
Lumpy Skin Disease Virus Infection Activates Autophagy and Endoplasmic Reticulum Stress-Related Cell Apoptosis in Primary Bovine Embryonic Fibroblast Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Virus Infection
2.2. Transmission Electron Microscope (TEM) Observation
2.3. Antibodies
2.4. Western Blotting
2.5. Cell Apoptosis Analysis
2.6. Cellular Indirect Immunofluorescence (IFA)
2.7. Statistical Analysis
3. Results
3.1. LSDV Elicits Ultrastructural Abnormalities of BEF Cells
3.2. LSDV Infection Induces BEF Cell Apoptosis
3.3. LSDV Facilitates the Activation of ER Stress-Related Apoptosis in BEF Cells
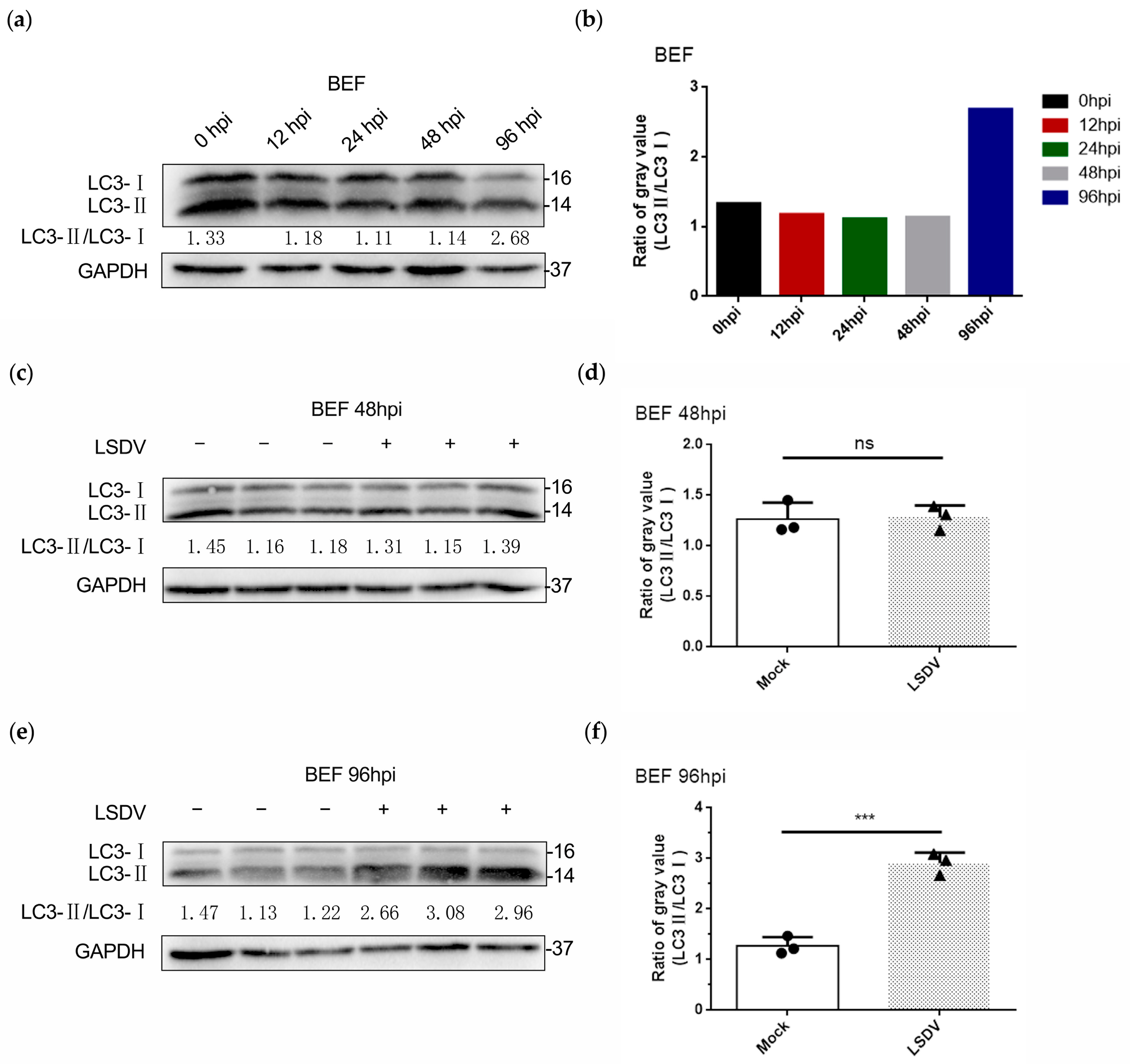
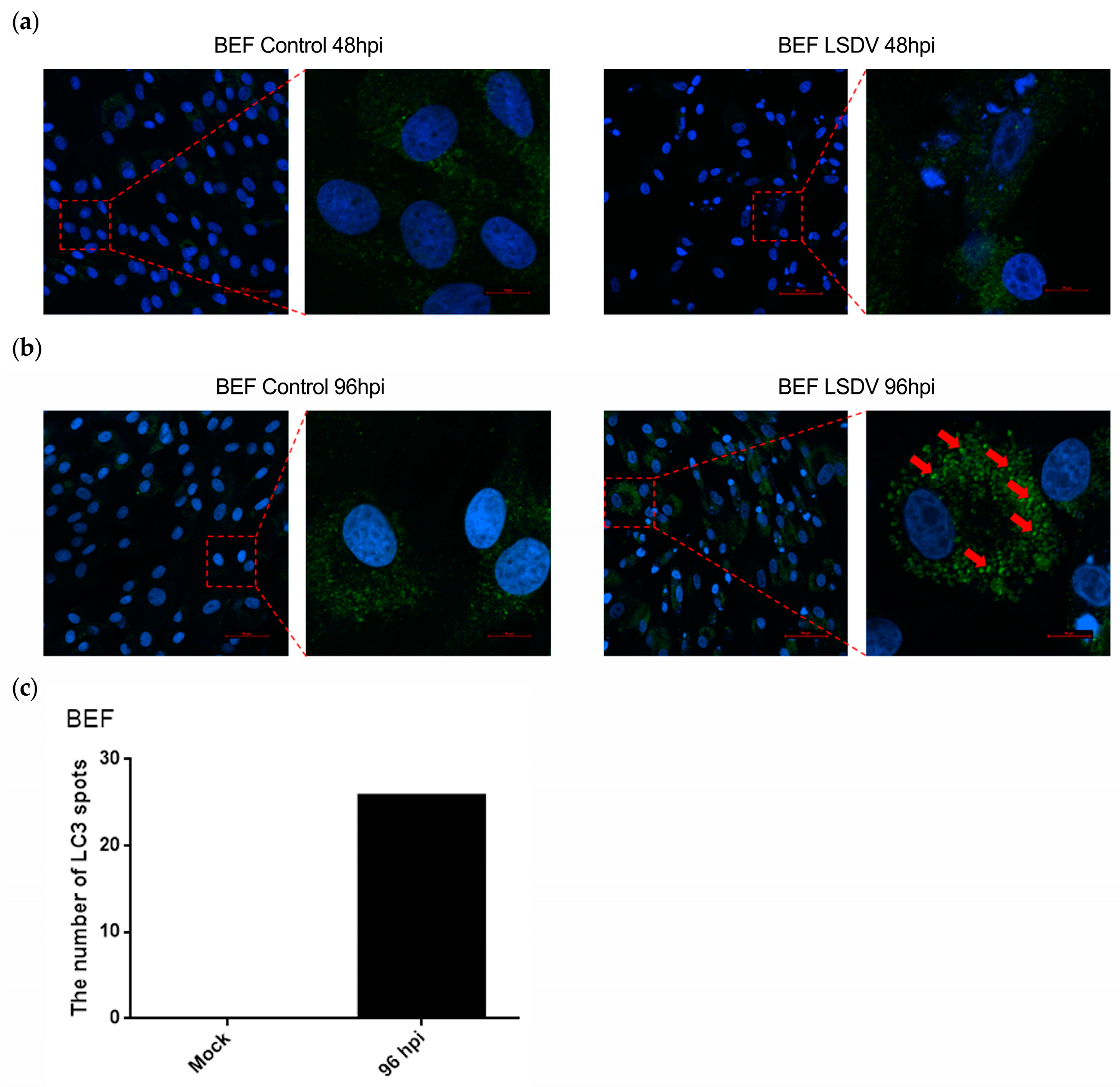
3.4. LSDV Infection Promotes Autophagy in BEF Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, Y.R.; Ali, A.; Hussain, K.; Ijaz, M.; Rabbani, A.H.; Khan, R.L.; Abbas, S.N.; Aziz, M.U.; Ghaffar, A.; Sajid, H.A. A review: Surveillance of lumpy skin disease (LSD) a growing problem in Asia. Microb. Pathog. 2021, 158, 105050. [Google Scholar] [CrossRef]
- Hunter, P.; Wallace, D. Lumpy skin disease in southern Africa: A review of the disease and aspects of control. J. S. Afr. Vet. Assoc. 2001, 72, 68–71. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.; Oura, C.A. Review: Lumpy skin disease: An emerging threat to Europe, the Middle East and Asia. Transbound. Emerg. Dis. 2012, 59, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Ratyotha, K.; Prakobwong, S.; Piratae, S. Lumpy skin disease: A newly emerging disease in Southeast Asia. Vet. World 2022, 15, 2764–2771. [Google Scholar] [CrossRef]
- Whittle, L.; Chapman, R.; Williamson, A.L. Lumpy Skin Disease-An Emerging Cattle Disease in Europe and Asia. Vaccines 2023, 11, 578. [Google Scholar] [CrossRef] [PubMed]
- Yeruham, I.; Nir, O.; Braverman, Y.; Davidson, M.; Grinstein, H.; Haymovitch, M.; Zamir, O. Spread of lumpy skin disease in Israeli dairy herds. Vet. Rec. 1995, 137, 91–93. [Google Scholar] [CrossRef]
- Tageldin, M.H.; Wallace, D.B.; Gerdes, G.H.; Putterill, J.F.; Greyling, R.R.; Phosiwa, M.N.; Al Busaidy, R.M.; Al Ismaaily, S.I. Lumpy skin disease of cattle: An emerging problem in the Sultanate of Oman. Trop. Anim. Health Prod. 2014, 46, 241–246. [Google Scholar] [CrossRef]
- Tsai, K.J.; Tu, Y.C.; Wu, C.H.; Huang, C.W.; Ting, L.J.; Huang, Y.L.; Pan, C.H.; Chang, C.Y.; Deng, M.C.; Lee, F. First detection and phylogenetic analysis of lumpy skin disease virus from Kinmen Island, Taiwan in 2020. J. Vet. Med. Sci. 2022, 84, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Abutarbush, S.M.; Ababneh, M.M.; Al Zoubi, I.G.; Al Sheyab, O.M.; Al Zoubi, M.G.; Alekish, M.O.; Al Gharabat, R.J. Lumpy Skin Disease in Jordan: Disease Emergence, Clinical Signs, Complications and Preliminary-associated Economic Losses. Transbound. Emerg. Dis. 2015, 62, 549–554. [Google Scholar] [CrossRef]
- Santhamani, R.; Yogisharadhya, R.; Venkatesan, G.; Shivachandra, S.B.; Pandey, A.B.; Ramakrishnan, M.A. Molecular characterization of Indian sheeppox and goatpox viruses based on RPO30 and GPCR genes. Virus Genes 2014, 49, 286–291. [Google Scholar] [CrossRef]
- Sumana, K.; Revanaiah, Y.; Shivachandra, S.B.; Mothay, D.; Apsana, R.; Saminathan, M.; Basavaraj, S.; Reddy, G.B.M. Molecular phylogeny of Capripoxviruses based on major immunodominant protein (P32) reveals circulation of host specific sheeppox and goatpox viruses in small ruminants of India. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2020, 85, 104472. [Google Scholar] [CrossRef] [PubMed]
- Sprygin, A.; Pestova, Y.; Bjadovskaya, O.; Prutnikov, P.; Zinyakov, N.; Kononova, S.; Ruchnova, O.; Lozovoy, D.; Chvala, I.; Kononov, A. Evidence of recombination of vaccine strains of lumpy skin disease virus with field strains, causing disease. PLoS ONE 2020, 15, e0232584. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, J.; Bamouh, Z.; Jazouli, M.; Boumart, Z.; Tadlaoui, K.O.; Fihri, O.F.; El Harrak, M. Experimental evaluation of the cross-protection between Sheeppox and bovine Lumpy skin vaccines. Sci. Rep. 2020, 10, 8888. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Barua, S.; Kumar, R.; Khandelwal, N.; Kumar, A.; Verma, A.; Singh, L.; Godara, B.; Chander, Y.; Kumar, G.; et al. Evaluation of the safety, immunogenicity and efficacy of a new live-attenuated lumpy skin disease vaccine in India. Virulence 2023, 14, 2190647. [Google Scholar] [CrossRef]
- Davies, F.G. Lumpy skin disease, an African capripox virus disease of cattle. Br. Vet. J. 1991, 147, 489–503. [Google Scholar] [CrossRef]
- Kononov, A.; Prutnikov, P.; Shumilova, I.; Kononova, S.; Nesterov, A.; Byadovskaya, O.; Pestova, Y.; Diev, V.; Sprygin, A. Determination of lumpy skin disease virus in bovine meat and offal products following experimental infection. Transbound. Emerg. Dis. 2019, 66, 1332–1340. [Google Scholar] [CrossRef]
- Hasib, F.M.Y.; Islam, M.S.; Das, T.; Rana, E.A.; Uddin, M.H.; Bayzid, M.; Nath, C.; Hossain, M.A.; Masuduzzaman, M.; Das, S.; et al. Lumpy skin disease outbreak in cattle population of Chattogram, Bangladesh. Vet. Med. Sci. 2021, 7, 1616–1624. [Google Scholar] [CrossRef]
- Tan, J.; Liu, Y.; Yang, F.; Chen, G.; Fang, Y.; He, X.; Lou, Z.; Jia, H.; Jing, Z.; Li, W. Emerging evidence for poxvirus-mediated unfolded protein response: Lumpy skin disease virus maintains self-replication by activating PERK and IRE1 signaling. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2023, 37, e22902. [Google Scholar] [CrossRef]
- Fu, X.; Liu, J.; Liu, D.; Zhou, Y.; Guo, Y.; Wang, Z.; Yang, S.; He, W.; Chen, P.; Wang, X.; et al. Glucose-regulated protein 78 modulates cell growth, epithelial-mesenchymal transition, and oxidative stress in the hyperplastic prostate. Cell Death Dis. 2022, 13, 78. [Google Scholar] [CrossRef]
- Hu, H.; Tian, M.; Ding, C.; Yu, S. The C/EBP Homologous Protein (CHOP) Transcription Factor Functions in Endoplasmic Reticulum Stress-Induced Apoptosis and Microbial Infection. Front. Immunol. 2018, 9, 3083. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, Y.; Wang, Z.; Liu, X.; Lu, S.; Hu, X. A Native Drug-Free Macromolecular Therapeutic to Trigger Mutual Reinforcing of Endoplasmic Reticulum Stress and Mitochondrial Dysfunction for Cancer Treatment. ACS Nano 2023, 17, 11023–11038. [Google Scholar] [CrossRef] [PubMed]
- Mareninova, O.A.; Jia, W.; Gretler, S.R.; Holthaus, C.L.; Thomas, D.D.H.; Pimienta, M.; Dillon, D.L.; Gukovskaya, A.S.; Gukovsky, I.; Groblewski, G.E. Transgenic expression of GFP-LC3 perturbs autophagy in exocrine pancreas and acute pancreatitis responses in mice. Autophagy 2020, 16, 2084–2097. [Google Scholar] [CrossRef]
- Vorou, R.M.; Papavassiliou, V.G.; Pierroutsakos, I.N. Cowpox virus infection: An emerging health threat. Curr. Opin. Infect. Dis. 2008, 21, 153–156. [Google Scholar] [CrossRef]
- Caravaglio, J.V.; Khachemoune, A. Orf Virus Infection in Humans: A Review with a Focus on Advances in Diagnosis and Treatment. J. Drugs Dermatol. JDD 2017, 16, 684–689. [Google Scholar] [PubMed]
- Petersen, E.; Kantele, A.; Koopmans, M.; Asogun, D.; Yinka-Ogunleye, A.; Ihekweazu, C.; Zumla, A. Human Monkeypox: Epidemiologic and Clinical Characteristics, Diagnosis, and Prevention. Infect. Dis. Clin. N. Am. 2019, 33, 1027–1043. [Google Scholar] [CrossRef] [PubMed]
- MacIntyre, C.R. Reevaluating the Risk of Smallpox Reemergence. Mil. Med. 2020, 185, e952–e957. [Google Scholar] [CrossRef] [PubMed]
- Meseda, C.A.; Weir, J.P. Third-generation smallpox vaccines: Challenges in the absence of clinical smallpox. Future Microbiol. 2010, 5, 1367–1382. [Google Scholar] [CrossRef]
- Thèves, C.; Biagini, P.; Crubézy, E. The rediscovery of smallpox. Clin. Microbiol. Infect. 2014, 20, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Ginglen, J.G.; Doyle, M.Q. Immunization. In StatPearls; StatPearls Publishing LLC.: St. Petersburg, FL, USA, 2023. [Google Scholar]
- Meyer, H.; Ehmann, R.; Smith, G.L. Smallpox in the Post-Eradication Era. Viruses 2020, 12, 138. [Google Scholar] [CrossRef]
- Zafar, S.J.; Shishido, A.A. Smallpox as a Bioagent: A Refresher and Update for the SOF Provider. J. Spec. Oper. Med. A Peer Rev. J. SOF Med. Prof. 2022, 22, 124–128. [Google Scholar] [CrossRef]
- Rahi, M.; Joy, S.; Sharma, A. Public Health Challenges in the Context of the Global Spread of Mpox Infections. Am. J. Trop. Med. Hyg. 2023, 108, 641–645. [Google Scholar] [CrossRef]
- Aden, D.; Zaheer, S.; Kumar, R.; Ranga, S. Monkeypox (Mpox) outbreak during COVID-19 pandemic—Past and the future. J. Med. Virol. 2023, 95, e28701. [Google Scholar] [CrossRef] [PubMed]
- Sohal, P.; Gupta, A.; Gupta, S.; Gupta, V.; Jain, R.; Jain, R. Monkeypox: Another pandemic in the making? Bayl. Univ. Med. Cent. Proc. 2023, 36, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, M.A.; El-Deeb, A.; El-Tholoth, M.; Hoffmann, D.; Czerny, C.P.; Hufert, F.T.; Weidmann, M.; Abd El Wahed, A. Recombinase polymerase amplification assay for rapid detection of lumpy skin disease virus. BMC Vet. Res. 2016, 12, 244. [Google Scholar] [CrossRef] [PubMed]
- Annandale, C.H.; Smuts, M.P.; Ebersohn, K.; du Plessis, L.; Thompson, P.N.; Venter, E.H.; Stout, T.A.E. Effect of using frozen-thawed bovine semen contaminated with lumpy skin disease virus on in vitro embryo production. Transbound. Emerg. Dis. 2019, 66, 1539–1547. [Google Scholar] [CrossRef] [PubMed]
- Milovanović, M.; Milićević, V.; Radojičić, S.; Valčić, M.; Hoffmann, B.; Dietze, K. Suitability of individual and bulk milk samples to investigate the humoral immune response to lumpy skin disease vaccination by ELISA. Virol. J. 2020, 17, 28. [Google Scholar] [CrossRef] [PubMed]
- Verfaillie, T.; Rubio, N.; Garg, A.D.; Bultynck, G.; Rizzuto, R.; Decuypere, J.P.; Piette, J.; Linehan, C.; Gupta, S.; Samali, A.; et al. PERK is required at the ER-mitochondrial contact sites to convey apoptosis after ROS-based ER stress. Cell Death Differ. 2012, 19, 1880–1891. [Google Scholar] [CrossRef]
- Ji, C.; Mehrian-Shai, R.; Chan, C.; Hsu, Y.H.; Kaplowitz, N. Role of CHOP in hepatic apoptosis in the murine model of intragastric ethanol feeding. Alcohol. Clin. Exp. Res. 2005, 29, 1496–1503. [Google Scholar] [CrossRef]
- Zhang, H.; Monken, C.E.; Zhang, Y.; Lenard, J.; Mizushima, N.; Lattime, E.C.; Jin, S. Cellular autophagy machinery is not required for vaccinia virus replication and maturation. Autophagy 2006, 2, 91–95. [Google Scholar] [CrossRef]
- Moloughney, J.G.; Monken, C.E.; Tao, H.; Zhang, H.; Thomas, J.D.; Lattime, E.C.; Jin, S. Vaccinia virus leads to ATG12–ATG3 conjugation and deficiency in autophagosome formation. Autophagy 2011, 7, 1434–1447. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, J.; Liu, Y.; Li, W.; Zhang, Y.; Chen, G.; Fang, Y.; He, X.; Jing, Z. Lumpy Skin Disease Virus Infection Activates Autophagy and Endoplasmic Reticulum Stress-Related Cell Apoptosis in Primary Bovine Embryonic Fibroblast Cells. Microorganisms 2023, 11, 1883. https://doi.org/10.3390/microorganisms11081883
Tan J, Liu Y, Li W, Zhang Y, Chen G, Fang Y, He X, Jing Z. Lumpy Skin Disease Virus Infection Activates Autophagy and Endoplasmic Reticulum Stress-Related Cell Apoptosis in Primary Bovine Embryonic Fibroblast Cells. Microorganisms. 2023; 11(8):1883. https://doi.org/10.3390/microorganisms11081883
Chicago/Turabian StyleTan, Jinlong, Yinju Liu, Weike Li, Yongzhi Zhang, Guohua Chen, Yongxiang Fang, Xiaobing He, and Zhizhong Jing. 2023. "Lumpy Skin Disease Virus Infection Activates Autophagy and Endoplasmic Reticulum Stress-Related Cell Apoptosis in Primary Bovine Embryonic Fibroblast Cells" Microorganisms 11, no. 8: 1883. https://doi.org/10.3390/microorganisms11081883
APA StyleTan, J., Liu, Y., Li, W., Zhang, Y., Chen, G., Fang, Y., He, X., & Jing, Z. (2023). Lumpy Skin Disease Virus Infection Activates Autophagy and Endoplasmic Reticulum Stress-Related Cell Apoptosis in Primary Bovine Embryonic Fibroblast Cells. Microorganisms, 11(8), 1883. https://doi.org/10.3390/microorganisms11081883