the fecal microbiota and their functions of Thoroughbred, Mongolian, and Hybrid horses. Front. Vet. Sci. 2022, 9, 920080. [Google Scholar] [CrossRef] [PubMed]
- Weinert-Nelson, J.R.; Biddle, A.S.; Sampath, H.; Williams, C.A. Fecal Microbiota, Forage Nutrients, and Metabolic Responses of Horses Grazing Warm- and Cool-Season Grass Pastures. Animals 2023, 13, 790. [Google Scholar] [CrossRef]
- Theelen, M.J.P.; Luiken, R.E.C.; Wagenaar, J.A.; Sloet van Oldruitenborgh-Oosterbaan, M.M.; Rossen, J.W.A.; Zomer, A.L. The Equine Faecal Microbiota of Healthy Horses and Ponies in The Netherlands: Impact of Host and Environmental Factors. Animals 2021, 11, 1762. [Google Scholar] [CrossRef]
- Metcalf, J.L.; Song, S.J.; Morton, J.T.; Weiss, S.; Seguin-Orlando, A.; Joly, F.; Feh, C.; Taberlet, P.; Coissac, E.; Amir, A.; et al. Evaluating the impact of domestication and captivity on the horse gut microbiome. Sci. Rep. 2017, 7, 15497. [Google Scholar] [CrossRef] [PubMed]
- Prišenk, J.; Filipič, N.; Rozman, Č.; Pažek, K.; Turk, J. Evaluation of Traditional and Indigenous Horse Breeds for Wider Intended Use: Case Study from Slovenia. Sustainability 2022, 14, 1971. [Google Scholar] [CrossRef]
- Mendelsohn, R. The challenge of conserving indigenous domesticated animals. Ecol. Econ. 2003, 45, 501–510. [Google Scholar] [CrossRef]
- Marsoner, T.; Vigl, L.E.; Manck, F.; Jaritz, G.; Tappeiner, U.; Tasser, E. Indigenous livestock breeds as indicators for cultural ecosystem services: A spatial analysis within the Alpine Space. Ecol. Indic. 2018, 94, 55–63. [Google Scholar] [CrossRef]
- Khrabrova, L.A. Preservation and use of the gene pool of native breeds of horses. Eff. Anim. Husb. 2016, 4, 33–35. (In Russian) [Google Scholar]
- Taishin, V.A.; Lkhasaranov, B.B.; James, R. Atlas of Nomadic Animals; Publishing House of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 1999; 284p. [Google Scholar]
- Khrabrova, L.A.; Blohina, N.V.; Bazaron, B.Z.; Khamiruev, T.N. Variability of mitochondrial DNA D-loop sequences in Zabaikalskaya horse breed. Vavilov J. Genet. Breed. 2021, 25, 486–491. [Google Scholar] [CrossRef]
- Kalashnikov, I.A. Scientific and Practical Aspects of Conservation, Selection and Use of Horses of Local Aboriginal Breeds (on the Example of the Buryatian Horse). Thesis of Doctor’s Degree, Moscow Timiryazev Agricultural Academy, Moscow, Russia, 1997; 36p. [Google Scholar]
- Schoenecker, K.A.; King, S.R.B.; Nordquist, M.K.; Nandintsetseg, D.; Cao, Q. Habitat and diet of equids. In Wild Equids: Ecology, Management, and Conservation; Ransom, J.I., Kaczensky, P., Eds.; EdsJohns Hopkins University Press: Baltimore, MD, USA, 2016; pp. 41–57. [Google Scholar]
- Kobayashi, Y.; Koike, S.; Miyaji, M.; Hata, H.; Tanaka, K. Hindgut microbes, fermentation and their seasonal variations in Hokkaido native horses compared to light horses. Ecol. Res. 2006, 21, 285–291. [Google Scholar] [CrossRef]
- Salem, S.E.; Maddox, T.W.; Berg, A.; Antczak, P.; Ketley, J.M.; Williams, N.J.; Archer, D.C. Variation in faecal microbiota in a group of horses managed on pasture over a 12-month period. Sci. Rep. 2018, 8, 8510. [Google Scholar] [CrossRef] [PubMed]
- Korolyuk, A.Y. Syntaxonomy of steppe vegetation of the Republic of Buryatia. Veg. Russ. 2017, 31, 3–32. [Google Scholar] [CrossRef]
- King, S.R.B.; Schoenecker, K.A. Comparison of Methods to Examine Diet of Feral Horses from Noninvasively Collected Fecal Samples. Rangel. Ecol. Manag. 2019, 72, 661–666. [Google Scholar] [CrossRef]
- Scasta, J.D.; Beck, J.L.; Anqwin, C.J. Meta-Analysis of Diet Composition and Potential Conflict of Wild Horses with Livestock and Wild Ungulates on Western Rangelands of North America. Rangel. Ecol. Manag. 2016, 69, 309–318. [Google Scholar] [CrossRef]
- Holechek, J.L.; Vavra, M.; Pieper, R.D. Botanical composition determination of range herbivore diets: A review. J. Range Manag. 1982, 35, 309–315. [Google Scholar] [CrossRef]
- Toshchakov, S.V.; Izotova, A.O.; Vinogradova, E.N.; Kachmazov, G.S.; Tuaeva, A.Y.; Abaev, V.T.; Evteeva, M.A.; Gunitseva, N.M.; Korzhenkov, A.A.; Elcheninov, A.G.; et al. Culture-Independent Survey of Thermophilic Microbial Communities of the North Caucasus. Biology 2021, 10, 1352. [Google Scholar] [CrossRef]
- Karnachuk, O.V.; Panova, I.A.; Panov, V.L.; Ikkert, O.P.; Kadnikov, V.V.; Rusanov, I.I.; Avakyan, M.P.; Glukhova, L.B.; Rakitin, A.V.; Begmatov, S.; et al. Active Sulfate-Reducing Bacterial Community in the Camel Gut. Microorganisms 2023, 11, 401. [Google Scholar] [CrossRef]
- Frey, B.; Rime, T.; Phillips, M.; Stierli, B.; Hajdas, I.; Widmer, F.; Hartmann, M. Microbial diversity in European alpine permafrost and active layers. FEMS Microbiol. Ecol. 2016, 92, fiw018. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Gohl, D.; Gohl, D.M.; MacLean, A.; Hauge, A.; Becker, A.; Walek, D.; Beckman, K.B. An Optimized Protocol for High-Throughput Amplicon-Based Microbiome Profiling. Protoc. Exch. 2016, 1–15. [Google Scholar] [CrossRef]
- Hugerth, L.W.; Wefer, H.A.; Lundin, S.; Jakobsson, H.E.; Lindberg, M.; Rodin, S.; Engstrand, L.; Andersson, A.F. DegePrime, a Program for Degenerate Primer Design for Broad-Taxonomic-Range PCR in Microbial Ecology Studies. Appl. Environ. Microbiol. 2014, 80, 5116–5123. [Google Scholar] [CrossRef]
- Merkel, A.Y.; Tarnovetskii, I.Y.; Podosokorskaya, O.A.; Toshchakov, S.V. Analysis of 16S RRNA Primer Systems for Profiling of Thermophilic Microbial Communities. Microbiology 2019, 88, 671–680. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Davis, N.M.; Proctor, D.M.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple Statistical Identification and Removal of Contaminant Sequences in Marker-Gene and Metagenomics Data. Microbiome 2018, 6, 226. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.C.; Ma, K.H.; Chao, A. INEXT: An R Package for Rarefaction and Extrapolation of Species Diversity (H Ill Numbers). Methods Eco. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Vegan: Community Ecology Package. Ordination Methods, Diversity Analysis and Other Functions for Community and Vegetation Ecologists. Available online: https://www.worldagroforestry.org/publication/vegan-community-ecology-package-ordination-methods-diversity-analysis-and-other (accessed on 19 November 2022).
- Lahti, L.; Shetty, S.; Blake, T.; Salojarvi, J. Microbiome; Version 1.0.2; Bioconductor: 2017. Available online: https://bioconductor.statistik.tutortmund.de/packages/3.6/bioc/html/microbiome.html (accessed on 19 November 2022).
- Zuur, A.F.; Ieno, E.N.; Smith, G.M. Analyzing Ecological Data; Springer: New York, NY, USA, 2007; p. 672. [Google Scholar]
- Peshkova, G.A. Florogenetic Analysis of the Steppe Flora of the Mountains of Southern Siberia; Science: Novosibirsk, Russia, 2001; 192p. (In Russian) [Google Scholar]
- Amarjargal, A.; Bayarkhuu, Y.; Bayarkhuu, B. Distribution and some results of chemical studies in Schizonepeta multifida L. Vestn. Tuvan State Univ. 2022, 3, 17–29. [Google Scholar]
- Zhao, X.; Zhou, M. Review on Chemical Constituents of Schizonepeta tenuifolia Briq. and Their Pharmacological Effects. Molecules 2022, 27, 5249. [Google Scholar] [CrossRef]
- Mojtahed Zadeh Asl, R.; Niakousari, M.; Hashemi Gahruie, H.; Saharkhiz, M.J.; Mousavi Khaneghah, A. Study of two-stage ohmic hydro-extraction of essential oil from Artemisia aucheri Boiss.: Antioxidant and antimicrobial characteristics. Food Res. Int. 2018, 107, 462–469. [Google Scholar] [CrossRef]
- Kļaviņa, A.; Keidāne, D.; Šukele, R.; Bandere, D.; Kovaļčuka, L. Traditional Latvian herbal medicinal plants used to treat parasite infections of small ruminants: A review. Vet. World 2021, 14, 1548–1558. [Google Scholar] [CrossRef]
- Lara, F.; Castro, R.; Thomson, P. Changes in the gut microbiome and colic in horses: Are they causes or consequences? Open Vet. J. 2022, 12, 242–249. [Google Scholar] [CrossRef]
- Qiu, X.; Qin, X.; Chen, L.; Chen, Z.; Hao, R.; Zhang, S.; Yang, S.; Wang, L.; Cui, Y.; Li, Y.; et al. Serum Biochemical Parameters, Rumen Fermentation, and Rumen Bacterial Communities Are Partly Driven by the Breed and Sex of Cattle When Fed High-Grain Diet. Microorganisms 2022, 10, 323. [Google Scholar] [CrossRef]
- Pereira, F.C.; Wasmund, K.; Cobankovic, I.; Jehmlich, N.; Herbold, C.W.; Lee, K.S.; Sziranyi, B.; Vesely, C.; Decker, T.; Stocker, R.; et al. Rational design of a microbial consortium of mucosal sugar utilizers reduces Clostridiodes difficile colonization. Nat. Commun. 2020, 11, 5104. [Google Scholar] [CrossRef]
- Dougal, K.; Harris, P.A.; Edwards, A.; Pachebat, J.A.; Blackmore, T.M.; Worgan, H.J.; Newbold, C.J. A comparison of the microbiome and the metabolome of different regions of the equine hindgut. FEMS Microbiol. Ecol. 2012, 82, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Fliegerova, K.; Mura, E.; Mrázek, J.; Moniello, G. A comparison of microbial profiles of different regions of the equine hindgut. Livest. Sci. 2016, 190, 16–19. [Google Scholar] [CrossRef]
- Lwin, K.O.; Matsui, H. Comparative analysis of the methanogen diversity in horse and pony by using mcrA gene and archaeal 16s rRNA gene clone libraries. Archaea 2014, 2014, 483574. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).