
Characterization of the Bacterial Communities Inhabiting Tropical Propolis of Puerto Rico


 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Areas and Hive Description and Sampling
2.2. Cultivation of Bacteria from Propolis
2.3. DNA Extraction from Isolated Bacteria, 16S rRNA Amplification, and Sequencing
2.4. Antimicrobial Activity of Propolis Extract
2.5. Antimicrobial Activity of Bacterial Isolates
2.6. Total DNA Extraction from Propolis and Library Preparation
2.7. Clustering of 16S rRNA Amplicons
2.8. Metataxonomic and Diversity Analyses
3. Results
3.1. Cultivation and Identification of Bacteria
3.2. Antimicrobial Activity of Propolis Extracts and Bacterial Isolates
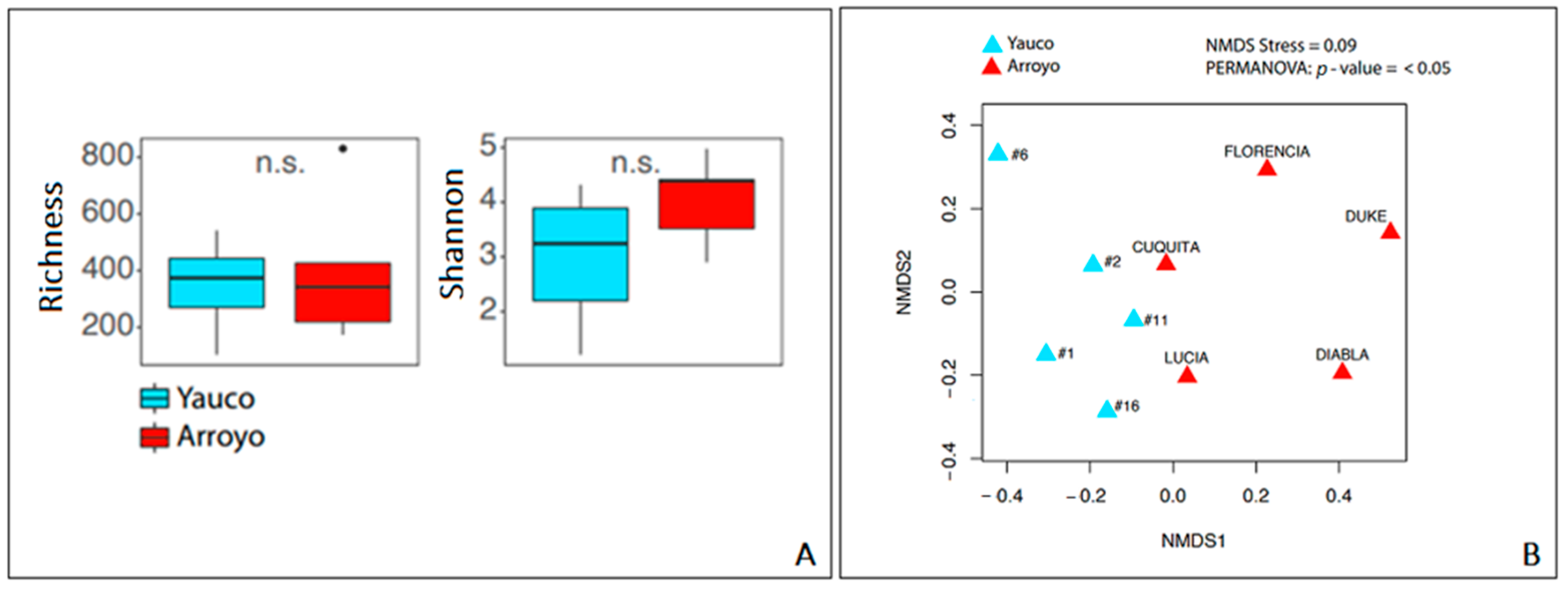
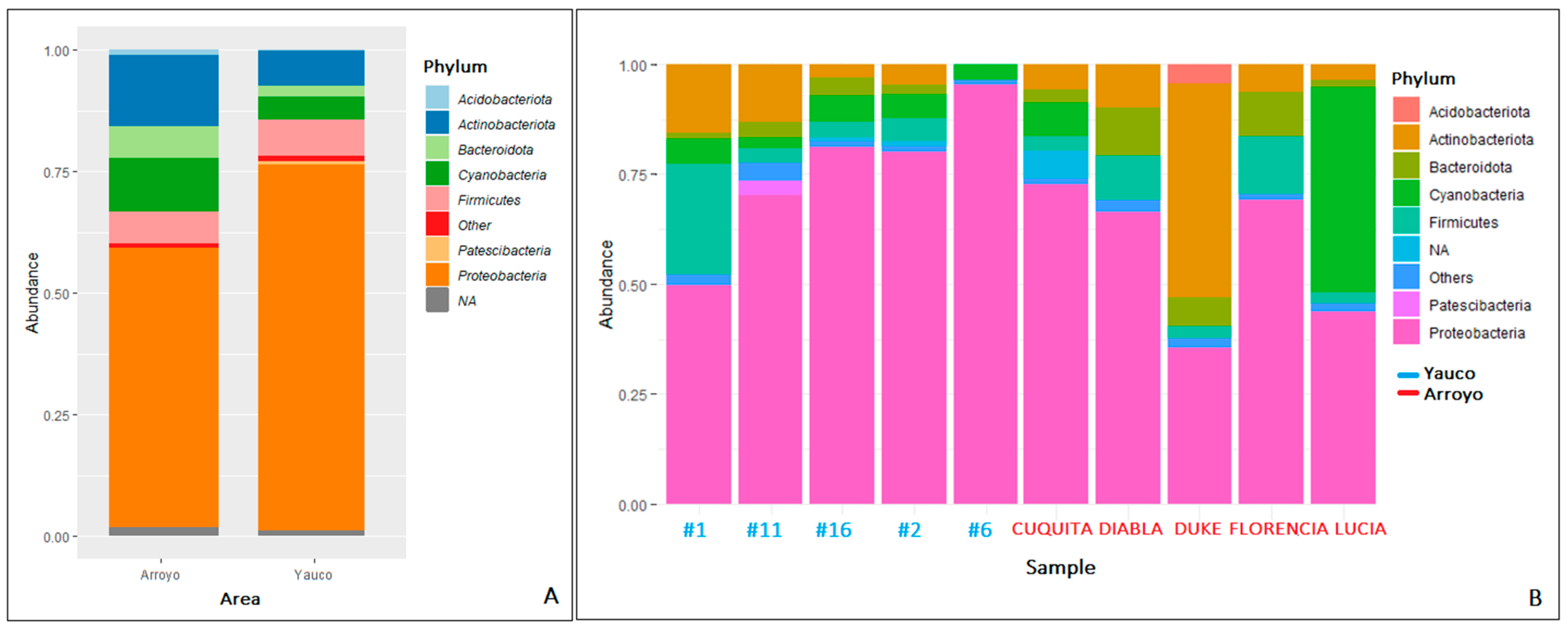
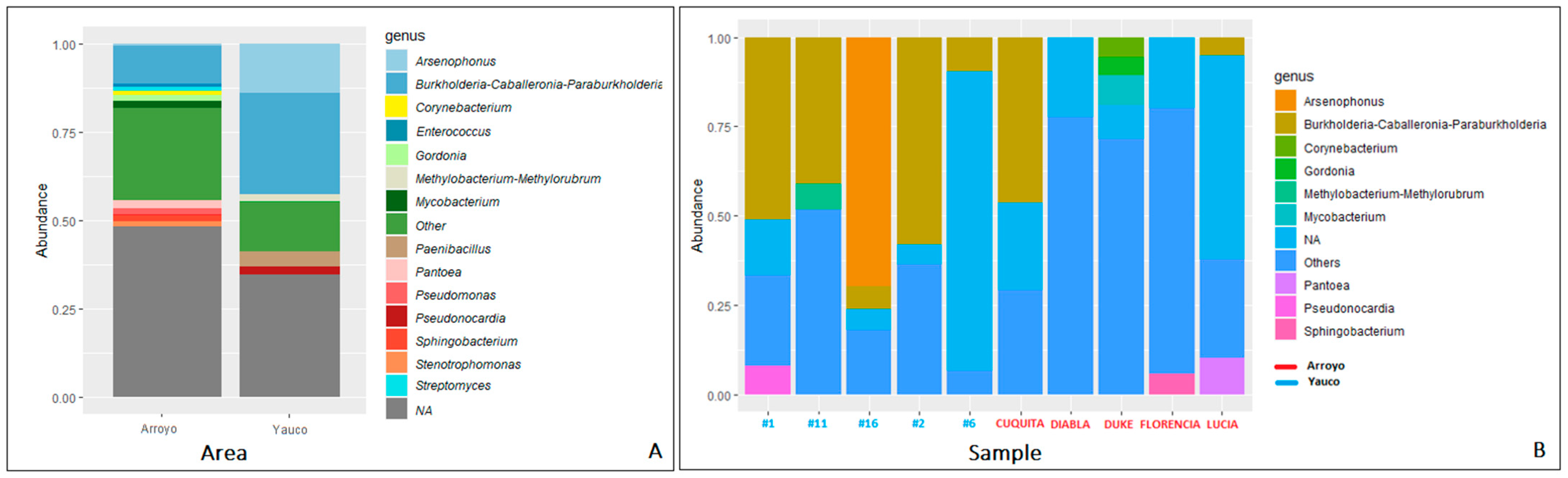
3.3. Diversity and Metataxonomic Analyses of the Bacterial Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simone-Finstrom, M.; Spivak, M. Propolis and bee health: The natural history and significance of resin use by honey bees. Apidologie 2010, 41, 295–311. [Google Scholar] [CrossRef]
- Przybyłek, I.; Karpiński, T.M. Antibacterial Properties of Propolis. Molecules 2019, 24, 2047. [Google Scholar] [CrossRef] [PubMed]
- Yosri, N.; Abd El-Wahed, A.A.; Ghonaim, R.; Khattab, O.M.; Sabry, A.; Ibrahim, M.A.A.; Moustafa, M.F.; Guo, Z.; Zou, X.; Algethami, A.F.M.; et al. Anti-Viral and Immunomodulatory Properties of Propolis: Chemical Diversity, Pharmacological Properties, Preclinical and Clinical Applications, and In Silico Potential against SARS-CoV-2. Foods 2021, 10, 1776. [Google Scholar] [CrossRef]
- Casalone, E.; Cavalieri, D.; Daly, G.; Vitali, F.; Perito, B. Propolis hosts a diverse microbial community. World J. Microbiol. Biotechnol. 2020, 36, 50. [Google Scholar] [CrossRef] [PubMed]
- Zabaiou, N.; Fouache, A.; Trousson, A.; Baron, S.; Zellagui, A.; Lahouel, M.; Lobaccaro, J.M.A. Biological properties of propolis extracts: Something new from an ancient product. Chem. Phys. Lipids 2017, 207, 214–222. [Google Scholar] [CrossRef]
- Kasote, D.; Bankova, V.; Viljoen, A.M. Propolis: Chemical diversity and challenges in quality control. Phytochem. Rev. 2022, 21, 1887–1911. [Google Scholar] [CrossRef]
- Silva-Carvalho, R.; Baltazar, F.; Almeida-Aguiar, C. Propolis: A Complex Natural Product with a Plethora of Biological Activities That Can Be Explored for Drug Development. Evid. Based Complement Altern. Med. 2015, 2015, 206439. [Google Scholar] [CrossRef]
- Kuropatnicki, A.K.; Szliszka, E.; Krol, W. Historical Aspects of Propolis Research in Modern Times. Evid. Based Complement Altern. Med. 2013, 2013, 964149. [Google Scholar] [CrossRef]
- Stojanović, S.T.; Najman, S.J.; Popov, B.B.; Najman, S.S. Propolis: Chemical composition, biological and pharmacological activity—A review. Acta Med. Medianae 2020, 59, 108–113. [Google Scholar] [CrossRef]
- Siheri, W.; Alenezi, S.; Tusiimire, J.; Watson, D.G. The Chemical and Biological Properties of Propolis. In Bee Products—Chemical and Biological Properties; Springer: Cham, Switzerland, 2017; pp. 137–178. [Google Scholar] [CrossRef]
- Simone-Finstrom, M. Social Immunity and the Superorganism: Behavioral Defenses Protecting Honey Bee Colonies from Pathogens and Parasites. Bee World 2017, 94, 21–29. [Google Scholar] [CrossRef]
- Saelao, P.; Borba, R.S.; Ricigliano, V.; Spivak, M.; Simone-Finstrom, M. Honeybee microbiome is stabilized in the presence of propolis. Biol. Lett. 2020, 16, 20200003. [Google Scholar] [CrossRef] [PubMed]
- Vojvodic, S.; Rehan, S.M.; Anderson, K.E. Microbial Gut Diversity of Africanized and European Honey Bee Larval Instars. PLoS ONE 2013, 8, e72106. [Google Scholar] [CrossRef] [PubMed]
- Raymann, K.; Moran, N.A. The role of the gut microbiome in health and disease of adult honey bee workers. Curr. Opin. Insect Sci. 2018, 26, 97–104. [Google Scholar] [CrossRef]
- Bonilla-Rosso, G.; Engel, P. Functional roles and metabolic niches in the honey bee gut microbiota. Curr. Opin. Microbiol. 2018, 43, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.E.; Sheehan, T.H.; Mott, B.M.; Maes, P.; Snyder, L.; Schwan, M.R.; Walton, A.; Jones, B.M.; Corby-Harris, V. Microbial Ecology of the Hive and Pollination Landscape: Bacterial Associates from Floral Nectar, the Alimentary Tract and Stored Food of Honey Bees (Apis mellifera). PLoS ONE 2013, 8, e83125. [Google Scholar] [CrossRef]
- Corby-Harris, V.; Maes, P.; Anderson, K.E. The Bacterial Communities Associated with Honey Bee (Apis mellifera) Foragers. PLoS ONE 2014, 9, e95056. [Google Scholar] [CrossRef]
- Donkersley, P.; Rhodes, G.; Pickup, R.W.; Jones, K.C.; Wilson, K. Bacterial communities associated with honeybee food stores are correlated with land use. Ecol. Evol. 2018, 8, 4743–4756. [Google Scholar] [CrossRef]
- Santorelli, L.A.; Wilkinson, T.; Abdulmalik, R.; Rai, Y.; Creevey, C.J.; Huws, S.; Gutierrez-Merino, J. Beehives possess their own distinct microbiomes. Environ. Microbiome 2023, 18, 1. [Google Scholar] [CrossRef]
- Grubbs, K.J.; Scott, J.J.; Budsberg, K.J.; Read, H.; Balser, T.C.; Currie, C.R. Unique Honey Bee (Apis mellifera) Hive Component-Based Communities as Detected by a Hybrid of Phospholipid Fatty-Acid and Fatty-Acid Methyl Ester Analyses. PLoS ONE 2015, 10, e0121697. [Google Scholar] [CrossRef]
- Garcia-Mazcorro, J.F.; Kawas, J.R.; Marroquin-Cardona, A.G. Descriptive Bacterial and Fungal Characterization of Propolis Using Ultra-High-Throughput Marker Gene Sequencing. Insects 2019, 10, 402. [Google Scholar] [CrossRef]
- Sallemi, S.; Lekired, A.; Korbi, N.; Saadouli, I.; Cherif, A.; Zidi, I.; Klibi, N.; Ouzari, H.-I.; Mosbah, A. Fungal Community Investigation from Propolis Natural Products: Diversity and Antibacterial Activities Evaluation. Evid. Based Complem. Altern. Med. 2022, 2022, 7151655. [Google Scholar] [CrossRef] [PubMed]
- Ersoy Omeroglu, E.; Keriman Arserim-Uçar, D.; Yegin, Z.; Çağlayan, N.; Nur Zafer Yurt, M.; Busra Tasbasi, B.; Esma Acar, E.; Ucak, S.; Cengiz Ozalp, V.; Sudagidan, M. Determination of Bacterial Diversity of Propolis Microbiota. Chem. Biodiv. 2023, 20, e202201182. [Google Scholar] [CrossRef] [PubMed]
- Kerr, W.E. The history of the introduction of African bees in Brazil. South African Bee J. 1967, 39, 33–35. [Google Scholar]
- Hall, H.G. Parental analysis of introgressive hybridization between African and European honeybees using nuclear DNA RFLPs. Genetics 1990, 125, 611–621. [Google Scholar] [CrossRef]
- Rivera-Marchand, B.; Oskay, D.; Giray, T. Gentle Africanized bees on an oceanic island. Evol. Appl. 2012, 5, 746–756. [Google Scholar] [CrossRef]
- Avalos, A.; Pan, H.; Li, C.; Acevedo-Gonzalez, J.P.; Rendon, G.; Fields, C.J.; Brown, P.J.; Giray, T.; Robinson, G.E.; Hudson, M.E.; et al. A soft selective sweep during rapid evolution of gentle behaviour in an Africanized honeybee. Nat. Commun. 2017, 8, 1550. [Google Scholar] [CrossRef]
- Hettiarachchi, S.A.; Lee, S.-J.; Lee, Y.; Kwon, Y.-K.; Zoysa, M.; de Moon, S.; Jo, E.; Kim, E.; Kang, D.-H.; Heo, S.-J.; et al. A Rapid and Efficient Screening Method for Antibacterial Compound-Producing Bacteria. J. Microbiol. Biotechnol. 2017, 27, 1441–1448. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Wright, E.S.; Yilmaz, L.S.; Noguera, D.R. DECIPHER, a Search-Based Approach to Chimera Identification for 16S rRNA Sequences. Appl. Environ. Microbiol. 2012, 78, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. PHYLOSEQ: A Bioconductor package for handling and analysis of high-throughput phylogenetic sequence data. In Biocomputing; World Scientific Publishing Co. Pte. Ltd.: Singapore, 2012; pp. 235–246. [Google Scholar] [CrossRef]
- Lahti, L.; Shetty, S. Introduction to the Microbiome R Package. Bioconductor. 2018. Available online: https://www.bioconductor.org/packages/release/bioc/html/microbiome.html (accessed on 15 October 2022).
- Ssekagiri, A.T.; Sloan, W.; Ijaz, U.Z. MicrobiomeSeq: An R package for analysis of microbial communities in an environmental context. In ISCB Africa; ASBCB: Entebbe, Uganda; ISCB: Kumasi, Ghana, 2017. [Google Scholar]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.A.; Al-Ghamdi, A.A.; Ghramh, H.A.; Ansari, M.J.; Ali, H.; Alamri, S.A.; Al-Kahtani, S.N.; Adgaba, N.; Qasim, M.; Hafeez, M. Structural diversity and functional variability of gut microbial communities associated with honeybees. Microb. Pathog. 2020, 138, 103793. [Google Scholar] [CrossRef]
- Karthick, P.; Mohanraju, R. Antimicrobial compounds produced by Lysinibacillus odysseyi epiphytic bacteria associated with red algae. Braz. J. Microbiol. 2020, 51, 1683–1690. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Gupta, R.S. A phylogenomic and comparative genomic framework for resolving the polyphyly of the genus Bacillus: Proposal for six new genera of Bacillus species, Peribacillus gen. nov., Cytobacillus gen. nov., Mesobacillus gen. nov., Neobacillus gen. nov., Metabacillus gen. nov. and Alkalihalobacillus gen. nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 406–438. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, S.; Merrill, S.A. Staphylococcus Saprophyticus; StatPearls: Tampa, FL, USA, 2022. [Google Scholar] [PubMed]
- Ganeshprasad, D.N.; Lone, J.K.; Jani, K.; Shouche, Y.S.; Khan, K.A.; Sayed, S.; Shukry, M.; Dar, S.A.; Mushtaq, M.; Sneharani, A.H. Gut Bacterial Flora of Open Nested Honeybee, Apis florea. Front. Ecol. Evol. 2022, 10, 837381. [Google Scholar] [CrossRef]
- Schäfer, J.; Martin, K.; Kämpfer, P. Citricoccus parietis sp. nov., isolated from a mould-colonized wall and emended description of Citricoccus alkalitolerans Li et al. 2005. Int. J. Syst. Evol. Microbiol. 2010, 60, 271–274. [Google Scholar] [CrossRef]
- Surek, M.; Fachi, M.M.; de Fátima Cobre, A.; de Oliveira, F.F.; Pontarolo, R.; Crisma, A.R.; de Souza, W.M.; Felipe, K.B. Chemical composition, cytotoxicity, and antibacterial activity of propolis from Africanized honeybees and three different Meliponini species. J. Ethnopharmacol. 2021, 269, 113662. [Google Scholar] [CrossRef]
- Seidel, V.; Peyfoon, E.; Watson, D.G.; Fearnley, J. Comparative study of the antibacterial activity of propolis from different geographical and climatic zones. Phytotherapy Res. 2008, 22, 1256–1263. [Google Scholar] [CrossRef]
- Brudzynski, K. Honey as an Ecological Reservoir of Antibacterial Compounds Produced by Antagonistic Microbial Interactions in Plant Nectars, Honey and Honey Bee. Antibiotics 2021, 10, 551. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, B.; Chávez, A.; Forero, A.; García-Huante, Y.; Romero, A.; Sánchez, M.; Rocha, D.; Sánchez, B.; Rodríguez-Sanoja, R.; Sánchez, S.; et al. Production of microbial secondary metabolites: Regulation by the carbon source. Crit. Rev. Microbiol. 2010, 36, 146–167. [Google Scholar] [CrossRef] [PubMed]
- El-Hamshary, O.I.M.; Khattab, A.A. Evaluation of antimicrobial activity of Bacillus subtilis and Bacillus cereus and their fusants against Fusarium solani. Res. J. Cell Mol. Biol. 2008, 2, 24–29. [Google Scholar]
- Risoen, P.A.; Ronning, P.; Hegna, I.K.; Kolsto, A.B. Characterization of a broad range antimicrobial substance from Bacillus cereus. J. Appl. Microbiol. 2004, 96, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, M.; Soran, H.; Beyatli, Y. Antimicrobial activities of some Bacillus spp. strains isolated from the soil. Microbiol. Res. 2006, 161, 127–131. [Google Scholar] [CrossRef]
- Genersch, E. American Foulbrood in honeybees and its causative agent, Paenibacillus larvae. J. Invertebr. Pathol. 2010, 103, S10–S19. [Google Scholar] [CrossRef]
- Dong, Z.X.; Li, H.Y.; Chen, Y.F.; Wang, F.; Deng, X.Y.; Lin, L.B.; Zhang, Q.L.; Li, J.L.; Guo, J. Colonization of the gut microbiota of honey bee (Apis mellifera) workers at different developmental stages. Microbiol. Res. 2020, 231, 126370. [Google Scholar] [CrossRef]
- Scheiner, R.; Strauß, S.; Thamm, M.; Farré-Armengol, G.; Junker, R.R. The Bacterium Pantoea ananatis Modifies Behavioral Responses to Sugar Solutions in Honeybees. Insects 2020, 11, 692. [Google Scholar] [CrossRef]
- Walterson, A.M.; Stavrinides, J. Pantoea: Insights into a highly versatile and diverse genus within the Enterobacteriaceae. FEMS Microbiol. Rev. 2015, 39, 968–984. [Google Scholar] [CrossRef]
- Estrada-de los Santos, P.; Palmer, M.; Chávez-Ramírez, B.; Beukes, C.; Steenkamp, E.; Briscoe, L.; Khan, N.; Maluk, M.; Lafos, M.; Humm, E.; et al. Whole genome analyses suggests that Burkholderia sensu lato contains two additional novel genera (Mycetohabitans gen. nov., and Trinickia gen. nov.): Implications for the evolution of diazotrophy and nodulation in the Burkholderiaceae. Genes 2018, 9, 389. [Google Scholar] [CrossRef]
- Yañez, O.; Gauthier, L.; Chantawannakul, P.; Neumann, P. Endosymbiotic bacteria in honey bees: Arsenophonus spp. are not transmitted transovarially. FEMS Microbiol. Lett. 2016, 363, fnw147. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.R.; Tarpy, D.R.; Van Engelsdorp, D.; Chauzat, M.P.; Cox-Foster, D.L.; Delaplane, K.S.; Neumann, P.; Pettis, J.S.; Rogers, R.E.L.; Shutler, D. Colony Collapse Disorder in context. BioEssays 2010, 32, 845–846. [Google Scholar] [CrossRef] [PubMed]
- Grubbs, K.J.; May, D.S.; Sardina, J.A.; Dermenjian, R.K.; Wyche, T.P.; Pinto-Tomás, A.A.; Clardy, J.; Currie, C.R. Pollen Streptomyces produce antibiotic that inhibits the Honey Bee pathogen Paenibacillus larvae. Front. Microbiol. 2021, 12, 632637. [Google Scholar] [CrossRef] [PubMed]
- Hartmans, S.; de Bont, J.A.M.; Stackebrandt, E. The Genus Mycobacterium—Nonmedical. In The Prokaryotes; Springer: New York, NY, USA, 2006; pp. 889–918. [Google Scholar] [CrossRef]
- Schulze-Röbbecke, R. Mycobacteria in the environment. Immunitat Infektion 1993, 21, 126–131. [Google Scholar] [PubMed]
- Olofsson, T.C.; Vásquez, A.; Sammataro, D.; Macharia, J. A scientific note on the lactic acid bacterial flora within the honeybee subspecies Apis mellifera (Buckfast), A. m. scutellata, A. m. mellifera, and A. m. monticola. Apidologie 2011, 42, 696–699. [Google Scholar] [CrossRef]
- Klein, S.; Cabirol, A.; Devaud, J.M.; Barron, A.B.; Lihoreau, M. Why Bees are so vulnerable to environmental stressors. Trends Ecol. Evol. 2017, 32, 268–278. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area | Sample (Hive) | Metataxonomic | Cultivation | Ethanolic Extracts |
|---|---|---|---|---|
| Yauco | #1 | ✓ | ✓ | ✓ |
| #2 | ✓ | ✓ | ✓ | |
| #4 | ✓ | |||
| #6 | ✓ | |||
| #8 | ✓ | ✓ | ||
| #11 | ✓ | ✓ | ||
| #13 | ✓ | ✓ | ||
| #16 | ✓ | |||
| Arroyo | AKIRA | ✓ | ||
| CUQUITA | ✓ | ✓ | ||
| DIABLA | ✓ | ✓ | ||
| DUKE | ✓ | ✓ | ||
| FLORENCIA | ✓ | ✓ | ✓ | |
| LUCIA | ✓ | ✓ | ✓ |
| Area | Hive | Untreated (CFU/g ± Sd) | Heat-Treated (CFU/g ± Sd) |
|---|---|---|---|
| Arroyo | Lucia | 1.0 × 103 ± 1.5 × 102 | nm |
| Florencia | 2.7 × 104 ± 6.1 × 103 | nd | |
| Yauco | #1 | 1.2 × 107 ± 3.1 × 106 | nm |
| #2 | 4.0 × 106 ± 9.8 × 105 | 2.5 × 104 ± 1.2 × 104 | |
| #8 | 1.0 × 106 ± 2.6 × 105 | 7.5 × 103 ± 1.1 × 104 | |
| #13 | 2.8 × 104 ± 2.0 × 103 | nm |
| Area | Strain/ Morphotype * | Organisms with the Most Similar 16S rRNA Sequences | Identity (%) |
|---|---|---|---|
| Yauco | #1C | Stenotrophomonas maltophilia | 98.59 |
| #2C | Staphyloccoccus saprophyticus | 98.30 | |
| #8C | Enterobacter ludwigii | 98.58 | |
| #13A | Curtobacterium luteum | 98.43 | |
| #13D | Oceanobacillus profundus | 98.24 | |
| #13F | Curtobacterium oceanosedimentum | 98.00 | |
| #13G | Rhodococcus baikonurensis | 99.02 | |
| Arroyo | FLORENCIA A | Luteimonas sp. | 98.09 |
| FLORENCIA B | Alkalihalobacillus xiaoxiensis | 98.58 | |
| FLORENCIA C | Citricoccus parietis | 98.06 | |
| FLORENCIA E | Staphylococcus kloosii | 99.53 | |
| FLORENCIA F | Bacillus cereus | 99.21 | |
| LUCIA A | Bacillus thuringensis | 97.21 | |
| LUCIA C | Oceanobacillus sojae | 97.48 | |
| LUCIA H | Lysinibacillus odysseyi | 98.82 | |
| LUCIA L | Bacillus toyonensis | 98.15 |
| Bacterial Strains from Propolis | Tester Strains | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E. coli | S. marcescens | K. rizophyla | K. pneumoniae | B. cereus | S. aureus | |||||||
| 4 | 10 | 4 | 10 | 4 | 10 | 4 | 10 | 4 | 10 | 4 | 10 | |
| #2C | + | + | +/− | +/− | +/− | + | − | − | − | + | +/− | + |
| #13A | + | + | − | +/− | − | − | − | − | − | − | − | − |
| #13D | + | + | +/− | +/− | + | + | − | − | − | +/− | +/− | + |
| #13F | + | + | +/− | +/− | − | − | − | − | − | − | − | − |
| #13G | + | + | +/− | + | +/− | +/− | − | − | − | + | − | + |
| FLORENCIA B | + | + | +/− | + | +/− | +/− | − | − | − | +/− | − | +/− |
| FLORENCIA C | + | + | + | + | + | + | − | +/− | − | + | − | + |
| FLORENCIA F | + | + | +/− | + | + | + | − | +/− | − | + | − | + |
| LUCIA A | + | + | +/− | + | +/− | + | − | − | − | + | + | + |
| LUCIA C | + | + | +/− | + | +/− | − | − | − | − | +/− | − | + |
| LUCIA H | + | + | +/− | + | +/− | +/− | − | − | − | − | − | − |
| LUCIA L | + | + | +/− | + | + | + | − | − | − | − | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez Matos, A.E.; Bacci, G.; Borruso, L.; Landolfi, M.; Petrocchi, D.; Renzi, S.; Perito, B. Characterization of the Bacterial Communities Inhabiting Tropical Propolis of Puerto Rico. Microorganisms 2023, 11, 1130. https://doi.org/10.3390/microorganisms11051130
Pérez Matos AE, Bacci G, Borruso L, Landolfi M, Petrocchi D, Renzi S, Perito B. Characterization of the Bacterial Communities Inhabiting Tropical Propolis of Puerto Rico. Microorganisms. 2023; 11(5):1130. https://doi.org/10.3390/microorganisms11051130
Chicago/Turabian StylePérez Matos, Ana E., Giovanni Bacci, Luigimaria Borruso, Maria Landolfi, Dominique Petrocchi, Sonia Renzi, and Brunella Perito. 2023. "Characterization of the Bacterial Communities Inhabiting Tropical Propolis of Puerto Rico" Microorganisms 11, no. 5: 1130. https://doi.org/10.3390/microorganisms11051130
APA StylePérez Matos, A. E., Bacci, G., Borruso, L., Landolfi, M., Petrocchi, D., Renzi, S., & Perito, B. (2023). Characterization of the Bacterial Communities Inhabiting Tropical Propolis of Puerto Rico. Microorganisms, 11(5), 1130. https://doi.org/10.3390/microorganisms11051130