
Effects of Different Yeast Selenium Levels on Rumen Fermentation Parameters, Digestive Enzyme Activity and Gastrointestinal Microflora of Sika Deer during Antler Growth


Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Material and Method
2.3. Sample Collection and Analysis
2.4. Statistical Analysis
3. Results
3.1. Digestibility of Fiber in Sika Deer under Different Selenium Levels
3.2. Fermentation Parameters of Rumen Fluid of Sika Deer at Velvet Antler Growth Stage under Different Selenium Levels
3.3. Rumen Digestive Enzyme Activities of Sika Deer at Velvet Antler Growth Stage under Different Selenium Levels
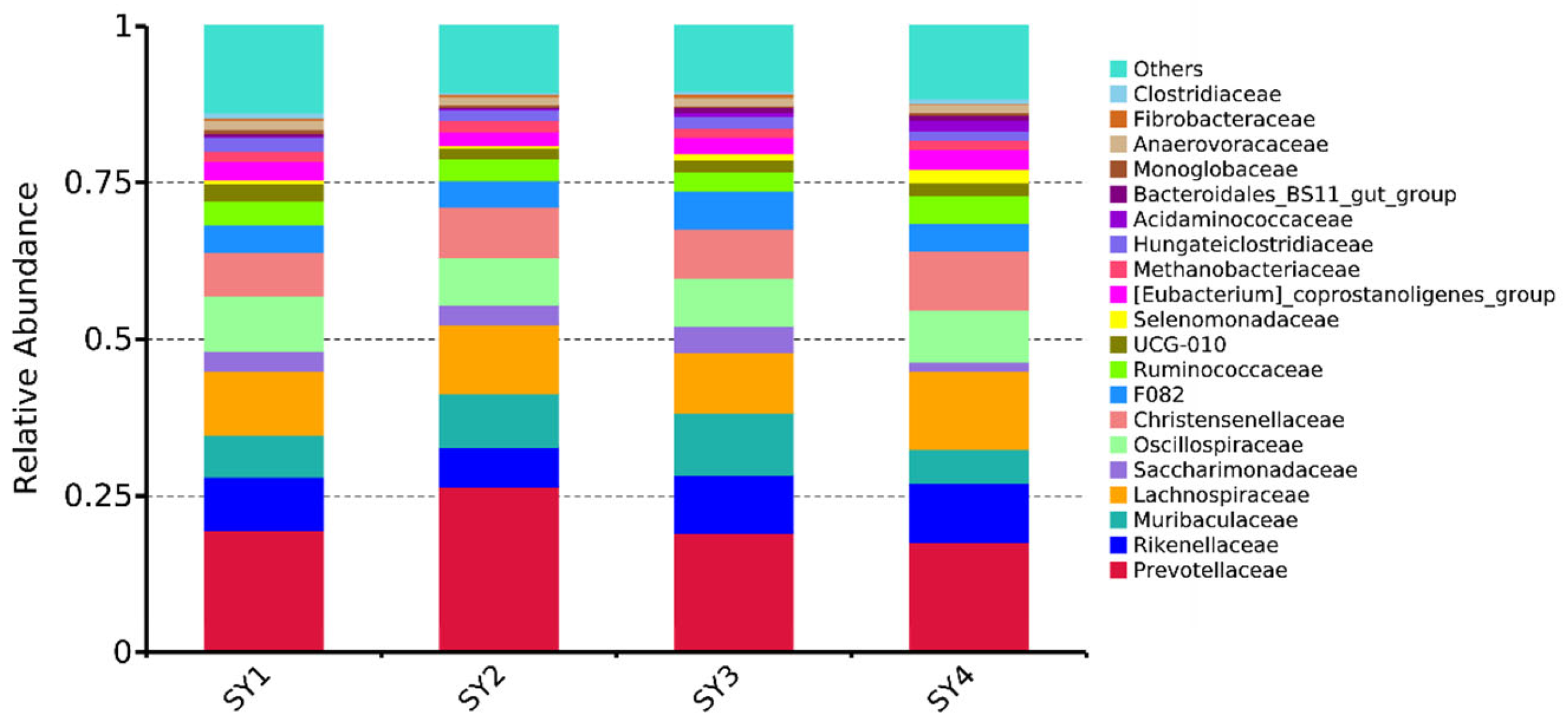
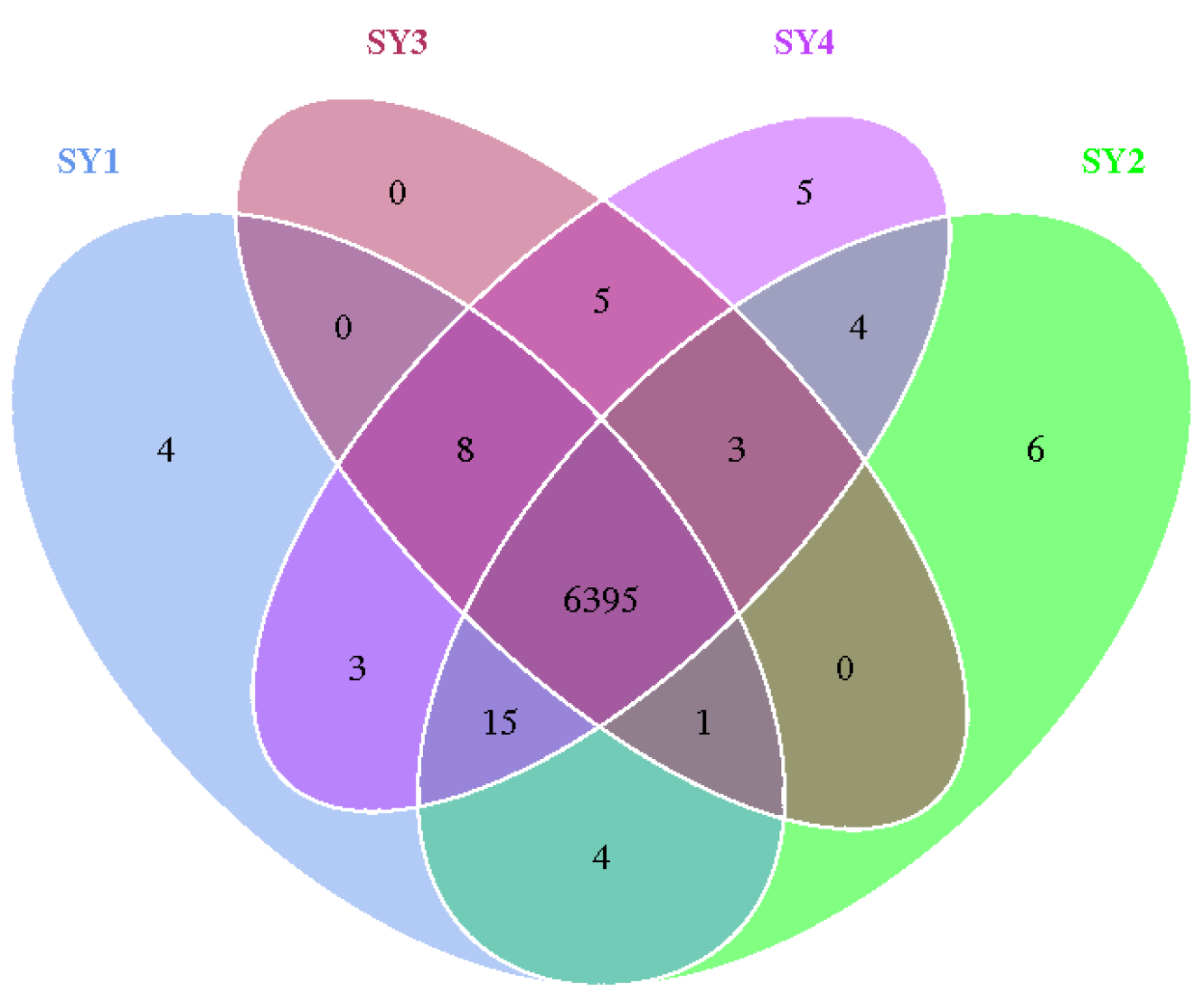
3.4. Rumen Microflora Relative Abundance of Sika Deer at Velvet Antler Growth Stage under Different Selenium Levels
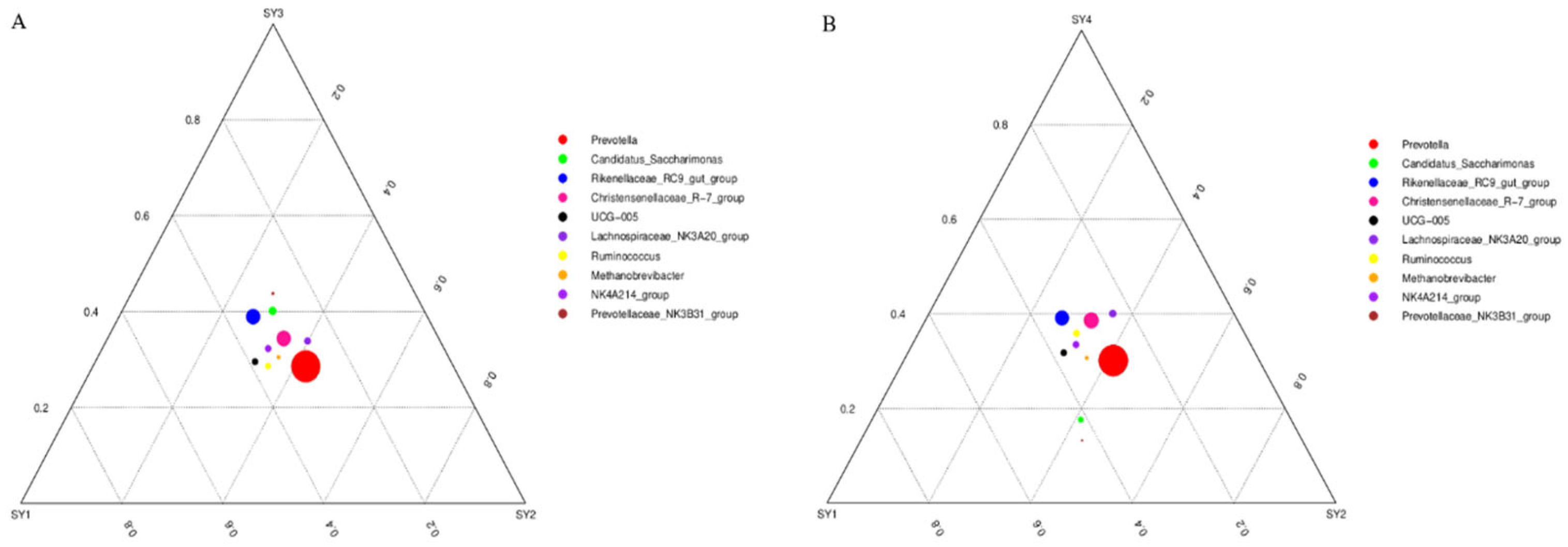
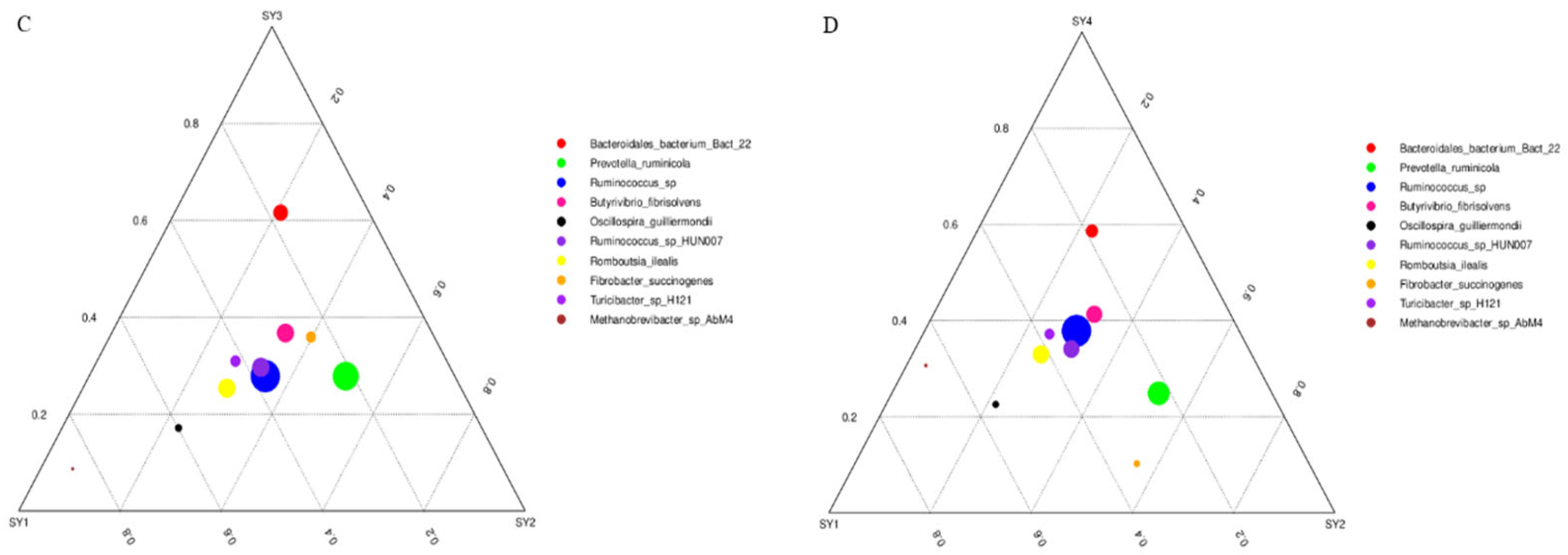
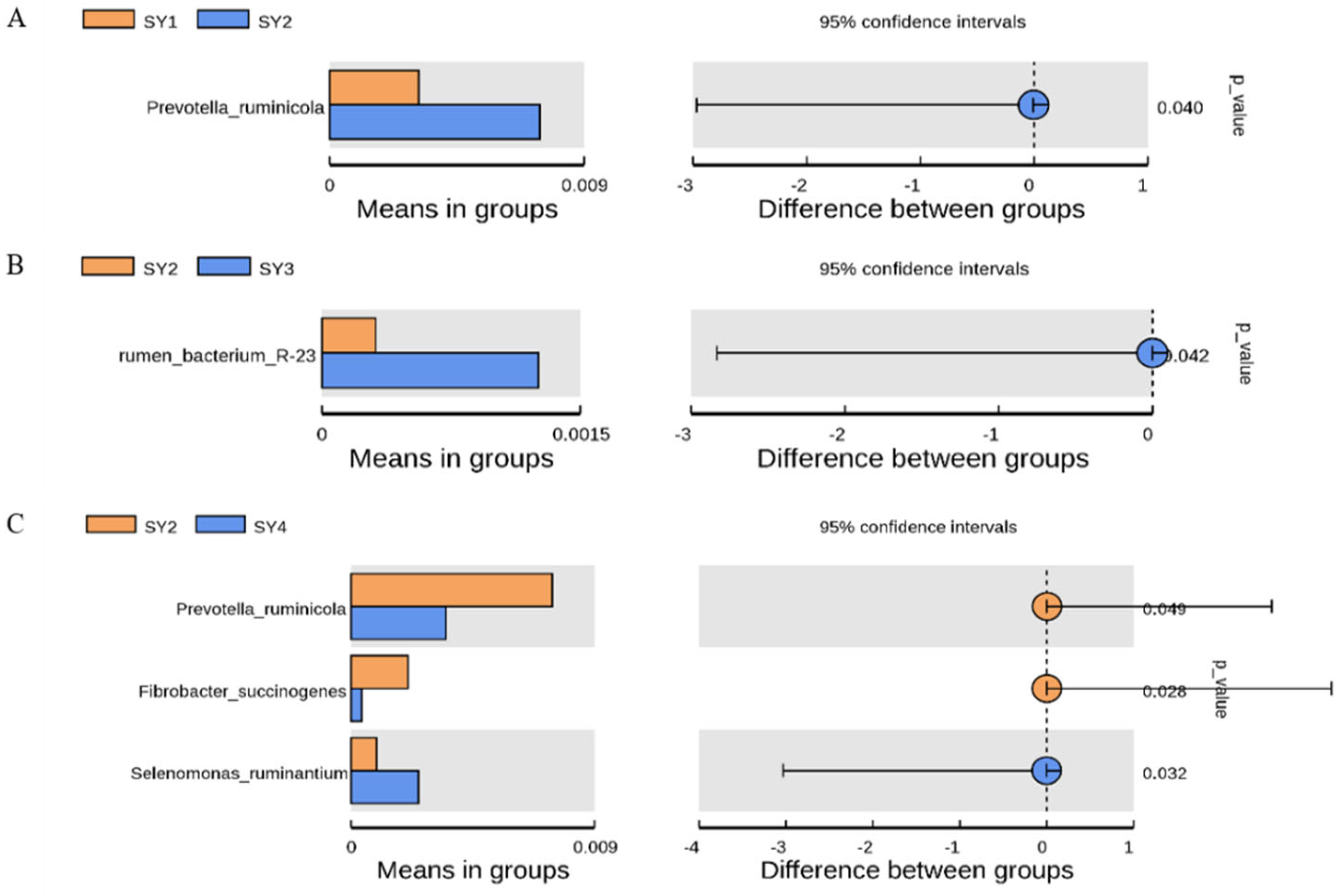
3.5. Analysis on the Difference of Dominant Strain Species under Different Selenium Levels
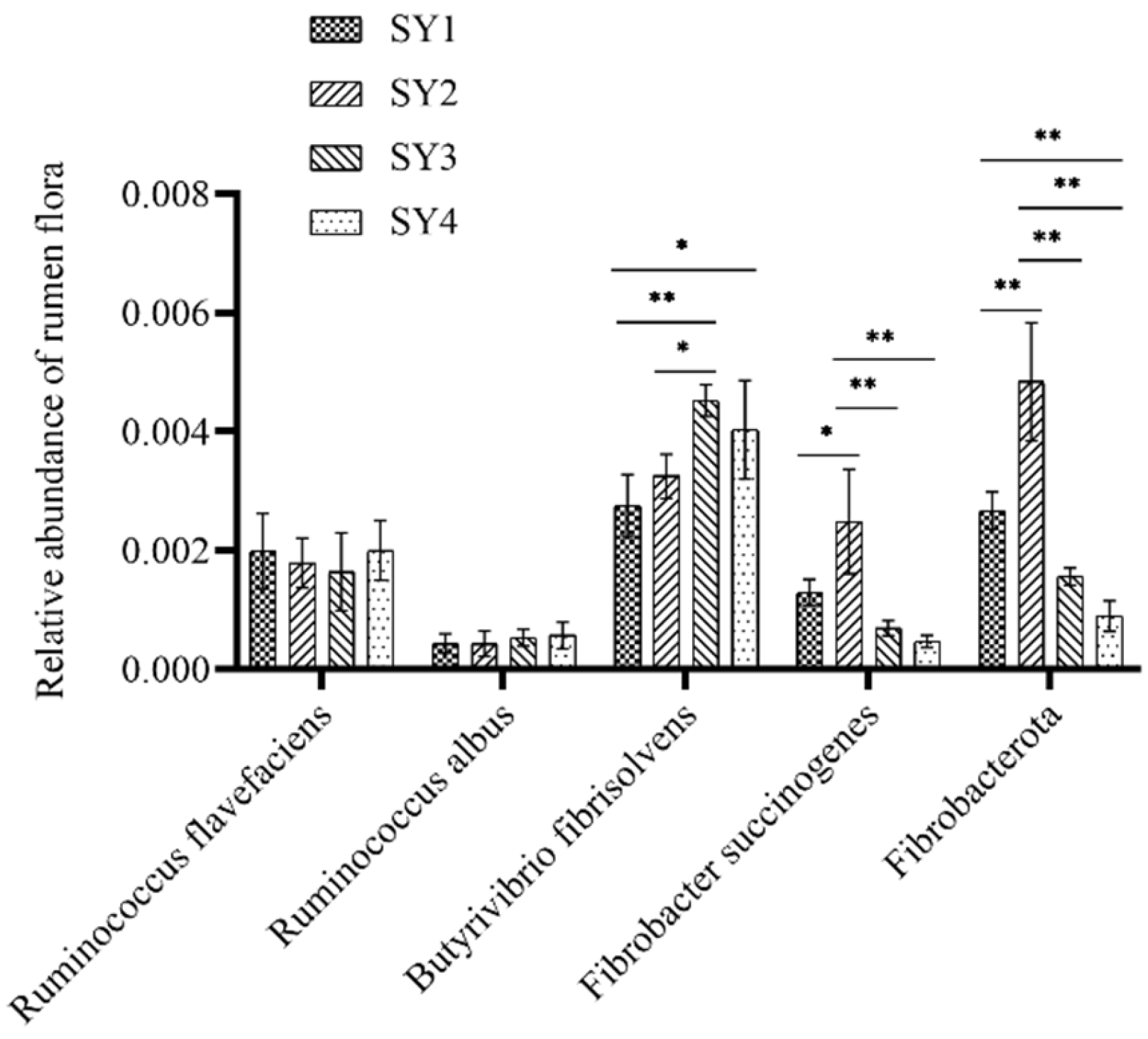
3.6. Changes in the Abundance of Rumen Fiber Degradation-Related Bacteria in Sika Deer during Velvet Antler Growth with Different Selenium Levels
3.7. Analysis of Rumen Differential Bacteria of Sika Deer at Velvet Antler Growth Stage under Different Selenium Levels
3.8. Changes of Rumen Microflora Abundance of Sika Deer during Velvet Antler Growth with Environmental Factors Intervention
3.9. Screening Differential Genes and Proteins in Rumen Microorganisms
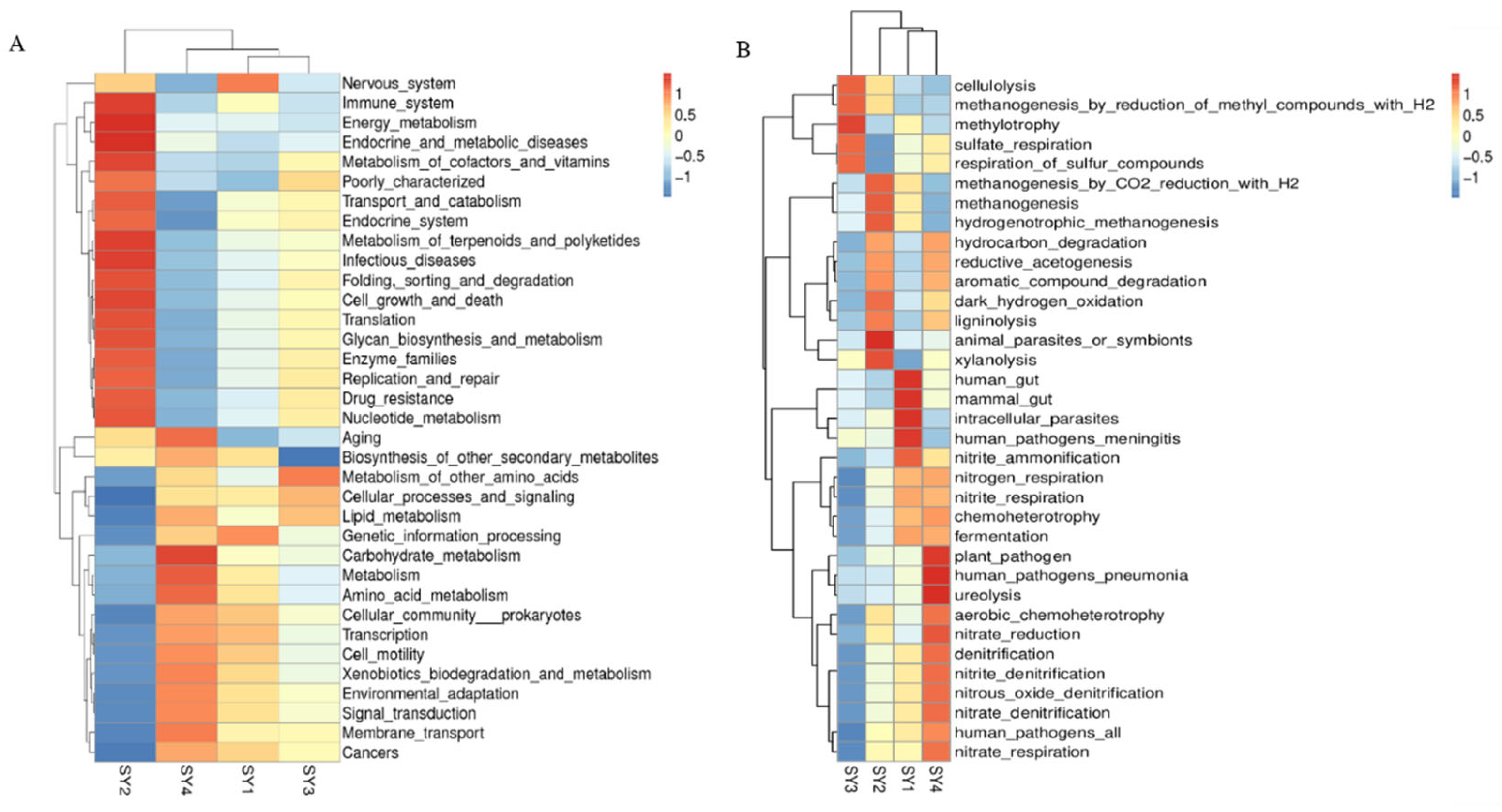
3.10. Prediction of Rumen Microflora Function of Sika Deer at Antler Growth Stage under Different Selenium Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wei, J.; Wang, J.; Liu, W.; Zhang, K.; Sun, P. Short communication: Effects of different selenium supplements on rumen fermentation and apparent nutrient and selenium digestibility of mid-lactation dairy cows. J. Dairy Sci. 2019, 102, 3131–3135. [Google Scholar] [CrossRef]
- Wang, C.; Liu, Q.; Yang, W.Z.; Dong, Q.; Yang, X.M.; He, D.C.; Zhang, P.; Dong, K.H.; Huang, Y.X. Effects of selenium yeast on rumen fermentation, lactation performance and feed digestibilities in lactating dairy cows. Livest Sci. 2009, 126, 239–244. [Google Scholar] [CrossRef]
- Wu, X.X. Effect of Sources and Levels of Selenium on in Vitro Ruminal Fermentation and Nutrient Degradation of Dairy Cows; Inner Mongolia Agricultural University: Hohhot, China, 2012. [Google Scholar]
- Hendawy, A.O.; Sugimura, S.; Sato, K.; Mansour, M.M.; Abd El-Aziz, A.H.; Samir, H.; Islam, A.; Bostami, A.B.M.R.; Mandour, A.S.; Elfadadny, A. Effects of Selenium Supplementation on Rumen Microbiota, Rumen Fermentation, and Apparent Nutrient Digestibility of Ruminant Animals: A Review. Fermentation 2022, 8, 4. [Google Scholar] [CrossRef]
- Faixová, Z.; Piešová, E.; Maková, Z.; Cobanova, K.; Faix, S. Effect of dietary supplementation with selenium-enriched yeast or sodium selenite on ruminal enzyme activities and blood chemistry in sheep. Acta. Vet. Brno. 2016, 85, 185–194. [Google Scholar] [CrossRef]
- Oberson, J.L.; Probst, S.; Schlegel, P. Magnesium absorption as influenced by the rumen passage kinetics in lactating dairy cows fed modified levels of fibre and protein. Animal 2019, 13, 1412–1420. [Google Scholar] [CrossRef]
- Turło, J.; Gutkowska, B.; Gajzlerska, W.; Dawidowski, M. Biological availability and preliminary selenium speciation in selenium-enriched mycelium of Lentinula edodes (Berk.). Food Biotechnol. 2011, 25, 16–29. [Google Scholar] [CrossRef]
- Van Soest, P.J.; McQueen, R.W. The chemistry and estimation of fibre. Proc. Nutr. Soc. 1973, 32, 123–130. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583e97. [Google Scholar] [CrossRef]
- A.O.A.C. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1990. [Google Scholar]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Algina, J.; Keselman, H.J. Comparing squared multiple correlation coefficients: Examination of a confidence interval and a test significance. Psychol. Methods 1999, 4, 76–83. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, G.; Zhang, P.; Ma, X.; Li, F.; Zhang, H.; Tao, X.; Ye, J.; Nabi, M. Rumen fluid fermentation for enhancement of hydrolysis and acidification of grass clipping. J. Environ. Manag. 2018, 220, 142–148. [Google Scholar] [CrossRef]
- Alimohamady, R.; Aliarabi, H.; Bahari, A.; Dezfoulian, A.H. Influence of different amounts and sources of selenium supplementation on performance, some blood parameters, and nutrient digestibility in lambs. Biol. Trace Elem. Res. 2013, 154, 45–54. [Google Scholar] [CrossRef]
- Quigley, J.D.; Hill, T.M.; Dennis, T.S.; Suarez-Mena, F.X.; Schlotterbeck, R.L. Effects of feeding milk replacer at 2 rates with pelleted, low-starch or texturized, high-starch starters on calf performance and digestion. J. Dairy Sci. 2018, 101, 5937–5948. [Google Scholar] [CrossRef]
- Shi, R.; Dong, S.; Fu, Y.; Wang, Y.; Cao, Z.; Li, S.; Zhao, G. Effects of Neutral Detergent Fiber Levels in Diet on Milk Performance, Nutrient Apparent Digestibility and Serum Indices of Dairy Cows during Peak Lactation Period. Anim. Nutr. 2015, 27, 2414–2422. [Google Scholar]
- Li, W.; Zhang, X.; Feng, Y.; Cui, Z. Research Advances on Application of Trace Element Selenium in Ruminants. Anim. Nutr. 2020, 32, 1499–1507. [Google Scholar]
- Mihaliková, K.; Gresáková, L.; Boldizárová, K.; Faix, S.; Leng, L.; Kisidayová, S. The effects of organic selenium supplementation on the rumen ciliate population in sheep. Folia Microbiol. 2005, 50, 353–356. [Google Scholar] [CrossRef]
- Xun, W.; Shi, L.; Yue, W.; Zhang, C.; Ren, Y.; Liu, Q. Effect of high-dose nano-selenium and selenium-yeast on feed digestibility, rumen fermentation, and purine derivatives in sheep. Biol. Trace Elem. Res. 2012, 150, 130–136. [Google Scholar] [CrossRef]
- Zhang, L.; Yao, J.; Tan, B.; Xu, Z. Effects of Different Supplementary Concentration Se-yeast in Dairy Ration on Digestibility. China Anim. Husb. Vet. Med. 2007, 8, 66–68. [Google Scholar]
- Zhang, L.; Xing, D.; Jiang, Q.; Sun, R.; Ren, N. Properties of cellulose-degrading and hydrogen-producing bacteria flora in rumen fluid at different pH. J. Dalian Polytech. Univ. 2017, 36, 328–332. [Google Scholar]
- Harshaw, L.T.; Larkin, I.V.; Staples, C.R.; Scott, K.C.; Hill, R.C. In Vivo Apparent Digestibility of Fiber in Florida Manatees (Trichechus manatus latirostris) Under Human Care. Aquat. Mamm. 2019, 45, 513–524. [Google Scholar] [CrossRef]
- Shi, L.; Ren, Y.; Zhang, C.; Yue, W.; Lei, F. Effects of maternal dietary selenium (Se-enriched yeast) on growth performance, antioxidant status and haemato-biochemical parameters of their male kids in Taihang Black Goats. Anim. Feed. Sci. Technol. 2017, 231, 67–75. [Google Scholar] [CrossRef]
- Sun, X.; Li, Q.; Cao, Y.; Yan, J.; Liu, T.; Zhang, Y.; Li, J.; Gao, Y.; Li, S.; Xie, P.; et al. Effects of Dietary Addition of N-carbamylglutamate on Growth Performance, Rumen Fermentation, Rumen Microflora and Methane Emission of Holstein Bulls. Acta Vet. Zootech. Sin. 2020, 51, 1903–1912. [Google Scholar]
- Yang, W.Z.; Beauchemin, K.A.; Rode, L.M. Effects of grain processing, forage to concentrate ratio, and forage particle size on rumen pH and digestion by dairy cows. J. Dairy Sci. 2001, 84, 2203–2216. [Google Scholar] [CrossRef]
- Del Bianco Benedeti, P.; Silva, B.C.; Pacheco, M.V.C.; Serão, N.V.L.; Carvalho Filho, I.; Lopes, M.M.; Marcondes, M.I.; Mantovani, H.C.; Valadares Filho, S.C.; Detmann, E.; et al. Effects of grain processing methods on the expression of genes involved in volatile fatty acid transport and pH regulation, and keratinization in rumen epithelium of beef cattle. PLoS ONE 2018, 13, e0198963. [Google Scholar] [CrossRef]
- Genther, O.N.; Hansen, S.L. The effect of trace mineral source and concentration on ruminal digestion and mineral solubility. J. Dairy Sci. 2015, 98, 566–573. [Google Scholar] [CrossRef]
- Cui, X.; Wang, Z.; Tan, Y.; Chang, S.; Zheng, H.; Wang, H.; Yan, T.; Guru, T.; Hou, F. Selenium Yeast Dietary Supplement Affects Rumen Bacterial Population Dynamics and Fermentation Parameters of Tibetan Sheep (Ovis aries) in Alpine Meadow. Front. Microbiol. 2021, 12, 663945. [Google Scholar] [CrossRef]
- Čobanová, K.; Faix, Š.; Plachá, I.; Mihaliková, K.; Váradyová, Z.; Kišidayová, S.; Grešáková, Ľ. Effects of Different Dietary Selenium Sources on Antioxidant Status and Blood Phagocytic Activity in Sheep. Biol. Trace Elem. Res. 2017, 175, 339–346. [Google Scholar] [CrossRef]
- Serra, A.B.; Nakamura, K.; Matsui, T.; Harumoto, T.; Fujihara, T. Inorganic selenium for sheep. II. Its influence on rumen bacterial yield, volatile fatty acid production and total tract digestion of timothy hay. Asian Australas. J. Anim. Sci. 1994, 7, 91–96. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, C.; Liu, Q.; Guo, G.; Huo, W.; Zhang, Y.; Pei, C.; Zhang, S.; Zhang, J. Effects of sodium selenite addition on ruminal fermentation, microflora and urinary excretion of purine derivatives in Holstein dairy bulls. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1719–1726. [Google Scholar] [CrossRef]
- Mcdonald, P. Animal Nutrition, 7th ed.; Pearson Education: London, UK, 2011. [Google Scholar]
- Dijkstra, J.; Tamminga, S. Simulation of the effects of diet on the contribution of rumen protozoa to degradation of fibre in the rumen. Br. J. Nutr. 1995, 74, 617–634. [Google Scholar] [CrossRef]
- Wang, Y.; Mcallister, T.A. Rumen Microbes, Enzymes and Feed Digestion-A Review. Asian Australas. J. Anim. Sci. 2002, 15, 1659–1676. [Google Scholar] [CrossRef]
- Moss, A.R.; Jouany, J.P.; Newbold, J. Methane production by ruminants: Its contribution to global warming. Ann. Zootech. 2000, 49, 231–253. [Google Scholar] [CrossRef]
- Cheng, K.; Wang, C.; Zhang, G.; Du, H.; Wu, Z.; Liu, Q.; Guo, G.; Huo, W.; Zhang, J.; Chen, L.; et al. Effects of betaine and rumen-protected folic acid supplementation on lactation performance, nutrient digestion, rumen fermentation and blood metabolites in dairy cows. Anim. Feed. Sci. Technol. 2020, 262, 114445. [Google Scholar] [CrossRef]
- Wereszka, K.; Michałowski, T. The ability of the rumen ciliate protozoan Diploplastron affine to digest and ferment starch. Folia Microbiol. 2012, 57, 375–377. [Google Scholar] [CrossRef]
- Grabber, J.H. How do lignin composition, structure, and cross-linking affect degradability? A review of cell wall model studies. Crop Sci. 2005, 45, 820–831. [Google Scholar] [CrossRef]
- Kovatcheva-Datchary, P.; Nilsson, A.; Akrami, R.; Lee, Y.S.; De Vadder, F.; Arora, T.; Hallen, A.; Martens, E.; Björck, I.; Bäckhed, F. Dietary Fiber-Induced Improvement in Glucose Metabolism Is Associated with Increased Abundance of Prevotella. Cell Metab. 2015, 22, 971–982. [Google Scholar] [CrossRef]
- Aakko, J.; Pietilä, S.; Toivonen, R.; Rokka, A.; Mokkala, K.; Laitinen, K.; Elo, L.; Hänninen, A. A carbohydrate-active enzyme (CAZy) profile links successful metabolic specialization of Prevotella to its abundance in gut microbiota. Sci. Rep. 2020, 10, 12411. [Google Scholar] [CrossRef]
- Dodd, D.; Kocherginskaya, S.A.; Spies, M.A.; Beery, K.E.; Abbas, C.A.; Mackie, R.I.; Cann, I.K. Biochemical analysis of a beta-D-xylosidase and a bifunctional xylanase-ferulic acid esterase from a xylanolytic gene cluster in Prevotella ruminicola 23. J. Bacteriol. 2009, 191, 3328–3338. [Google Scholar] [CrossRef]
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef]
- Krause, D.O.; Denman, S.E.; Mackie, R.I.; Morrison, M.; Rae, A.L.; Attwood, G.T.; McSweeney, C.S. Opportunities to improve fiber degradation in the rumen: Microbiology, ecology, and genomics. FEMS Microbiol. Rev. 2003, 27, 663–693. [Google Scholar] [CrossRef]
- Yoshida, S.; Mackie, R.I.; Cann, I.K. Biochemical and domain analyses of FSUAxe6B, a modular acetyl xylan esterase, identify a unique carbohydrate binding module in Fibrobacter succinogenes S85. J. Bacteriol. 2010, 192, 483–493. [Google Scholar] [CrossRef]
- Raut, M.P.; Couto, N.; Karunakaran, E.; Biggs, C.A.; Wright, P.C. Deciphering the unique cellulose degradation mechanism of the ruminal bacterium Fibrobacter succinogenes S85. Sci. Rep. 2019, 9, 16542. [Google Scholar] [CrossRef]
- Bryant, M.P.; Doetsch, R.N. A study of actively cellulolytic rod-shaped bacteria of the bovine rumen. J. Dairy Sci. 1954, 37, 1176–1183. [Google Scholar] [CrossRef]
- Nouaille, R.; Matulova, M.; Pätoprstý, V.; Delort, A.M.; Forano, E. Production of oligosaccharides and cellobionic acid by Fibrobacter succinogenes S85 growing on sugars, cellulose and wheat straw. Appl. Microbiol. Biotechnol. 2009, 83, 425–433. [Google Scholar] [CrossRef]
- Neumann, A.P.; Weimer, P.J.; Suen, G. A global analysis of gene expression in Fibrobacter succinogenes S85 grown on cellulose and soluble sugars at different growth rates. Biotechnol. Biofuels 2018, 11, 295. [Google Scholar] [CrossRef]
- Bryant, M.P.; Robinson, I.M.; Chu, H. Observations on the nutrition of Bacteroides succinogenes—A ruminal cellulolytic bacterium. J. Dairy Sci. 1959, 42, 1831–1847. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Content |
| Corn | 42.57 |
| Soybean meal | 25.54 |
| Maize germ | 8.51 |
| Wheat bran | 5.96 |
| Silage corn stalks | 13.86 |
| CaHPO4 | 1.28 |
| NaHCO3 | 0.43 |
| NaCl | 0.85 |
| Premix (1) | 1 |
| Total | 100.00 |
| Nutrient level (2) | |
| Dry matter | 89.72 |
| Metabolic energy/(MJ/kg) | 10.50 |
| Crude protein | 19.07 |
| Ether extract | 2.31 |
| Neutral detergent fiber | 60.41 |
| Acid detergent fiber | 16.62 |
| Calcium | 0.78 |
| Phosphorus | 0.58 |
| Selenium/(mg/kg) | 0.04 |
| Items | Time(d) | Group | p Value | |||
|---|---|---|---|---|---|---|
| SY1 | SY2 | SY3 | SY4 | |||
| Neutral detergent fiber (NDF) | 8 | 73.82 ± 1.75 | 70.92 ± 0.92 | 70.52 ± 2.09 | 73.77 ± 1.61 | 0.0154 |
| 30 | 75.66 ± 0.61 Bb | 79.42 ± 1.15 Aa | 73.98 ± 1.51 Bb | 75.84 ± 1.41 Bb | <0.0001 | |
| 90 | 79.92 ± 1.11 Aa | 77.04 ± 1.76 Ab | 78.46 ± 1.35 Aab | 76.79 ± 1.05 Ab | 0.0266 | |
| Acid detergent fiber (ADF) | 8 | 45.76 ± 2.84 Aa | 41.41 ± 2.54 Aab | 39.57 ± 3.79 Ab | 45.00 ± 2.85 Aab | 0.0331 |
| 30 | 49.09 ± 1.37 Bb | 55.78 ± 1.84 Aa | 46.70 ± 1.23 Bb | 48.20 ± 1.01 Bb | <0.0001 | |
| 90 | 54.47 ± 1.95 Aa | 50.29 ± 1.95 Ab | 51.79 ± 1.71 Aab | 50.24 ± 2.03 Aab | 0.0300 | |
| Hemicellulose | 8 | 84.47 ± 1.36 Aab | 82.11 ± 0.37 Ab | 82.26 ± 1.48 Aab | 84.69 ± 1.27 Aa | 0.0099 |
| 30 | 85.75 ± 0.53 ABab | 88.39 ± 1.11 Aa | 84.34 ± 1.89 Bb | 86.33 ± 1.82 ABab | 0.0094 | |
| 90 | 89.58 ± 1.20 | 87.18 ± 1.78 | 88.59 ± 1.24 | 86.87 ± 0.76 | 0.0519 | |
| Cellulose | 8 | 55.53 ± 3.30 | 49.28 ± 2.77 | 49.66 ± 6.15 | 51.55 ± 2.94 | 0.1523 |
| 30 | 59.34 ± 1.03 Bb | 66.49 ± 1.40 Aa | 54.86 ± 0.92 Cc | 55.61 ± 1.75 Cc | <0.0001 | |
| 90 | 63.13 ± 2.86 Aa | 57.93 ± 2.56 Ab | 59.81 ± 1.57 Aab | 56.91 ± 2.22 Ab | 0.0172 | |
| Crude fiber (CF) | 8 | 51.99 ± 5.30 Aa | 49.04 ± 4.62 Aab | 37.14 ± 7.19 Ab | 43.88 ± 3.02 Aab | 0.0270 |
| 30 | 51.65 ± 4.95 ABb | 63.25 ± 5.86 Aa | 42.56 ± 4.99 Bb | 45.99 ± 4.14 Bb | 0.0008 | |
| 90 | 53.30 ± 5.07 | 53.43 ± 1.32 | 54.36 ± 4.03 | 54.24 ± 8.52 | 0.6724 | |
| Acid detergent lignin | 8 | 41.70 ± 6.24 ABab | 41.01 ± 4.41 ABab | 35.85 ± 6.36 Bb | 51.24 ± 4.81 Aa | 0.0133 |
| 30 | 45.53 ± 4.94 | 54.87 ± 5.98 | 48.61 ± 7.43 | 53.61 ± 1.49 | 0.0895 | |
| 90 | 59.60 ± 3.48 | 56.27 ± 4.58 | 57.36 ± 3.93 | 59.40 ± 3.77 | 0.6109 | |
| Items | Groups | p Value | |||
|---|---|---|---|---|---|
| SY1 | SY2 | SY3 | SY4 | ||
| pH value | 7.21 ± 0.09 Ab | 7.32 ± 0.09 Aab | 7.21 ± 0.14 Ab | 7.52 ± 0.13 Aa | 0.0177 |
| NH3-N/(mg·dL−1) | 47.83 ± 6.17 | 44.88 ± 5.67 | 43.33 ± 3.68 | 48.75 ± 6.40 | 0.6977 |
| MCP/(mg·mL−1) | 2.80 ± 0.75 | 1.86 ± 0.47 | 1.98 ± 0.06 | 1.81 ± 0.44 | 0.0746 |
| TVFA/(mmol·L−1) | 18.62 ± 2.96 ABab | 23.59 ± 2.78 Aa | 22.67 ± 1.84 Aa | 13.17 ± 1.15 Bb | 0.0009 |
| Acetate/% | 11.94 ± 1.86 ABbc | 16.07 ± 2.00 Aa | 15.09 ± 1.09 Aab | 8.70 ± 0.72 Bc | 0.0006 |
| Propionate/% | 3.02 ± 0.46 ABbc | 4.05 ± 0.50 Aa | 3.99 ± 0.42 Aab | 2.13 ± 0.13 Bc | 0.0004 |
| Acetate/Propionate | 3.95 ± 0.14 | 3.97 ± 0.24 | 3.80 ± 0.13 | 4.09 ± 0.18 | 0.4126 |
| Isobutyrate/% | 0.58 ± 0.08 Aa | 0.53 ± 0.05 Aab | 0.55 ± 0.01 Aa | 0.40 ± 0.05 Ab | 0.0124 |
| Butyrate/% | 2.07 ± 0.46 Aab | 2.11 ± 0.25 Aa | 2.13 ± 0.32 Aab | 1.29 ± 0.24 Ab | 0.0294 |
| Isovalerate/% | 0.76 ± 0.11 Aa | 0.59 ± 0.07 ABab | 0.65 ± 0.01 ABab | 0.48 ± 0.07 Bb | 0.0093 |
| Valerate/% | 0.26 ± 0.04 | 0.24 ± 0.04 | 0.26 ± 0.02 | 0.18 ± 0.03 | 0.0588 |
| Items | Groups | p Value | |||
|---|---|---|---|---|---|
| SY1 | SY2 | SY3 | SY4 | ||
| α-amylase/(U·dL−1) | 50.84 ± 1.40 | 52.15 ± 2.15 | 49.81 ± 2.28 | 51.55 ± 1.31 | 0.4784 |
| Protease/(U·mL−1) | 3.15 ± 0.30 Bb | 2.38 ± 0.11 Bc | 3.97 ± 0.22 Aa | 3.08 ± 0.17 Bb | 0.0003 |
| Cellulase/(U·mg−1) | 34.71 ± 1.29 | 24.15 ± 1.30 | 30.01 ± 6.61 | 26.34 ± 8.03 | 0.2793 |
| Groups | KO Number | Enzyme/Protein | Enzyme Classification |
|---|---|---|---|
| SY1 | K11389 | glyceraldehyde-3-phosphate dehydrogenase (ferredoxin) | EC:1.2.7.6 |
| K02092 | allophycocyanin alpha subunit | ||
| K02975 | small subunit ribosomal protein S25e | ||
| K07547 | 2-[hydroxy(phenyl)methyl]-succinyl-CoA dehydrogenase BbsC subunit | EC:1.1.1.35 | |
| SY2 | K12073 | 1,4-dihydroxy-2-naphthoyl-CoA hydrolase | EC:3.1.2.28 |
| K11214 | sedoheptulokinase | EC:2.7.1.14 | |
| K00211 | prephenate dehydrogenase (NADP+) | EC:1.3.1.13 | |
| K01663 | glutamine amidotransferase/cyclase | EC:2.4.2.- 4.1.3.- | |
| K02384 | flagellar protein FlbC | ||
| K13713 | fusion protein PurCD | EC:6.3.2.6 6.3.4.13 | |
| SY4 | K14331 | fatty aldehyde decarbonylase | EC:4.1.99.5 |
| K12535 | outer membrane protein RsaF | ||
| K12533 | ATP-binding cassette, subfamily C, bacterial RsaD | ||
| K05348 | 2-hydroxycinnamic acid beta-D-glucosylisomelase | EC:5.2.1.- | |
| K13585 | holdfast attachment protein HfaA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, W.; Shi, H.; Gong, C.; Liu, K.; Li, G. Effects of Different Yeast Selenium Levels on Rumen Fermentation Parameters, Digestive Enzyme Activity and Gastrointestinal Microflora of Sika Deer during Antler Growth. Microorganisms 2023, 11, 1444. https://doi.org/10.3390/microorganisms11061444
Sun W, Shi H, Gong C, Liu K, Li G. Effects of Different Yeast Selenium Levels on Rumen Fermentation Parameters, Digestive Enzyme Activity and Gastrointestinal Microflora of Sika Deer during Antler Growth. Microorganisms. 2023; 11(6):1444. https://doi.org/10.3390/microorganisms11061444
Chicago/Turabian StyleSun, Weili, Hongpeng Shi, Chengyan Gong, Keyuan Liu, and Guangyu Li. 2023. "Effects of Different Yeast Selenium Levels on Rumen Fermentation Parameters, Digestive Enzyme Activity and Gastrointestinal Microflora of Sika Deer during Antler Growth" Microorganisms 11, no. 6: 1444. https://doi.org/10.3390/microorganisms11061444
APA StyleSun, W., Shi, H., Gong, C., Liu, K., & Li, G. (2023). Effects of Different Yeast Selenium Levels on Rumen Fermentation Parameters, Digestive Enzyme Activity and Gastrointestinal Microflora of Sika Deer during Antler Growth. Microorganisms, 11(6), 1444. https://doi.org/10.3390/microorganisms11061444