


In Silico Identification and In Vitro Validation of Repurposed Compounds Targeting the RSV Polymerase




Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of the RSV Polymerase Structure
2.2. Small Molecule Procurement
2.3. Binding Affinity Measurements
2.4. Inhibition Measurements
2.5. Hydrogen Bond Analysis
2.6. In Vitro RNA Synthesis Assay Screening Inhibitors
3. Results
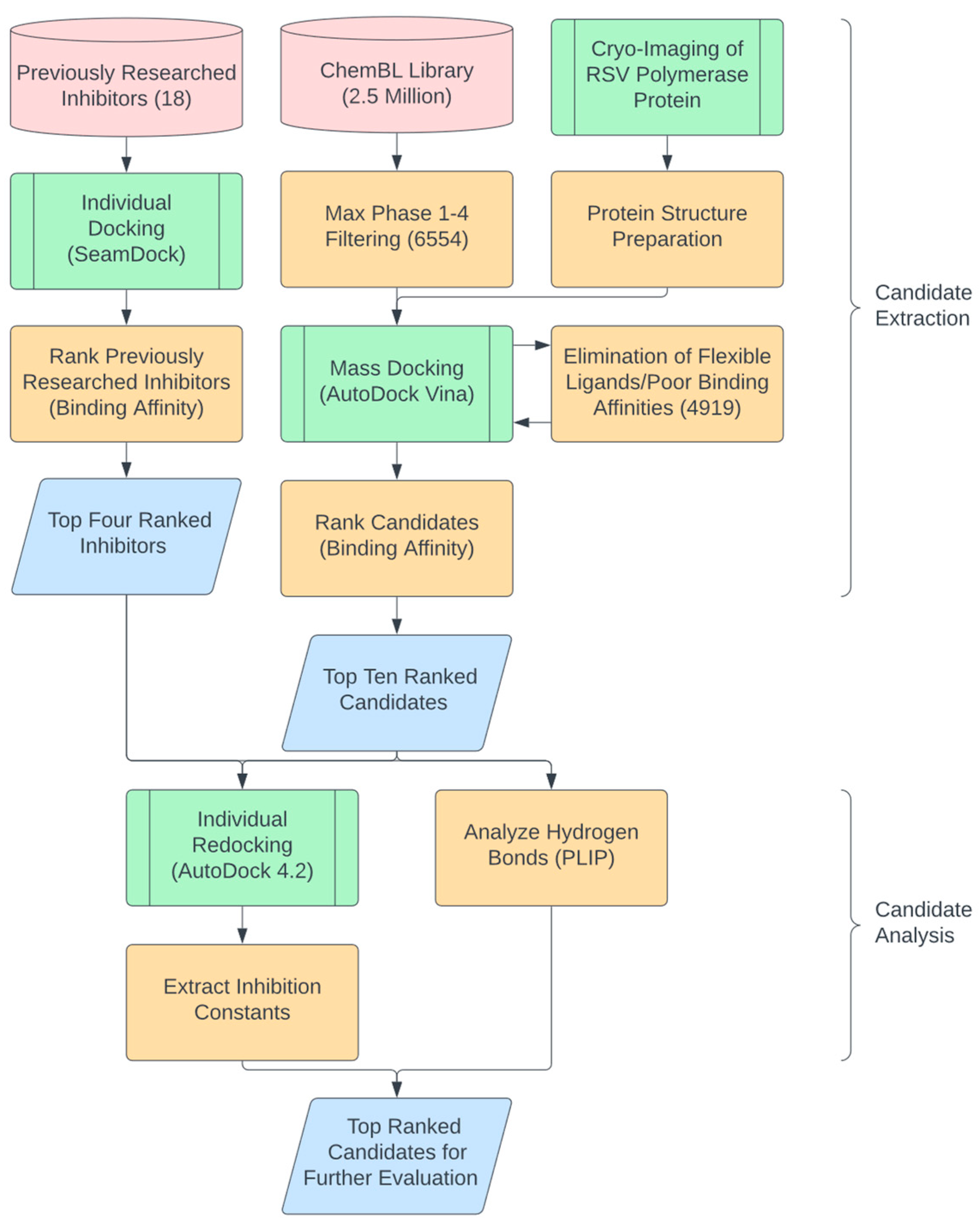
3.1. In Silico Candidate Extraction Procedure
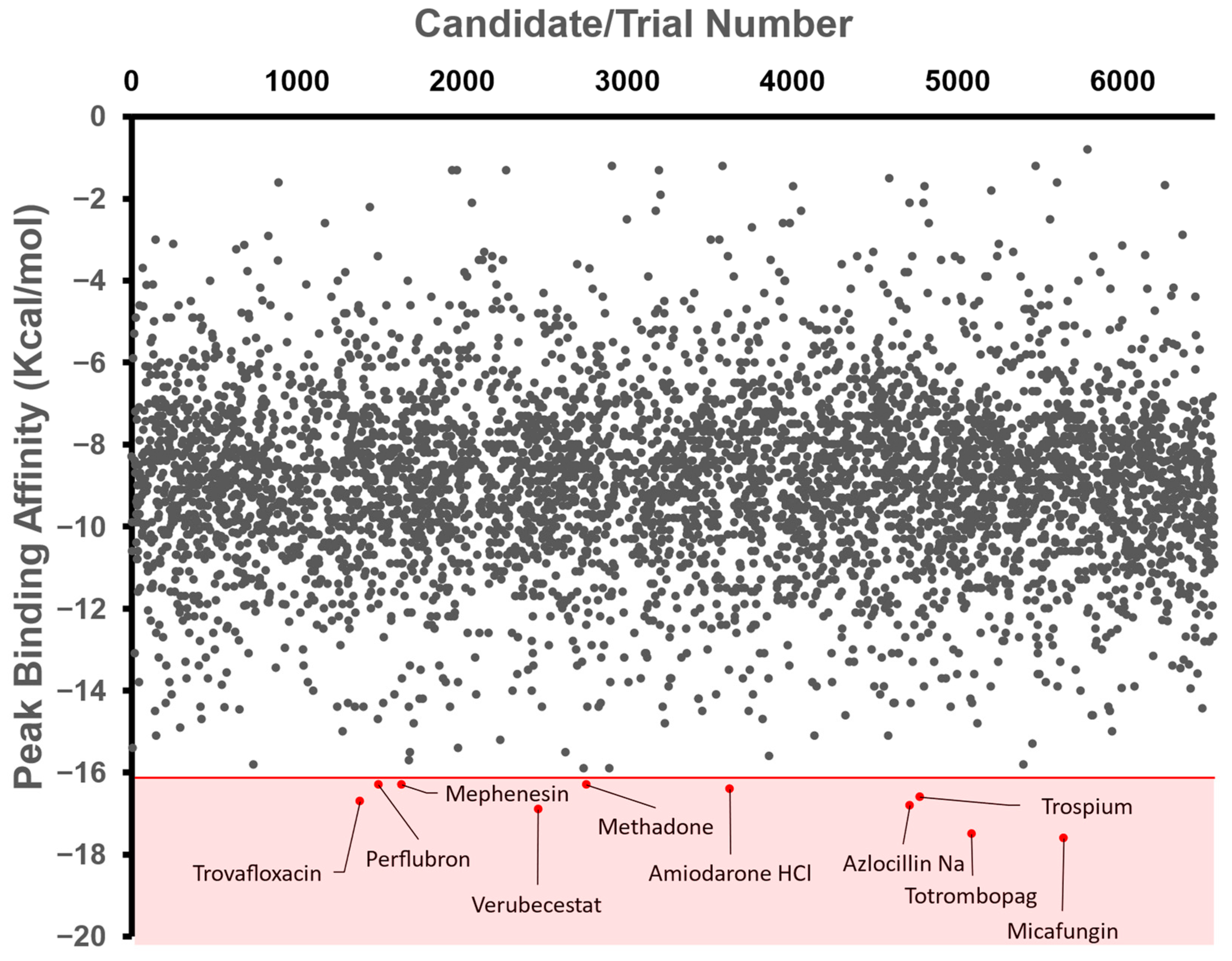
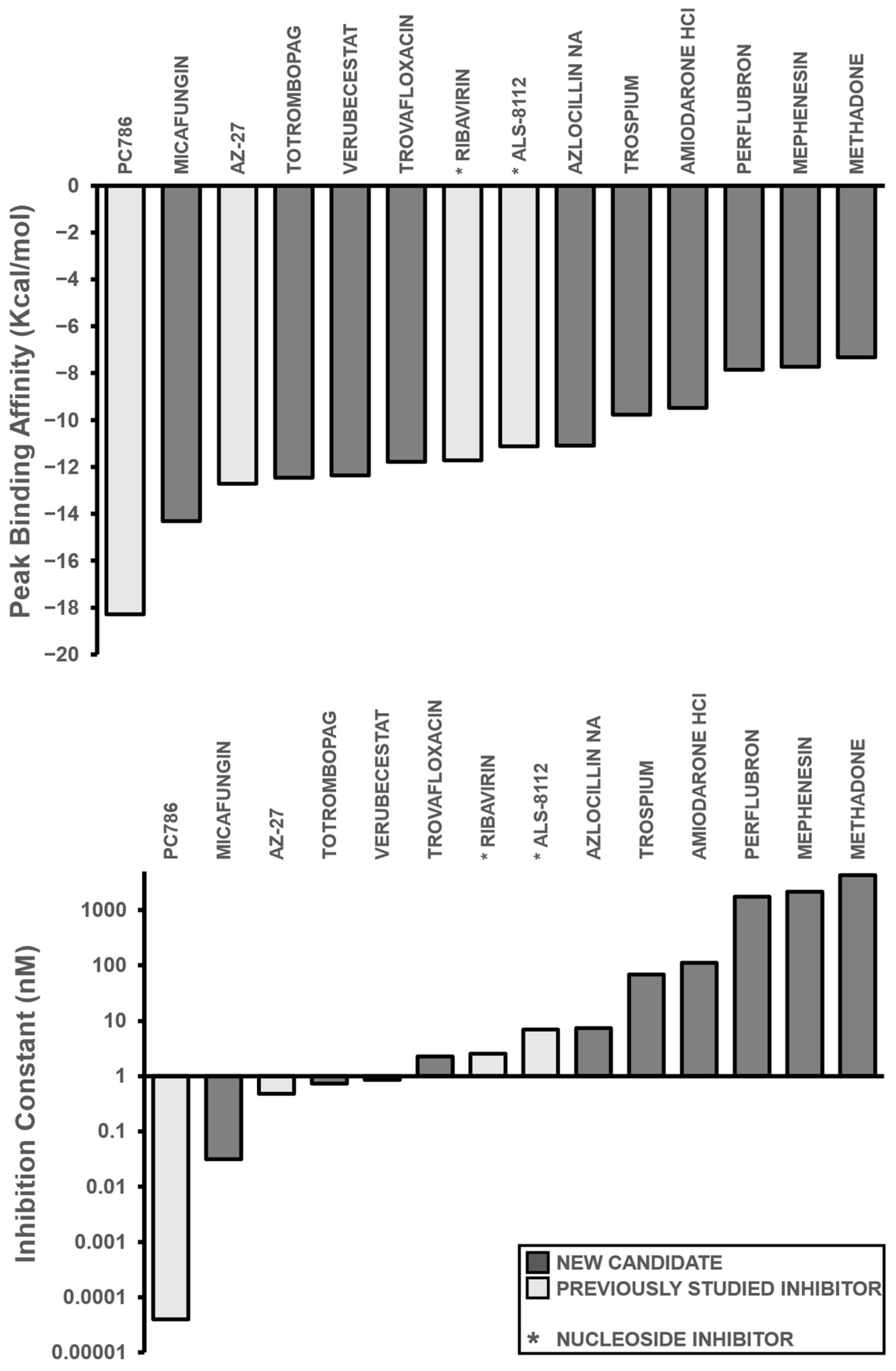
3.2. In Silico Candidate Results
3.3. Potential Molecular Actions of Top Repurposed Compound Candidates
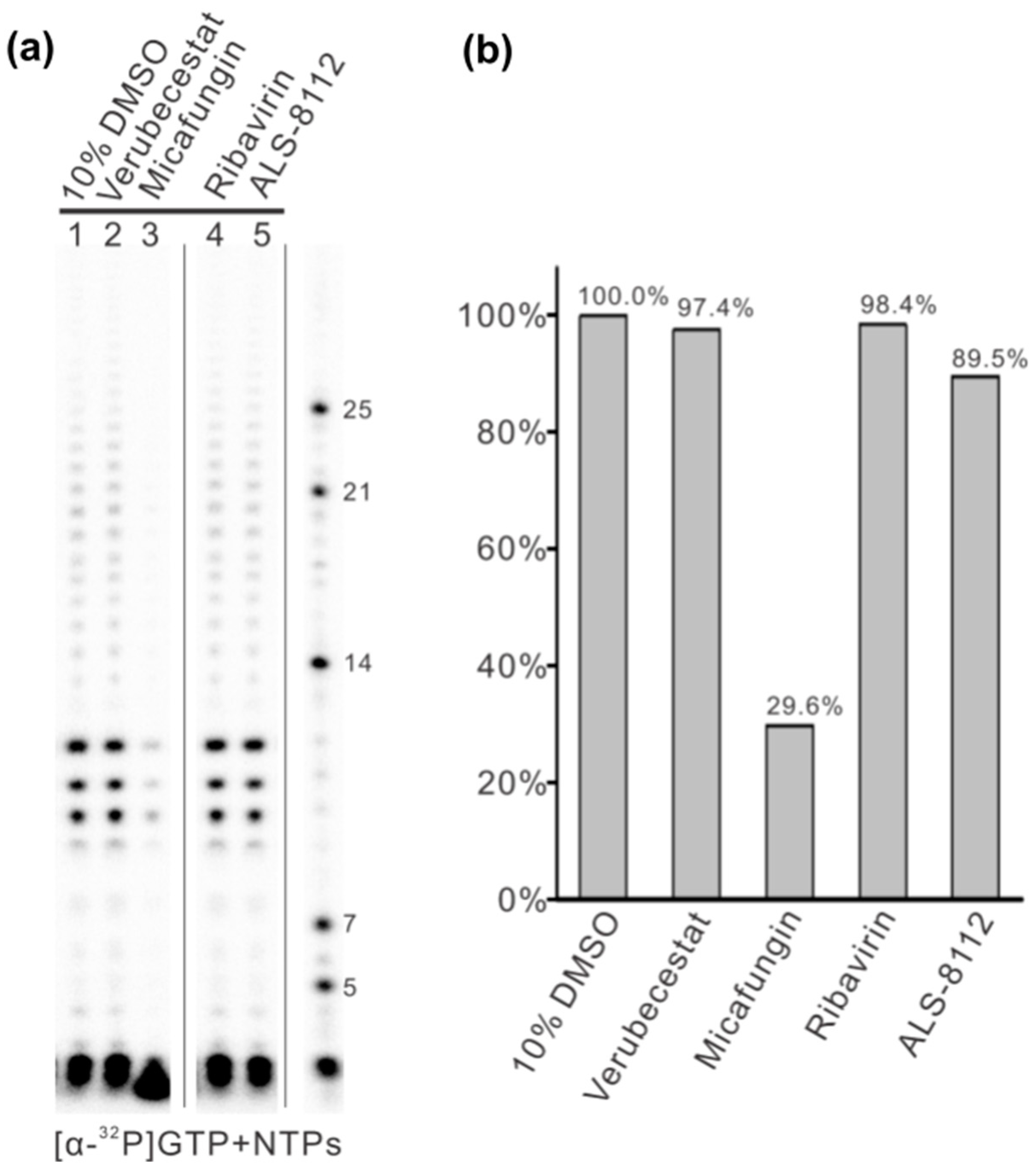
3.4. In Vitro Assay Validation
4. Discussions
5. Patents
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cowton, V.M.; McGivern, D.R.; Fearns, R. Unravelling the complexities of respiratory syncytial virus RNA synthesis. J. Gen. Virol. 2006, 87 Pt 7, 1805–1821. [Google Scholar] [CrossRef]
- Fearns, R.; Plemper, R.K. Polymerases of paramyxoviruses and pneumoviruses. Virus Res. 2017, 234, 87–102. [Google Scholar] [CrossRef]
- Noton, S.L.; Nagendra, K.; Dunn, E.F.; Mawhorter, M.E.; Yu, Q.; Fearns, R. Respiratory Syncytial Virus Inhibitor AZ-27 Differentially Inhibits Different Polymerase Activities at the Promoter. J. Virol. 2015, 89, 7786–7798. [Google Scholar] [CrossRef]
- Cox, R.; Plemper, R.K. The paramyxovirus polymerase complex as a target for next-generation anti-paramyxovirus therapeutics. Front. Microbiol. 2015, 6, 459. [Google Scholar] [CrossRef] [PubMed]
- Agamah, F.E.; Mazandu, G.K.; Hassan, R.; Bope, C.D.; Thomford, N.E.; Ghansah, A.; Chimusa, E.R. Computational/in silico methods in drug target and lead prediction. Brief. Bioinform. 2020, 21, 1663–1675. [Google Scholar] [CrossRef]
- Meewan, I.; Zhang, X.; Roy, S.; Ballatore, C.; O’Donoghue, A.J.; Schooley, R.T.; Abagyan, R. Discovery of New Inhibitors of Hepatitis C Virus NS3/4A Protease and Its D168A Mutant. ACS Omega 2019, 4, 16999–17008. [Google Scholar] [CrossRef] [PubMed]
- Underwood, M.R.; Johns, B.A.; Sato, A.; Martin, J.N.; Deeks, S.G.; Fujiwara, T. The activity of the integrase inhibitor dolutegravir against HIV-1 variants isolated from raltegravir-treated adults. J. Acquir. Immune Defic. Syndr. 2012, 61, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Gao, Y.; Roesler, C.; Rice, S.; D’Cunha, P.; Zhuang, L.; Slack, J.; Domke, M.; Antonova, A.; Romanelli, S.; et al. Cryo-EM structure of the respiratory syncytial virus RNA polymerase. Nat. Commun. 2020, 11, 368. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open Babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef] [PubMed]
- Adasme, M.F.; Linnemann, K.L.; Bolz, S.N.; Kaiser, F.; Salentin, S.; Haupt, V.J.; Schroeder, M. PLIP 2021: Expanding the scope of the protein-ligand interaction profiler to DNA and RNA. Nucleic Acids Res. 2021, 49, W530–W534. [Google Scholar] [CrossRef] [PubMed]
- Sourimant, J.; Lieber, C.M.; Aggarwal, M.; Cox, R.M.; Wolf, J.D.; Yoon, J.J.; Toots, M.; Ye, C.; Sticher, Z.; Kolykhalov, A.A.; et al. 4’-Fluorouridine is an oral antiviral that blocks respiratory syncytial virus and SARS-CoV-2 replication. Science 2022, 375, 161–167. [Google Scholar] [CrossRef]
- Chung, D.H.; Moore, B.P.; Matharu, D.S.; Golden, J.E.; Maddox, C.; Rasmussen, L.; Sosa, M.I.; Ananthan, S.; White, E.L.; Jia, F.; et al. A cell based high-throughput screening approach for the discovery of new inhibitors of respiratory syncytial virus. Virol. J. 2013, 10, 19. [Google Scholar] [CrossRef]
- Siegel, D.; Hui, H.C.; Doerffler, E.; Clarke, M.O.; Chun, K.; Zhang, L.; Neville, S.; Carra, E.; Lew, W.; Ross, B.; et al. Discovery and Synthesis of a Phosphoramidate Prodrug of a Pyrrolo[2,1-f][triazin-4-amino] Adenine C-Nucleoside (GS-5734) for the Treatment of Ebola and Emerging Viruses. J. Med. Chem. 2017, 60, 1648–1661. [Google Scholar] [CrossRef]
- Wang, G.; Deval, J.; Hong, J.; Dyatkina, N.; Prhavc, M.; Taylor, J.; Fung, A.; Jin, Z.; Stevens, S.K.; Serebryany, V.; et al. Discovery of 4′-chloromethyl-2′-deoxy-3′,5′-di-O-isobutyryl-2′-fluorocytidine (ALS-8176), a first-in-class RSV polymerase inhibitor for treatment of human respiratory syncytial virus infection. J. Med. Chem. 2015, 58, 1862–1878. [Google Scholar] [CrossRef]
- Liuzzi, M.; Mason, S.W.; Cartier, M.; Lawetz, C.; McCollum, R.S.; Dansereau, N.; Bolger, G.; Lapeyre, N.; Gaudette, Y.; Lagace, L.; et al. Inhibitors of respiratory syncytial virus replication target cotranscriptional mRNA guanylylation by viral RNA-dependent RNA polymerase. J. Virol. 2005, 79, 13105–13115. [Google Scholar] [CrossRef]
- Fearns, R.; Deval, J. New antiviral approaches for respiratory syncytial virus and other mononegaviruses: Inhibiting the RNA polymerase. Antivir. Res. 2016, 134, 63–76. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, Y.; Zheng, W.; Shang, W.; Wu, Y.; Li, N.; Xiong, J.; Jiang, H.; Shen, J.; Xiao, G.; et al. Oral remdesivir derivative VV116 is a potent inhibitor of respiratory syncytial virus with efficacy in mouse model. Signal Transduct. Target Ther. 2022, 7, 123. [Google Scholar] [CrossRef]
- Coates, M.; Brookes, D.; Kim, Y.I.; Allen, H.; Fordyce, E.A.F.; Meals, E.A.; Colley, T.; Ciana, C.L.; Parra, G.F.; Sherbukhin, V.; et al. Preclinical Characterization of PC786, an Inhaled Small-Molecule Respiratory Syncytial Virus L Protein Polymerase Inhibitor. Antimicrob. Agents Chemother. 2017, 61, e00737-17. [Google Scholar] [CrossRef] [PubMed]
- Turner, T.L.; Kopp, B.T.; Paul, G.; Landgrave, L.C.; Hayes, D., Jr.; Thompson, R. Respiratory syncytial virus: Current and emerging treatment options. Clin. Outcomes Res. 2014, 6, 217–225. [Google Scholar] [CrossRef]
- Veber, D.F.; Johnson, S.R.; Cheng, H.Y.; Smith, B.R.; Ward, K.W.; Kopple, K.D. Molecular properties that influence the oral bioavailability of drug candidates. J. Med. Chem. 2002, 45, 2615–2623. [Google Scholar] [CrossRef]
- Jarvis, B.; Figgitt, D.P.; Scott, L.J. Micafungin. Drugs 2004, 64, 969–982; discussion 983–984. [Google Scholar] [CrossRef]
- Garzon, A.M.; Mitchell, W.B. Use of Thrombopoietin Receptor Agonists in Childhood Immune Thrombocytopenia. Front. Pediatr. 2015, 3, 70. [Google Scholar] [CrossRef]
- Haria, M.; Lamb, H.M. Trovafloxacin. Drugs 1997, 54, 435–445; discussion 446. [Google Scholar] [CrossRef]
- Woolf, R.A.; Koup, J.R.; Smith, A.L.; Hilman, B.C.; Goldmann, D.; Williams-Warren, J. Pharmacokinetics of high-dose azlocillin sodium in patients with cystic fibrosis. Clin. Pharm. 1985, 4, 664–669. [Google Scholar] [PubMed]
- Chapple, C. New once-daily formulation for trospium in overactive bladder. Int. J. Clin. Pract. 2010, 64, 1535–1540. [Google Scholar] [CrossRef] [PubMed]
- Sloskey, G.E. Amiodarone: A unique antiarrhythmic agent. Clin. Pharm. 1983, 2, 330–340. [Google Scholar] [PubMed]
- Fourches, D.; Barnes, J.C.; Day, N.C.; Bradley, P.; Reed, J.Z.; Tropsha, A. Cheminformatics analysis of assertions mined from literature that describe drug-induced liver injury in different species. Chem. Res. Toxicol. 2010, 23, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Crankshaw, D.P.; Raper, C. Mephenesin, methocarbamol, chlordiazepoxide and diazepam: Actions on spinal reflexes and ventral root potentials. Br. J. Pharmacol. 1970, 38, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Durrani, M.; Bansal, K. Methadone. In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2023. [Google Scholar]
- Tiong-Yip, C.L.; Aschenbrenner, L.; Johnson, K.D.; McLaughlin, R.E.; Fan, J.; Challa, S.; Xiong, H.; Yu, Q. Characterization of a respiratory syncytial virus L protein inhibitor. Antimicrob. Agents Chemother. 2014, 58, 3867–3873. [Google Scholar] [CrossRef] [PubMed]
- Deval, J.; Hong, J.; Wang, G.; Taylor, J.; Smith, L.K.; Fung, A.; Stevens, S.K.; Liu, H.; Jin, Z.; Dyatkina, N.; et al. Molecular Basis for the Selective Inhibition of Respiratory Syncytial Virus RNA Polymerase by 2′-Fluoro-4′-Chloromethyl-Cytidine Triphosphate. PLoS Pathog. 2015, 11, e1004995. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, H.; Banks, G.; Smith, R. Ribavirin: A clinical overview. Eur. J. Epidemiol. 1986, 2, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekar, P.H.; Sobel, J.D. Micafungin: A new echinocandin. Clin. Infect. Dis. 2006, 42, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Oblak, A.L.; Cope, Z.A.; Quinney, S.K.; Pandey, R.S.; Biesdorf, C.; Masters, A.R.; Onos, K.D.; Haynes, L.; Keezer, K.J.; Meyer, J.A.; et al. Prophylactic evaluation of verubecestat on disease- and symptom-modifying effects in 5XFAD mice. Alzheimers Dement. 2022, 8, e12317. [Google Scholar] [CrossRef] [PubMed]
- Cosconati, S.; Forli, S.; Perryman, A.L.; Harris, R.; Goodsell, D.S.; Olson, A.J. Virtual Screening with AutoDock: Theory and Practice. Expert Opin. Drug Discov. 2010, 5, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, R.; Smith, J.C. Speed vs Accuracy: Effect on Ligand Pose Accuracy of Varying Box Size and Exhaustiveness in AutoDock Vina. Mol. Inform. 2023, 42, e2200188. [Google Scholar] [CrossRef]
- Kim, C.; Kang, H.; Kim, D.E.; Song, J.H.; Choi, M.; Kang, M.; Lee, K.; Kim, H.S.; Shin, J.S.; Jeong, H.; et al. Antiviral activity of micafungin against enterovirus 71. Virol. J. 2016, 13, 99. [Google Scholar] [CrossRef]
- Lu, J.W.; Chen, Y.C.; Huang, C.K.; Lin, K.C.; Ho, Y.J. Synergistic in-vitro antiviral effects of combination treatment using anidulafungin and T-1105 against Zika virus infection. Antiviral Res. 2021, 195, 105188. [Google Scholar] [CrossRef]
- Chen, Y.C.; Lu, J.W.; Yeh, C.T.; Lin, T.Y.; Liu, F.C.; Ho, Y.J. Micafungin Inhibits Dengue Virus Infection through the Disruption of Virus Binding, Entry, and Stability. Pharmaceuticals 2021, 14, 338. [Google Scholar] [CrossRef]
- Nakajima, S.; Ohashi, H.; Akazawa, D.; Torii, S.; Suzuki, R.; Fukuhara, T.; Watashi, K. Antiviral Activity of Micafungin and Its Derivatives against SARS-CoV-2 RNA Replication. Viruses 2023, 15, 452. [Google Scholar] [CrossRef] [PubMed]
- Liang, B. Structures of the Mononegavirales Polymerases. J. Virol. 2020, 94, e00175-20. [Google Scholar] [CrossRef] [PubMed]
- Sugita, Y.; Matsunami, H.; Kawaoka, Y.; Noda, T.; Wolf, M. Cryo-EM structure of the Ebola virus nucleoprotein-RNA complex at 3.6 A resolution. Nature 2018, 563, 137–140. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Chemical Structure | Inhibition Constant (nM) | Peak Binding Affinity (kcal/mol) | Number of Hydrogen Bonds | Nucleoside/ Non-Nucleoside |
|---|---|---|---|---|---|
| PC786 * |  | 39.9 × 10−6 | −18.28 | 6 | Non-Nucleoside |
| MICAFUNGIN |  | 0.0317 | −14.32 | 0 | Non-Nucleoside |
| AZ-27 * |  | 0.47997 | −12.71 | 5 | Non-Nucleoside |
| TOTROMBOPAG |  | 0.73929 | −12.46 | 6 | Non-Nucleoside |
| VERUBECESTAT |  | 0.85487 | −12.37 | 5 | Non-Nucleoside |
| TROVAFLOXACIN |  | 2.27 | −11.79 | 7 | Non-Nucleoside |
| RIBAVIRIN * |  | 2.56 | −11.72 | 6 | Nucleoside |
| ALS-8112 * |  | 6.98 | −11.13 | 1 | Nucleoside |
| AZLOCILLIN NA |  | 7.47 | −11.09 | 1 | Non-Nucleoside |
| TROSPIUM |  | 68.41 | −9.77 | 5 | Non-Nucleoside |
| AMIODARONE HCL |  | 111.15 | −9.49 | 5 | Non-Nucleoside |
| PERFLUBRON |  | 1760 | −7.85 | 1 | Non-Nucleoside |
| MEPHENESIN |  | 2160 | −7.73 | 3 | Non-Nucleoside |
| METHADONE |  | 4300 | −7.32 | 9 | Non-Nucleoside |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, E.; Park, S.; Calderon, J.; Cao, D.; Liang, B. In Silico Identification and In Vitro Validation of Repurposed Compounds Targeting the RSV Polymerase. Microorganisms 2023, 11, 1608. https://doi.org/10.3390/microorganisms11061608
Xu E, Park S, Calderon J, Cao D, Liang B. In Silico Identification and In Vitro Validation of Repurposed Compounds Targeting the RSV Polymerase. Microorganisms. 2023; 11(6):1608. https://doi.org/10.3390/microorganisms11061608
Chicago/Turabian StyleXu, Eric, Seohyun Park, Juan Calderon, Dongdong Cao, and Bo Liang. 2023. "In Silico Identification and In Vitro Validation of Repurposed Compounds Targeting the RSV Polymerase" Microorganisms 11, no. 6: 1608. https://doi.org/10.3390/microorganisms11061608
APA StyleXu, E., Park, S., Calderon, J., Cao, D., & Liang, B. (2023). In Silico Identification and In Vitro Validation of Repurposed Compounds Targeting the RSV Polymerase. Microorganisms, 11(6), 1608. https://doi.org/10.3390/microorganisms11061608