
Old Folks, Bad Boon: Antimicrobial Resistance in the Infant Gut Microbiome

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. The Study of Antimicrobial Resistance in the Human Microbiome
2.1. Omics Techniques for Monitoring ARG in the Microbiome
2.2. Challenges in Identifying ARGs in the Microbiome
3. The Infant Intestinal Resistome
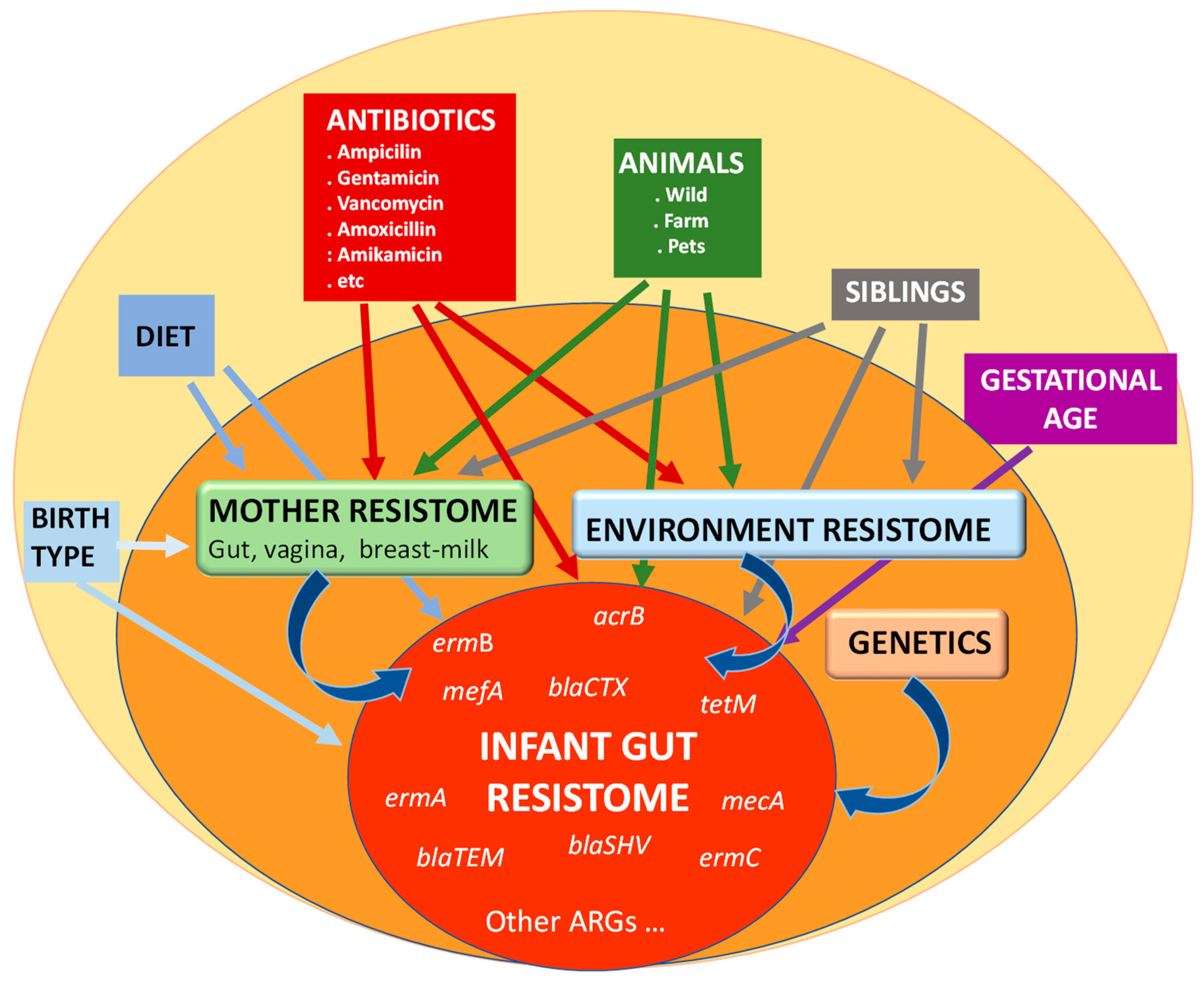
4. The Origin of Infant Resistome: Vertical Transmission of ARGs
4.1. Pregnancy Microbiota Changes
4.2. ARGs in Pregnancy and Lactation
4.3. The Maternal–Filial Transfer Route as a Source of ARGs for the Neonate
5. Impact of Antibiotics on the Developing Microbiota Composition
5.1. Antibiotics Use in Early Life
5.1.1. Use of Antibiotics in Neonatology
5.1.2. Use of Antibiotics in Infants and Older Children
5.2. Impact of Antibiotic Use on Microbiota Composition in Early Life
5.3. Impact of Antibiotics on the Developing Infant Resistome
6. Impact of Increased Levels of ARGs on Health and Mitigation Strategies
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Milani, C.; Duranti, S.; Bottacini, F.; Casey, E.; Turroni, F.; Mahony, J.; Belzer, C.; Delgado Palacio, S.; Arboleya Montes, S.; Mancabelli, L.; et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol. Mol. Biol. Rev. 2017, 81, e00036-17. [Google Scholar] [CrossRef]
- Martin, R.; Nauta, A.; Ben Amor, K.; Knippels, L.; Knol, J.; Garssen, J. Early life: Gut microbiota and immune development in infancy. Benef. Microbes. 2010, 1, 367–382. [Google Scholar] [CrossRef]
- Lebeaux, R.M.; Karalis, D.B.; Lee, J.; Whitehouse, H.C.; Madan, J.C.; Karagas, M.R.; Hoen, A.G. The association between early life antibiotic exposure and the gut resistome of young children: A systematic review. Gut Microbes 2022, 14, 2120743. [Google Scholar] [CrossRef]
- Kounnavong, S.; Yan, W.; Sihavong, A.; Sychareun, V.; Eriksen, J.; Hanson, C.; Chaleunvong, K.; Keohavong, B.; Vongsouvath, M.; Mayxay, M.; et al. Antibiotic knowledge, attitudes and reported practice during pregnancy and six months after birth: A follow- up study in Lao PDR. BMC Pregnancy Childbirth 2022, 22, 701. [Google Scholar] [CrossRef]
- Cantey, J.B.; Wozniak, P.S.; Sanchez, P.J. Prospective surveillance of antibiotic use in the neonatal intensive care unit: Results from the SCOUT study. Pediatr. Infect. Dis. J. 2015, 34, 267–272. [Google Scholar] [CrossRef]
- Dekker, A.R.J.; Verheij, T.J.M.; van der Velden, A.W. Antibiotic management of children with infectious diseases in Dutch primary care. Fam. Pract. 2017, 34, 169–174. [Google Scholar] [CrossRef]
- Taine, M.; Offredo, L.; Dray-Spira, R.; Weill, A.; Chalumeau, M.; Zureik, M. Paediatric outpatient prescriptions in France between 2010 and 2019: A nationwide population-based study: Paediatric outpatient prescriptions in France, 2010 to 2019. Lancet Reg Health Eur. 2021, 7, 100129. [Google Scholar] [CrossRef]
- Baron, R.; Taye, M.; der Vaart, I.B.; Ujcic-Voortman, J.; Szajewska, H.; Seidell, J.C.; Verhoeff, A. The relationship of prenatal antibiotic exposure and infant antibiotic administration with childhood allergies: A systematic review. BMC Pediatr. 2020, 20, 312. [Google Scholar] [CrossRef]
- Li, G.; Bielicki, J.A.; Ahmed, A.S.M.N.U.; Islam, M.S.; Berezin, E.N.; Gallacci, C.B.; Guinsburg, R.; da Silva Figueiredo, C.E.; Santarone Vieira, R.; Silva, A.R.; et al. Towards understanding global patterns of antimicrobial use and resistance in neonatal sepsis: Insights from the NeoAMR network. Arch. Dis. Child. 2020, 105, 26–31. [Google Scholar] [CrossRef]
- WHO. 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 1 June 2023).
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Antimicrobial Resistance in the EU/EEA: A One Health Response. OECD 2022. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/antimicrobial-resistance-policy-brief-2022.pdf (accessed on 1 June 2023).
- Roope, L.S.J.; Smith, R.D.; Pouwels, K.B.; Buchanan, J.; Abel, L.; Eibich, P.; Butler, C.C.; Tan, P.S.; Walker, A.S.; Robotham, J.V.; et al. The challenge of antimicrobial resistance: What economics can contribute. Science 2019, 364, eaau4679. [Google Scholar] [CrossRef]
- Bartoloni, A.; Pallecchi, L.; Rodríguez, H.; Fernandez, C.; Mantella, A.; Bartalesi, F.; Strohmeyer, M.; Kristiansson, C.; Gotuzzo, E.; Paradisi, F.; et al. Antibiotic resistance in a very remote Amazonas community. Int. J. Antimicrob. Agents 2009, 33, 125–129. [Google Scholar] [CrossRef]
- Hollis, A.; Ahmed, Z. The path of least resistance: Paying for antibiotics in non-human uses. Health Policy 2014, 118, 264–270. [Google Scholar] [CrossRef]
- Browne, A.J.; Chipeta, M.G.; Haines-Woodhouse, G.; Kumaran, E.P.A.; Hamadani, B.H.K.; Zaraa, S.; Henry, N.J.; Deshpande, A.; Reiner, R.C., Jr.; Day, N.P.J.; et al. Global antibiotic consumption and usage in humans, 2000–2018: A spatial modelling study. Lancet Plan Health 2021, 5, e893–e904. [Google Scholar] [CrossRef]
- Rahman, S.; Kesselheim, A.S.; Hollis, A. Persistence of resistance: A panel data analysis of the effect of antibiotic usage on the prevalence of resistance. J. Antibiot. 2023, 76, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Cha, C.J. Antibiotic resistome from the One-Health perspective: Understanding and controlling antimicrobial resistance transmission. Exp. Mol. Med. 2021, 53, 301–309. [Google Scholar] [CrossRef]
- Jorgensen, J.H. Laboratory issues in the detection and reporting of antibacterial resistance. Infect. Dis. Clin. N. Am. 1997, 11, 785. [Google Scholar] [CrossRef]
- Lu, N.; Hu, Y.; Zhu, L.; Yang, X.; Yin, Y.; Lei, F.; Zhu, Y.; Du, Q.; Wang, X.; Meng, Z.; et al. DNA microarray analysis reveals that antibiotic resistance-gene diversity in human gut microbiota is age related. Sci. Rep. 2014, 4, 4302. [Google Scholar] [CrossRef]
- Whon, T.W.; Shin, N.R.; Kim, J.Y.; Roh, S.W. Omics in gut microbiome analysis. J. Microbiol. 2021, 59, 292. [Google Scholar] [CrossRef] [PubMed]
- Finegold, S.M. Interaction of antimicrobial therapy and intestinal flora. Am. J. Clin. Nutr. 1970, 23, 1466. [Google Scholar] [CrossRef]
- Archer, G.L.; Penell, E. Detection of methicillin resistance in Staphylococci by using a DNA probe. Antimicrob. Agents Chemother. 1990, 34, 1720. [Google Scholar] [CrossRef]
- Gosalbes, M.J.; Vallès, Y.; Jiménez-Hernández, N.; Balle, C.; Riva, P.; Miravet-Verde, S.; de Vries, L.E.; Llop, S.; Agersø, Y.; Sørensen, S.J.; et al. High frequencies of antibiotic resistance genes in infants’ meconium and early fecal samples. J. Dev. Orig. Health Dis. 2016, 7, 35. [Google Scholar] [CrossRef]
- Klassert, T.E.; Zubiria-Barrera, C.; Kankel, S.; Stock, M.; Neubert, R.; Lorenzo-Diaz, F.; Doehring, N.; Driesch, D.; Fischer, D.; Slevogt, H. Early bacterial colonization and antibiotic resistance gene acquisition in newborns. Front. Cell. Infect. Microbiol. 2020, 10, 332. [Google Scholar] [CrossRef]
- Lai, F.Y.; Muziasari, W.; Virta, M.; Wiberg, K.; Ahrens, L. Profiles of environmental antibiotic resistomes in the urban aquatic recipients of Sweden using high-throughput quantitative PCR analysis. Environ. Pol. 2021, 287, 117651. [Google Scholar] [CrossRef]
- Pehrsson, E.C.; Forsberg, K.J.; Gibson, M.K.; Ahmadi, S.; Dantas, G. Novel resistance functions uncovered using functional metagenomic investigations of resistance reservoirs. Front. Microbiol. 2013, 4, 145. [Google Scholar] [CrossRef]
- Kim, D.W.; Thawng, C.N.; Choi, J.H.; Lee, K.; Cha, C.J. Polymorphism of antibiotic-inactivating enzyme driven by ecology expands the environmental resistome. ISME J. 2018, 12, 267. [Google Scholar] [CrossRef]
- Böhm, M.E.; Razavi, M.; Marathe, N.P.; Flach, C.F.; Larsson, D.G.J. Discovery of a novel integron-borne aminoglycoside resistance gene present in clinical pathogens by screening environmental bacterial communities. Microbiome 2020, 8, 41. [Google Scholar] [CrossRef]
- Ma, L.; Xia, Y.; Li, B.; Yang, Y.; Li, L.G.; Tiedje, J.M.; Zhang, G. Metagenomic assembly reveals hosts of antibiotic resistance genes and the shared resistome in pig, chicken, and human feces. Environ. Sci. Technol. 2016, 50, 420. [Google Scholar] [CrossRef]
- Suzuki, S.; Horinouchi, T.; Furusawa, C. Prediction of antibiotic resistance by gene expression profiles. Nat. Commun. 2014, 5, 5792. [Google Scholar] [CrossRef]
- Felden, B.; Cattoir, V. Bacterial adaptation to antibiotics through regulatory RNAs. Antimicrob. Agents Chemother. 2018, 62, e02503. [Google Scholar] [CrossRef]
- Le Neindre, K.; Dejoies, L.; Reissier, S.; Guérin, F.; Felden, B.; Cattoir, V. Small RNA-mediated regulation of the tet(M) resistance gene expression in Enterococcus faecium. Res. Microbiol. 2022, 173, 103941. [Google Scholar] [CrossRef] [PubMed]
- Boolchandani, M.; D’Souza, A.W.; Dantas, G. Sequencing-based methods and resources to study antimicrobial resistance. Nat. Rev. Genet. 2019, 20, 356. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.J.; Boisvert, S.; Brettin, T.; Kenyon, R.W.; Mao, C.; Olson, R.; Overbeek, R.; Santerre, J.; Shukla, M.; Wattam, A.R.; et al. Antimicrobial resistance prediction in PATRIC and RAST. Sci. Rep. 2016, 6, 27930. [Google Scholar] [CrossRef]
- Arango-Argoty, G.A.; Dai, D.; Pruden, A.; Vikesland, P.; Heath, L.S.; Zhang, L. NanoARG: A web service for detecting and contextualizing antimicrobial resistance genes from nanopore-derived metagenomes. Microbiome 2019, 7, 88. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Chung, J.; Battaglia, T.; Henderson, N.; Jay, M.; Li, H.; Lieber, A.; Wu, F.; Perez-Perez, G.I.; Chen, Y.; et al. Antibiotics, birth mode, and diet shape microbiome maturation during early life. Sci. Transl. Med. 2016, 8, 343ra82. [Google Scholar] [CrossRef]
- Penders, J.; Stobberingh, E.E.; Savelkoul, P.H.; Wolffs, P.F. The human microbiome as a reservoir of antimicrobial resistance. Front. Microbiol. 2013, 4, 87. [Google Scholar] [CrossRef]
- Leo, S.; Curtis, N.; Zimmermann, P. The neonatal intestinal resistome and factors that influence it-a systematic review. Clin. Microbiol. Infect. 2022, 28, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Thänert, R.; Sawhney, S.S.; Schwartz, D.J.; Dantas, G. The resistance within: Antibiotic disruption of the gut microbiome and resistome dynamics in infancy. Cell Host Microbe 2022, 30, 675–683. [Google Scholar] [CrossRef]
- Li, X.; Stokholm, J.; Brejnrod, A.; Vestergaard, G.A.; Russel, J.; Trivedi, U.; Thorsen, J.; Gupta, S.; Hjelmsø, M.H.; Shah, S.A.; et al. The infant gut resistome associates with E. coli, environmental exposures, gut microbiome maturity, and asthma-associated bacterial composition. Cell Host Microbe 2021, 29, 975–987. [Google Scholar] [CrossRef]
- Reyman, M.; van Houten, M.A.; van Baarle, D.; Bosch, A.A.T.M.; Man, W.H.; Chu, M.L.J.N.; Arp, K.; Watson, R.L.; Sanders, E.A.M.; Fuentes, S.; et al. Impact of delivery mode-associated gut microbiota dynamics on health in the first year of life. Nat. Commun. 2019, 10, 4997. [Google Scholar] [CrossRef]
- Li, W.; Tapiainen, T.; Brinkac, L.; Lorenzi, H.A.; Moncera, K.; Tejesvi, M.V.; Salo, J.; Nelson, K.E. Vertical transmisión of gut microbiome and antimicrobial resistance genes in infants exposed to antibiotics at birth. J. Infect. Dis. 2021, 224, 1236–1246. [Google Scholar] [CrossRef]
- Gasparrini, A.J.; Wang, B.; Sun, X.; Kennedy, E.A.; Hernandez-Leyva, A.; Ndao, I.M.; Tarr, P.I.; Warner, B.B.; Dantas, G.l. Persistent metagenomic signatures of early-life hospitalization and antibiotic treatment in the infant gut microbiota and resistome. Nat. Microbiol. 2019, 4, 2285–2297. [Google Scholar] [CrossRef]
- Pärnänen, K.; Karkman, A.; Hultman, J.; Lyra, C.; Bengtsson-Palme, J.; Larsson, D.G.J.; Rautava, S.; Isolauri, E.; Salminen, S.; Kumar, H.; et al. Maternal gut and breast milk microbiota affect infant gut antibiotic resistome and mobile genetic elements. Nat. Commun. 2018, 9, 3891. [Google Scholar] [CrossRef]
- Socha-Banasiak, A.; Pawłowska, M.; Czkwianianc, E.; Pierzynowska, K. From Intrauterine to Extrauterine Life-The Role of Endogenous and Exogenous Factors in the Regulation of the Intestinal Microbiota Community and Gut Maturation in Early Life. Front. Nutr. 2021, 8, 696966. [Google Scholar] [CrossRef]
- Gibson, M.K.; Wang, B.; Ahmadi, S.; Burnham, C.A.; Tarr, P.I.; Warner, B.B.; Dantas, G. Developmental dynamics of the preterm infant gut microbiota and antibiotic resistome. Nat. Microbiol. 2016, 1, 16024. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, L.; Chen, H.; Zhang, S.; Zhuang, J.; Li, Q.; Feng, Z. Intestinal Microbiota in Early Life and Its Implications on Childhood Health. Genom. Proteom. Bioinform. 2019, 17, 13–25. [Google Scholar] [CrossRef]
- Hill, C.J.; Lynch, D.B.; Murphy, K.; Ulaszewska, M.; Jeffery, I.B.; O'Shea, C.A.; Watkins, C.; Dempsey, E.; Mattivi, F.; Tuohy, K.; et al. Evolution of gut microbiota composition from birth to 24 weeks in the INFANTMET Cohort. Microbiome 2017, 5, 4. [Google Scholar] [CrossRef] [PubMed]
- Mesa, M.D.; Loureiro, B.; Iglesia, I.; Fernandez Gonzalez, S.; Llurba Olivé, E.; García Algar, O.; Solana, M.J.; Cabero Perez, M.J.; Sainz, T.; Martinez, L.; et al. The Evolving Microbiome from Pregnancy to Early Infancy: A Comprehensive Review. Nutrients 2020, 12, 133. [Google Scholar] [CrossRef] [PubMed]
- Neuman, H.; Koren, O. The Pregnancy Microbiome. Nestlé Nutr. Inst. Work. Ser. 2017, 88, 1–9. [Google Scholar] [CrossRef]
- Khan, I.; Yasir, M.; Farman, M.; Kumosani, T.; AlBasri, S.F.; Bajouh, O.S.; Azhar, E.I. Evaluation of gut bacterial community composition and antimicrobial resistome in pregnant and non-pregnant women from Saudi population. Infect. Drug Resist. 2019, 12, 1749. [Google Scholar] [CrossRef]
- Sosa-Moreno, A.; Comstock, S.S.; Sugino, K.Y.; Ma, T.F.; Paneth, N.; Davis, Y.; Olivero, R.; Schein, R.; Maurer, J.; Zhang, L. Perinatal risk factors for fecal antibiotic resistance gene patterns in pregnant women and their infants. PLoS ONE 2020, 15, e0234751. [Google Scholar] [CrossRef]
- Gupta, P.; Singh, M.P.; Goyal, K. Diversity of Vaginal Microbiome in Pregnancy: Deciphering the Obscurity. Front. Public Health 2020, 8, 326. [Google Scholar] [CrossRef]
- Severgnini, M.; Camboni, T.; Ceccarani, C.; Morselli, S.; Cantiani, A.; Zagonari, S.; Patuelli, G.; Pedna, M.F.; Sambri, V.; Foschi, C.; et al. Distribution of ermb, ermf, tet(W), and tet(m) resistance genes in the vaginal ecosystem of women during pregnancy and puerperium. Pathogens 2021, 10, 1546. [Google Scholar] [CrossRef]
- Maestre-Carballa, L.; Navarro-López, V.; Martinez-Garcia, M.A. Resistome Roadmap: From the Human Body to Pristine Environments. Front. Microbiol. 2022, 13, 858831. [Google Scholar] [CrossRef]
- Roachford, O.S.E.; Alleyne, A.T.; Kuelbs, C.; Torralba, M.G.; Nelson, K.E. The cervicovaginal microbiome and its resistome in a random selection of Afro-Caribbean women. Human. Microb. J. 2021, 20, 100079. [Google Scholar] [CrossRef]
- Perera, M.; Al-Hebshi, N.N.; Speicher, D.J.; Perera, I.; Johnson, N.W. Emerging role of bacteria in oral carcinogenesis: A review with special reference to perio-pathogenic bacteria. J. Oral. Microbiol. 2016, 8, 32762. [Google Scholar] [CrossRef] [PubMed]
- Borgo, P.V.; Rodrigues, V.A.A.; Feitosa, A.C.R.; Xavier, K.C.B.; Avila-Campos, M.J. Association between periodontal condition and subgingival microbiota in women during pregnancy: A longitudinal study. J. Appl. Oral. Sci. 2014, 22, 528. [Google Scholar] [CrossRef]
- Fujiwara, N.; Tsuruda, K.; Iwamoto, Y.; Kato, F.; Odaki, T.; Yamane, N.; Hori, Y.; Harashima, Y.; Sakoda, A.; Tagaya, A.; et al. Significant increase of oral bacteria in the early pregnancy period in Japanese women. J. Investig. Clin. Dent. 2017, 8, e12189. [Google Scholar] [CrossRef]
- Gohir, W.; Kennedy, K.M.; Wallace, J.G.; Saoi, M.; Bellissimo, C.J.; Britz-McKibbin, P.; Petrik, J.J.; Surette, M.G.; Sloboda, D.M. High-fat diet intake modulates maternal intestinal adaptations to pregnancy and results in placental hypoxia, as well as altered fetal gut barrier proteins and immune markers. J. Physiol. 2019, 597, 3029–3051. [Google Scholar] [CrossRef]
- Serrano, M.G.; Parikh, H.I.; Brooks, J.P.; Edwards, D.J.; Arodz, T.J.; Edupuganti, L.; Huang, B.; Girerd, P.H.; Bokhari, Y.A.; Bradley, S.P.-; et al. Racioethnic diversity in the dynamics of the vaginal microbiome during pregnancy. Nat. Med. 2019, 25, 1001–1011. [Google Scholar] [CrossRef]
- Carvalho, M.J.; Sands, K.; Thomson, K.; Portal, E.; Mathias, J.; Milton, R.; Gillespie, D.; Dyer, C.; Akpulu, C.; Boostrom, I.; et al. Antibiotic resistance genes in the gut microbiota of mothers and linked neonates with or without sepsis from low- and middle-income countries. Nat. Microbiol. 2022, 7, 1337–1347. [Google Scholar] [CrossRef] [PubMed]
- Yassour, M.; Jason, E.; Hogstrom, L.J.; Arthur, T.D.; Tripathi, S.; Siljander, H.; Selvenius, J.; Oikarinen, S.; Hyöty, H.; Virtanen, S.M.; et al. Strain-level analysis of mother-to-child bacterial transmission during the first few months of life. Cell Host Microbe 2018, 24, 146. [Google Scholar] [CrossRef] [PubMed]
- Kozak, K.; Charbonneau, D.; Sanozky-Dawes, R.; Klaenhammer, T. Characterization of bacterial isolates from the microbiota of mothers’ breast milk and their infants. Gut Microbes 2015, 6, 341. [Google Scholar] [CrossRef]
- Patangia, D.V.; Ryan, C.A.; Dempsey, E.; Stanton, C.; Ross, R.P. Vertical transfer of antibiotics and antibiotic resistant strains across the mother/baby axis. Trends Microbiol. 2022, 30, 47–56. [Google Scholar] [CrossRef]
- Dubois, V.; De Barbeyrac, B.; Rogues, A.M.; Arpin, C.; Coulange, L.; Andre, C.; M'zali, F.; Megraud, F.; Quentin, C. CTX-M-producing Escherichia coli in a maternity ward: A likely community importation and evidence of mother-to-neonate transmission. J. Antimicrob. Chemother. 2010, 65, 1368–1371. [Google Scholar] [CrossRef]
- Alicea-Serrano, A.M.; Contreras, M.; Magris, M.; Hidalgo, G.; Dominguez-Bello, M.G. Tetracycline resistance genes acquired at birth. Arch. Microbiol. 2013, 195, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Karami, N.; Nowrouzian, F.; Adlerberth, I.; Wold, A.E. Tetracycline resistance in Escherichia coli and persistence in the infantile colonic microbiota. Antimicrob. Agents Chemother. 2006, 50, 156–161. [Google Scholar] [CrossRef]
- Das, L.; Virmani, R.; Sharma, V.; Rawat, D.; Singh, Y. Human Milk Microbiota: Transferring the Antibiotic Resistome to Infants. Ind. J. Microbiol. 2019, 59, 410–416. [Google Scholar] [CrossRef]
- Huang, M.S.; Cheng, C.C.; Tseng, S.Y.; Lin, Y.L.; min Lo, H.; Chen, P.W. Most commensally bacterial strains in human milk of healthy mothers display multiple antibiotic resistance. Microbiologyopen 2019, 8, e00618. [Google Scholar] [CrossRef]
- Nadimpalli, M.L.; Bourke, C.D.; Robertson, R.C.; Delarocque-Astagneau, E.; Manges, A.R.; Pickering, A.J. Can breastfeeding protect against antimicrobial resistance? BMC Med. 2020, 18, 392. [Google Scholar] [CrossRef]
- Li, X.; Zhou, Y.; Zhan, X.; Huang, W.; Wang, X. Breast milk is a potential reservoir for livestock-associated Staphylococcus aureus and community-associated Staphylococcus aureus in Shanghai, China. Front. Microbiol. 2018, 8, 2639. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.W.; Tseng, S.Y.; Huang, M.S. Antibiotic Susceptibility of Commensal Bacteria from Human Milk. Curr. Microbiol. 2016, 72, 113–119. [Google Scholar] [CrossRef] [PubMed]
- White, N.D. Drug-Induced Microbiome Changes: Considerations in Pregnancy. Am. J. Lifestyle Med. 2022, 17, 50–53. [Google Scholar] [CrossRef]
- Brigadoi, G.; Rossin, S.; Visentin, D.; Barbieri, E.; Giaquinto, C.; Da Dalt, L.; Donà, D. The impact of Antimicrobial Stewardship Programmes in paediatric emergency departments and primary care: A systematic review. Ther. Adv. Infect. Dis. 2023, 10. [Google Scholar] [CrossRef]
- Calle-Miguel, L.; Pérez-Méndez, C.; García-García, E.; Moreno-Pavón, B.; Solís-Sánchez, G. Trends and Pattern of Antibiotic Use in Children in Northern Spain, Interpreting Data about Antibiotic Consumption in Pediatric Outpatients. Children 2022, 9, 442. [Google Scholar] [CrossRef] [PubMed]
- Poole, N.M.; Shapiro, D.J.; Fleming-Dutra, K.E.; Hicks, L.A.; Hersh, A.L.; Kronman, M.P. Antibiotic Prescribing for Children in United States Emergency Departments: 2009–2014. Pediatrics 2019, 143, e20181056. [Google Scholar] [CrossRef]
- García-Moreno, F.J.; Escobar-Castellanos, M.; Marañón, R.; Rivas-García, A.; Manrique-Rodríguez, S.; Mora-Capín, A.; Fernández-Llamazares, C.M. Adequacy of pediatric antimicrobial prescribing in the Emergency Department at discharge. An. Pediatr. (Engl. Ed.) 2022, 96, 179–189. [Google Scholar] [CrossRef]
- WHO. Collaborating Center for Drug Statistics Methodology. In Guidelines for ATC Classification and DDD Assignment; Norwegian Institute of Public Health: Oslo, Norway, 2022. [Google Scholar]
- Same, R.G. The Current State and Future Directions of Inpatient Pediatric Antimicrobial Stewardship. Infect. Dis. Clin. N. Am. 2022, 36, 173–186. [Google Scholar] [CrossRef]
- Glaser, M.A.; Hughes, L.M.; Jnah, A.; Newberry, D. Neonatal Sepsis: A Review of Pathophysiology and Current Management Strategies. Adv. Neonatal Care 2021, 21, 49–60. [Google Scholar] [CrossRef]
- Fleiss, N.; Schwabenbauer, K.; Randis, T.M.; Polin, R.A. What's new in the management of neonatal early-onset sepsis? Arch. Dis. Child. Fetal Neonatal Ed. 2023, 108, 10–14. [Google Scholar] [CrossRef]
- Cailes, B.; Kortsalioudaki, C.; Buttery, J.; Pattnayak, S.; Greenough, A.; Matthes, J.; Bedford Russell, A.; Kennea, N.; Heath, P.T.; neonIN network. Epidemiology of UK neonatal infections: The neonIN infection surveillance network. Arch. Dis. Child. Fetal Neonatal Ed. 2018, 103, F547–F553. [Google Scholar] [CrossRef] [PubMed]
- Paul, S.P.; Khattak, H.; Kini, P.K.; Heaton, P.A.; Goel, N. NICE guideline review: Neonatal infection: Antibiotics for prevention and treatment (NG195). Arch. Dis. Child. Educ. Pract. Ed. 2022, 107, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Winzor, G.; Atabani, S.F. How and when to use CSF to investigate neonates and children with possible central nervous system infection. Arch. Dis. Child. Educ. Pract. Ed. 2022, 107, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Siddiq, S.; Grainger, J. The diagnosis and management of acute otitis media: American Academy of Pediatrics Guidelines 2013. Arch. Dis. Child. Educ. Pract. Ed. 2015, 100, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.N.; Marikar, D. Antibiotic prescribing for upper respiratory tract infections: NICE guidelines. Arch. Dis. Child. Educ. Pract. Ed. 2020, 105, 104–106. [Google Scholar] [CrossRef]
- Autore, G.; Bernardi, L.; La Scola, C.; Ghidini, F.; Marchetti, F.; Pasini, A.; Pierantoni, L.; Castellini, C.; Gatti, C.; Malaventura, C.; et al. The Uti-Ped-Er Study Group. Management of Pediatric Urinary Tract Infections: A Delphi Study. Antibiotics 2022, 11, 1122. [Google Scholar] [CrossRef]
- Yi, J.; Wood, J.B.; Creech, C.B.; Williams, D.; Jimenez-Truque, N.; Yildirim, I.; Sederdahl, B.; Daugherty, M.; Hussaini, L.; Munye, M.; et al. Clinical Epidemiology and Outcomes of Pediatric Musculoskeletal Infections. J. Pediatr. 2021, 234, 236–244. [Google Scholar] [CrossRef]
- Posadas, E.; Fisher, J. Pediatric bacterial meningitis: An update on early identification and management. Pediatr. Emer. Med. Pract. 2018, 15, 1–20. [Google Scholar]
- Zimmermann, P.; Curtis, N. The effect of antibiotics on the composition of the intestinal microbiota—A systematic review. J. Infect. 2019, 79, 471–489. [Google Scholar] [CrossRef]
- Fjalstad, J.W.; Esaiassen, E.; Juvet, L.K.; van den Anker, J.N.; Klingenberg, C. Antibiotic therapy in neonates and impact on gut microbiota and antibiotic resistance development: A systematic review. J. Antimicrob. Chemother. 2018, 73, 569–580. [Google Scholar] [CrossRef]
- Yassour, M.; Vatanen, T.; Siljander, H.; Hämäläinen, A.M.; Härkönen, T.; Ryhänen, S.J.; Franzosa, E.A.; Vlamakis, H.; Huttenhower, C.; Gevers, D.; et al. Natural history of the infant gut microbiome and impact of antibiotic treatment on bacterial strain diversity and stability. Sci. Transl. Med. 2016, 8, 343ra81. [Google Scholar] [CrossRef] [PubMed]
- Fouhy, F.; Guinane, C.M.; Hussey, S.; Wall, R.; Ryan, C.A.; Dempsey, E.M.; Murphy, B.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C.; et al. High-throughput sequencing reveals the incomplete, short-term recovery of infant gut microbiota following parenteral antibiotic treatment with ampicillin and gentamicin. Antimicrob. Agents Chemother. 2012, 56, 5811–5820. [Google Scholar] [CrossRef] [PubMed]
- Uzan-Yulzari, A.; Turta, O.; Belogolovski, A.; Ziv, O.; Kunz, C.; Perschbacher, S.; Neuman, H.; Pasolli, E.; Oz, A.; Ben-Amram, H.; et al. Neonatal antibiotic exposure impairs child growth during the first six years of life by perturbing intestinal microbial colonization. Nat. Commun. 2021, 12, 443. [Google Scholar] [CrossRef] [PubMed]
- Reyman, M.; van Houten, M.A.; Watson, R.L.; Chu, M.L.J.N.; Arp, K.; de Waal, W.J.; Schiering, I.; Plötz, F.B.; Willems, R.J.L.; van Schaik, W.; et al. Effects of early-life antibiotics on the developing infant gut microbiome and resistome: A randomized trial. Nat. Commun. 2022, 13, 893. [Google Scholar] [CrossRef]
- Gibson, M.K.; Crofts, T.S.; Dantas, G. Antibiotics and the developing infant gut microbiota and resistome. Curr. Opin. Microbiol. 2015, 27, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Zwittink, R.D.; Renes, I.B.; van Lingen, R.A.; van Zoeren-Grobben, D.; Konstanti, P.; Norbruis, O.F.; Martin, R.; Groot Jebbink, L.J.M.; Knol, J.; Belzer, C. Association between duration of intravenous antibiotic administration and early-life microbiota development in late-preterm infants. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Kuppala, V.S.; Meinzen-Derr, J.; Morrow, A.L.; Schibler, K.R. Prolonged initial empirical antibiotic treatment is associated with adverse outcomes in premature infants. J. Pediatr. 2011, 159, 720–725. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, C.; Morrow, A.L.; Lagomarcino, A.J.; Altaye, M.; Taft, D.H.; Yu, Z.; Newburg, D.S.; Ward, D.V.; Schibler, K.R. Early empiric antibiotic use in preterm infants is associated with lower bacterial diversity and higher relative abundance of Enterobacter. J. Pediatr. 2014, 165, 23–29. [Google Scholar] [CrossRef]
- Rooney, A.M.; Timberlake, K.; Brown, K.A.; Bansal, S.; Tomlinson, C.; Lee, K.S.; Science, M.; Coburn, B. Each Additional Day of Antibiotics Is Associated with Lower Gut Anaerobes in Neonatal Intensive Care Unit Patients. Clin. Infect. Dis. 2020, 70, 2553–2560. [Google Scholar] [CrossRef]
- Schwartz, D.J.; Langdon, A.E.; Dantas, G. Understanding the impact of antibiotic perturbation on the human microbiome. Genome Med. 2020, 12, 82. [Google Scholar] [CrossRef] [PubMed]
- Duong, Q.A.; Pittet, L.F.; Curtis, N.; Zimmermann, P. Antibiotic exposure and adverse long-term health outcomes in children: A systematic review and meta-analysis. J. Infect. 2022, 85, 213–300. [Google Scholar] [CrossRef]
- Saari, A.; Virta, L.J.; Sankilampi, U.; Dunkel, L.; Saxen, H. Antibiotic exposure in infancy and risk of being overweight in the first 24 months of life. Pediatrics 2015, 135, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Cox, L.M.; Yamanishi, S.; Sohn, J.; Alekseyenko, A.V.; Leung, J.M.; Cho, I.; Kim, S.G.; Li, H.; Gao, Z.; Mahana, D.; et al. Altering the intestinal microbiota during a critical developmental window has lasting metabolic consequences. Cell 2014, 158, 705–721. [Google Scholar] [CrossRef] [PubMed]
- Samarra, A.; Esteban-Torres, M.; Cabrera-Rubio, R.; Bernabeu, M.; Arboleya, S.; Gueimonde, M.; Collado, M.C. Maternal-infant antibiotic resistance genes transference: What do we know? Gut Microbes 2023, 15, 2194797. [Google Scholar] [CrossRef] [PubMed]
- Gueimonde, M.; Salminen, S.; Isolauri, E. Presence of specific antibiotic (tet) resistance genes in infant faecal microbiota. FEMS Immunol. Med. Microbiol. 2006, 48, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Stam, J.; van Stuijvenberg, M.; Grüber, C.; Mosca, F.; Arslanoglu, S.; Chirico, G.; Braegger, C.P.; Riedler, J.; Boehm, G.; Sauer, P.J.; et al. Antibiotic use in infants in the first year of life in five European countries. Acta Paediatr. 2012, 101, 929–934. [Google Scholar] [CrossRef]
- Loo, E.X.L.; Zain, A.; Yap, G.C.; Purbojati, R.W.; Drautz-Moses, D.I.; Koh, Y.Q.; Chong, Y.S.; Tan, K.H.; Gluckman, P.D.; Yap, F.; et al. Longitudinal assessment of antibiotic resistance gene profiles in gut microbiomes of infants at risk of eczema. BMC Infect. Dis. 2020, 20, 312. [Google Scholar] [CrossRef]
- Nogacka, A.M.; Salazar, N.; Arboleya, S.; Suárez, M.; Fernández, N.; Solís, G.; de Los Reyes-Gavilán, C.G.; Gueimonde, M. Early microbiota, antibiotics and health. Cell. Mol. Life Sci. 2018, 75, 83–91. [Google Scholar] [CrossRef]
- Raabe, V.N.; Shane, A.L. Group B Streptococcus (Streptococcus agalactiae). Microbiol. Spect. 2019, 7, 17. [Google Scholar] [CrossRef]
- Arboleya, S.; Sánchez, B.; Milani, C.; Duranti, S.; Solís, G.; Fernández, N.; de los Reyes-Gavilán, C.G.; Ventura, M.; Margolles, A.; Gueimonde, M. Intestinal microbiota development in preterm neonates and effect of perinatal antibiotics. J. Pediatr. 2015, 166, 538–544. [Google Scholar] [CrossRef]
- Nogacka, A.; Salazar, N.; Suárez, M.; Milani, C.; Arboleya, S.; Solís, G.; Fernández, N.; Alaez, L.; Hernández-Barranco, A.M.; de Los Reyes-Gavilán, C.G.; et al. Impact of intrapartum antimicrobial prophylaxis upon the intestinal microbiota and the prevalence of antibiotic resistance genes in vaginally delivered full-term neonates. Microbiome 2017, 5, 93. [Google Scholar] [CrossRef] [PubMed]
- Arboleya, S.; Saturio, S.; Gueimonde, M. Impact of intrapartum antibiotics on the developing microbiota: A review. Microb. Res. Rep. 2022, 1, 22. [Google Scholar] [CrossRef]
- Gilstrap, L.C.; Ramin, S.M. Urinary tract infections during pregnancy. Obstet. Gynecol. Clin. N. Am. 2001, 28, 581–591. [Google Scholar] [CrossRef]
- Lebeaux, R.M.; Madan, J.C.; Nguyen, Q.P.; Coker, M.O.; Dade, E.F.; Moroishi, Y.; Palys, T.J.; Ross, B.D.; Pettigrew, M.M.; Morrison, H.G.; et al. Impact of antibiotics on off-target infant gut microbiota and resistance genes in cohort studies. Pediatr. Res. 2022, 92, 1757–1766. [Google Scholar] [CrossRef] [PubMed]
- Crits-Christoph, A.; Hallowell, H.A.; Koutouvalis, K.; Suez, J. Good microbes, bad genes? The dissemination of antimicrobial resistance in the human microbiome. Gut Microbes 2022, 14, 2055944. [Google Scholar] [CrossRef]
- Stecher, B.; Denzler, R.; Maier, L.; Bernet, F.; Sanders, M.J.; Pickard, D.J.; Barthel, M.; Westendorf, A.M.; Krogfelt, K.A.; Walker, A.W.; et al. Gut inflammation can boost horizontal gene transfer between pathogenic and commensal Enterobacteriaceae. Proc. Natl. Acad. Sci. USA 2012, 109, 1269–1274. [Google Scholar] [CrossRef] [PubMed]
- Goren, M.G.; Carmeli, Y.; Schwaber, M.J.; Chmelnitsky, I.; Schechner, V.; Navon-Venezia, S. Transfer of carbapenem-resistant plasmid from Klebsiella pneumoniae ST258 to Escherichia coli in patient. Emerg. Infect. Dis. 2010, 16, 1014–1017. [Google Scholar] [CrossRef]
- ARTITUS 2013. Antibiotic Resistance Threats in the United States. 2013. Available online: https://www.cdc.gov/drugresistance/pdf/ar-threats-2013-508.pdf (accessed on 1 June 2023).
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; The Review on Antimicrobial Resistance; Government of the United Kingdom: London, UK, 2016. [Google Scholar]
- Cifuentes, S.G.; Graham, J.; Loayza, F.; Saraiva, C.; Salinas, L.; Trueba, G.; Cárdenas, P.A. Evaluation of changes in the faecal resistome associated with children’s exposure to domestic animals and food animal production. J. Glob. Antimicrob. Resist. 2022, 31, 212–215. [Google Scholar] [CrossRef]
- Wu, G.; Zhang, C.; Wang, J.; Zhang, F.; Wang, R.; Shen, J.; Wang, L.; Pang, X.; Zhang, X.; Zhao, L.; et al. Diminution of the gut resistome after a gut microbiota-targeted dietary intervention in obese children. Sci. Rep. 2016, 6, 24030. [Google Scholar] [CrossRef]
- Guitor, A.K.; Yousuf, E.I.; Raphenya, A.R.; Hutton, E.K.; Morrison, K.M.; McArthur, A.G.; Wright, G.D.; Stearns, J.C. Capturing the antibiotic resistome of preterm infants reveals new benefits of probiotic supplementation. Microbiome 2022, 10, 136. [Google Scholar] [CrossRef]
- Bargheet, A.; Klingenberg, C.; Esaiassen, E.; Hjerde, E.; Cavanagh, J.P.; Bengtsson-Palme, J.; Pettersen, V.K. Development of early life gut resistome and mobilome across gestational ages and microbiota-modifying treatments. EBioMedicine 2023, 92, 104613. [Google Scholar] [CrossRef]
- Montassier, E.; Valdés-Mas, R.; Batard, E.; Zmora, N.; Dori-Bachash, M.; Suez, J.; Elinav, E. Probiotics impact the antibiotic resistance gene reservoir along the human GI tract in a person-specific and antibiotic-dependent manner. Nat. Microbiol. 2021, 6, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- Karbalaei, M.; Keikha, M. Probiotics and intestinal decolonization of antibiotic-resistant microorganisms; A reality or fantasy? Ann. Med. Surg. 2022, 80, 104269. [Google Scholar] [CrossRef] [PubMed]
- Merrick, B.; Sergaki, C.; Edwards, L.; Moyes, D.L.; Kertanegara, M.; Prossomariti, D.; Shawcross, D.L.; Goldenberg, S.D. Modulation of the Gut Microbiota to Control Antimicrobial Resistance (AMR)—A Narrative Review with a Focus on Faecal Microbiota Transplantation (FMT). Infect. Dis. Rep. 2023, 15, 238–254. [Google Scholar] [CrossRef]
- Corbellino, M.; Kieffer, N.; Kutateladze, M.; Balarjishvili, N.; Leshkasheli, L.; Askilashvili, L.; Tsertsvadze, G.; Rimoldi, S.G.; Nizharadze, D.; Hoyle, N.; et al. Eradication of a Multidrug-Resistant, Carbapenemase-Producing Klebsiella pneumoniae Isolate Following Oral and Intra-rectal Therapy with a Custom Made, Lytic Bacteriophage Preparation. Clin. Infect. Dis. 2019, 70, 1998–2001. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, S.; Ruth, M.M.; Mientjes, M.; de Sévaux, R.G.L.; van Ingen, J.A. Dutch Case Report of Successful Treatment of Chronic Relapsing Urinary Tract Infection with Bacteriophages in a Renal Transplant Patient. Antimicrob. Agents Chemother. 2019, 64, e01281-19. [Google Scholar] [CrossRef]
- Millan, B.; Park, H.; Hotte, N.; Mathieu, O.; Burguiere, P.; Tompkins, T.A.; Kao, D.; Madsen, K.L. Fecal Microbial Transplants Reduce Antibiotic-resistant Genes in Patients with Recurrent Clostridium difficile Infection. Clin. Infect. Dis. 2016, 62, 1479–1486. [Google Scholar] [CrossRef]
- Bilsen, M.P.; Lambregts, M.M.C.; van Prehn, J.; Kuijper, E.J. Faecal microbiota replacement to eradicate antimicrobial resistant bacteria in the intestinal tract—A systematic review. Curr. Opin. Gastroenterol. 2022, 38, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Merli, P.; Putignani, L.; Ruggeri, A.; Del Chierico, F.; Gargiullo, L.; Galaverna, F.; Gaspari, S.; Pagliara, D.; Russo, A.; Pane, S.; et al. Decolonization of multidrug resistant bacteria by fecal microbiota transplantation in five pediatric patients before allogeneic hematopoietic stem cell transplantation: Gut microbiota profiling, infectious and clinical outcomes. Haematologica 2020, 105, 2686–2690. [Google Scholar] [CrossRef]


{kind=link}
| NEONATAL PERIOD | ||
|---|---|---|
| Most Frequent Bacteria | Usual Antibiotic Treatment | |
| Vertical infections | Streptococcus agalactiae, Listeria monocytogenes, Enterococcus faecalis, Staphylococcus aureus, Escherichia coli | Ampicillin IV plus Gentamicin IV |
| Nosocomial infections | Staphylococcus epidermidis, coagulase negative Staphylococcus, Staphylococcus aureus, Klebsiella pneumoniae, Pseudomonas aeruginosa, Serratia marcescens, Enterobacter spp. | Vancomycin IV plus Amikacin IV |
| Vertical or Nosocomial with meningitis | Any of the previous | Cefotaxime IV plus Gentamicin IV |
| Community infections: | Chlamydia trachomatis, Streptococcus pneumonia, | Oral azithromycin or |
| - Respiratory | Bordetella pertussis | Ampicillin IV |
| - Urinary | E. coli, Enterococcus faecalis | Ampicillin IV plus Gentamicin IV |
| - Soft tissues | Staphylococcus aureus | Cloxacillin IV |
| TODDLER–CHILD PERIOD | ||
| - Otitis/sinusitis - Tonsillopharyngitis | Streptococcus pneumoniae Haemophilus influenzae Streptococcus pyogenes | Amoxicillin Amoxicillin–clavulanate Penicillin |
| - Pneumonia (typical) - Pneumonia (atypical) | Streptococcus pneumoniae Haemophilus influenzae Mycoplasma pneumoniae Bordetella pertussis | Amoxicillin Amoxicillin clavulanic Macrolides: azithromycin or clarithromycin |
| Urinary infections | E. coli, Enterococcus faecalis, Proteus | Amoxicillin–clavulanic, Cefixime, Cefuroxime, Cefotaxime, Ceftriaxone |
| Other infections - Osteomuscular - Meningitis and sepsis | Staphylococcus aureus, Kingella kingae Streptococcus pneumoniae, Neisseria meningitidis, Haemophilus influenzae | Cloxacillin, Cefotaxime Cefotaxime |
| Antibiotic | Doses (mg/kg) | Time | |
|---|---|---|---|
| Neonate | Ampicillin (IV) | 25–50 | Every 6–8–12 h |
| Gentamicin (IV) | 4–5 | Every 24–48 h | |
| Vancomycin (IV) | 10–15 | Every 8–12 h | |
| Teicoplamin (IV) | 10 | Every 12–24 h | |
| Amikacin (IV) | 15–18 | Every 24–48 h | |
| Cefotaxime (IV) | 50–100 | Every 8–12 h | |
| Cloxacillin (IV) | 25–30 | Every 6–12 h | |
| Toddler–Child | Phenoxymethylpenicillin (oral) | 6–12 | Every 6–8 h |
| Benzylpenicillin (IV IM) | 25,000–50,000 * | Every 6 h | |
| Amoxicillin (oral) | 15–25 | Every 8 h | |
| Amoxicilin clavulanic (oral or IV) | 15–25 ** | Every 8 h | |
| Ampicillin (IV) | 25–50 | Every 6 h | |
| Cloxacillin (IV or oral) | 25 | Every 6 h | |
| Cefixime (oral) | 4–8 | Every 12–24 h | |
| Cefuroxime (oral) | 10 | Every 12 h | |
| Cefuroxime (IV) | 33 | Every 8 h | |
| Cefotaxime (IV) | 25–50 | Every 6–8 h | |
| Ceftriaxone (IM or IV) | 50–100 | Every 12–24 h | |
| Gentamicin (IV) | 5 | Every 24 h | |
| Azithromycin (oral) | 10 | Every 24 h | |
| Clarithromycin (oral) | 7.5–15 | Every 12 h |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saturio, S.; Rey, A.; Samarra, A.; Collado, M.C.; Suárez, M.; Mantecón, L.; Solís, G.; Gueimonde, M.; Arboleya, S. Old Folks, Bad Boon: Antimicrobial Resistance in the Infant Gut Microbiome. Microorganisms 2023, 11, 1907. https://doi.org/10.3390/microorganisms11081907
Saturio S, Rey A, Samarra A, Collado MC, Suárez M, Mantecón L, Solís G, Gueimonde M, Arboleya S. Old Folks, Bad Boon: Antimicrobial Resistance in the Infant Gut Microbiome. Microorganisms. 2023; 11(8):1907. https://doi.org/10.3390/microorganisms11081907
Chicago/Turabian StyleSaturio, Silvia, Alejandra Rey, Anna Samarra, Maria Carmen Collado, Marta Suárez, Laura Mantecón, Gonzalo Solís, Miguel Gueimonde, and Silvia Arboleya. 2023. "Old Folks, Bad Boon: Antimicrobial Resistance in the Infant Gut Microbiome" Microorganisms 11, no. 8: 1907. https://doi.org/10.3390/microorganisms11081907
APA StyleSaturio, S., Rey, A., Samarra, A., Collado, M. C., Suárez, M., Mantecón, L., Solís, G., Gueimonde, M., & Arboleya, S. (2023). Old Folks, Bad Boon: Antimicrobial Resistance in the Infant Gut Microbiome. Microorganisms, 11(8), 1907. https://doi.org/10.3390/microorganisms11081907