


Isolation and Characterization of Bacterial Endophytes from Small Nodules of Field-Grown Peanut

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of Bacterial Isolates from Small Nodules of Peanut Roots
2.2. Identification and Molecular Characterization of Bacterial Isolates
2.3. Biochemical Analysis of Isolates for Growth-Promoting Traits
2.3.1. Ammonia (NH3) Production Assay
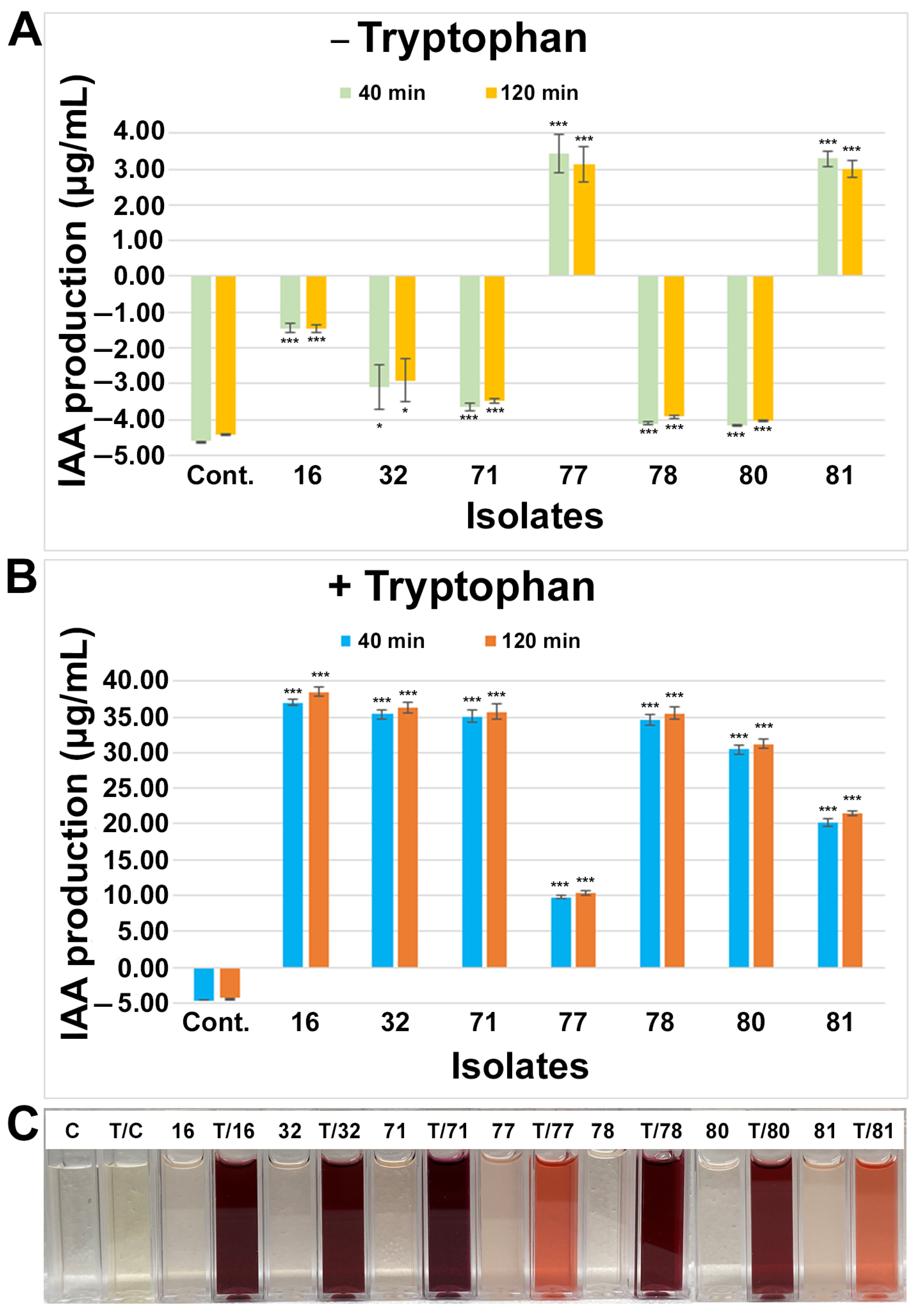
2.3.2. Indole-3-Acetic Acid (IAA) Production Assay
2.3.3. P-Solubilization Assay (Agar Plate Method)
2.3.4. Siderophore Production Assay
2.3.5. ACCD Activity (Plate Assay)
2.3.6. Tolerance of Isolates to High Temperature, NaCl, and Low pH
2.4. Evaluation of Plant Growth Promotion (PGP) Ability with the Selected Endophytes
3. Results
3.1. Isolation of Bacterial Endophytes from Small Nodules of Peanut Roots
3.2. Molecular Identification of Small Nodule Endophytes of Peanut Roots
3.3. Characterization of Growth Promoting Traits of Small Nodule Endophytes from Peanut Roots
3.4. Plant Growth Promotion (PGP) Assay
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guo, K.; Yang, J.; Yu, N.; Luo, L.; Wang, E. Biological Nitrogen Fixation in Cereal Crops: Progress, Strategies, and Perspectives. Plant Commun. 2023, 4, 100499. [Google Scholar] [CrossRef] [PubMed]
- Peix, A.; Ramírez-Bahena, M.H.; Velázquez, E.; Bedmar, E.J. Bacterial Associations with Legumes. Crit. Rev. Plant Sci. 2015, 34, 17–42. [Google Scholar] [CrossRef]
- Lòpez-Fernàndez, S.; Mazzoni, V.; Pedrazzoli, F.; Pertot, I.; Campisano, A. A Phloem-Feeding Insect Transfers Bacterial Endophytic Communities between Grapevine Plants. Front. Microbiol. 2017, 8, 834. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, P.; Alves, A.; Silveira, P.; Sá, C.; Fidalgo, C.; Freitas, R.; Figueira, E. Bacteria from Nodules of Wild Legume Species: Phylogenetic Diversity, Plant Growth Promotion Abilities and Osmotolerance. Sci. Total Environ. 2018, 645, 1094–1102. [Google Scholar] [CrossRef]
- Rajendran, G.; Patel, M.H.; Joshi, S.J. Isolation and Characterization of Nodule-Associated Exiguobacterium Sp. from the Root Nodules of Fenugreek (Trigonella Foenum-Graecum) and Their Possible Role in Plant Growth Promotion. Int. J. Microbiol. 2012, 2012, 1–8. [Google Scholar] [CrossRef]
- Ferchichi, N.; Toukabri, W.; Boularess, M.; Smaoui, A.; Mhamdi, R.; Trabelsi, D. Isolation, Identification and Plant Growth Promotion Ability of Endophytic Bacteria Associated with Lupine Root Nodule Grown in Tunisian Soil. Arch. Microbiol. 2019, 201, 1333–1349. [Google Scholar] [CrossRef]
- Preyanga, R.; Anandham, R.; Krishnamoorthy, R.; Senthilkumar, M.; Gopal, N.O.; Vellaikumar, A.; Meena, S. Groundnut (Arachis Hypogaea) Nodule Rhizobium and Passenger Endophytic Bacterial Cultivable Diversity and Their Impact on Plant Growth Promotion. Rhizosphere 2021, 17, 100309. [Google Scholar] [CrossRef]
- Ibañez, F.; Arroyo, M.E.; Angelini, J.; Tonelli, M.L.; Muñoz, V.; Ludueña, L.; Valetti, L.; Fabra, A. Non-Rhizobial Peanut Nodule Bacteria Promote Maize (Zea Mays L.) and Peanut (Arachis Hypogaea L.) Growth in a Simulated Crop Rotation System. Appl. Soil Ecol. 2014, 84, 208–212. [Google Scholar] [CrossRef]
- Mansour, S.; Elhaloos, B.; Abdel-Lateif, K. Phenotypic and Genetic Diversity of Native Rhizobium Isolated from Root Nodules of Leguminous Plants Grown in Reclaimed Soil, Egypt. SABRAOJBG 2023, 52, 344–357. [Google Scholar] [CrossRef]
- Patra, D.; Mandal, S. Non-Rhizobia Are the Alternative Sustainable Solution for Growth and Development of the Nonlegume Plants. Biotechnol. Genet. Eng. Rev. 2022, 2022, 1–30. [Google Scholar] [CrossRef]
- Dudeja, S.S.; Giri, R.; Saini, R.; Suneja-Madan, P.; Kothe, E. Interaction of Endophytic Microbes with Legumes. J. Basic Microbiol. 2012, 52, 248–260. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H. Root Nodules of Legumes: A Suitable Ecological Niche for Isolating Non-Rhizobial Bacteria with Biotechnological Potential in Agriculture. Curr. Res. Biotechnol. 2022, 4, 78–86. [Google Scholar] [CrossRef]
- Ahmad, E.; Sharma, P.K.; Khan, M.S. IAA Biosynthesis in Bacteria and Its Role in Plant-Microbe Interaction for Drought Stress Management. In Plant Stress Mitigators; Vaishnav, A., Arya, S.S., Choudhary, D.K., Eds.; Springer Nature Singapore: Singapore, 2022; ISBN 9789811677588. [Google Scholar]
- Bhutani, N.; Maheshwari, R.; Negi, M.; Suneja, P. Optimization of IAA Production by Endophytic Bacillus Spp. from Vigna Radiata for Their Potential Use as Plant Growth Promoters. Isr. J. Plant Sci. 2018, 65, 83–96. [Google Scholar] [CrossRef]
- Pandya, M.; Rajput, M.; Rajkumar, S. Exploring Plant Growth Promoting Potential of Non Rhizobial Root Nodules Endophytes of Vigna Radiata. Microbiology 2015, 84, 80–89. [Google Scholar] [CrossRef]
- Kramer, J.; Özkaya, Ö.; Kümmerli, R. Bacterial Siderophores in Community and Host Interactions. Nat. Rev. Microbiol. 2020, 18, 152–163. [Google Scholar] [CrossRef]
- Saleem, A.R.; Brunetti, C.; Khalid, A.; Della Rocca, G.; Raio, A.; Emiliani, G.; De Carlo, A.; Mahmood, T.; Centritto, M. Drought Response of Mucuna Pruriens (L.) DC. Inoculated with ACC Deaminase and IAA Producing Rhizobacteria. PLoS ONE 2018, 13, e0191218. [Google Scholar] [CrossRef]
- Marra, L.M.; de Oliveira-Longatti, S.M.; Soares, C.R.F.S.; Olivares, F.L.; Moreira, F.M.D.S. The Amount of Phosphate Solubilization Depends on the Strain, C-Source, Organic Acids and Type of Phosphate. Geomicrobiol. J. 2019, 36, 232–242. [Google Scholar] [CrossRef]
- Jha, B.; Gontia, I.; Hartmann, A. The Roots of the Halophyte Salicornia Brachiata Are a Source of New Halotolerant Diazotrophic Bacteria with Plant Growth-Promoting Potential. Plant Soil 2012, 356, 265–277. [Google Scholar] [CrossRef]
- Mukhtar, S.; Hirsch, A.M.; Khan, N.; Malik, K.A.; Humm, E.A.; Pellegrini, M.; Shi, B.; Briscoe, L.; Huntemann, M.; Clum, A.; et al. Impact of Soil Salinity on the Cowpea Nodule-Microbiome and the Isolation of Halotolerant PGPR Strains to Promote Plant Growth under Salinity Stress. Phytobiomes J. 2020, 4, 364–374. [Google Scholar] [CrossRef]
- Ruiz-Díez, B.; Fajardo, S.; Puertas-Mejía, M.A.; de Felipe, M.D.R.; Fernández-Pascual, M. Stress Tolerance, Genetic Analysis and Symbiotic Properties of Root-Nodulating Bacteria Isolated from Mediterranean Leguminous Shrubs in Central Spain. Arch Microbiol. 2009, 191, 35–46. [Google Scholar] [CrossRef]
- Hammons, R.O.; Herman, D.; Stalker, H.T. Origin and Early History of the Peanut. In Peanuts; Elsevier: Amsterdam, The Netherlands, 2016; ISBN 978-1-63067-038-2. [Google Scholar]
- Desmae, H.; Janila, P.; Okori, P.; Pandey, M.K.; Motagi, B.N.; Monyo, E.; Mponda, O.; Okello, D.; Sako, D.; Echeckwu, C.; et al. Genetics, Genomics and Breeding of Groundnut (Arachis Hypogaea L.). Plant Breed 2019, 138, 425–444. [Google Scholar] [CrossRef] [PubMed]
- Tajima, R.; Lee, O.N.; Abe, J.; Lux, A.; Morita, S. Nitrogen-Fixing Activity of Root Nodules in Relation to Their Size in Peanut (Arachis Hypogaea L.). Plant Prod. Sci. 2007, 10, 423–429. [Google Scholar] [CrossRef]
- Hossain, M.S.; DeLaune, P.B.; Gentry, T.J. Microbiome Analysis Revealed Distinct Microbial Communities Occupying Different Sized Nodules in Field-Grown Peanut. Front. Microbiol. 2023, 14, 1075575. [Google Scholar] [CrossRef] [PubMed]
- Haldar, S.; Sengupta, S. Impact of Plant Development on the Rhizobacterial Population of Arachis Hypogaea: A Multifactorial Analysis: Bacterial Community Dynamics in Arachis Rhizosphere. J. Basic Microbiol. 2015, 55, 922–928. [Google Scholar] [CrossRef]
- Dai, L.; Zhang, G.; Yu, Z.; Ding, H.; Xu, Y.; Zhang, Z. Effect of Drought Stress and Developmental Stages on Microbial Community Structure and Diversity in Peanut Rhizosphere Soil. IJMS 2019, 20, 2265. [Google Scholar] [CrossRef]
- Hinsu, A.T.; Panchal, K.J.; Pandit, R.J.; Koringa, P.G.; Kothari, R.K. Characterizing Rhizosphere Microbiota of Peanut (Arachis Hypogaea L.) from Pre-Sowing to Post-Harvest of Crop under Field Conditions. Sci. Rep. 2021, 11, 17457. [Google Scholar] [CrossRef]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.-Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M.; et al. Evaluation of 16S RRNA Gene Sequencing for Species and Strain-Level Microbiome Analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glockner, F.O. SILVA: A Comprehensive Online Resource for Quality Checked and Aligned Ribosomal RNA Sequence Data Compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, Interactive Sequence Choice and Visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Singh, P.; Singh, R.K.; Li, H.-B.; Guo, D.-J.; Sharma, A.; Lakshmanan, P.; Malviya, M.K.; Song, X.-P.; Solanki, M.K.; Verma, K.K.; et al. Diazotrophic Bacteria Pantoea Dispersa and Enterobacter Asburiae Promote Sugarcane Growth by Inducing Nitrogen Uptake and Defense-Related Gene Expression. Front. Microbiol. 2021, 11, 600417. [Google Scholar] [CrossRef]
- Mohite, B. Isolation and Characterization of Indole Acetic Acid (IAA) Producing Bacteria from Rhizospheric Soil and Its Effect on Plant Growth. J. Soil Sci. Plant Nutr. 2013, 13, 638–649. [Google Scholar] [CrossRef]
- Gordon, S.A.; Weber, R.P. Colorimetric estimation of indoleacetic acid. Plant Physiol. 1951, 26, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhao, G.; Wei, Y.; Dong, Y.; Hou, L.; Jiao, R. Isolation and Screening of Multifunctional Phosphate Solubilizing Bacteria and Its Growth-Promoting Effect on Chinese Fir Seedlings. Sci. Rep. 2021, 11, 9081. [Google Scholar] [CrossRef]
- Pikovskaya, R.I. Mobilization of Phosphorus in Soil Connection with the Vital Activity of Some Microbial Species. Microbiology 1984, 17, 362–370. [Google Scholar]
- Pande, A.; Pandey, P.; Mehra, S.; Singh, M.; Kaushik, S. Phenotypic and Genotypic Characterization of Phosphate Solubilizing Bacteria and Their Efficiency on the Growth of Maize. J. Genet. Eng. Biotechnol. 2017, 15, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Virpiranta, H.; Banasik, M.; Taskila, S.; Leiviskä, T.; Halttu, M.; Sotaniemi, V.-H.; Tanskanen, J. Isolation of Efficient Metal-Binding Bacteria from Boreal Peat Soils and Development of Microbial Biosorbents for Improved Nickel Scavenging. Water 2020, 12, 2000. [Google Scholar] [CrossRef]
- Maheshwari, R.; Bhutani, N.; Suneja, P. Isolation and Characterization of ACC Deaminase Producing Endophytic Bacillus Mojavensis PRN2 from Pisum Sativum. Iran J. Biotch. 2020, 18, 2308. [Google Scholar] [CrossRef]
- Dworkin, M.; Foster, J.W. Experiments with some microorganisms which utilize ethane and hydrogen. J. Bacteriol. 1958, 75, 592–603. [Google Scholar] [CrossRef]
- Sobolev, V.S.; Orner, V.A.; Arias, R.S. Distribution of Bacterial Endophytes in Peanut Seeds Obtained from Axenic and Control Plant Material under Field Conditions. Plant Soil 2013, 371, 367–376. [Google Scholar] [CrossRef]
- Huang, Y.; Kuang, Z.; Deng, Z.; Zhang, R.; Cao, L. Endophytic Bacterial and Fungal Communities Transmitted from Cotyledons and Germs in Peanut (Arachis Hypogaea L.) Sprouts. Environ. Sci. Pollut. Res. 2017, 24, 16458–16464. [Google Scholar] [CrossRef]
- Li, L.; Zhang, Z.; Pan, S.; Li, L.; Li, X. Characterization and Metabolism Effect of Seed Endophytic Bacteria Associated With Peanut Grown in South China. Front. Microbiol. 2019, 10, 2659. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.S.; Zhao, L.F.; Kong, Z.Y.; Yang, W.Q.; Lindström, K.; Wang, E.T.; Wei, G.H. Diversity of Endophytic Bacteria within Nodules of the Sphaerophysa Salsula in Different Regions of Loess Plateau in China: Sphaerophysa Salsula Endophytic Bacteria. FEMS Microbiol. Ecol. 2011, 76, 463–475. [Google Scholar] [CrossRef]
- Martínez-Hidalgo, P.; Hirsch, A.M. The Nodule Microbiome: N 2 -Fixing Rhizobia Do Not Live Alone. Phytobiomes J. 2017, 1, 70–82. [Google Scholar] [CrossRef]
- Aserse, A.A.; Räsänen, L.A.; Aseffa, F.; Hailemariam, A.; Lindström, K. Diversity of Sporadic Symbionts and Nonsymbiotic Endophytic Bacteria Isolated from Nodules of Woody, Shrub, and Food Legumes in Ethiopia. Appl. Microbiol. Biotechnol. 2013, 97, 10117–10134. [Google Scholar] [CrossRef]
- Wigley, K.; Moot, D.; Wakelin, S.A.; Laugraud, A.; Blond, C.; Seth, K.; Ridgway, H. Diverse Bacterial Taxa Inhabit Root Nodules of Lucerne (Medicago Sativa L.) in New Zealand Pastoral Soils. Plant Soil 2017, 420, 253–262. [Google Scholar] [CrossRef]
- Omar Ahmed Idris, A.; Alamin Gibreel Noh, E. Legumes and nodule associated bacteria interaction as key factor for abiotic stresses impact mitigation. In Legumes Research—Volume 1; Jimenez-Lopez, C.J., Clemente, A., Eds.; IntechOpen: London, UK, 2022; ISBN 978-1-83969-490-5. [Google Scholar]
- Gupta, S.; Pandey, S.; Kotra, V.; Kumar, A. Assessing the Role of ACC Deaminase-Producing Bacteria in Alleviating Salinity Stress and Enhancing Zinc Uptake in Plants by Altering the Root Architecture of French Bean (Phaseolus vulgaris) Plants. Planta 2023, 258, 3. [Google Scholar] [CrossRef] [PubMed]
- Ratnaningsih, H.R.; Noviana, Z.; Dewi, T.K.; Loekito, S.; Wiyono, S.; Gafur, A.; Antonius, S. IAA and ACC Deaminase Producing-Bacteria Isolated from the Rhizosphere of Pineapple Plants Grown under Different Abiotic and Biotic Stresses. Heliyon 2023, 9, e16306. [Google Scholar] [CrossRef] [PubMed]
- Amy, C.; Avice, J.-C.; Laval, K.; Bressan, M. Are Native Phosphate Solubilizing Bacteria a Relevant Alternative to Mineral Fertilizations for Crops? Part I. When Rhizobacteria Meet Plant P Requirements. Rhizosphere 2022, 21, 100476. [Google Scholar] [CrossRef]
- Kumar, A.; Tripti; Maleva, M.; Bruno, L.B.; Rajkumar, M. Synergistic Effect of ACC Deaminase Producing Pseudomonas Sp. TR15a and Siderophore Producing Bacillus Aerophilus TR15c for Enhanced Growth and Copper Accumulation in Helianthus Annuus L. Chemosphere 2021, 276, 130038. [Google Scholar] [CrossRef]
- Aeron, A.; Maheshwari, D.K.; Meena, V.S. Endophytic Bacteria Promote Growth of the Medicinal Legume Clitoria Ternatea L. by Chemotactic Activity. Arch. Microbiol. 2020, 202, 1049–1058. [Google Scholar] [CrossRef]
- Gupta, S.; Pandey, S. ACC Deaminase Producing Bacteria With Multifarious Plant Growth Promoting Traits Alleviates Salinity Stress in French Bean (Phaseolus vulgaris) Plants. Front. Microbiol. 2019, 10, 1506. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate ID | % Identity with ASVs | ASV Fragment Size (bp) | Isolate Fragment Size (bp) | Genus |
|---|---|---|---|---|
| 1 | 100.00 | 445 | 1107 | Pantoea |
| 2 | 100.00 | 445 | 1098 | Pantoea |
| 16 | 100.00 | 420 | 1145 | Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium |
| 17 | 100.00 | 445 | 1195 | Burkholderia-Caballeronia-Paraburkholderia |
| 18 | 100.00 | 445 | 1117 | Mitsuaria |
| 22 | 100.00 | 420 | 1165 | Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium |
| 23 | 100.00 | 445 | 1104 | Herbaspirillum |
| 25 | 100.00 | 445 | 1159 | Mitsuaria |
| 30 | 100.00 | 445 | 1113 | Pseudomonas |
| 32 | 100.00 | 445 | 1154 | Mitsuaria |
| 33 | 100.00 | 420 | 1140 | Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium |
| 39 | 100.00 | 446 | 1118 | Bacillus |
| 42 | 100.00 | 446 | 1029 | Bacillus |
| 43 | 100.00 | 445 | 1004 | Mitsuaria |
| 49 | 100.00 | 446 | 1030 | Bacillus |
| 50 | 100.00 | 446 | 1022 | Bacillus |
| 56 | 100.00 | 446 | 1067 | Bacillus |
| 57 | 100.00 | 446 | 1075 | Bacillus |
| 58 | 100.00 | 446 | 1104 | Bacillus |
| 59 | 100.00 | 446 | 1087 | Bacillus |
| 60 | 100.00 | 446 | 1123 | Bacillus |
| 61 | 100.00 | 446 | 1029 | Bacillus |
| 62 | 100.00 | 446 | 1075 | Bacillus |
| 63 | 100.00 | 446 | 1053 | Bacillus |
| 64 | 100.00 | 446 | 1088 | Bacillus |
| 65 | 100.00 | 446 | 1071 | Bacillus |
| 66 | 100.00 | 446 | 1122 | Bacillus |
| 67 | 100.00 | 446 | 1095 | Bacillus |
| 68 | 100.00 | 446 | 1159 | Bacillus |
| 69 | 100.00 | 446 | 1095 | Bacillus |
| 77 | 100.00 | 420 | 1143 | Rhizobium sp. |
| 84 | 100.00 | 420 | 1232 | Rhizobium sp. CT 6-01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, M.S.; Frith, C.; Bhattacharyya, S.S.; DeLaune, P.B.; Gentry, T.J. Isolation and Characterization of Bacterial Endophytes from Small Nodules of Field-Grown Peanut. Microorganisms 2023, 11, 1941. https://doi.org/10.3390/microorganisms11081941
Hossain MS, Frith C, Bhattacharyya SS, DeLaune PB, Gentry TJ. Isolation and Characterization of Bacterial Endophytes from Small Nodules of Field-Grown Peanut. Microorganisms. 2023; 11(8):1941. https://doi.org/10.3390/microorganisms11081941
Chicago/Turabian StyleHossain, Md Shakhawat, Christine Frith, Siddhartha Shankar Bhattacharyya, Paul B. DeLaune, and Terry J. Gentry. 2023. "Isolation and Characterization of Bacterial Endophytes from Small Nodules of Field-Grown Peanut" Microorganisms 11, no. 8: 1941. https://doi.org/10.3390/microorganisms11081941
APA StyleHossain, M. S., Frith, C., Bhattacharyya, S. S., DeLaune, P. B., & Gentry, T. J. (2023). Isolation and Characterization of Bacterial Endophytes from Small Nodules of Field-Grown Peanut. Microorganisms, 11(8), 1941. https://doi.org/10.3390/microorganisms11081941