
Emergence of Azithromycin-Resistant Neisseria gonorrhoeae Isolates Belonging to the NG-MAST Genogroup 12302 in Russia

 ,
, 
 ,
, 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection and Characterization of Russian N. gonorrhoeae Isolates: Antimicrobial Susceptibility Testing
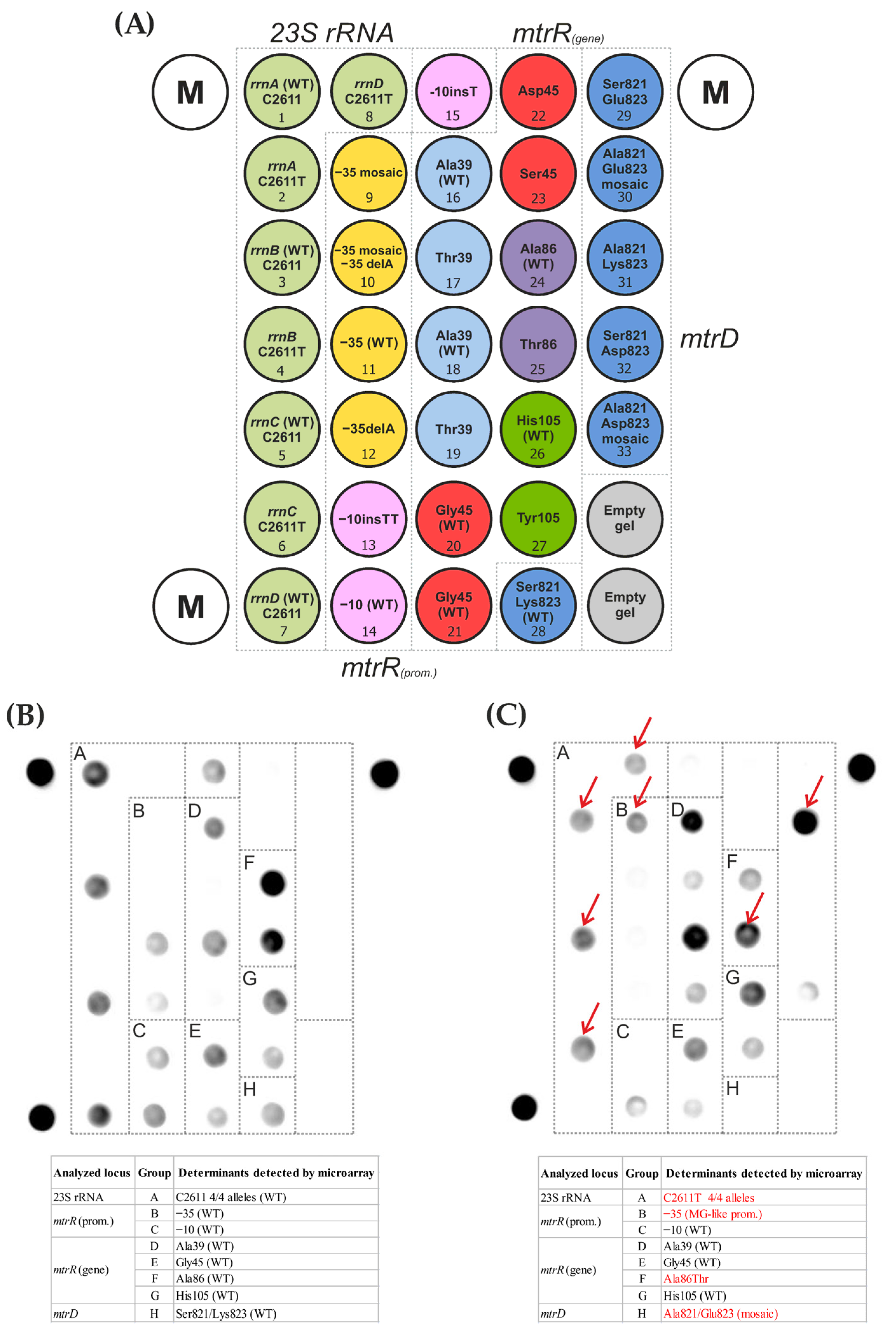
2.2. Detection of Mutations Associated with Azithromycin Resistance Using an Oligonucleotide Hydrogel Microarray
2.3. Target Sequencing of N. gonorrhoeae DNA
2.4. NG-MAST Typing of the Russian N. gonorrhoeae Isolates and Definition of the Genogroups
2.5. Analysis of the Collection of the Russian and European Samples
2.6. Construction of Phylogenetic Trees
3. Results
3.1. Azithromycin Susceptibility and Genetic Resistance Determinants of Isolates Collected in Russia in 2018–2021
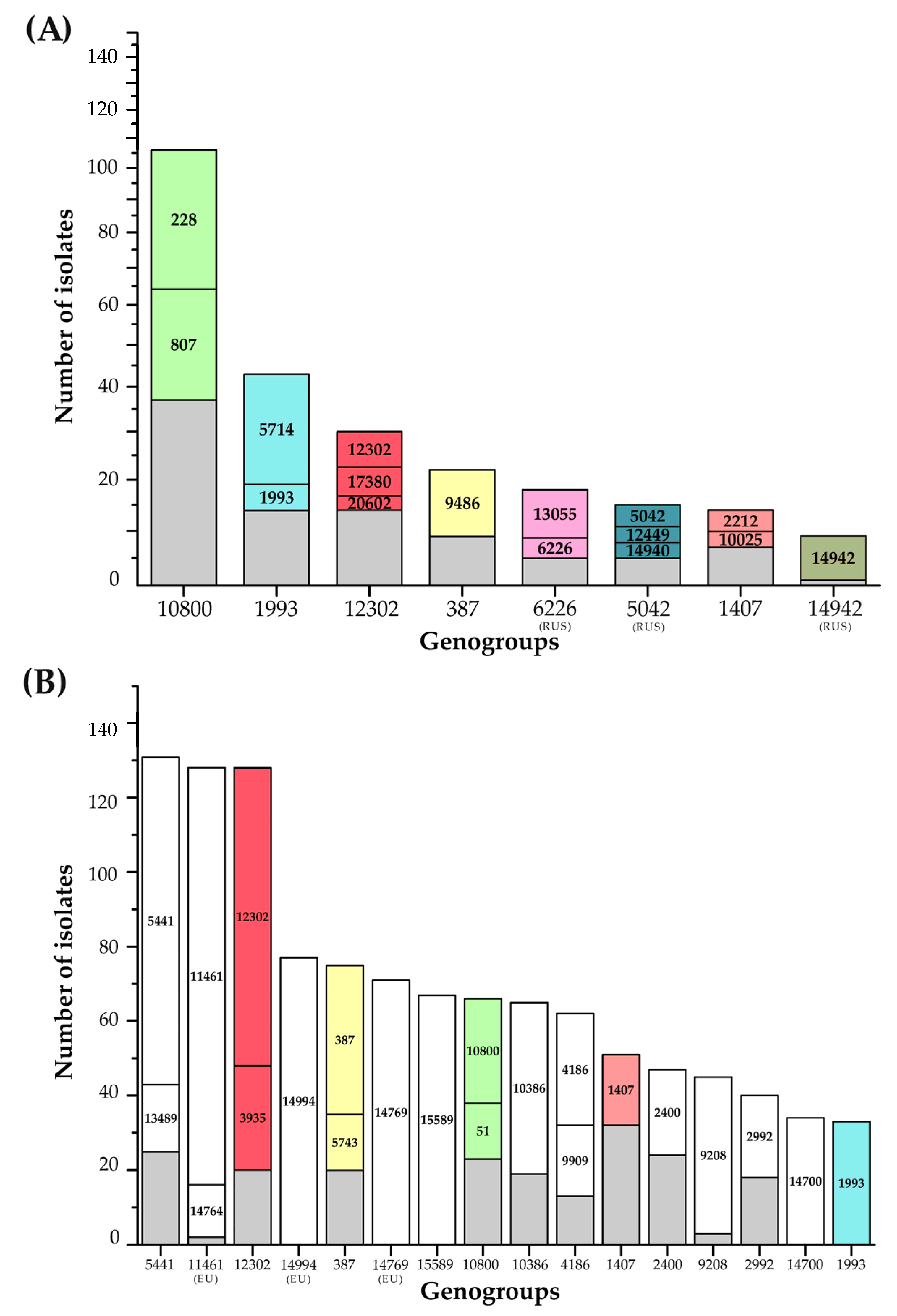
3.2. Comparison of the NG-MAST Types in the Russian and European N. gonorrhoeae Populations
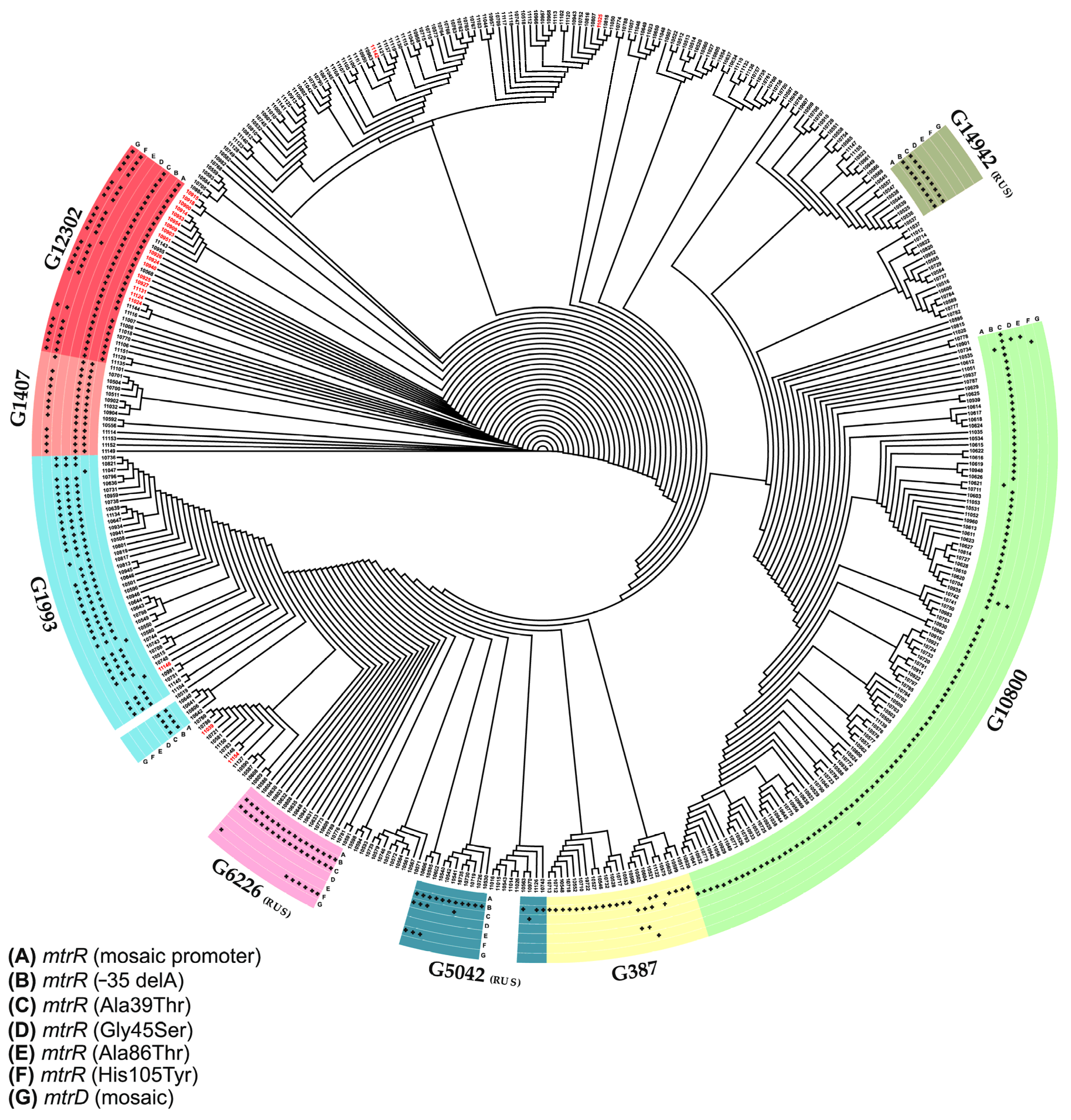
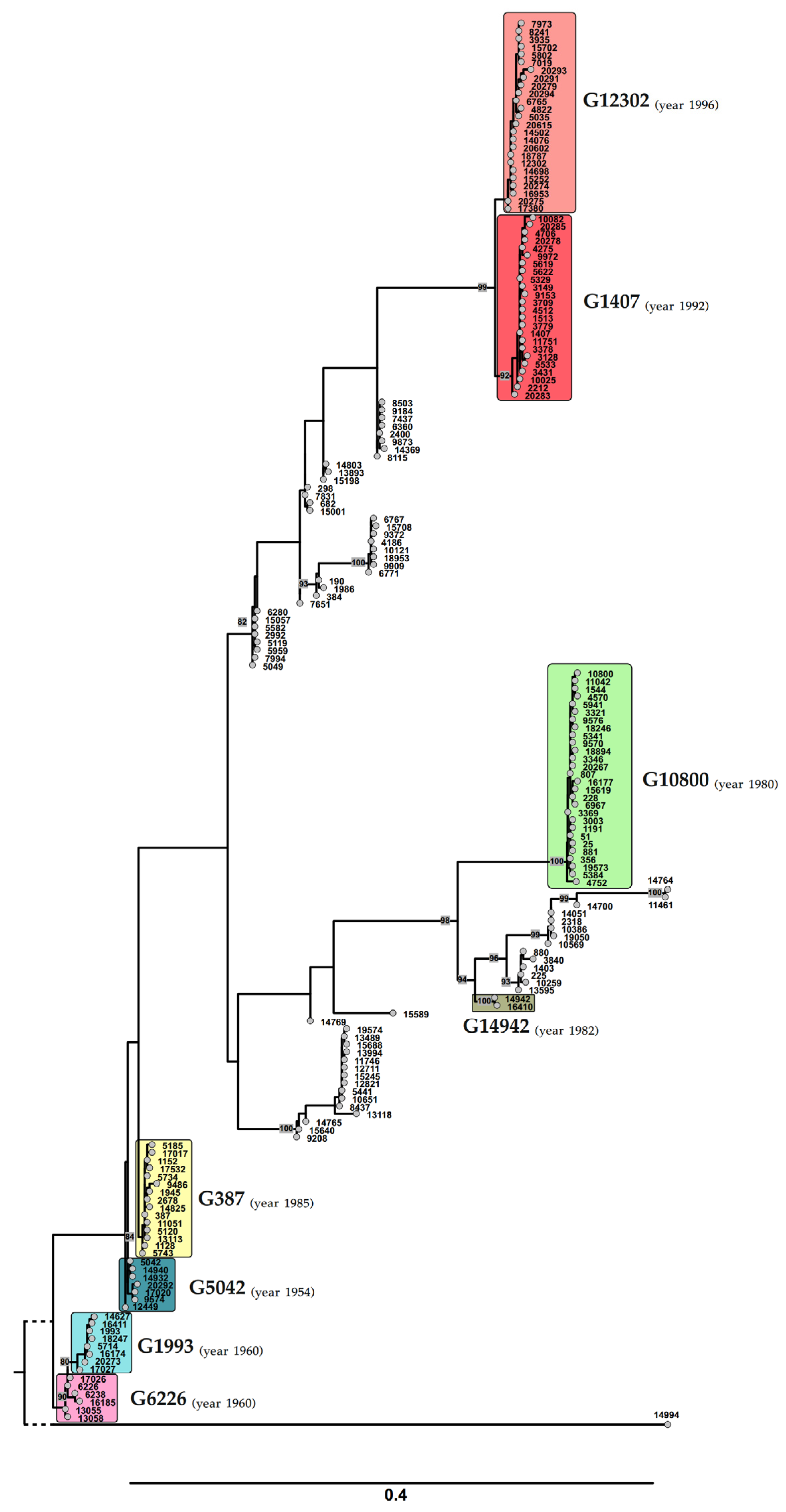
3.3. Phylogenetic Study of the Russian Isolates

4. Discussion

5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Global Progress Report on HIV, Viral Hepatitis and Sexually Transmitted Infections. 2021. Available online: https://www.who.int/publications/i/item/9789240027077 (accessed on 12 April 2023).
- Kubanov, A.A.; Bogdanova, E.V. Performance results of medical organizations providing medical care in the field of dermatovenereology in 2019–2021 in the Russian Federation. Vestn. Dermatol. Venerol. 2022, 98, 18–33. (In Russian) [Google Scholar] [CrossRef]
- Unemo, M.; Ross, J.; Serwin, A.B.; Gomberg, M.; Cusini, M.; Jensen, J.S. Background review for the ‘2020 European guideline for the diagnosis and treatment of gonorrhoea in adults’. Int. J. STD AIDS 2021, 32, 108–126. [Google Scholar] [CrossRef] [PubMed]
- Barbee, L.A.; St Cyr, S.B. Management of Neisseria gonorrhoeae in the United States: Summary of evidence from the development of the 2020 Gonorrhea Treatment Recommendations and the 2021 Centers for Disease Control and Prevention Sexually Transmitted Infection Treatment Guidelines. Clin. Infect. Dis. 2022, 74 (Suppl. S2), S95–S111. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Chen, W.; Yu, Y.; Lan, Y.; Xie, Q.; Liao, Y.; Wu, X.; Tang, S.; Qin, X.; Zheng, H. Emergence and genomic characterization of Neisseria gonorrhoeae isolates with high levels of ceftriaxone and azithromycin resistance in Guangdong, China, from 2016 to 2019. Microbiol. Spectr. 2022, 10, e0157022. [Google Scholar] [CrossRef]
- WHO Guidelines for the Treatment of Neisseria gonorrhoeae. 1 January 2016. Available online: https://www.who.int/publications/i/item/9789241549691 (accessed on 12 April 2023).
- Chisholm, S.A.; Unemo, M.; Quaye, N.; Johansson, E.; Cole, M.J.; Ison, C.A.; Van de Laar, M.J. Molecular epidemiological typing within the European Gonococcal Antimicrobial Resistance Surveillance Programme reveals predominance of a multidrug-resistant clone. Euro Surveill. 2013, 183, 20358. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Molecular Typing of Neisseria gonorrhoeae—A Study of 2013 Isolates; ECDC: Stockholm, Sweden, 2018; Available online: https://www.ecdc.europa.eu/sites/portal/files/documents/Molecular-typing-N-gonorrhoeae-web.pdf (accessed on 12 April 2023).
- Harris, S.R.; Cole, M.J.; Spiteri, G.; Sánchez-Busó, L.; Golparian, D.; Jacobsson, S.; Goater, R.; Abudahab, K.; Yeats, C.A.; Bercot, B.; et al. Public health surveillance of multidrug-resistant clones of Neisseria gonorrhoeae in Europe: A genomic survey. Lancet Infect. Dis. 2018, 18, 758–768. [Google Scholar] [CrossRef]
- Sánchez-Busó, L.; Cole, M.J.; Spiteri, G.; Day, M.; Jacobsson, S.; Golparian, D.; Sajedi, N.; Yeats, C.A.; Abudahab, K.; Underwood, A.; et al. Europe-wide expansion and eradication of multidrug-resistant Neisseria gonorrhoeae lineages: A genomic surveillance study. Lancet Microbe 2022, 3, e452–e463. [Google Scholar] [CrossRef]
- Shaskolskiy, B.; Dementieva, E.; Kandinov, I.; Chestkov, A.; Kubanov, A.; Deryabin, D.; Gryadunov, D. Genetic diversity of Neisseria gonorrhoeae multi-antigen sequence types in Russia and Europe. Int. J. Infect. Dis. 2020, 93, 1–8. [Google Scholar] [CrossRef]
- Shaskolskiy, B.L.; Kandinov, I.D.; Chestkov, A.V.; Solomka, V.S.; Kubanov, A.A.; Deryabin, D.G.; Gryadunov, D.A.; Dementieva, E.I. Comparative phylogenetic analysis of Neisseria gonorrhoeae clinical isolates in Russia, European Union, and Japan. Bull. Russ. State Med. Univ. 2020, 1, 5–13. [Google Scholar] [CrossRef]
- Kandinov, I.; Shaskolskiy, B.; Kravtsov, D.; Vinokurova, A.; Gorshkova, S.; Kubanov, A.; Solomka, V.; Shagabieva, J.; Deryabin, D.; Dementieva, E.; et al. Azithromycin susceptibility testing and molecular investigation of Neisseria gonorrhoeae isolates collected in Russia, 2020–2021. Antibiotics 2023, 12, 170. [Google Scholar] [CrossRef]
- Kubanov, A.; Solomka, V.; Plakhova, X.; Chestkov, A.; Petrova, N.; Shaskolskiy, B.; Dementieva, E.; Leinsoo, A.; Gryadunov, D.; Deryabin, D. Summary and trends of the Russian Gonococcal Antimicrobial Surveillance Programme, 2005 to 2016. J. Clin. Microbiol. 2019, 57, e02024-18. [Google Scholar] [CrossRef]
- Shaskolskiy, B.; Dementieva, E.; Kandinov, I.; Filippova, M.; Petrova, N.; Plakhova, X.; Chestkov, A.; Kubanov, A.; Deryabin, D.; Gryadunov, D. Resistance of Neisseria gonorrhoeae isolates to beta-lactams (benzylpenicillin and ceftriaxone) in Russia, 2015-2017. PLoS ONE 2019, 14, e0220339. [Google Scholar] [CrossRef]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 13.0. 2023. Available online: https://www.eucast.org/clinical_breakpoints (accessed on 12 April 2023).
- Shaskolskiy, B.; Kandinov, I.; Kravtsov, D.; Vinokurova, A.; Gorshkova, S.; Filippova, M.; Kubanov, A.; Solomka, V.; Deryabin, D.; Dementieva, E.; et al. Hydrogel droplet microarray for genotyping antimicrobial resistance determinants in Neisseria gonorrhoeae isolates. Polymers 2021, 13, 3889. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, S.A.; Dave, J.; Ison, C.A. High-level azithromycin resistance occurs in Neisseria gonorrhoeae as a result of a single point mutation in the 23S rRNA genes. Antimicrob. Agents Chemother. 2010, 54, 3812–3816. [Google Scholar] [CrossRef] [PubMed]
- Cousin, S., Jr.; Whittington, W.L.; Roberts, M.C. Acquired macrolide resistance genes in pathogenic Neisseria spp. isolated between 1940 and 1987. Antimicrob. Agents Chemother. 2003, 47, 3877–3880. [Google Scholar] [CrossRef] [PubMed]
- Whiley, D.M.; Kundu, R.L.; Jennison, A.V.; Buckley, C.; Limnios, A.; Hogan, T.; Enriquez, R.; El Nasser, J.; George, C.R.; Lahra, M.M. Azithromycin-resistant Neisseria gonorrhoeae spreading amongst men who have sex with men (MSM) and heterosexuals in New South Wales, Australia, 2017. J. Antimicrob. Chemother. 2018, 73, 1242–1246. [Google Scholar] [CrossRef] [PubMed]
- Belkacem, A.; Jacquier, H.; Goubard, A.; Mougari, F.; La Ruche, G.; Patey, O.; Micaëlo, M.; Semaille, C.; Cambau, E.; Bercot, B. Molecular epidemiology and mechanisms of resistance of azithromycin-resistant Neisseria gonorrhoeae isolated in France during 2013-14. J. Antimicrob. Chemother. 2016, 71, 2471–2478. [Google Scholar] [CrossRef]
- Kandinov, I.; Dementieva, E.; Kravtsov, D.; Chestkov, A.; Kubanov, A.; Solomka, V.; Deryabin, D.; Gryadunov, D.; Shaskolskiy, B. Molecular typing of Neisseria gonorrhoeae clinical isolates in Russia, 2018–2019: A link between penA alleles and NG-MAST types. Pathogens 2020, 9, 941. [Google Scholar] [CrossRef]
- Zhou, Q.; Liu, J.; Chen, S.; Xu, W.; Han, Y.; Yin, Y. The accuracy of molecular detection targeting the mutation C2611T for detecting moderate-level azithromycin resistance in Neisseria gonorrhoeae: A systematic review and meta-analysis. Antibiotics 2021, 10, 1027. [Google Scholar] [CrossRef]
- Trembizki, E.; Doyle, C.; Jennison, A.; Smith, H.; Bates, J.; Lahra, M.; Whiley, D. A Neisseria gonorrhoeae strain with a meningococcal mtrR sequence. J. Med. Microbiol. 2014, 63, 1113–1115. [Google Scholar] [CrossRef]
- Demczuk, W.; Martin, I.; Peterson, S.; Bharat, A.; Van Domselaar, G.; Graham, M.; Lefebvre, B.; Allen, V.; Hoang, L.; Tyrrell, G.; et al. Genomic epidemiology and molecular resistance mechanisms of azithromycin-resistant Neisseria gonorrhoeae in Canada from 1997 to 2014. J. Clin Microbiol. 2016, 54, 1304–1313. [Google Scholar] [CrossRef]
- Bignell, C.; Unemo, M. European guideline on the diagnosis and treatment of gonorrhoea in adults. Int. J. STD AIDS 2013, 24, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, J.; Lippert, K.; Pleininger, S.; Stöger, A.; Hasenberger, P.; Stadlbauer, S.; Heger, F.; Eigentler, A.; Geusau, A.; Indra, A.; et al. Association of phylogenomic relatedness among Neisseria gonorrhoeae strains with antimicrobial resistance, Austria, 2016–2020. Emerg. Infect. Dis. 2022, 28, 1694–1698. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.J.; Thomas, J.C.; Schmerer, M.W.; Cartee, J.C.; St Cyr, S.; Schlanger, K.; Kersh, E.N.; Raphael, B.H.; Gernert, K.M. Global emergence and dissemination of Neisseria gonorrhoeae ST-9363 isolates with reduced susceptibility to azithromycin. Genome Biol. Evol. 2022, 14, evab287. [Google Scholar] [CrossRef] [PubMed]
- Sawatzky, P.; Demczuk, W.; Lefebvre, B.; Allen, V.; Diggle, M.; Hoang, L.; Van Caeseele, P.; Haldane, D.; Minion, J.; Mulvey, M.R.; et al. Increasing azithromycin resistance in Neisseria gonorrhoeae due to NG-MAST 12302 clonal spread in Canada, 2015 to 2018. Antimicrob. Agents Chemother. 2022, 66, e0168821. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyzed Locus | Detected Determinants | Proportion of Isolates | |||
|---|---|---|---|---|---|
| Year 2018 | Year 2019 | Year 2020 | Year 2021 | ||
| 23S rRNA | C2611 (4/4 alleles) | 100% | 100% | 98% | 100% |
| C2611T (4/4 alleles) | - | - | 2% | - | |
| mtrR (promoter) | −35 (wt) | 82.8% | 90% | 76% | 64.8% |
| −35 delA | 16.5% | 10% | 10% | 13% | |
| −35 mosaic | - | - | - | 3.7% | |
| −35 mosaic/delA | 0.7% | - | 14% | 18.5% | |
| −10 (wt) | 99.3% | 100% | 100% | 100% | |
| −10 insT | 0.7% | - | - | - | |
| mtrR (coding region) | Ala39 (wt) | 83% | 78% | 80% | 55.5% |
| Thr39 | 17% | 22% | 20% | 44.5% | |
| Gly45 (wt) | 83% | 79% | 83% | 90.8% | |
| Asp45 | 8% | 9% | 8% | 3.7% | |
| Ser45 | 9% | 12% | 9% | 5.5% | |
| Ala86 (wt) | 24.5% | 24% | 10% | 13% | |
| Thr86 | 75.5% | 76% | 90% | 87% | |
| His105 (wt) | 99.3% | 98.4% | 82% | 67% | |
| Tyr105 | 0.7% | 1.6% | 18% | 33% | |
| mtrD | Ser821/Lys823 (wt) | 99.3% | 98.4% | 82% | 67% |
| Ala821/Glu823 (mosaic) | 0.7% | 1.6% | 18% | 33% | |
| Total number of isolates for each year | 151 | 122 | 101 | 54 | |
| Number of phenotypically azithromycin-resistant isolates | 0 | 0 | 17 (16.8%) | 5 (9.3%) | |
| Geno- Group | Number of Isolates (%) | NG-MAST (Number of Isolates) | Azithromycin Susceptibility | Median MICazm, mg/L |
|---|---|---|---|---|
| G10800 | 103 (24.0%) | 228 (35); 807 (34); 1544 (3); 3321 (2); 3369 (1); 4570 (2); 5941 (5); 6967 (1); 9570 (6); 9576 (3); 15619 (1); 16177 (1); 18246 (2); 18894 (5); 19573 (1); 20267 (1) | S (all isolates) | 0.12 |
| G1993 | 43 (10.0%) | 1993 (23); 5714 (6); 14627 (3); 16174 (2); 16411 (2); 17027 (4); 18247 (1); 20273 (2) | S (42 isolates) R (1 isolate) | 0.12 |
| G12302 | 30 (7.0%) | 6765 (1); 12302 (7); 14502 (2); 16953 (2); 17380 (6); 18787 (2); 20274 (1); 20275 (1); 20279 (1); 20291 (1); 20293 (1) 20294 (1); 20602 (3); 20615 (1) | S (13 isolates) R (17 isolates) | 2.0 |
| G387 | 22 (5.1%) | 1152 (2); 5185 (1); 5734 (2); 9486 (13); 14825 (1); 17017 (2); 17532 (1) | S (all isolates) | 0.06 |
| G1407 | 14 (3.3%) | 2212 (3); 3149 (1); 4706 (2); 5622 (1); 10025 (4); 20278 (1); 20283 (1); 20285 (1) | S (all isolates) | 0.25 |
| G14020 | 9 (2.1%) | 14020 (9) | S (all isolates) | 0.06 |
| G12542 | 3 (0.7%) | 12542 (3) | S (all isolates) | - |
| G5441 | 2 (0.5%) | 13994 (1); 19574 (1) | S (all isolates) | - |
| G799 | 2 (0.5%) | 19588 (2) | S (all isolates) | - |
| G4186 | 2 (0.5%) | 18953 (2) | S (all isolates) | - |
| G5624 | 2 (0.5%) | 5624 (2) | S (1 isolate), R (1 isolate) | - |
| G9918 | 2 (0.5%) | 20614 (2) | S (all isolates) | - |
| G10386 | 2 (0.5%) | 19050 (2) | S (all isolates) | - |
| G14700 | 2 (0.5%) | 14700 (2) | S (all isolates) | - |
| G15589 | 2 (0.5%) | 15589 (2) | S (all isolates) | - |
| G5 | 1 (0.2%) | 21 (1) | S | - |
| G758 | 1 (0.2%) | 20280 (1) | S | - |
| G3785 | 1 (0.2%) | 18131 (1) | S | - |
| G2400 | 1 (0.2%) | 9184 (1) | S | - |
| G12818 | 1 (0.2%) | 18245 (1) | S | - |
| G6226(RUS) | 18 (4.2%) | 6226 (9); 13055 (4); 13058 (2); 6238 (1); 16185 (1); 17026 (1) | S (all isolates) | 0.12 |
| G5042(RUS) | 15 (3.5%) | 5042 (4); 9574 (1); 12449 (3); 14940 (3); 17020 (2); 14932 (1); 20292 (1) | S (all isolates) | 0.09 |
| G14942(RUS) | 9 (2.1%) | 14942 (8); 16410 (1) | S (all isolates) | 0.12 |
| G19572(RUS) | 7 (1.6%) | 19572 (6), 19576 (1) | S (6 isolates), R (1 isolate) | 0.12 |
| Un-grouped | 133 (33.2%) | - | S (131 isolates) R (2 isolates) | 0.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kandinov, I.; Dementieva, E.; Filippova, M.; Vinokurova, A.; Gorshkova, S.; Kubanov, A.; Solomka, V.; Shagabieva, J.; Deryabin, D.; Shaskolskiy, B.; et al. Emergence of Azithromycin-Resistant Neisseria gonorrhoeae Isolates Belonging to the NG-MAST Genogroup 12302 in Russia. Microorganisms 2023, 11, 1226. https://doi.org/10.3390/microorganisms11051226
Kandinov I, Dementieva E, Filippova M, Vinokurova A, Gorshkova S, Kubanov A, Solomka V, Shagabieva J, Deryabin D, Shaskolskiy B, et al. Emergence of Azithromycin-Resistant Neisseria gonorrhoeae Isolates Belonging to the NG-MAST Genogroup 12302 in Russia. Microorganisms. 2023; 11(5):1226. https://doi.org/10.3390/microorganisms11051226
Chicago/Turabian StyleKandinov, Ilya, Ekaterina Dementieva, Marina Filippova, Alexandra Vinokurova, Sofya Gorshkova, Alexey Kubanov, Victoria Solomka, Julia Shagabieva, Dmitry Deryabin, Boris Shaskolskiy, and et al. 2023. "Emergence of Azithromycin-Resistant Neisseria gonorrhoeae Isolates Belonging to the NG-MAST Genogroup 12302 in Russia" Microorganisms 11, no. 5: 1226. https://doi.org/10.3390/microorganisms11051226
APA StyleKandinov, I., Dementieva, E., Filippova, M., Vinokurova, A., Gorshkova, S., Kubanov, A., Solomka, V., Shagabieva, J., Deryabin, D., Shaskolskiy, B., & Gryadunov, D. (2023). Emergence of Azithromycin-Resistant Neisseria gonorrhoeae Isolates Belonging to the NG-MAST Genogroup 12302 in Russia. Microorganisms, 11(5), 1226. https://doi.org/10.3390/microorganisms11051226