


Distribution Patterns of Benthic Protist Communities Depending on Depth Revealed by Environmental Sequencing—From the Sublittoral to the Deep Sea


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
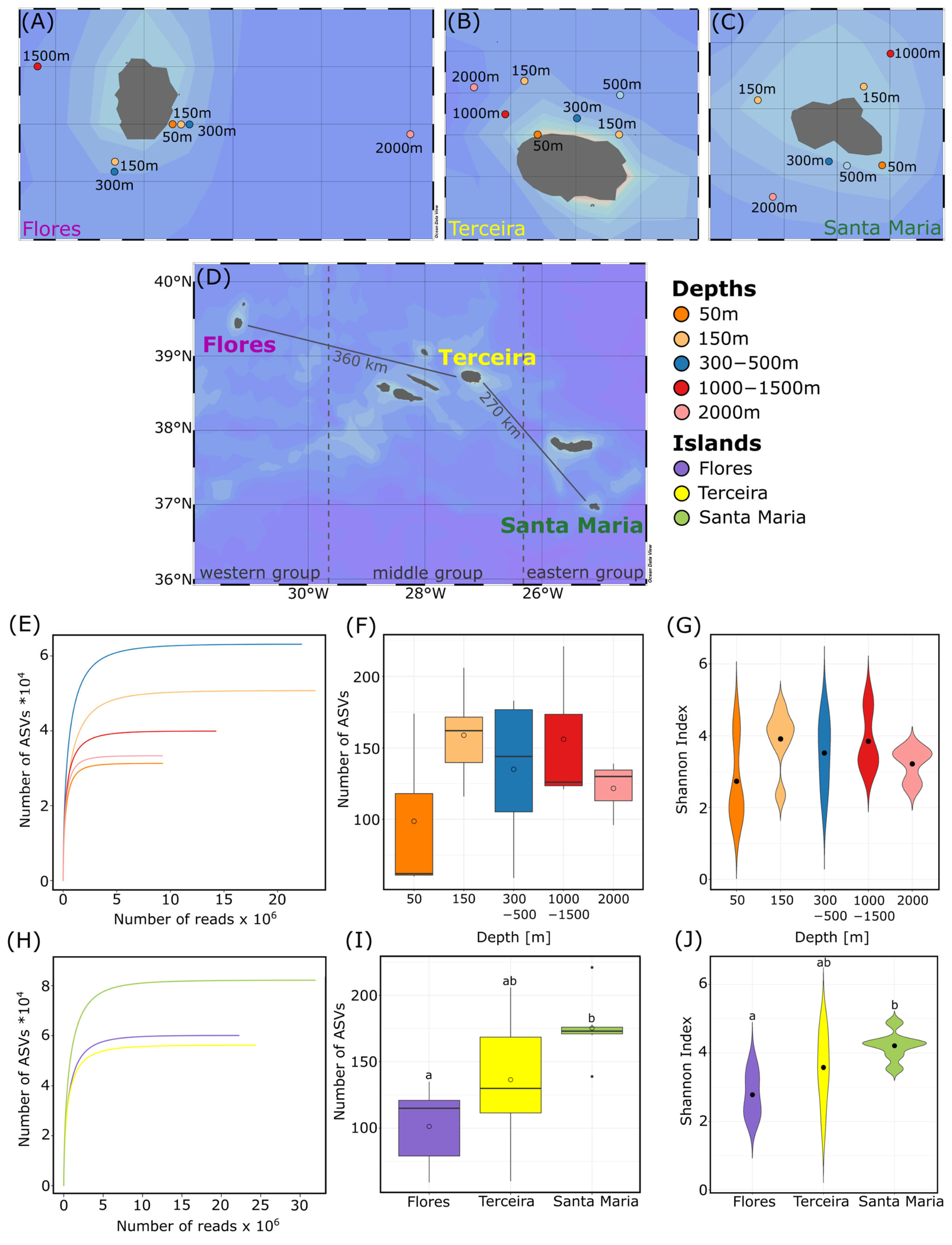
2.1. Sampling
2.2. DNA Extraction, PCR Amplification and High-Throughput Sequencing
2.3. Bioinformatic Processing
2.4. Statistical Analyses
3. Results and Discussion
3.1. Alpha Diversity
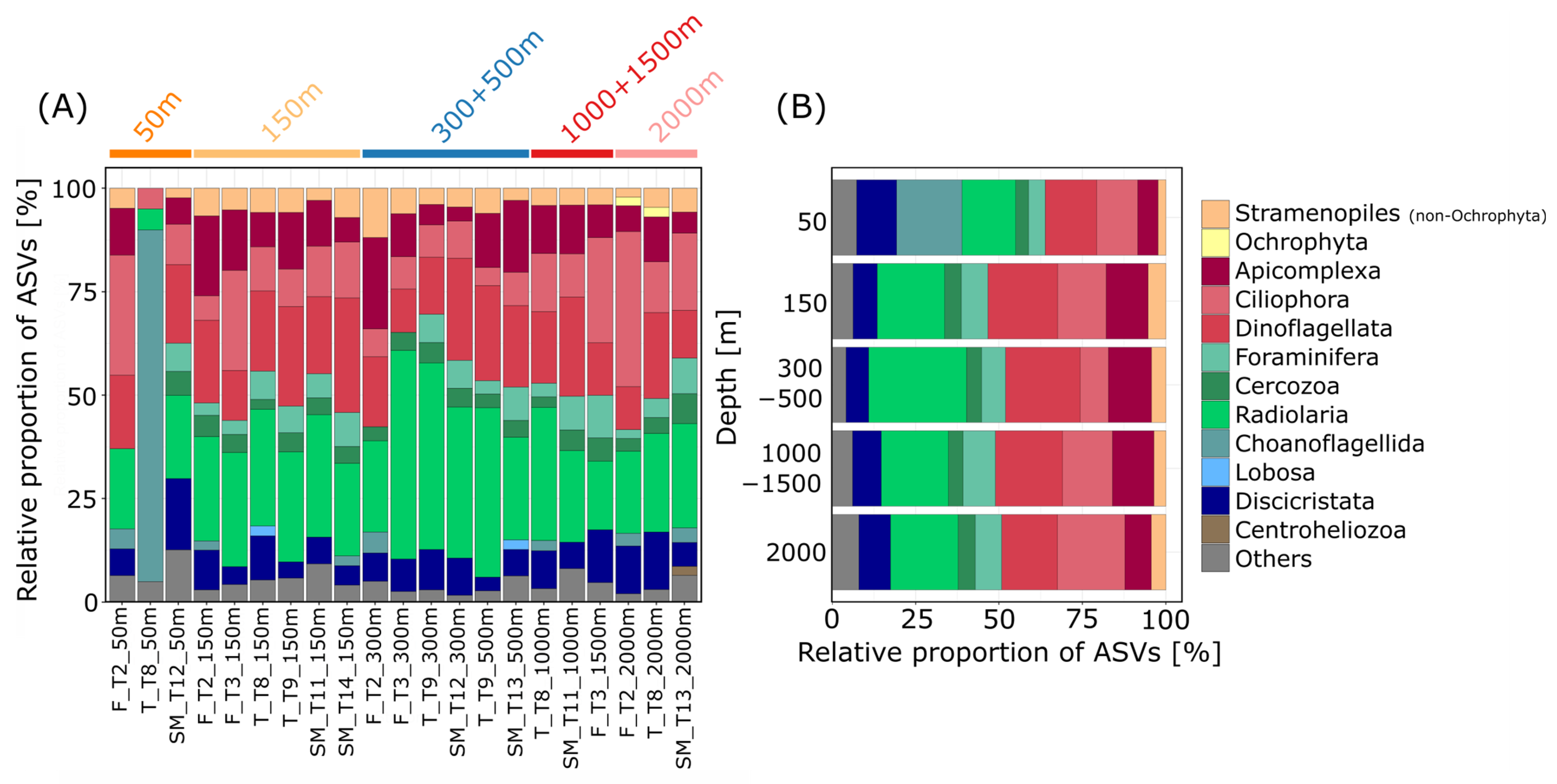
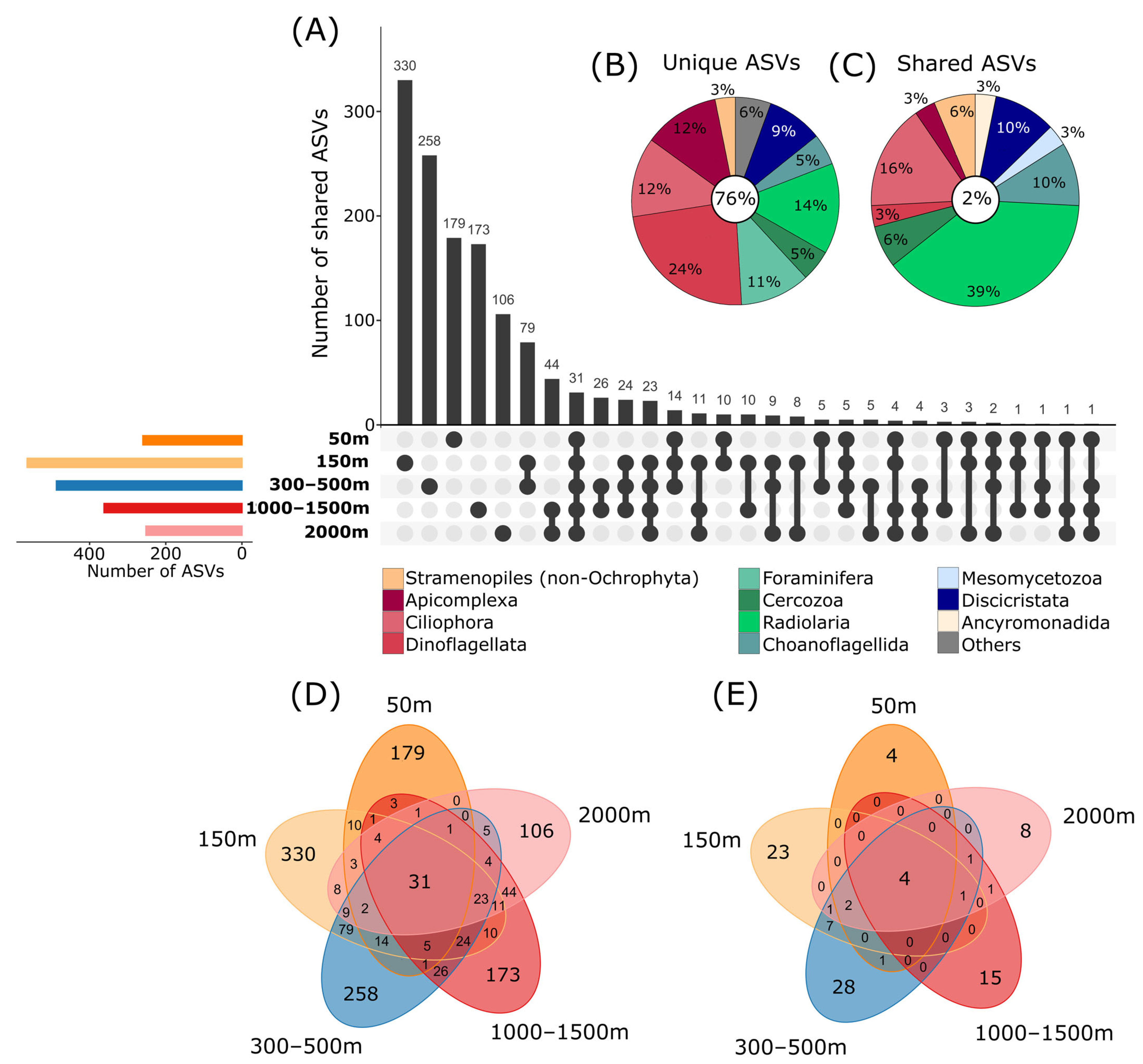
3.2. Taxonomic Composition of Communities in Different Depths
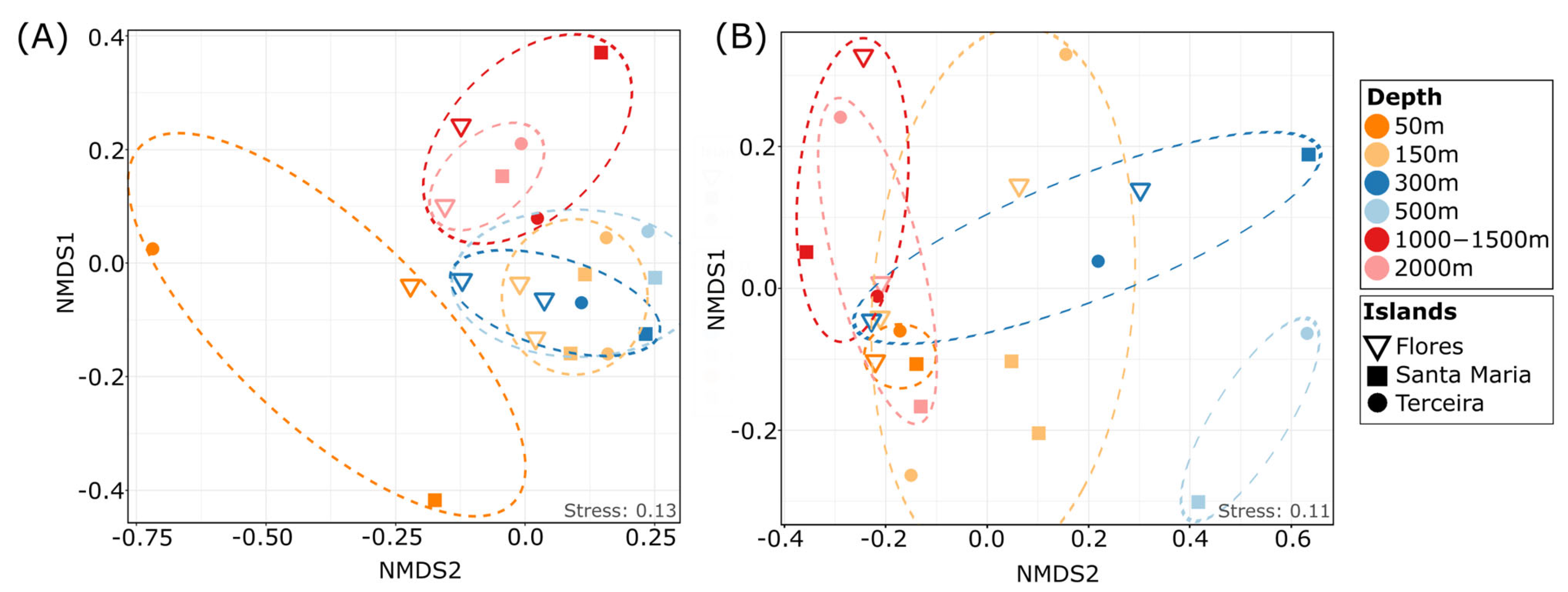
3.3. Depth-Dependent Patterns of Protist Communities
3.4. Genotype Distribution of Cultivated Flagellates
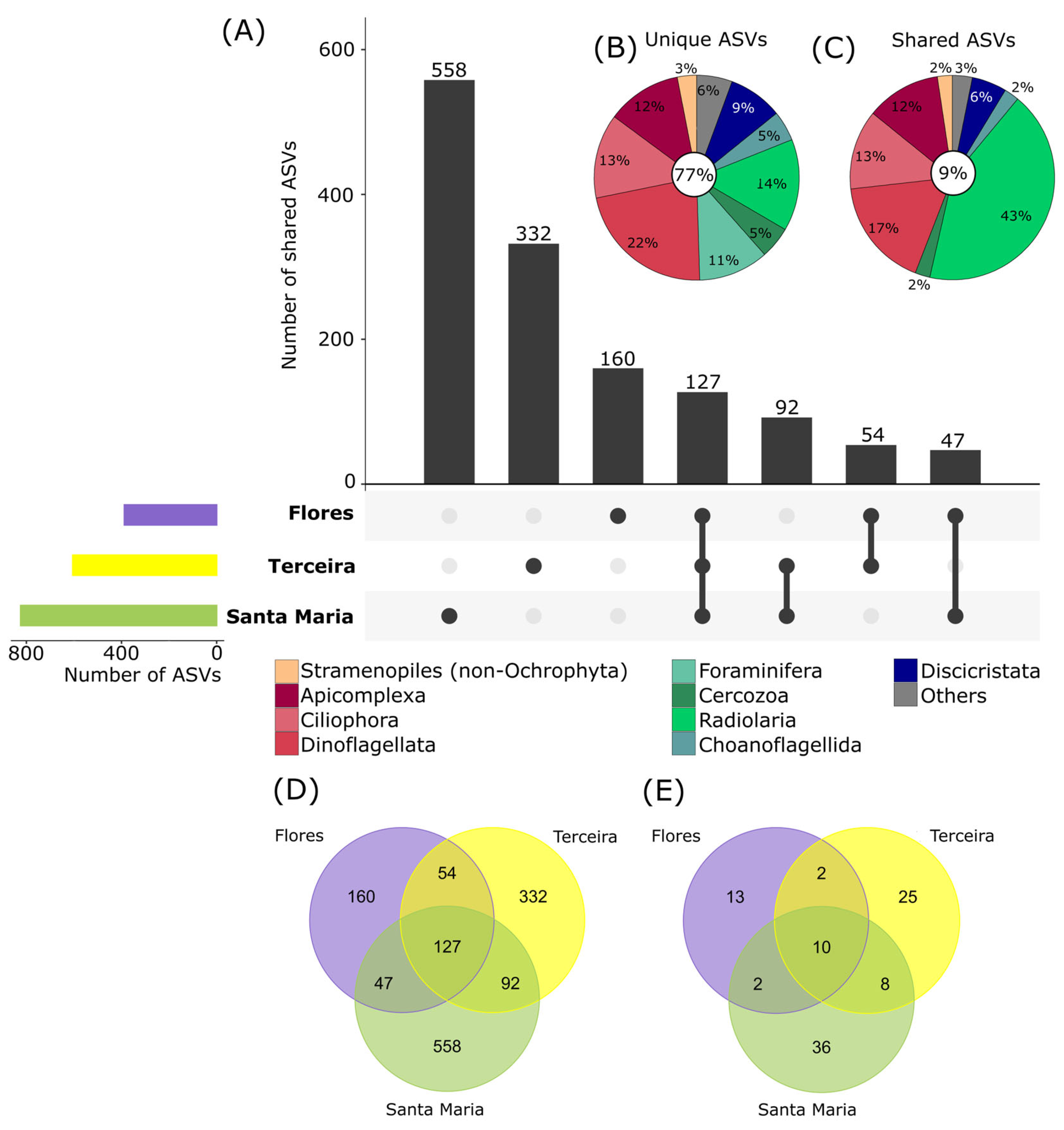
3.5. Patterns of Protist Communities at Different Islands
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Vargas, C.; Audic, S.; Henry, N.; Decelle, J.; Mahé, F.; Logares, R.; Lara, E.; Berney, C.; Le Boscot, N.; Probert, I.; et al. Eukaryotic Plankton Diversity in the Sunlit Ocean. Science 2015, 348, 1261605. [Google Scholar] [CrossRef]
- Pernice, M.C.; Giner, C.R.; Logares, R.; Perera-Bel, J.; Acinas, S.G.; Duarte, C.M.; Gasol, J.M.; Massana, R. Large Variability of Bathypelagic Microbial Eukaryotic Communities across the World’s Oceans. ISME J. 2016, 10, 945–958. [Google Scholar] [CrossRef] [PubMed]
- Worden, A.Z.; Follows, M.J.; Giovannoni, S.J.; Wilken, S.; Zimmerman, A.E.; Keeling, P.J. Rethinking the Marine Carbon Cycle: Factoring in the Multifarious Lifestyles of Microbes. Science 2015, 347, 1257594. [Google Scholar] [CrossRef] [PubMed]
- Cordier, T.; Angeles, I.B.; Henry, N.; Lejzerowicz, F.; Berney, C.; Morard, R.; Brandt, A.; Cambon-Bonavita, M.-A.; Guidi, L.; Lombard, F.; et al. Patterns of Eukaryotic Diversity from the Surface to the Deep-Ocean Sediment. Sci. Adv. 2022, 8, eabj9309. [Google Scholar] [CrossRef] [PubMed]
- Schoenle, A.; Hohlfeld, M.; Hermanns, K.; Mahé, F.; de Vargas, C.; Nitsche, F.; Arndt, H. High and Specific Diversity of Protists in the Deep-Sea Basins Dominated by Diplonemids, Kinetoplastids, Ciliates and Foraminiferans. Commun. Biol. 2021, 4, 501. [Google Scholar] [CrossRef]
- Gooday, A.J.; Schoenle, A.; Dolan, J.R.; Arndt, H. Protist Diversity and Function in the Dark Ocean—Challenging the Paradigms of Deep-Sea Ecology with Special Emphasis on Foraminiferans and Naked Protists. Eur. J. Protistol. 2020, 75, 125721. [Google Scholar] [CrossRef]
- Hohlfeld, M.; Schoenle, A.; Arndt, H. Horizontal and Vertical Small-Scale Patterns of Protist Communities at the Atlantic Deep-Sea Floor. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2021, 171, 103515. [Google Scholar] [CrossRef]
- Gong, J.; Shi, F.; Ma, B.; Dong, J.; Pachiadaki, M.; Zhang, X.; Edgcomb, V.P. Depth Shapes α- and β-Diversities of Microbial Eukaryotes in Surficial Sediments of Coastal Ecosystems. Environ. Microbiol. 2015, 17, 3722–3737. [Google Scholar] [CrossRef]
- George, K.H. Faunistic Research on Metazoan Meiofauna from Seamounts—A Review. Meiofauna Mar. 2013, 20, 1–32. [Google Scholar]
- Whittaker, R.J.; Triantis, K.A.; Ladle, R.J. A General Dynamic Theory of Oceanic Island Biogeography. J. Biogeogr. 2008, 35, 977–994. [Google Scholar] [CrossRef]
- Salani, F.S.; Arndt, H.; Hausmann, K.; Nitsche, F.; Scheckenbach, F. Analysis of the Community Structure of Abyssal Kinetoplastids Revealed Similar Communities at Larger Spatial Scales. ISME J. 2012, 6, 713–723. [Google Scholar] [CrossRef]
- Alongi, D.M. Bathymetric Patterns of Deep-Sea Benthic Communities from Bathyal to Abyssal Depths in the Western South Pacific (Solomon and Coral Seas). Deep Sea Res. Part A Oceanogr. Res. Pap. 1992, 39, 549–565. [Google Scholar] [CrossRef]
- Arndt, H.; Hausmann, K.; Wolf, M. Deep-Sea Heterotrophic Nanoflagellates of the Eastern Mediterranean Sea: Qualitative and Quantitative Aspects of Their Pelagic and Benthic Occurrence. Mar. Ecol. Prog. Ser. 2003, 256, 45–56. [Google Scholar] [CrossRef]
- Bak, R.P.M.; Nieuwland, G. Seasonal Variation in Bacterial and Flagellate Communities of Deep-Sea Sediments in a Monsoonal Upwelling System. Deep Sea Res. Part II 1997, 44, 1281–1292. [Google Scholar] [CrossRef]
- Hohlfeld, M.; Arndt, H. Changes in the Abundance and Taxonomic Composition of Benthic Heterotrophic Protists from Atlantic Sublittoral to Deep-Sea Sediments. Diversity 2022, 14, 164. [Google Scholar] [CrossRef]
- Hachich, N.F.; Bonsall, M.; Arraut, E.; Barneche, D.; Lewinsohn, T.; Floeter, S. Island Biogeography: Patterns of Marine Shallow-water Organisms in the Atlantic Ocean. J. Biogeogr. 2015, 42, 1871–1882. [Google Scholar] [CrossRef]
- Mazzei, E.F.; Pinheiro, H.T.; Simon, T.; Moura, R.L.; Macieira, R.M.; Pimentel, C.R.; Teixeira, J.B.; Floeter, S.R.; Ferreira, C.E.L.; Ghisolfi, R.D.; et al. Mechanisms of Dispersal and Establishment Drive a Stepping Stone Community Assembly on Seamounts and Oceanic Islands. Mar. Biol. 2021, 168, 109. [Google Scholar] [CrossRef]
- Pinheiro, H.T.; Bernardi, G.; Simon, T.; Joyeux, J.-C.; Macieira, R.M.; Gasparini, J.L.; Rocha, C.; Rocha, L.A. Island Biogeography of Marine Organisms. Nature 2017, 549, 82–85. [Google Scholar] [CrossRef]
- Finlay, B.J. Global Dispersal of Free-Living Microbial Eukaryote Species. Science 2002, 296, 1061–1063. [Google Scholar] [CrossRef]
- George, K.; Arndt, H.; Wehrmann, A.; Baptista, L.; Berning, B.; Bruhn, M.; Carvalho, F.; Cordeiro, R.; Creemers, M.; Defise, A.; et al. Controls in Benthic and Pelagic BIODIversity of the AZores BIODIAZ; METEOR-Berichte; Senckenberg am Meer: Wilhelmshaven, Germany, 2018. [Google Scholar]
- Danovaro, R. Methods for the Study of Deep-Sea Sediments, Their Functioning and Biodiversity; CRC Press: Boca Raton, FL, USA; Taylor & Francis Group: Boca Raton, FL, USA, 2009. [Google Scholar]
- Fortin, N.; Beaumier, D.; Lee, K.; Greer, C.W. Soil Washing Improves the Recovery of Total Community DNA from Polluted and High Organic Content Sediments. J. Microbiol. Methods 2004, 56, 181–191. [Google Scholar] [CrossRef]
- Amaral-Zettler, L.A.; McCliment, E.A.; Ducklow, H.W.; Huse, S.M. A Method for Studying Protistan Diversity Using Massively Parallel Sequencing of V9 Hypervariable Regions of Small-Subunit Ribosomal RNA Genes. PLoS ONE 2009, 4, e6372. [Google Scholar] [CrossRef]
- Michu, E.; Mráčková, M.; Vyskot, B.; Žlŭvová, J. Reduction of Heteroduplex Formation in PCR Amplification. Biol. Plant. 2010, 54, 173–176. [Google Scholar] [CrossRef]
- Lennartz née Rybarski, A.E.L.; Nitsche, F.; Schoenle, A.; Voigt, C.; Staubwasser, M.; Arndt, H. High Diversity and Isolated Distribution of Aquatic Heterotrophic Protists in Salars of the Atacama Desert at Different Salinities. Eur. J. Protistol. 2023, 89, 125987. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Guillou, L.; Bachar, D.; Audic, S.; Bass, D.; Berney, C.; Bittner, L.; Boutte, C.; Burgaud, G.; de Vargas, C.; Decelle, J.; et al. The Protist Ribosomal Reference Database (PR2): A Catalog of Unicellular Eukaryote Small Sub-Unit RRNA Sequences with Curated Taxonomy. Nucleic Acids Res. 2013, 41, D597–D604. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Orksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-2; R Core Team: Vienna, Austria, 2018. [Google Scholar]
- Chen, H.; Boutros, P.C. VennDiagram: A Package for the Generation of Highly-Customizable Venn and Euler Diagrams in R. BMC Bioinform. 2011, 12, 35. [Google Scholar] [CrossRef]
- Conway, J.R.; Lex, A.; Gehlenborg, N. UpSetR: An R Package for the Visualization of Intersecting Sets and Their Properties. Bioinformatics 2017, 33, 2938–2940. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Mahé, F.; Mayor, J.; Bunge, J.; Chi, J.; Siemensmeyer, T.; Stoeck, T.; Wahl, B.; Paprotka, T.; Filker, S.; Dunthorn, M. Comparing High-Throughput Platforms for Sequencing the V4 Region of SSU-RDNA in Environmental Microbial Eukaryotic Diversity Surveys. J. Eukaryot. Microbiol. 2015, 62, 338–345. [Google Scholar] [CrossRef]
- Choi, J.; Park, J.S. Comparative Analyses of the V4 and V9 Regions of 18S RDNA for the Extant Eukaryotic Community Using the Illumina Platform. Sci. Rep. 2020, 10, 6519. [Google Scholar] [CrossRef]
- Biard, T. Diversity and Ecology of Radiolaria in Modern Oceans. Environ. Microbiol. 2022, 24, 2179–2200. [Google Scholar] [CrossRef]
- Coats, D.W. Parasitic Life Styles of Marine Dinoflagellates. J. Eukaryot. Microbiol. 1999, 46, 402–409. [Google Scholar] [CrossRef]
- Guillou, L.; Viprey, M.; Chambouvet, A.; Welsh, R.M.; Kirkham, A.R.; Massana, R.; Scanlan, D.J.; Worden, A.Z. Widespread Occurrence and Genetic Diversity of Marine Parasitoids Belonging to Syndiniales (Alveolata). Environ. Microbiol. 2008, 10, 3349–3365. [Google Scholar] [CrossRef]
- Siano, R.; Alves-de-Souza, C.; Foulon, E.; Bendif, E.M.; Simon, N.; Guillou, L.; Not, F. Distribution and Host Diversity of Amoebophryidae Parasites across Oligotrophic Waters of the Mediterranean Sea. Biogeosciences 2011, 8, 267–278. [Google Scholar] [CrossRef]
- Gómez, F.; Moreira, D.; López-García, P. Molecular Phylogeny of Noctilucoid Dinoflagellates (Noctilucales, Dinophyceae). Protist 2010, 161, 466–478. [Google Scholar] [CrossRef]
- Janouškovec, J.; Gavelis, G.; Burki, F.; Dinh, D.; Bachvaroff, T.; Gornik, S.; Bright, K.; Imanian, B.; Strom, S.; Delwiche, C.; et al. Major Transitions in Dinoflagellate Evolution Unveiled by Phylotranscriptomics. PNAS 2016, 114, E171–E180. [Google Scholar] [CrossRef]
- Schoenle, A.; Nitsche, F.; Werner, J.; Arndt, H. Deep-Sea Ciliates: Recorded Diversity and Experimental Studies on Pressure Tolerance. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2017, 128, 55–66. [Google Scholar] [CrossRef]
- Flegontova, O.; Flegontov, P.; Londoño, P.A.C.; Walczowski, W.; Šantić, D.; Edgcomb, V.P.; Lukeš, J.; Horák, A. Environmental Determinants of the Distribution of Planktonic Diplonemids and Kinetoplastids in the Oceans. Environ. Microbiol. 2020, 22, 4014–4031. [Google Scholar] [CrossRef]
- Flegontova, O.; Flegontov, P.; Malviya, S.; Audic, S.; Wincker, P.; de Vargas, C.; Bowler, C.; Lukeš, J.; Horák, A. Extreme Diversity of Diplonemid Eukaryotes in the Ocean. Curr. Biol. 2016, 26, 3060–3065. [Google Scholar] [CrossRef]
- Obiol, A.; Giner, C.R.; Sánchez, P.; Duarte, C.M.; Acinas, S.G.; Massana, R. A Metagenomic Assessment of Microbial Eukaryotic Diversity in the Global Ocean. Mol. Ecol. Resour. 2020, 20, 718–731. [Google Scholar] [CrossRef] [PubMed]
- Tashyreva, D.; Simpson, A.G.B.; Prokopchuk, G.; Škodová-Sveráková, I.; Butenko, A.; Hammond, M.; George, E.E.; Flegontova, O.; Záhonová, K.; Faktorová, D.; et al. Diplonemids—A Review on “New” Flagellates on the Oceanic Block. Protist 2022, 173, 125868. [Google Scholar] [CrossRef] [PubMed]
- Kent, M.; Elston, R.A. An Isonema-like Flagellate (Protozoa: Mastigophora) Infection in Larval Geoduck Clams, Panope Abrupta. J. Invertebr. Pathol. 1987, 50, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Lukeš, J.; Flegontova, O.; Horák, A. Diplonemids. Curr. Biol. 2015, 25, R702–R704. [Google Scholar] [CrossRef]
- Schoenle, A.; Hohlfeld, M.; Rosse, M.; Filz, P.; Wylezich, C.; Nitsche, F.; Arndt, H. Global Comparison of Bicosoecid Cafeteria-like Flagellates from the Deep Ocean and Surface Waters, with Reorganization of the Family Cafeteriaceae. Eur. J. Protistol. 2020, 73, 125665. [Google Scholar] [CrossRef]
- Schoenle, A.; Hohlfeld, M.; Rybarski, A.; Sachs, M.; Freches, E.; Wiechmann, K.; Nitsche, F.; Arndt, H. Cafeteria in Extreme Environments: Investigations on C. burkhardae and Three New Species from the Atacama Desert and the Deep Ocean. Eur. J. Protistol. 2022, 85, 125905. [Google Scholar] [CrossRef]
- Leadbeater, B.S.C. Choanoflagellate Lorica Construction and Assembly: The Nudiform Condition. I. Savillea Species. Protist 2008, 159, 259–268. [Google Scholar] [CrossRef]
- Schiwitza, S.; Arndt, H.; Nitsche, F. First Description of an Euryoecious Acanthoecid Choanoflagellate Species, Enibas tolerabilis Gen. et Sp. Nov. from a Salar in the Chilean Andes Based on Morphological and Transcriptomic Data. Eur. J. Protistol. 2019, 67, 106–113. [Google Scholar] [CrossRef]
- Decelle, J.; Romac, S.; Sasaki, E.; Not, F.; Mahé, F. Intracellular Diversity of the V4 and V9 Regions of the 18S RRNA in Marine Protists (Radiolarians) Assessed by High-Throughput Sequencing. PLoS ONE 2014, 9, e104297. [Google Scholar] [CrossRef]
- Galluzzi, L.; Penna, A.; Bertozzini, E.; Vila, M.; Garcés, E.; Magnani, M. Development of a Real-Time PCR Assay for Rapid Detection and Quantification of Alexandrium minutum (a Dinoflagellate). Appl. Environ. Microbiol. 2004, 70, 1199–1206. [Google Scholar] [CrossRef]
- Godhe, A.; Asplund, M.E.; Härnström, K.; Saravanan, V.; Tyagi, A.; Karunasagar, I. Quantification of Diatom and Dinoflagellate Biomasses in Coastal Marine Seawater Samples by Real-Time PCR. Appl. Environ. Microbiol. 2008, 74, 7174–7182. [Google Scholar] [CrossRef]
- Gong, J.; Dong, J.; Liu, X.; Massana, R. Extremely High Copy Numbers and Polymorphisms of the rDNA Operon Estimated from Single Cell Analysis of Oligotrich and Peritrich Ciliates. Protist 2013, 164, 369–379. [Google Scholar] [CrossRef]
- Prescott, D.M. The DNA of Ciliated Protozoa. Microbiol. Rev. 1994, 58, 233–267. [Google Scholar] [CrossRef]
- Milivojević, T.; Rahman, S.N.; Raposo, D.; Siccha, M.; Kucera, M.; Morard, R. High Variability in SSU rDNA Gene Copy Number among Planktonic Foraminifera Revealed by Single-Cell qPCR. ISME Commun. 2021, 1, 63. [Google Scholar] [CrossRef]
- Corinaldesi, C.; Barucca, M.; Luna, G.M.; Dell’Anno, A. Preservation, Origin and Genetic Imprint of Extracellular DNA in Permanently Anoxic Deep-Sea Sediments. Mol. Ecol. 2011, 20, 642–654. [Google Scholar] [CrossRef]
- Pawlowski, J.; Christen, R.; Lecroq, B.; Bachar, D.; Shahbazkia, H.R.; Amaral-Zettler, L.; Guillou, L. Eukaryotic Richness in the Abyss: Insights from Pyrotag Sequencing. PLoS ONE 2011, 6, e18169. [Google Scholar] [CrossRef]
- Guardiola, M.; Wangensteen, O.S.; Taberlet, P.; Coissac, E.; Uriz, M.J.; Turon, X. Spatio-Temporal Monitoring of Deep-Sea Communities Using Metabarcoding of Sediment DNA and RNA. PeerJ 2016, 4, e2807. [Google Scholar] [CrossRef]
- Brandt, A.; Gooday, A.J.; Brandão, S.N.; Brix, S.; Brökeland, W.; Cedhagen, T.; Choudhury, M.; Cornelius, N.; Danis, B.; De Mesel, I.; et al. First Insights into the Biodiversity and Biogeography of the Southern Ocean Deep Sea. Nature 2007, 447, 307–311. [Google Scholar] [CrossRef]
- Hohlfeld, M.; Meyer, C.; Schoenle, A.; Nitsche, F.; Arndt, H. Biogeography, Autecology, and Phylogeny of Percolomonads Based on Newly Described Species. J. Eukaryot. Microbiol. 2022, 70, e12930. [Google Scholar] [CrossRef]
- Borges, P.A.V.; Amorim, I.R.; Gabriel, R.; Cunha, R.; Martins, A.F.; Silva, L.; Costa, A.; Vieira, V. Azores. In Encyclopedia of Islands; Gillespie, R.G., Clague, D.A., Eds.; University of California Press: Berkeley, CA, USA, 2009. [Google Scholar]
- McArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Fiore-Donno, A.M.; Rixen, C.; Rippin, M.; Glaser, K.; Samolov, E.; Karsten, U.; Becker, B.; Bonkowski, M. New Barcoded Primers for Efficient Retrieval of Cercozoan Sequences in High-Throughput Environmental Diversity Surveys, with Emphasis on Worldwide Biological Soil Crusts. Mol. Ecol. Resour. 2018, 18, 229–239. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dünn, M.; Arndt, H. Distribution Patterns of Benthic Protist Communities Depending on Depth Revealed by Environmental Sequencing—From the Sublittoral to the Deep Sea. Microorganisms 2023, 11, 1664. https://doi.org/10.3390/microorganisms11071664
Dünn M, Arndt H. Distribution Patterns of Benthic Protist Communities Depending on Depth Revealed by Environmental Sequencing—From the Sublittoral to the Deep Sea. Microorganisms. 2023; 11(7):1664. https://doi.org/10.3390/microorganisms11071664
Chicago/Turabian StyleDünn, Manon, and Hartmut Arndt. 2023. "Distribution Patterns of Benthic Protist Communities Depending on Depth Revealed by Environmental Sequencing—From the Sublittoral to the Deep Sea" Microorganisms 11, no. 7: 1664. https://doi.org/10.3390/microorganisms11071664
APA StyleDünn, M., & Arndt, H. (2023). Distribution Patterns of Benthic Protist Communities Depending on Depth Revealed by Environmental Sequencing—From the Sublittoral to the Deep Sea. Microorganisms, 11(7), 1664. https://doi.org/10.3390/microorganisms11071664