

Expression of Envelope Protein Encoded by Endogenous Retrovirus K102 in Rheumatoid Arthritis Neutrophils

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Human Subjects
2.2. Isolation of Polymorphonuclear and Mononuclear Blood Leukocytes
2.3. RNA Isolation and RNA-Seq
2.4. Bioinformatics of Retrotransposons
2.5. Real-Time and Ordinary Polymerase Chain Reaction (PCR)
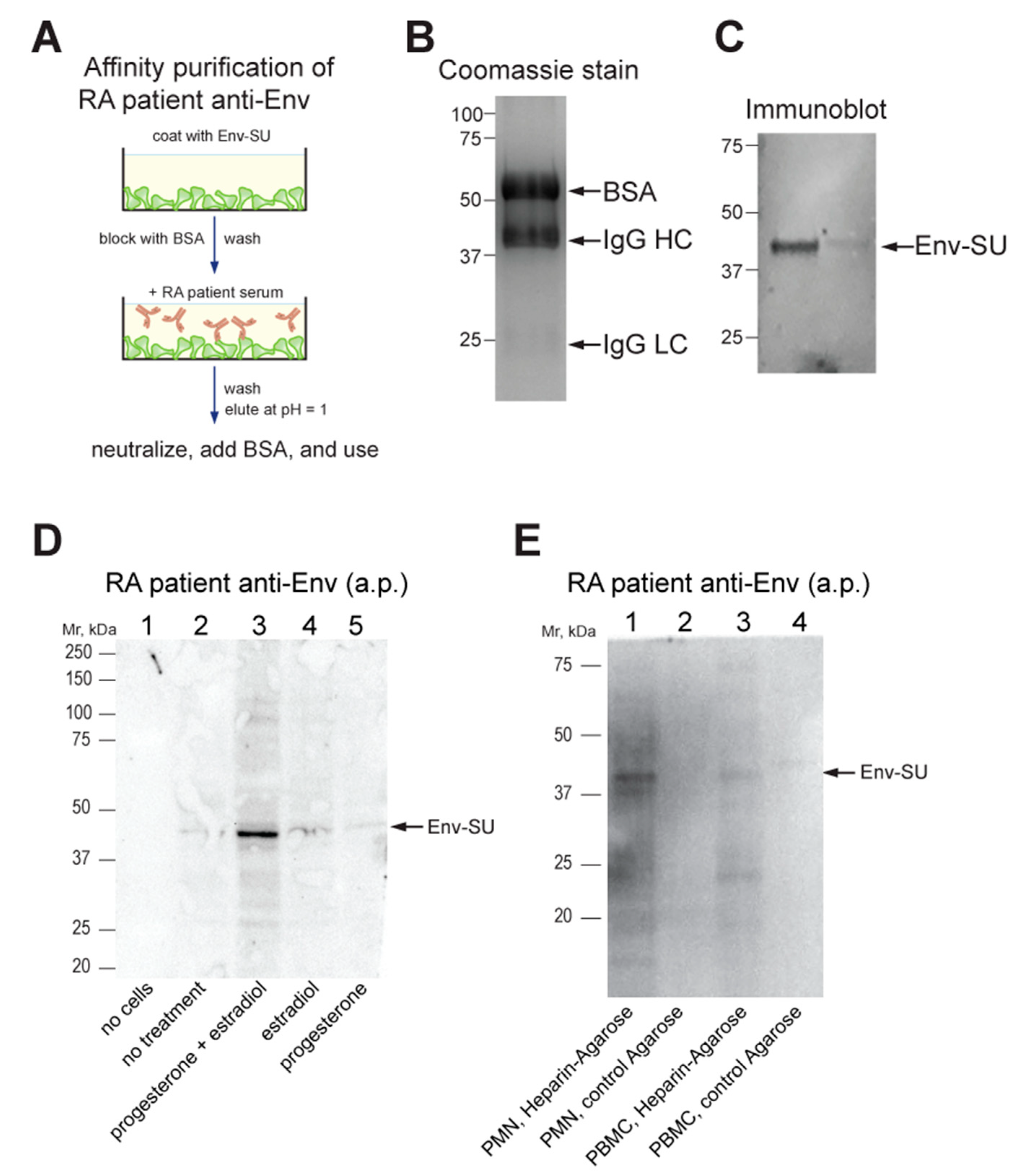
2.6. Affinity-Purification of Patient Antibodies against Env
2.7. Breast Cancer Cells and Hormone Treatment to Induce HERV-K Expression
2.8. Enrichment in Env from RA Leukocytes
2.9. Transient Expression and Immunofluorescence Staining
2.10. Gel Electrophoresis and Immunoblotting
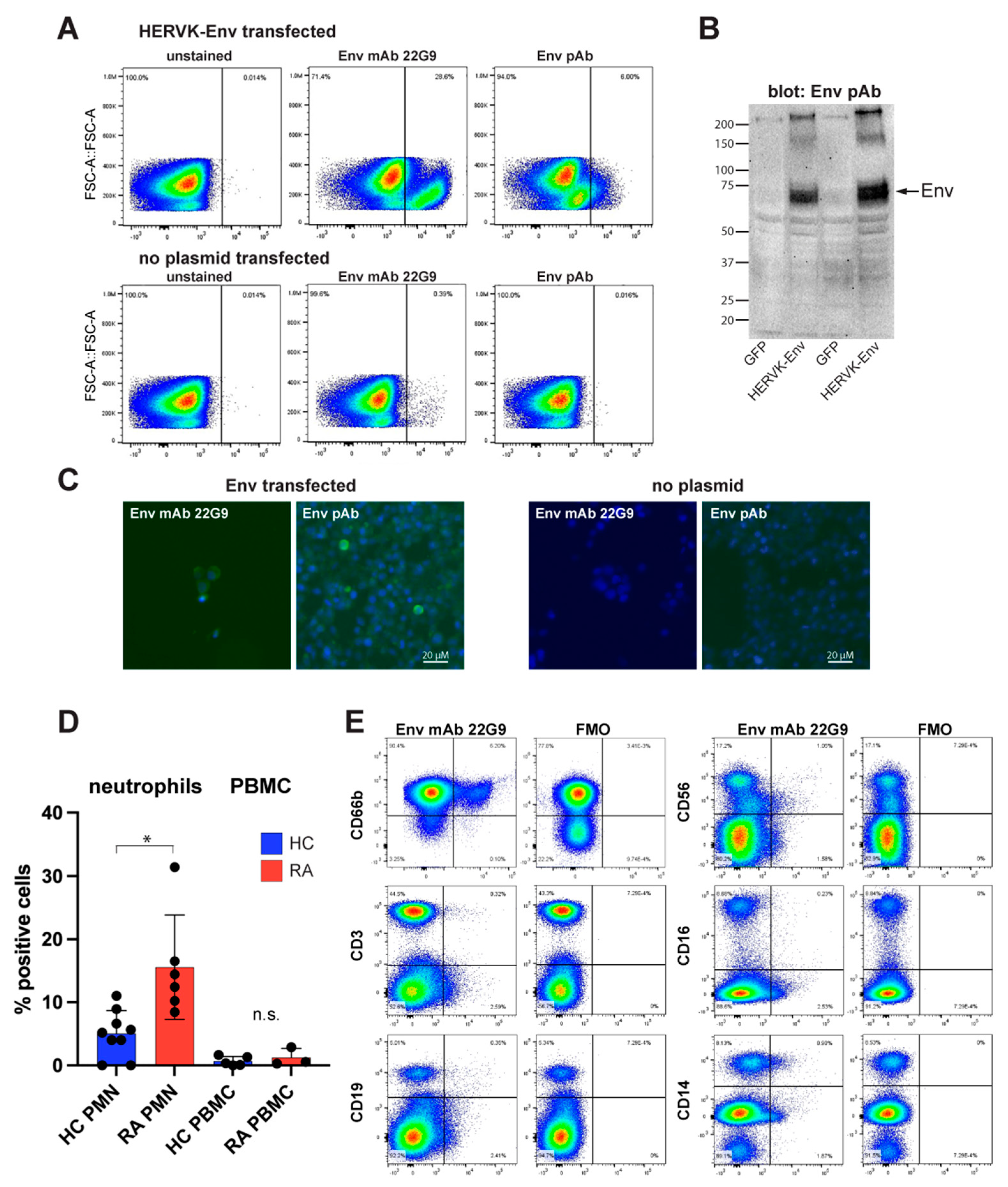
2.11. Generation of a Monoclonal Antibody against Env-SU
2.12. Flow Cytometry
2.13. Statistical Analysis
3. Results
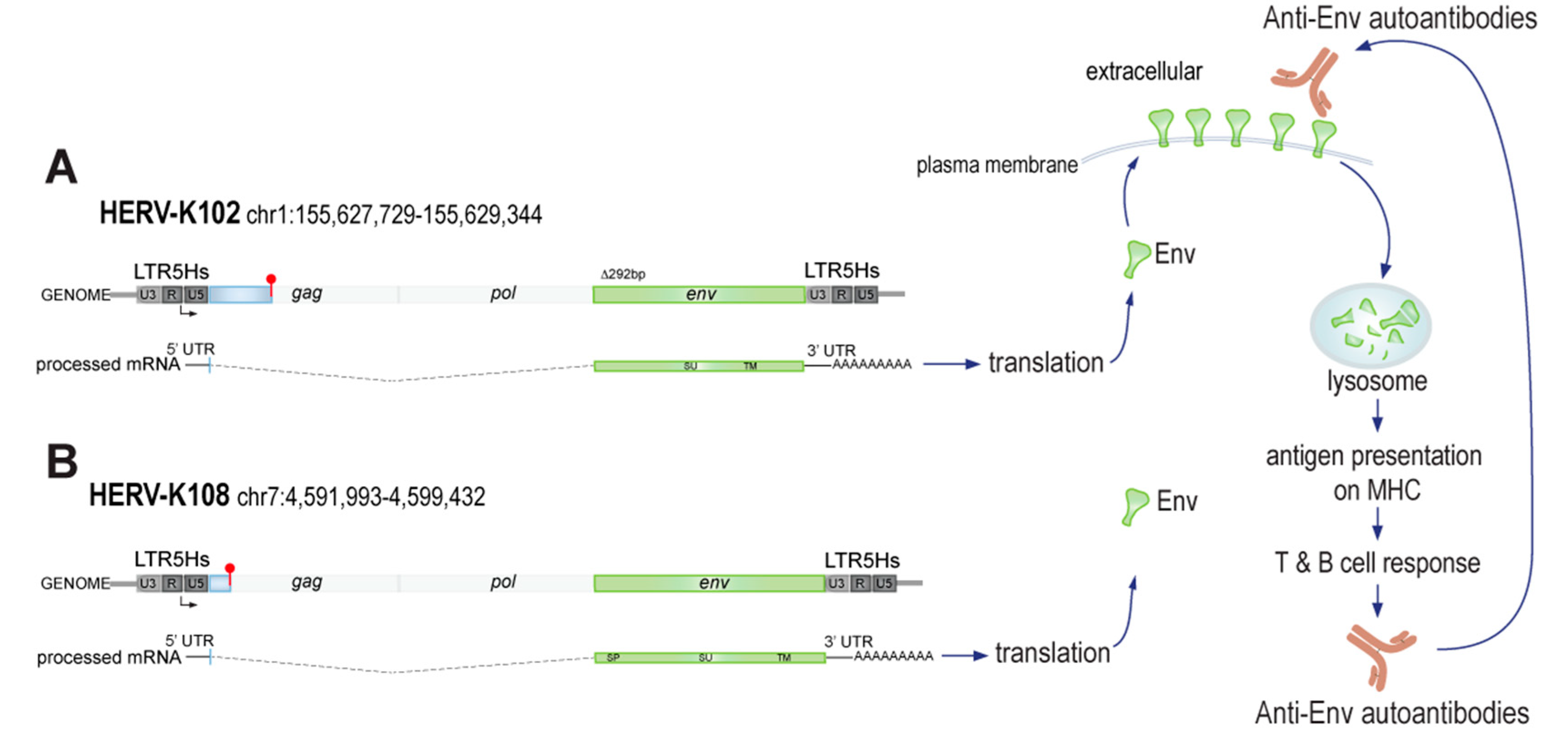
3.1. Transcripts from HERV-K Loci That Can Produce Env in RA
3.2. Validation of HERV-K Transcripts in a Public RA Data Set
3.3. Quantitation of K102 and K108 Transcripts in RA Neutrophils and PBMC
3.4. Anti-Env Autoantibodies from RA Patients Recognize Cell Surface-Exposed Env
3.5. Endogenously Expressed Env in RA Neutrophils
3.6. Generation and Validation of Antibodies Specific for Env for Flow Cytometry
3.7. Endogenously Expressed Env in Immune Cells from RA Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goronzy, J.J.; Weyand, C.M. Rheumatoid arthritis. Immunol. Rev. 2005, 204, 55–73. [Google Scholar] [CrossRef] [PubMed]
- Schellekens, G.A.; Visser, H.; de Jong, B.A.; van den Hoogen, F.H.; Hazes, J.M.; Breedveld, F.C.; van Venrooij, W.J. The diagnostic properties of rheumatoid arthritis antibodies recognizing a cyclic citrullinated peptide. Arthritis Rheum. 2000, 43, 155–163. [Google Scholar] [CrossRef] [PubMed]
- De Rycke, L.; Nicholas, A.P.; Cantaert, T.; Kruithof, E.; Echols, J.D.; Vandekerckhove, B.; Veys, E.M.; De Keyser, F.; Baeten, D. Synovial intracellular citrullinated proteins colocalizing with peptidyl arginine deiminase as pathophysiologically relevant antigenic determinants of rheumatoid arthritis-specific humoral autoimmunity. Arthritis Rheum. 2005, 52, 2323–2330. [Google Scholar] [CrossRef] [PubMed]
- Too, C.L.; Murad, S.; Dhaliwal, J.S.; Larsson, P.; Jiang, X.; Ding, B.; Alfredsson, L.; Klareskog, L.; Padyukov, L. Polymorphisms in peptidylarginine deiminase associate with rheumatoid arthritis in diverse Asian populations: Evidence from MyEIRA study and meta-analysis. Arthritis Res. Ther. 2012, 14, R250. [Google Scholar] [CrossRef]
- Suzuki, A.; Yamada, R.; Chang, X.; Tokuhiro, S.; Sawada, T.; Suzuki, M.; Nagasaki, M.; Nakayama-Hamada, M.; Kawaida, R.; Ono, M.; et al. Functional haplotypes of PADI4, encoding citrullinating enzyme peptidylarginine deiminase 4, are associated with rheumatoid arthritis. Nat. Genet. 2003, 34, 395–402. [Google Scholar] [CrossRef]
- Hashemi, M.; Zakeri, Z.; Taheri, H.; Bahari, G.; Taheri, M. Association between Peptidylarginine Deiminase Type 4 rs1748033 Polymorphism and Susceptibility to Rheumatoid Arthritis in Zahedan, Southeast Iran. Iran. J. Allergy Asthma Immunol. 2015, 14, 255–260. [Google Scholar] [PubMed]
- Hua, J.; Huang, W. Peptidylarginine deiminase 4 -104C/T polymorphism and risk of rheumatoid arthritis: A pooled analysis based on different populations. PLoS ONE 2018, 13, e0193674. [Google Scholar] [CrossRef]
- Nielen, M.M.; van der Horst, A.R.; van Schaardenburg, D.; van der Horst-Bruinsma, I.E.; van de Stadt, R.J.; Aarden, L.; Dijkmans, B.A.; Hamann, D. Antibodies to citrullinated human fibrinogen (ACF) have diagnostic and prognostic value in early arthritis. Ann. Rheum. Dis. 2005, 64, 1199–1204. [Google Scholar] [CrossRef]
- Paalanen, K.; Puolakka, K.; Nikiphorou, E.; Hannonen, P.; Sokka, T. Is seronegative rheumatoid arthritis true rheumatoid arthritis? A nationwide cohort study. Rheumatology 2021, 60, 2391–2395. [Google Scholar] [CrossRef]
- Herve, C.A.; Lugli, E.B.; Brand, A.; Griffiths, D.J.; Venables, P.J. Autoantibodies to human endogenous retrovirus-K are frequently detected in health and disease and react with multiple epitopes. Clin. Exp. Immunol. 2002, 128, 75–82. [Google Scholar] [CrossRef]
- Mameli, G.; Erre, G.L.; Caggiu, E.; Mura, S.; Cossu, D.; Bo, M.; Cadoni, M.L.; Piras, A.; Mundula, N.; Colombo, E.; et al. Identification of a HERV-K env surface peptide highly recognized in Rheumatoid Arthritis (RA) patients: A cross-sectional case-control study. Clin. Exp. Immunol. 2017, 189, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Nelson, P.N.; Roden, D.; Nevill, A.; Freimanis, G.L.; Trela, M.; Ejtehadi, H.D.; Bowman, S.; Axford, J.; Veitch, A.M.; Tugnet, N.; et al. Rheumatoid arthritis is associated with IgG antibodies to human endogenous retrovirus gag matrix: A potential pathogenic mechanism of disease? J. Rheumatol. 2014, 41, 1952–1960. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hefton, A.; Ni, K.; Ukadike, K.C.; Bowen, M.A.; Eckert, M.; Stevens, A.; Lood, C.; Mustelin, T. Autoantibodies Against Unmodified and Citrullinated Human Endogenous Retrovirus K Envelope Protein in Patients With Rheumatoid Arthritis. J. Rheumatol. 2022, 49, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Fraziano, M.; Montesano, C.; Lombardi, V.R.; Sammarco, I.; De Pisa, F.; Mattei, M.; Valesini, G.; Pittoni, V.; Colizzi, V. Epitope specificity of anti-HIV antibodies in human and murine autoimmune diseases. AIDS Res. Hum. Retrovir. 1996, 12, 491–496. [Google Scholar] [CrossRef]
- Ejtehadi, H.D.; Freimanis, G.L.; Ali, H.A.; Bowman, S.; Alavi, A.; Axford, J.; Callaghan, R.; Nelson, P.N. The potential role of human endogenous retrovirus K10 in the pathogenesis of rheumatoid arthritis: A preliminary study. Ann. Rheum. Dis. 2006, 65, 612–616. [Google Scholar] [CrossRef]
- Reynier, F.; Verjat, T.; Turrel, F.; Imbert, P.E.; Marotte, H.; Mougin, B.; Miossec, P. Increase in human endogenous retrovirus HERV-K (HML-2) viral load in active rheumatoid arthritis. Scand. J. Immunol. 2009, 70, 295–299. [Google Scholar] [CrossRef]
- Freimanis, G.; Hooley, P.; Ejtehadi, H.D.; Ali, H.A.; Veitch, A.; Rylance, P.B.; Alawi, A.; Axford, J.; Nevill, A.; Murray, P.G.; et al. A role for human endogenous retrovirus-K (HML-2) in rheumatoid arthritis: Investigating mechanisms of pathogenesis. Clin. Exp. Immunol. 2010, 160, 340–347. [Google Scholar] [CrossRef]
- Ehlhardt, S.; Seifert, M.; Schneider, J.; Ojak, A.; Zang, K.D.; Mehraein, Y. Human endogenous retrovirus HERV-K(HML-2) Rec expression and transcriptional activities in normal and rheumatoid arthritis synovia. J. Rheumatol. 2006, 33, 16–23. [Google Scholar]
- Subramanian, R.P.; Wildschutte, J.H.; Russo, C.; Coffin, J.M. Identification, characterization, and comparative genomic distribution of the HERV-K (HML-2) group of human endogenous retroviruses. Retrovirology 2011, 8, 90. [Google Scholar] [CrossRef]
- Boller, K.; Schonfeld, K.; Lischer, S.; Fischer, N.; Hoffmann, A.; Kurth, R.; Tonjes, R.R. Human endogenous retrovirus HERV-K113 is capable of producing intact viral particles. J. Gen. Virol. 2008, 89, 567–572. [Google Scholar] [CrossRef]
- Dewannieux, M.; Harper, F.; Richaud, A.; Letzelter, C.; Ribet, D.; Pierron, G.; Heidmann, T. Identification of an infectious progenitor for the multiple-copy HERV-K human endogenous retroelements. Genome Res. 2006, 16, 1548–1556. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Montojo, M.; Doucet-O’Hare, T.; Henderson, L.; Nath, A. Human endogenous retrovirus-K (HML-2): A comprehensive review. Crit. Rev. Microbiol. 2018, 44, 715–738. [Google Scholar] [CrossRef] [PubMed]
- Geis, F.K.; Goff, S.P. Silencing and Transcriptional Regulation of Endogenous Retroviruses: An Overview. Viruses 2020, 12, 884. [Google Scholar] [CrossRef] [PubMed]
- Grow, E.J.; Flynn, R.A.; Chavez, S.L.; Bayless, N.L.; Wossidlo, M.; Wesche, D.J.; Martin, L.; Ware, C.B.; Blish, C.A.; Chang, H.Y.; et al. Intrinsic retroviral reactivation in human preimplantation embryos and pluripotent cells. Nature 2015, 522, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Goering, W.; Ribarska, T.; Schulz, W.A. Selective changes of retroelement expression in human prostate cancer. Carcinogenesis 2011, 32, 1484–1492. [Google Scholar] [CrossRef]
- Buscher, K.; Trefzer, U.; Hofmann, M.; Sterry, W.; Kurth, R.; Denner, J. Expression of human endogenous retrovirus K in melanomas and melanoma cell lines. Cancer Res. 2005, 65, 4172–4180. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Maldarelli, F.; Mellors, J.; Coffin, J.M. HIV-1 infection leads to increased transcription of human endogenous retrovirus HERV-K (HML-2) proviruses in vivo but not to increased virion production. J. Virol. 2014, 88, 11108–11120. [Google Scholar] [CrossRef]
- Bowen, L.N.; Tyagi, R.; Li, W.; Alfahad, T.; Smith, B.; Wright, M.; Singer, E.J.; Nath, A. HIV-associated motor neuron disease: HERV-K activation and response to antiretroviral therapy. Neurology 2016, 87, 1756–1762. [Google Scholar] [CrossRef]
- Contreras-Galindo, R.; Almodovar-Camacho, S.; Gonzalez-Ramirez, S.; Lorenzo, E.; Yamamura, Y. Comparative longitudinal studies of HERV-K and HIV-1 RNA titers in HIV-1-infected patients receiving successful versus unsuccessful highly active antiretroviral therapy. AIDS Res. Hum. Retrovir. 2007, 23, 1083–1086. [Google Scholar] [CrossRef]
- Contreras-Galindo, R.; Gonzalez, M.; Almodovar-Camacho, S.; Gonzalez-Ramirez, S.; Lorenzo, E.; Yamamura, Y. A new Real-Time-RT-PCR for quantitation of human endogenous retroviruses type K (HERV-K) RNA load in plasma samples: Increased HERV-K RNA titers in HIV-1 patients with HAART non-suppressive regimens. J. Virol. Methods 2006, 136, 51–57. [Google Scholar] [CrossRef]
- Contreras-Galindo, R.; Kaplan, M.H.; Markovitz, D.M.; Lorenzo, E.; Yamamura, Y. Detection of HERV-K(HML-2) viral RNA in plasma of HIV type 1-infected individuals. AIDS Res. Hum. Retrovir. 2006, 22, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Galindo, R.; Lopez, P.; Velez, R.; Yamamura, Y. HIV-1 infection increases the expression of human endogenous retroviruses type K (HERV-K) in vitro. AIDS Res. Hum. Retrovir. 2007, 23, 116–122. [Google Scholar] [CrossRef] [PubMed]
- de Mulder, M.; SenGupta, D.; Deeks, S.G.; Martin, J.N.; Pilcher, C.D.; Hecht, F.M.; Sacha, J.B.; Nixon, D.F.; Michaud, H.A. Anti-HERV-K (HML-2) capsid antibody responses in HIV elite controllers. Retrovirology 2017, 14, 41. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Hernandez, M.J.; Swanson, M.D.; Contreras-Galindo, R.; Cookinham, S.; King, S.R.; Noel, R.J., Jr.; Kaplan, M.H.; Markovitz, D.M. Expression of human endogenous retrovirus type K (HML-2) is activated by the Tat protein of HIV-1. J. Virol. 2012, 86, 7790–7805. [Google Scholar] [CrossRef]
- Tokuyama, M.; Gunn, B.M.; Venkataraman, A.; Kong, Y.; Kang, I.; Rakib, T.; Townsend, M.J.; Costenbader, K.H.; Alter, G.; Iwasaki, A. Antibodies against human endogenous retrovirus K102 envelope activate neutrophils in systemic lupus erythematosus. J. Exp. Med. 2021, 218, e20191766. [Google Scholar] [CrossRef]
- Khadjinova, A.I.; Wang, X.; Laine, A.; Ukadike, K.; Eckert, M.; Stevens, A.; Bengtsson, A.A.; Lood, C.; Mustelin, T. Autoantibodies against the envelope proteins of endogenous retroviruses K102 and K108 in patients with systemic lupus erythematosus correlate with active disease. Clin. Exp. Rheumatol. 2021, 40, 1306–1312. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Wang, L.; Wang, S.; Li, W. RSeQC: Quality control of RNA-seq experiments. Bioinformatics 2012, 28, 2184–2185. [Google Scholar] [CrossRef]
- Jin, Y.; Tam, O.H.; Paniagua, E.; Hammell, M. TEtranscripts: A package for including transposable elements in differential expression analysis of RNA-seq datasets. Bioinformatics 2015, 31, 3593–3599. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Tarazona, S.; Garcia-Alcalde, F.; Dopazo, J.; Ferrer, A.; Conesa, A. Differential expression in RNA-seq: A matter of depth. Genome Res. 2011, 21, 2213–2223. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Kawakami, M.; Ushikubo, H. Stimulation of expression of the human endogenous retrovirus genome by female steroid hormones in human breast cancer cell line T47D. J. Virol. 1987, 61, 2059–2062. [Google Scholar] [CrossRef] [PubMed]
- Mayer, J.; Sauter, M.; Racz, A.; Scherer, D.; Mueller-Lantzsch, N.; Meese, E. An almost-intact human endogenous retrovirus K on human chromosome 7. Nat. Genet. 1999, 21, 257–258. [Google Scholar] [CrossRef] [PubMed]
- Reus, K.; Mayer, J.; Sauter, M.; Scherer, D.; Muller-Lantzsch, N.; Meese, E. Genomic organization of the human endogenous retrovirus HERV-K(HML-2.HOM) (ERVK6) on chromosome 7. Genomics 2001, 72, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Shchetynsky, K.; Diaz-Gallo, L.M.; Folkersen, L.; Hensvold, A.H.; Catrina, A.I.; Berg, L.; Klareskog, L.; Padyukov, L. Discovery of new candidate genes for rheumatoid arthritis through integration of genetic association data with expression pathway analysis. Arthritis Res. Ther. 2017, 19, 19. [Google Scholar] [CrossRef] [PubMed]
- Wang-Johanning, F.; Frost, A.R.; Johanning, G.L.; Khazaeli, M.B.; LoBuglio, A.F.; Shaw, D.R.; Strong, T.V. Expression of human endogenous retrovirus k envelope transcripts in human breast cancer. Clin. Cancer Res. 2001, 7, 1553–1560. [Google Scholar] [PubMed]
- Montesion, M.; Williams, Z.H.; Subramanian, R.P.; Kuperwasser, C.; Coffin, J.M. Promoter expression of HERV-K (HML-2) provirus-derived sequences is related to LTR sequence variation and polymorphic transcription factor binding sites. Retrovirology 2018, 15, 57. [Google Scholar] [CrossRef]
- Beimforde, N.; Hanke, K.; Ammar, I.; Kurth, R.; Bannert, N. Molecular cloning and functional characterization of the human endogenous retrovirus K113. Virology 2008, 371, 216–225. [Google Scholar] [CrossRef]
- Robinson-McCarthy, L.R.; McCarthy, K.R.; Raaben, M.; Piccinotti, S.; Nieuwenhuis, J.; Stubbs, S.H.; Bakkers, M.J.G.; Whelan, S.P.J. Reconstruction of the cell entry pathway of an extinct virus. PLoS Pathog. 2018, 14, e1007123. [Google Scholar] [CrossRef]
- Wang-Johanning, F.; Radvanyi, L.; Rycaj, K.; Plummer, J.B.; Yan, P.; Sastry, K.J.; Piyathilake, C.J.; Hunt, K.K.; Johanning, G.L. Human endogenous retrovirus K triggers an antigen-specific immune response in breast cancer patients. Cancer Res. 2008, 68, 5869–5877. [Google Scholar] [CrossRef]
- Wang-Johanning, F.; Li, M.; Esteva, F.J.; Hess, K.R.; Yin, B.; Rycaj, K.; Plummer, J.B.; Garza, J.G.; Ambs, S.; Johanning, G.L. Human endogenous retrovirus type K antibodies and mRNA as serum biomarkers of early-stage breast cancer. Int. J. Cancer 2014, 134, 587–595. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RA Patients (n = 17) | |||
| Age (average ± S.D.) | 52.1 | ±14.5 | Range 32–73 |
| Sex | 4 male | 13 female | |
| Ethnicity/race | 9 white, 4 hispanic | 1 black, 1 Pacific Is | 1 Native Am, 1 n.d. * |
| Smoking status | 2 current | 3 former | 12 never |
| CCP/RF # | 3 seronegative | 14 seroposite | |
| Erosions | 8 not present | 9 present | |
| Clinical Disease Activity Index | 10.1 | ±9.9 | Range 0–31 |
| Current medication | 4 prednisone, 5 methotrexate | 8 anti-TNF, 1 other | 1 JAK ihibitor |
| Healthy controls (n = 10) | |||
| Age (average ± S.D.) | 35.3 | ±15.9 | Range 21–63 |
| Sex | 3 male | 7 female |
| Genomic Locus * | Read Counts in HC | Read Counts in RA | Ratio RA/HC |
|---|---|---|---|
| ERVK_chr1:155,627,729–155,629,344 (K102) | 16.2 | 30.5 | 1.9 |
| ERVK_chr1:155,629,344–155,634,877 (K102) | 168.5 | 207.9 | 1.2 |
| ERVK_chr12:8,558,451–8,562,470 | 140.8 | 154.5 | 1.1 |
| ERVK_chr14:69,811,901–69,814,583 | 25.3 | 29.4 | 1.2 |
| ERVK_chr16:2,661,288–2,668,373 | 4.9 | 7.1 | 1.4 |
| ERVK_chr17:43,339,412–43,345,762 | 8.3 | 10.1 | 1.2 |
| ERVK_chr19:21,622,130–21,625,081 | 2.4 | 6.8 | 2.9 |
| ERVK_chr19:58,307,323–58,310,256 | 271.6 | 242.2 | 0.9 |
| ERVK_chr19:58,311,249–58,313,278 | 151.8 | 130.8 | 0.9 |
| ERVK_chr2:71,388,647–71,395,356 | 17.3 | 31.9 | 1.8 |
| ERVK_chr3:101,692,862–101,698,340 | 48.7 | 59.5 | 1.2 |
| ERVK_chr3:101,698,340–101,699,951 | 10.7 | 20.6 | 1.9 |
| ERVK_chr4:153,688,875–153,693,253 | 24.3 | 17.4 | 0.7 |
| ERVK_chr4:69,192,349–69,195,436 | 13.0 | 22.4 | 1.7 |
| ERVK_chr4:69,196,348–69,198,210 | 22.1 | 37.5 | 1.7 |
| ERVK_chr4:9,658,974–9,666,434 | 3.4 | 15.1 | 4.5 |
| ERVK_chr5:1,581,574–1,584,921 | 23.9 | 25.4 | 1.1 |
| ERVK_chr5:71,573,256–71,577,918 | 12.3 | 12.3 | 1.0 |
| ERVK_chr6:29,876,165–29,880,725 | 45.4 | 35.5 | 0.8 |
| ERVK_chr7:4,591,993–4,599,432 (K108) | 11.7 | 8.5 | 0.7 |
| ERVK_chrX:75,326,408–75,329,359 | 14.3 | 15.0 | 1.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laine, A.; Wang, X.; Ni, K.; Smith, S.E.B.; Najjar, R.; Whitmore, L.S.; Yacoub, M.; Bays, A.; Gale, M., Jr.; Mustelin, T. Expression of Envelope Protein Encoded by Endogenous Retrovirus K102 in Rheumatoid Arthritis Neutrophils. Microorganisms 2023, 11, 1310. https://doi.org/10.3390/microorganisms11051310
Laine A, Wang X, Ni K, Smith SEB, Najjar R, Whitmore LS, Yacoub M, Bays A, Gale M Jr., Mustelin T. Expression of Envelope Protein Encoded by Endogenous Retrovirus K102 in Rheumatoid Arthritis Neutrophils. Microorganisms. 2023; 11(5):1310. https://doi.org/10.3390/microorganisms11051310
Chicago/Turabian StyleLaine, Amanda, Xiaoxing Wang, Kathryn Ni, Sarah E. B. Smith, Rayan Najjar, Leanne S. Whitmore, Michael Yacoub, Alison Bays, Michael Gale, Jr., and Tomas Mustelin. 2023. "Expression of Envelope Protein Encoded by Endogenous Retrovirus K102 in Rheumatoid Arthritis Neutrophils" Microorganisms 11, no. 5: 1310. https://doi.org/10.3390/microorganisms11051310
APA StyleLaine, A., Wang, X., Ni, K., Smith, S. E. B., Najjar, R., Whitmore, L. S., Yacoub, M., Bays, A., Gale, M., Jr., & Mustelin, T. (2023). Expression of Envelope Protein Encoded by Endogenous Retrovirus K102 in Rheumatoid Arthritis Neutrophils. Microorganisms, 11(5), 1310. https://doi.org/10.3390/microorganisms11051310