
Thanksgiving to Yeast, the HMGB Proteins History from Yeast to Cancer
 , ,
, , 
{kind=link}
{kind=link}
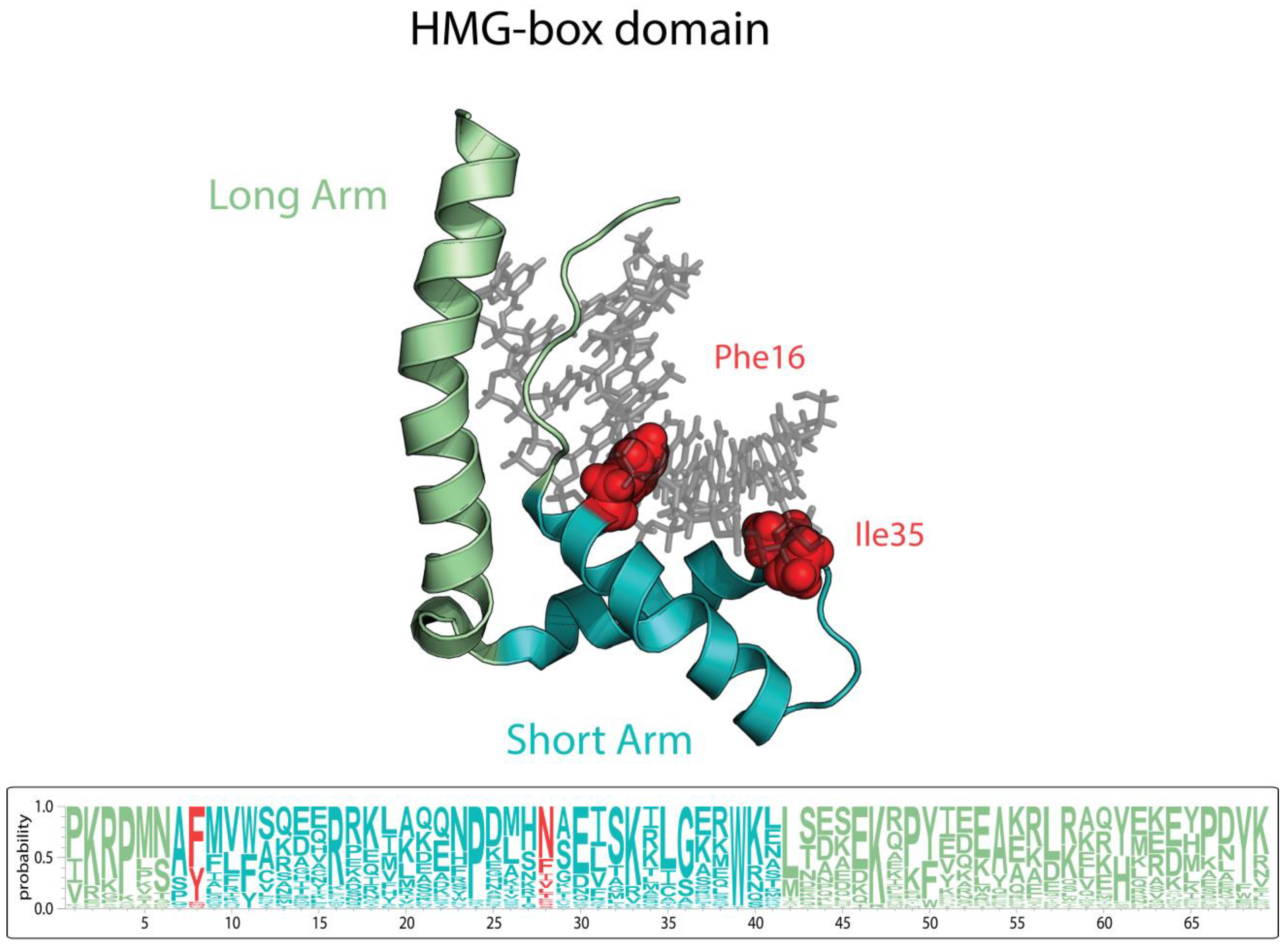{kind=link}
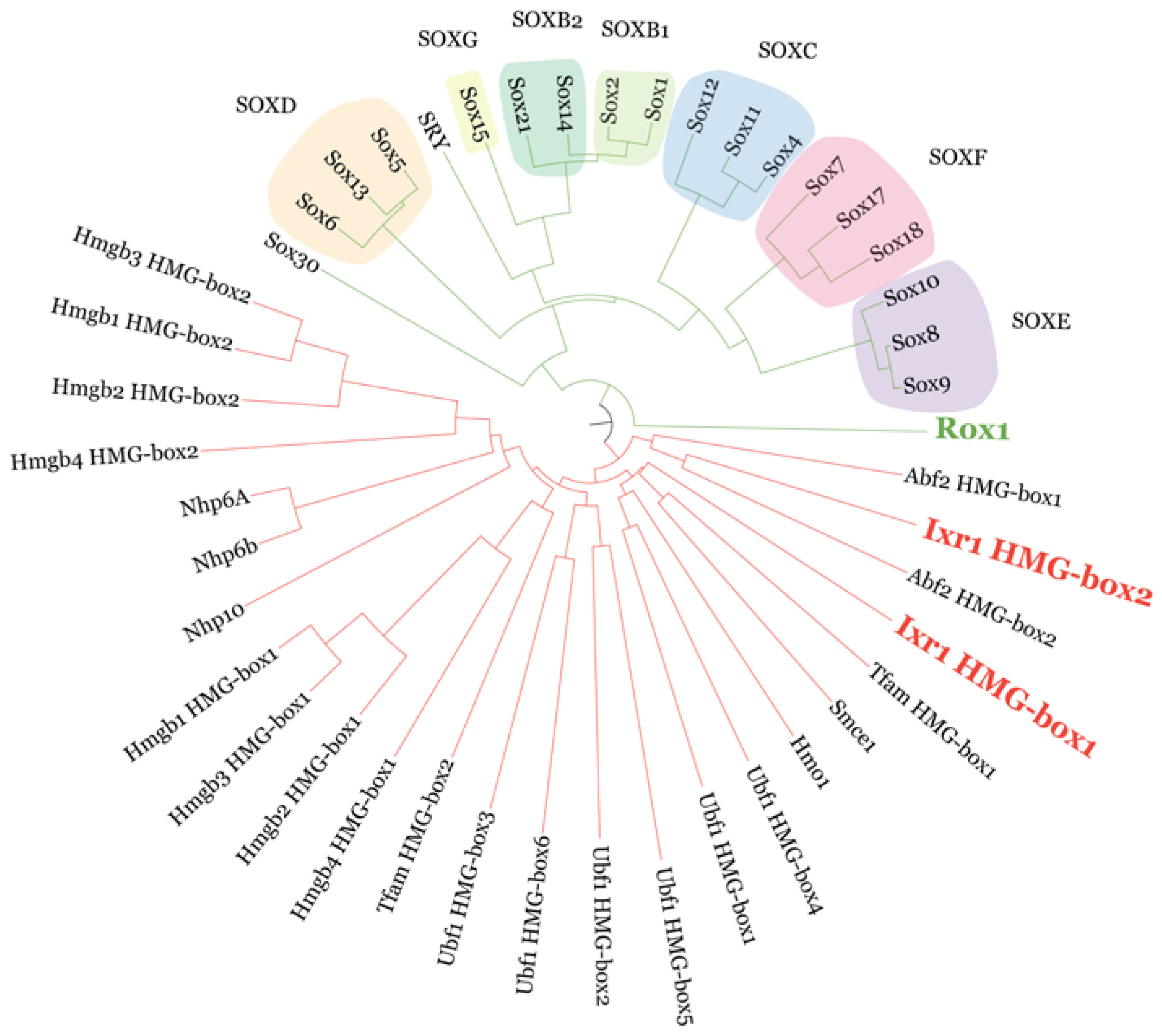{kind=link}
Abstract
1. Introduction
2. Conventional and “Non-Conventional” Yeast Models
3. Yeasts as Biological Tools
4. Outstanding Milestones for Yeast Thanksgiving and Their Relation to Cancer Research
5. The HMGB Proteins in Cancer
6. Studying HMGB Proteins: From Yeasts to Cancer
7. Ongoing Yeast Perspectives in Biomedicine
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matthews, B.J.; Vosshall, L.B. How to Turn an Organism into a Model Organism in 10 ‘Easy’ Steps. J. Exp. Biol. 2020, 223, jeb218198. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Ye, C. Microbial Cell Factories Based on Filamentous Bacteria, Yeasts, and Fungi. Microb. Cell Fact. 2023, 22, 20. [Google Scholar] [CrossRef] [PubMed]
- Tullio, V. Yeast Genomics and its Applications in Biotechnological Processes: What is our Present and Near Future? J. Fungi 2022, 8, 752. [Google Scholar] [CrossRef] [PubMed]
- Bach, S.; Colas, P.; Blondel, M. Budding Yeast, a Model and a Tool... also for Biomedical Research. Med. Sci. 2020, 36, 504–514. [Google Scholar] [CrossRef]
- Goffeau, A. The Yeast Genome. Pathol. Biol. 1998, 46, 96–97. [Google Scholar]
- Goffeau, A.; Barrell, B.G.; Bussey, H.; Davis, R.W.; Dujon, B.; Feldmann, H.; Galibert, F.; Hoheisel, J.D.; Jacq, C.; Johnston, M.; et al. Life with 6000 Genes. Science 1996, 274, 546–567. [Google Scholar] [CrossRef]
- Winzeler, E.A.; Shoemaker, D.D.; Astromoff, A.; Liang, H.; Anderson, K.; Andre, B.; Bangham, R.; Benito, R.; Boeke, J.D.; Bussey, H.; et al. Functional Characterization of the S. cerevisiae Genome by Gene Deletion and Parallel Analysis. Science 1999, 285, 901–906. [Google Scholar] [CrossRef]
- Lampitt, L.H. Nitrogen Metabolism in Saccharomyces cerevisiae. Biochem. J. 1919, 13, 459–486. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, A.; Ferminan, E.; Sanchez, M.; Gonzalez, F.J.; Perez-Campo, F.M.; Garcia, S.; Herrero, A.B.; San Vicente, A.; Cabello, J.; Prado, M.; et al. Non-Conventional Yeasts as Hosts for Heterologous Protein Production. Int. Microbiol. 1998, 1, 131–142. [Google Scholar] [PubMed]
- Yi, X.; Alper, H.S. Considering Strain Variation and Non-Type Strains for Yeast Metabolic Engineering Applications. Life 2022, 12, 510. [Google Scholar] [CrossRef] [PubMed]
- Wiese, E.K.; Hitosugi, T. Tyrosine Kinase Signaling in Cancer Metabolism: PKM2 Paradox in the Warburg Effect. Front. Cell Dev. Biol. 2018, 6, 79. [Google Scholar] [CrossRef] [PubMed]
- Gancedo, J.M. Yeast Carbon Catabolite Repression. Microbiol. Mol. Biol. Rev. 1998, 62, 334–361. [Google Scholar] [CrossRef]
- Viale, A.; Pettazzoni, P.; Lyssiotis, C.A.; Ying, H.; Sanchez, N.; Marchesini, M.; Carugo, A.; Green, T.; Seth, S.; Giuliani, V.; et al. Oncogene Ablation-Resistant Pancreatic Cancer Cells Depend on Mitochondrial Function. Nature 2014, 514, 628–632. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Lim, S.K.; Liang, Q.; Iyer, S.V.; Wang, H.; Wang, Z.; Xie, X.; Sun, D.; Chen, Y.; Tabar, V.; et al. Gboxin is an Oxidative Phosphorylation Inhibitor that Targets Glioblastoma. Nature 2019, 567, 341–346. [Google Scholar] [CrossRef]
- Snyder, V.; Reed-Newman, T.C.; Arnold, L.; Thomas, S.M.; Anant, S. Cancer Stem Cell Metabolism and Potential Therapeutic Targets. Front. Oncol. 2018, 8, 203. [Google Scholar] [CrossRef]
- Fontanesi, F.; Viola, A.M.; Ferrero, I. Heterologous Complementation of the Klaac Null Mutation of Kluyveromyces lactis by the Saccharomyces cerevisiae AAC3 Gene Encoding the ADP/ATP Carrier. FEMS Yeast Res. 2006, 6, 414–420. [Google Scholar] [CrossRef][Green Version]
- González-Siso, M.I.; Freire-Picos, M.A.; Ramil, E.; González-Domínguez, M.; Rodríguez Torres, A.; Cerdán, M.E. Respirofermentative Metabolism in Kluyveromyces lactis: Insights and Perspectives. Enzyme Microb. Technol. 2000, 26, 699–705. [Google Scholar] [CrossRef]
- Sewalt, V.; LaMarta, J.; Shanahan, D.; Gregg, L.; Carrillo, R. Letter to the Editor regarding “GRAS from the Ground Up: Review of the Interim Pilot Program for GRAS Notification” by. Food Chem. Toxicol. 2017, 107, 520–521. [Google Scholar] [CrossRef]
- Fields, S.; Song, O. A Novel Genetic System to Detect Protein-Protein Interactions. Nature 1989, 340, 245–246. [Google Scholar] [CrossRef]
- Paiano, A.; Margiotta, A.; De Luca, M.; Bucci, C. Yeast Two-Hybrid Assay to Identify Interacting Proteins. Curr. Protoc. Protein Sci. 2019, 95, e70. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.H.; Chiou, C.J.; Hayward, G.S. Evaluation and Mapping of the DNA Binding and Oligomerization Domains of the IE2 Regulatory Protein of Human Cytomegalovirus using Yeast One and Two Hybrid Interaction Assays. Gene 1998, 210, 25–36. [Google Scholar] [CrossRef]
- Licitra, E.J.; Liu, J.O. A Three-Hybrid System for Detecting Small Ligand-Protein Receptor Interactions. Proc. Natl. Acad. Sci. USA 1996, 93, 12817–12821. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.; Mukhtar, M.S. Mapping Protein-Protein Interaction using High-Throughput Yeast 2-Hybrid. Methods Mol. Biol. 2017, 1610, 217–230. [Google Scholar] [CrossRef]
- Dai, X.; Yuan, M.; Lu, Y.; Zhu, X.; Liu, C.; Zheng, Y.; Si, S.; Yuan, L.; Zhang, J.; Li, Y. Identification of a Small Molecule that Inhibits the Interaction of LPS Transporters LptA and LptC. Antibiotics 2022, 11, 1385. [Google Scholar] [CrossRef]
- Teymennet-Ramirez, K.V.; Martinez-Morales, F.; Trejo-Hernandez, M.R. Yeast Surface Display System: Strategies for Improvement and Biotechnological Applications. Front. Bioeng. Biotechnol. 2022, 9, 794742. [Google Scholar] [CrossRef]
- Han, L.; Zhao, Y.; Cui, S.; Liang, B. Redesigning of Microbial Cell Surface and its Application to Whole-Cell Biocatalysis and Biosensors. Appl. Biochem. Biotechnol. 2018, 185, 396–418. [Google Scholar] [CrossRef]
- Schreuder, M.P.; Brekelmans, S.; van den Ende, H.; Klis, F.M. Targeting of a Heterologous Protein to the Cell Wall of Saccharomyces Cerevisiae. Yeast 1993, 9, 399–409. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Zhang, L.; Li, L.; Liu, D.; Cheng, H.; Wang, H.; Nawaz, M.Z.; Cheng, H.; Deng, Z. An Alternative Approach to Synthesizing Galactooligosaccharides by Cell-Surface Display of Beta-Galactosidase on Yarrowia lipolytica. J. Agric. Food Chem. 2016, 64, 3819–3827. [Google Scholar] [CrossRef]
- Zhao, N.; Xu, Y.; Wang, K.; Zheng, S. Synthesis of Isomalto-Oligosaccharides by Pichia Pastoris Displaying the Aspergillus Niger Alpha-Glucosidase. J. Agric. Food Chem. 2017, 65, 9468–9474. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Lv, X.; Wang, X.; Wang, J.; Wang, R.; Wang, T. Cell-Surface Displayed Expression of Trehalose Synthase from Pseudomonas putida ATCC 47054 in Pichia pastoris using Pir1p as an Anchor Protein. Front. Microbiol. 2017, 8, 2583. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, H.; Zhu, Y.; Zhang, Y.; Li, X.; Wang, F. Saccharomyces cerevisiae Cell Surface Display Technology: Strategies for Improvement and Applications. Front. Bioeng. Biotechnol. 2022, 10, 1056804. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Liu, M.; Liu, H.; Li, C.; Feng, X. Direct Yeast Surface Codisplay of Sequential Enzymes with Complementary Anchor Motifs: Enabling Enhanced Glycosylation of Natural Products. ACS Synth. Biol. 2023, 12, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, G.S.; Graham, J.D.; Jackson, T.A.; Tung, L.; Powell, R.L.; Horwitz, L.D.; Horwitz, K.B. Tamoxifen Resistant Breast Cancer: Coregulators Determine the Direction of Transcription by Antagonist-Occupied Steroid Receptors. J. Steroid Biochem. Mol. Biol. 1999, 69, 45–50. [Google Scholar] [CrossRef]
- Khazak, V.; Eyrisch, S.; Kato, J.; Tamanoi, F.; Golemis, E.A. A Two-Hybrid Approach to Identify Inhibitors of the RAS-RAF Interaction. Enzymes 2013, 33 Pt A, 213–248. [Google Scholar] [CrossRef] [PubMed]
- Barreiro-Alonso, A.; Camara-Quilez, M.; Salamini-Montemurri, M.; Lamas-Maceiras, M.; Vizoso-Vazquez, A.; Rodriguez-Belmonte, E.; Quindos-Varela, M.; Martinez-Iglesias, O.; Figueroa, A.; Cerdan, M.E. Characterization of HMGB1/2 Interactome in Prostate Cancer by Yeast Two Hybrid Approach: Potential Pathobiological Implications. Cancers 2019, 11, 1729. [Google Scholar] [CrossRef]
- Mahdavi, S.Z.B.; Oroojalian, F.; Eyvazi, S.; Hejazi, M.; Baradaran, B.; Pouladi, N.; Tohidkia, M.R.; Mokhtarzadeh, A.; Muyldermans, S. An Overview on Display Systems (Phage, Bacterial, and Yeast Display) for Production of Anticancer Antibodies; Advantages and Disadvantages. Int. J. Biol. Macromol. 2022, 208, 421–442. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The Hallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer. Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Stock, D.; Leslie, A.G.; Walker, J.E. Molecular Architecture of the Rotary Motor in ATP Synthase. Science 1999, 286, 1700–1705. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, Y.; Guo, Y.; Shi, X.; Chen, X.; Feng, W.; Wu, L.; Zhang, J.; Yu, S.; Wang, Y.; et al. An Overview: The Diversified Role of Mitochondria in Cancer Metabolism. Int. J. Biol. Sci. 2023, 19, 897–915. [Google Scholar] [CrossRef]
- Ben-Shem, A.; Garreau de Loubresse, N.; Melnikov, S.; Jenner, L.; Yusupova, G.; Yusupov, M. The Structure of the Eukaryotic Ribosome at 3.0 A Resolution. Science 2011, 334, 1524–1529. [Google Scholar] [CrossRef]
- Derenzini, M.; Montanaro, L.; Trere, D. Ribosome Biogenesis and Cancer. Acta Histochem. 2017, 119, 190–197. [Google Scholar] [CrossRef]
- Woolford, J.L.J.; Baserga, S.J. Ribosome Biogenesis in the Yeast Saccharomyces cerevisiae. Genetics 2013, 195, 643–681. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, R.D. The Molecular Basis of Eukaryotic Transcription. Proc. Natl. Acad. Sci. USA 2007, 104, 12955–12961. [Google Scholar] [CrossRef]
- Dotson, M.R.; Yuan, C.X.; Roeder, R.G.; Myers, L.C.; Gustafsson, C.M.; Jiang, Y.W.; Li, Y.; Kornberg, R.D.; Asturias, F.J. Structural Organization of Yeast and Mammalian Mediator Complexes. Proc. Natl. Acad. Sci. USA 2000, 97, 14307–14310. [Google Scholar] [CrossRef]
- Myers, L.C.; Gustafsson, C.M.; Bushnell, D.A.; Lui, M.; Erdjument-Bromage, H.; Tempst, P.; Kornberg, R.D. The Med Proteins of Yeast and their Function through the RNA Polymerase II Carboxy-Terminal Domain. Genes Dev. 1998, 12, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, R. The Molecular Basis of Eukaryotic Transcription (Nobel Lecture). Angew. Chem. Int. Ed. Engl. 2007, 46, 6956–6965. [Google Scholar] [CrossRef] [PubMed]
- Aguilera, A. The Connection between Transcription and Genomic Instability. EMBO J. 2002, 21, 195–201. [Google Scholar] [CrossRef]
- Datta, A.; Jinks-Robertson, S. Association of Increased Spontaneous Mutation Rates with High Levels of Transcription in Yeast. Science 1995, 268, 1616–1619. [Google Scholar] [CrossRef]
- Voelkel-Meiman, K.; Keil, R.L.; Roeder, G.S. Recombination-Stimulating Sequences in Yeast Ribosomal DNA Correspond to Sequences Regulating Transcription by RNA Polymerase I. Cell 1987, 48, 1071–1079. [Google Scholar] [CrossRef]
- Bowry, A.; Kelly, R.D.W.; Petermann, E. Hypertranscription and Replication Stress in Cancer. Trends Cancer. 2021, 7, 863–877. [Google Scholar] [CrossRef] [PubMed]
- Kadyrov, F.A.; Holmes, S.F.; Arana, M.E.; Lukianova, O.A.; O’Donnell, M.; Kunkel, T.A.; Modrich, P. Saccharomyces cerevisiae MutLalpha is a Mismatch Repair Endonuclease. J. Biol. Chem. 2007, 282, 37181–37190. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Adebali, O.; Yang, Y.; Selby, C.P.; Sancar, A. Single-Nucleotide Resolution Dynamic Repair Maps of UV Damage in Saccharomyces cerevisiae Genome. Proc. Natl. Acad. Sci. USA 2018, 115, E3408–E3415. [Google Scholar] [CrossRef]
- Prado, F.; Cortes-Ledesma, F.; Huertas, P.; Aguilera, A. Mitotic Recombination in Saccharomyces cerevisiae. Curr. Genet. 2003, 42, 185–198. [Google Scholar] [CrossRef]
- Rondon, A.G.; Aguilera, A. What Causes an RNA-DNA Hybrid to Compromise Genome Integrity? DNA Repair 2019, 81, 102660. [Google Scholar] [CrossRef]
- Cleaver, J.E. Profile of Tomas Lindahl, Paul Modrich, and Aziz Sancar, 2015 Nobel Laureates in Chemistry. Proc. Natl. Acad. Sci. USA 2016, 113, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, S.G.; Shah, P.; Khattar, E. Recent Advances in Therapeutic Application of DNA Damage Response Inhibitors Against Cancer. Anti-Cancer Agents Med. Chem. 2022, 22, 469–484. [Google Scholar] [CrossRef]
- Forsburg, S.L.; Nurse, P. Cell Cycle Regulation in the Yeasts Saccharomyces cerevisiae and Schizosaccharomyces pombe. Annu. Rev. Cell Biol. 1991, 7, 227–256. [Google Scholar] [CrossRef]
- Nurse, P.; Masui, Y.; Hartwell, L. Understanding the Cell Cycle. Nat. Med. 1998, 4, 1103–1106. [Google Scholar] [CrossRef]
- Lee, M.G.; Nurse, P. Complementation used to Clone a Human Homologue of the Fission Yeast Cell Cycle Control Gene cdc2. Nature 1987, 327, 31–35. [Google Scholar] [CrossRef]
- Paulovich, A.G.; Hartwell, L.H. A Checkpoint Regulates the Rate of Progression through S Phase in S. Cerevisiae in Response to DNA Damage. Cell 1995, 82, 841–847. [Google Scholar] [CrossRef]
- Nurse, P.M. Nobel Lecture. Cyclin Dependent Kinases and Cell Cycle Control. Biosci. Rep. 2002, 22, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Hartwell, L.H. Yeast and Cancer. Biosci. Rep. 2004, 24, 523–544. [Google Scholar] [CrossRef]
- Nadav, E.; Shmueli, A.; Barr, H.; Gonen, H.; Ciechanover, A.; Reiss, Y. A Novel Mammalian Endoplasmic Reticulum Ubiquitin Ligase Homologous to the Yeast Hrd1. Biochem. Biophys. Res. Commun. 2003, 303, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Hoeller, D.; Hecker, C.; Dikic, I. Ubiquitin and Ubiquitin-Like Proteins in Cancer Pathogenesis. Nat. Rev. Cancer. 2006, 6, 776–788. [Google Scholar] [CrossRef] [PubMed]
- Kimata, Y. APC/C Ubiquitin Ligase: Coupling Cellular Differentiation to G1/G0 Phase in Multicellular Systems. Trends Cell Biol. 2019, 29, 591–603. [Google Scholar] [CrossRef]
- He, W.; Meng, J. CDC20: A Novel Therapeutic Target in Cancer. Am. J. Transl. Res. 2023, 15, 678–693. [Google Scholar]
- Varela, E.; Blasco, M.A. 2009 Nobel Prize in Physiology Or Medicine: Telomeres and Telomerase. Oncogene 2010, 29, 1561–1565. [Google Scholar] [CrossRef]
- Sandell, L.L.; Zakian, V.A. Loss of a Yeast Telomere: Arrest, Recovery, and Chromosome Loss. Cell 1993, 75, 729–739. [Google Scholar] [CrossRef]
- Lundblad, V.; Szostak, J.W. A Mutant with a Defect in Telomere Elongation Leads to Senescence in Yeast. Cell 1989, 57, 633–643. [Google Scholar] [CrossRef]
- Lendvay, T.S.; Morris, D.K.; Sah, J.; Balasubramanian, B.; Lundblad, V. Senescence Mutants of Saccharomyces cerevisiae with a Defect in Telomere Replication Identify Three Additional EST Genes. Genetics 1996, 144, 1399–1412. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.S.; Gottschling, D.E. TLC1: Template RNA Component of Saccharomyces Cerevisiae Telomerase. Science 1994, 266, 404–409. [Google Scholar] [CrossRef]
- Wanat, J.J.; Johnson, F.B. Telomere Stability and Carcinogenesis: An Off-again, on-again Relationship. J. Clin. Investig. 2012, 122, 1962–1965. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lundblad, V.; Blackburn, E.H. An Alternative Pathway for Yeast Telomere Maintenance Rescues est1-Senescence. Cell 1993, 73, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Schekman, R. Charting the Secretory Pathway in a Simple Eukaryote. Mol. Biol. Cell 2010, 21, 3781–3784. [Google Scholar] [CrossRef]
- Brose, N. All Roads Lead to Neuroscience: The 2013 Nobel Prize in Physiology Or Medicine. Neuron 2014, 81, 723–727. [Google Scholar] [CrossRef]
- Goldenring, J.R. A Central Role for Vesicle Trafficking in Epithelial Neoplasia: Intracellular Highways to Carcinogenesis. Nat. Rev. Cancer. 2013, 13, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Ohsumi, Y. Autophagy from Beginning to End. Interview by Caitlin Sedwick. J. Cell Biol. 2012, 197, 164–165. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N. Autophagy: Process and Function. Genes Dev. 2007, 21, 2861–2873. [Google Scholar] [CrossRef] [PubMed]
- Yun, C.W.; Lee, S.H. The Roles of Autophagy in Cancer. Int. J. Mol. Sci. 2018, 19, 3466. [Google Scholar] [CrossRef]
- Comel, A.; Sorrentino, G.; Capaci, V.; Del Sal, G. The Cytoplasmic Side of p53′s Oncosuppressive Activities. FEBS Lett. 2014, 588, 2600–2609. [Google Scholar] [CrossRef] [PubMed]
- Choi, A.M.K.; Ryter, S.W.; Levine, B. Autophagy in Human Health and Disease. N. Engl. J. Med. 2013, 368, 651–662. [Google Scholar] [CrossRef]
- Wang, J.; Sheng, Z.; Cai, Y. Effects of microRNA-513b on Cell Proliferation, Apoptosis, Invasion, and Migration by Targeting HMGB3 through Regulation of mTOR Signaling Pathway in Non-Small-Cell Lung Cancer. J. Cell. Physiol. 2019, 234, 10934–10941. [Google Scholar] [CrossRef]
- Deng, X.; Niu, Z.; Hao, C.; Lin, J.; Yao, W. HMGB1 Coordinates with Brahma-Related Gene 1 to Promote Epithelial-Mesenchymal Transition Via the PI3K/Akt/mTOR Pathway in BEAS-2B Cells. Exp. Cell Res. 2023, 424, 113522. [Google Scholar] [CrossRef]
- Alao, J.; Legon, L.; Dabrowska, A.; Tricolici, A.; Kumar, J.; Rallis, C. Interplays of AMPK and TOR in Autophagy Regulation in Yeast. Cells 2023, 12, 519. [Google Scholar] [CrossRef] [PubMed]
- Bjedov, I.; Rallis, C. The Target of Rapamycin Signalling Pathway in Ageing and Lifespan Regulation. Genes 2020, 11, 1043. [Google Scholar] [CrossRef] [PubMed]
- Chiu, M.I.; Katz, H.; Berlin, V. RAPT1, a Mammalian Homolog of Yeast Tor, Interacts with the FKBP12/rapamycin Complex. Proc. Natl. Acad. Sci. USA 1994, 91, 12574–12578. [Google Scholar] [CrossRef] [PubMed]
- Rangel, M.; Kong, J.; Bhatt, V.; Khayati, K.; Guo, J.Y. Autophagy and Tumorigenesis. FEBS J. 2022, 289, 7177–7198. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Liao, M.; Qin, R.; Zhu, S.; Peng, C.; Fu, L.; Chen, Y.; Han, B. Regulated Cell Death (RCD) in Cancer: Key Pathways and Targeted Therapies. Signal. Transduct. Target Ther. 2022, 7, 286. [Google Scholar] [CrossRef]
- Ángel, V.-V.; Aida, B.-A.; Agustín, R.-D.; Mónica, L.-M.; Esther, R.-B.; Manuel, B.; Isabel, G.-S.M.; Esperanza, C.M. HMGB Proteins from Yeast to Human. Gene Regulation, DNA Repair and Beyond. In Old Yeasts—New Questions; InTech: London, UK, 2017; pp. 139–165. ISBN 978-953-51-3677-4. [Google Scholar]
- Cámara-Quílez, M.; Barreiro-Alonso, A.; Vizoso-Vazquez, A.; Rodriguez-Belmonte, E.; Quindós-Varela, M.; Lamas-Maceiras, M.; Cerdán, M.E. The HMGB1-2 Ovarian Cancer Interactome. the Role of HMGB Proteins and their Interacting Partners MIEN1 and NOP53 in Ovary Cancer and Drug-Response. Cancers 2020, 12, 2435. [Google Scholar] [CrossRef]
- Andersson, U.; Antoine, D.J. The Functions of HMGB1 Depend on Molecular Localization and Post-Translational Modifications. J. Intern. Med. 2014, 276, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Kang, R.; Tang, D. The Mechanism of HMGB1 Secretion and Release. Exp. Mol. Med. 2022, 54, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Barreiro-Alonso, A.; Lamas-Maceiras, M.; Rodríguez-Belmonte, E.; Vizoso-Vazquez, A.; Quindos, M.; Cerdan, M.E. High Mobility Group B Proteins, their Partners, and Other Redox Sensors in Ovarian and Prostate Cancer. Oxid Med. Cell. Longev. 2016, 2016, 5845061. [Google Scholar] [CrossRef]
- Rapoport, B.L.; Steel, H.C.; Theron, A.J.; Heyman, L.; Smit, T.; Ramdas, Y.; Anderson, R. High Mobility Group Box 1 in Human Cancer. Cells 2020, 9, 1664. [Google Scholar] [CrossRef]
- Taverna, S.; Tonacci, A.; Ferraro, M.; Cammarata, G.; Cuttitta, G.; Bucchieri, S.; Pace, E.; Gangemi, S. High Mobility Group Box 1: Biological Functions and Relevance in Oxidative Stress Related Chronic Diseases. Cells 2022, 11, 849. [Google Scholar] [CrossRef] [PubMed]
- Voong, C.K.; Goodrich, J.A.; Kugel, J.F. Interactions of HMGB Proteins with the Genome and the Impact on Disease. Biomolecules 2021, 11, 1451. [Google Scholar] [CrossRef] [PubMed]
- Kang, R.; Livesey, K.M.; Zeh, H.J.; Loze, M.T.; Tang, D. HMGB1: A Novel Beclin 1-Binding Protein Active in Autophagy. Autophagy 2010, 6, 1209–1211. [Google Scholar] [CrossRef]
- Tang, D.; Kang, R.; Livesey, K.M.; Kroemer, G.; Billiar, T.R.; Van Houten, B.; Zeh, H.J., III; Lotze, M.T. High-Mobility Group Box 1 is Essential for Mitochondrial Quality Control. Cell Metab. 2011, 13, 701–711. [Google Scholar] [CrossRef]
- Ito, H.; Fujita, K.; Tagawa, K.; Chen, X.; Homma, H.; Sasabe, T.; Shimizu, J.; Shimizu, S.; Tamura, T.; Muramatsu, S.; et al. HMGB1 Facilitates Repair of Mitochondrial DNA Damage and Extends the Lifespan of Mutant Ataxin-1 Knock-in Mice. EMBO Mol. Med. 2015, 7, 78–101. [Google Scholar] [CrossRef]
- Andersson, U.; Yang, H.; Harris, H. High-Mobility Group Box 1 Protein (HMGB1) Operates as an Alarmin Outside as Well as Inside Cells. Semin. Immunol. 2018, 38, 40–48. [Google Scholar] [CrossRef]
- Bianchi, M.E.; Crippa, M.P.; Manfredi, A.A.; Mezzapelle, R.; Rovere Querini, P.; Venereau, E. High-Mobility Group Box 1 Protein Orchestrates Responses to Tissue Damage Via Inflammation, Innate and Adaptive Immunity, and Tissue Repair. Immunol. Rev. 2017, 280, 74–82. [Google Scholar] [CrossRef]
- Wang, J.; Li, R.; Peng, Z.; Hu, B.; Rao, X.; Li, J. HMGB1 Participates in LPS-induced Acute Lung Injury by Activating the AIM2 Inflammasome in Macrophages and Inducing Polarization of M1 Macrophages Via TLR2, TLR4, and RAGE/NF-kappaB Signaling Pathways. Int. J. Mol. Med. 2020, 45, 61–80. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, Y. HMGB1 in Inflammation and Cancer. J. Hematol. Oncol. 2020, 13, 116. [Google Scholar] [CrossRef] [PubMed]
- Nagatani, G.; Nomoto, M.; Takano, H.; Ise, T.; Kato, K.; Imamura, T.; Izumi, H.; Makishima, K.; Kohno, K. Transcriptional Activation of the Human HMG1 Gene in Cisplatin-Resistant Human Cancer Cells. Cancer Res. 2001, 61, 1592–1597. [Google Scholar] [PubMed]
- Kang, R.; Zhang, Q.; Zeh, H.J., III; Lotze, M.T.; Tang, D. HMGB1 in Cancer: Good, Bad, Or both? Clin. Cancer Res. 2013, 19, 4046–4057. [Google Scholar] [CrossRef] [PubMed]
- Tohme, S.; Yazdani, H.O.; Liu, Y.; Loughran, P.; van der Windt, D.J.; Huang, H.; Simmons, R.L.; Shiva, S.; Tai, S.; Tsung, A. Hypoxia Mediates Mitochondrial Biogenesis in Hepatocellular Carcinoma to Promote Tumor Growth through HMGB1 and TLR9 Interaction. Hepatology 2017, 66, 182–197. [Google Scholar] [CrossRef] [PubMed]
- Kang, R.; Xie, Y.; Zhang, Q.; Hou, W.; Jiang, Q.; Zhu, S.; Liu, J.; Zeng, D.; Wang, H.; Bartlett, D.L.; et al. Intracellular HMGB1 as a Novel Tumor Suppressor of Pancreatic Cancer. Cell Res. 2017, 27, 916–932. [Google Scholar] [CrossRef]
- Kang, R.; Tang, D. The Dual Role of HMGB1 in Pancreatic Cancer. J. Pancreatol. 2018, 1, 19–24. [Google Scholar] [CrossRef]
- Dormoy-Raclet, V.; Cammas, A.; Celona, B.; Lian, X.J.; van der Giessen, K.; Zivojnovic, M.; Brunelli, S.; Riuzzi, F.; Sorci, G.; Wilhelm, B.T.; et al. HuR and miR-1192 Regulate Myogenesis by Modulating the Translation of HMGB1 mRNA. Nat. Commun. 2013, 4, 2388. [Google Scholar] [CrossRef] [PubMed]
- Kwak, M.S.; Kim, H.S.; Lee, B.; Kim, Y.H.; Son, M.; Shin, J. Immunological Significance of HMGB1 Post-Translational Modification and Redox Biology. Front. Immunol. 2020, 11, 1189. [Google Scholar] [CrossRef]
- Li, G.; Liang, X.; Lotze, M.T. HMGB1: The Central Cytokine for all Lymphoid Cells. Front. Immunol. 2013, 4, 68. [Google Scholar] [CrossRef] [PubMed]
- Venereau, E.; Casalgrandi, M.; Schiraldi, M.; Antoine, D.J.; Cattaneo, A.; De Marchis, F.; Liu, J.; Antonelli, A.; Preti, A.; Raeli, L.; et al. Mutually Exclusive Redox Forms of HMGB1 Promote Cell Recruitment Or Proinflammatory Cytokine Release. J. Exp. Med. 2012, 209, 1519–1528. [Google Scholar] [CrossRef]
- Galluzzi, L.; Buque, A.; Kepp, O.; Zitvogel, L.; Kroemer, G. Immunogenic Cell Death in Cancer and Infectious Disease. Nat. Rev. Immunol. 2017, 17, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, H.; Luo, Y.; Kuniyasu, H. Non-Histone Nuclear Factor HMGB1 as a Therapeutic Target in Colorectal Cancer. Expert Opin. Ther. Targets 2011, 15, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhang, Q.; Cheng, Y.; Chen, X.; Wang, G.; Shi, M.; Zhang, T.; Cao, Y.; Pan, H.; Zhang, L.; et al. Tumor-Derived Exosomal HMGB1 Fosters Hepatocellular Carcinoma Immune Evasion by Promoting TIM-1(+) Regulatory B Cell Expansion. J. Immunother. Cancer 2018, 6, 145. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Li, T.; Wang, C.; Ma, Y.; Liu, Y.; Zheng, M.; Liu, Z.; Chen, J.; Li, K.; Sun, S.; et al. Synergy of Nanodiamond-Doxorubicin Conjugates and PD-L1 Blockade Effectively Turns Tumor-Associated Macrophages Against Tumor Cells. J. Nanobiotechnol. 2021, 19, 268. [Google Scholar] [CrossRef]
- Alsaab, H.O.; Sau, S.; Alzhrani, R.; Tatiparti, K.; Bhise, K.; Kashaw, S.K.; Iyer, A.K. PD-1 and PD-L1 Checkpoint Signaling Inhibition for Cancer Immunotherapy: Mechanism, Combinations, and Clinical Outcome. Front. Pharmacol. 2017, 8, 561. [Google Scholar] [CrossRef]
- Zitomer, R.S.; Carrico, P.; Deckert, J. Regulation of Hypoxic Gene Expression in Yeast. Kidney Int. 1997, 51, 507–513. [Google Scholar] [CrossRef][Green Version]
- Cerdán, M.E.; Zitomer, R.S. Oxygen-Dependent Upstream Activation Sites of Saccharomyces cerevisiae Cytochrome c Genes are Related Forms of the Same Sequence. Mol. Cell. Biol. 1988, 8, 2275–2279. [Google Scholar] [CrossRef]
- Lowry, C.V.; Cerdán, M.E.; Zitomer, R.S. A Hypoxic Consensus Operator and a Constitutive Activation Region Regulate the ANB1 Gene of Saccharomyces cerevisiae. Mol. Cell. Biol. 1990, 10, 5921–5926. [Google Scholar] [CrossRef]
- Deckert, J.; Rodriguez Torres, A.M.; Simon, J.T.; Zitomer, R.S. Mutational Analysis of Rox1, a DNA-Bending Repressor of Hypoxic Genes in Saccharomyces cerevisiae. Mol. Cell. Biol. 1995, 15, 6109–6117. [Google Scholar] [CrossRef]
- She, Z.Y.; Yang, W.X. SOX Family Transcription Factors Involved in Diverse Cellular Events during Development. Eur. J. Cell Biol. 2015, 94, 547–563. [Google Scholar] [CrossRef]
- Lambert, J.R.; Bilanchone, V.W.; Cumsky, M.G. The ORD1 Gene Encodes a Transcription Factor Involved in Oxygen Regulation and is Identical to IXR1, a Gene that Confers Cisplatin Sensitivity to Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1994, 91, 7345–7349. [Google Scholar] [CrossRef] [PubMed]
- Bourdineaud, J.P.; De Sampaio, G.; Lauquin, G.J. A Rox1-Independent Hypoxic Pathway in Yeast. Antagonistic Action of the Repressor Ord1 and Activator Yap1 for Hypoxic Expression of the SRP1/TIR1 Gene. Mol. Microbiol. 2000, 38, 879–890. [Google Scholar] [CrossRef] [PubMed]
- Castro-Prego, R.; Lamas Maceiras, M.; Soengas, P.; Carneiro, I.; González-Siso, I.; Cerdán, M.E. Regulatory Factors Controlling Transcription of Saccharomyces cerevisiae IXR1 by Oxygen Levels: A Model of Transcriptional Adaptation from Aerobiosis to Hypoxia Implicating ROX1and IXR1cross-Regulation. Biochem. J. 2010, 425, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Vizoso-Vázquez, A.; Lamas-Maceiras, M.; Becerra, M.; González-Siso, M.I.; Rodráguez-Belmonte, E.; Cerdán, M.E. Ixr1p and the Control of the Saccharomyces cerevisiae Hypoxic Response. Appl. Microbiol. Biotechnol. 2012, 94, 173–184. [Google Scholar] [CrossRef]
- Barreiro-Alonso, A.; Lamas-Maceiras, M.; Cerdán, E.M.; Vizoso-Vázquez, A. The HMGB Protein Ixr1 Interacts with Ssn8 and Tdh3 Involved in Transcriptional Regulation. FEMS Yeast Res. 2018, 18, foy013. [Google Scholar] [CrossRef]
- Kuchin, S.; Yeghiayan, P.; Carlson, M. Cyclin-Dependent Protein Kinase and Cyclin Homologs SSN3 and SSN8 Contribute to Transcriptional Control in Yeast. Proc. Natl. Acad. Sci. USA 1995, 92, 4006–4010. [Google Scholar] [CrossRef]
- Brown, S.J.; Kellett, P.J.; Lippard, S.J. Ixr1, a Yeast Protein that Binds to Platinated DNA and Confers Sensitivity to Cisplatin. Science 1993, 261, 603–605. [Google Scholar] [CrossRef]
- McA’Nulty, M.M.; Lippard, S.J. The HMG-Domain Protein Ixr1 Blocks Excision Repair of Cisplatin-DNA Adducts in Yeast. Mutat. Res. DNA Repair 1996, 362, 75–86. [Google Scholar] [CrossRef]
- McA’Nulty, M.M.; Whitehead, J.P.; Lippard, S.J. Binding of Ixr1, a Yeast HMG-Domain Protein, to Cisplatin-DNA Adducts in Vitro and in Vivo. Biochemistry 1996, 35, 6089–6099. [Google Scholar] [CrossRef] [PubMed]
- Vizoso-Vázquez, A.; Lamas-Maceiras, M.; Fernandez-Leiro, R.; Rico-Diaz, A.; Becerra, M.; Cerdán, M.E. Dual Function of Ixr1 in Transcriptional Regulation and Recognition of Cisplatin-DNA Adducts is Caused by Differential Binding through its Two HMG-Boxes. Biochim. Biophys. Acta Gene Regul. Mech. 2017, 1860, 256–269. [Google Scholar] [CrossRef] [PubMed]
- Kiers, J.; Zeeman, A.M.; Luttik, M.; Thiele, C.; Castrillo, J.I.; Steensma, H.Y.; van Dijken, J.P.; Pronk, J.T. Regulation of Alcoholic Fermentation in Batch and Chemostat Cultures of Kluyveromyces lactis CBS 2359. Yeast 1998, 14, 459–469. [Google Scholar] [CrossRef]
- Snoek, I.S.I.; Steensma, H.Y. Why does Kluyveromyces lactis Not Grow Under Anaerobic Conditions? Comparison of Essential Anaerobic Genes of Saccharomyces cerevisiae with the Kluyveromyces lactis Genome. FEMS Yeast Res. 2006, 6, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.M.R.; Maceiras, M.L.; Belmonte, E.R.; Naveira, L.N.; Calvo, M.B.; Cerdan, M.E. KlRox1p Contributes to Yeast Resistance to Metals and is Necessary for KlYCF1 Expression in the Presence of Cadmium. Gene 2012, 497, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Rico-Diaz, A.; Barreiro-Alonso, A.; Rey-Souto, C.; Becerra, M.; Lamas-Maceiras, M.; Cerdan, M.E.; Vizoso-Vazquez, A. The HMGB Protein KlIxr1, a DNA Binding Regulator of Kluyveromyces lactis Gene Expression Involved in Oxidative Metabolism, Growth, and dNTP Synthesis. Biomolecules 2021, 11, 1392. [Google Scholar] [CrossRef]
- Vizoso-Vázquez, A.; Lamas-Maceiras, M.; González-Siso, M.I.; Cerdán, M.E. Ixr1 Regulates Ribosomal Gene Transcription and Yeast Response to Cisplatin. Sci. Rep. 2018, 8, 3090. [Google Scholar] [CrossRef]
- Delgado-Roman, I.; Munoz-Centeno, M.C. Coupling between Cell Cycle Progression and the Nuclear RNA Polymerases System. Front. Mol. Biosci. 2021, 8, 691636. [Google Scholar] [CrossRef]
- Sato, M.; Irie, K.; Suda, Y.; Mizuno, T.; Irie, K. The RNA-Binding Protein Puf5 and the HMGB Protein Ixr1 Contribute to Cell Cycle Progression through the Regulation of Cell Cycle-Specific Expression of CLB1 in Saccharomyces cerevisiae. PLoS Genet. 2022, 18, e1010340. [Google Scholar] [CrossRef]
- Soussi, T.; Beroud, C. Assessing TP53 Status in Human Tumours to Evaluate Clinical Outcome. Nat. Rev. Cancer. 2001, 1, 233–240. [Google Scholar] [CrossRef]
- Sermeus, A.; Michiels, C. Reciprocal Influence of the p53 and the Hypoxic Pathways. Cell Death Dis. 2011, 2, e164. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.B.; Schumacher, B. P53 in the DNA-Damage-Repair Process. Cold Spring Harb. Perspect. Med. 2016, 6, a026070. [Google Scholar] [CrossRef]
- Lessard, F.; Brakier-Gingras, L.; Ferbeyre, G. Ribosomal Proteins Control Tumor Suppressor Pathways in Response to Nucleolar Stress. Bioessays 2019, 41, e1800183. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Quaas, M.; Steiner, L.; Engeland, K. The p53-p21-DREAM-CDE/CHR Pathway Regulates G2/M Cell Cycle Genes. Nucleic Acids Res. 2016, 44, 164–174. [Google Scholar] [CrossRef]
- Holzel, M.; Burger, K.; Muhl, B.; Orban, M.; Kellner, M.; Eick, D. The Tumor Suppressor p53 Connects Ribosome Biogenesis to Cell Cycle Control: A Double-Edged Sword. Oncotarget 2010, 1, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Rowell, J.P.; Simpson, K.L.; Stott, K.; Watson, M.; Thomas, J.O. HMGB1-Facilitated p53 DNA Binding Occurs Via HMG-Box/p53 Transactivation Domain Interaction, Regulated by the Acidic Tail. Structure 2012, 20, 2014–2024. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shao, S.; Han, D.; Xu, Y.; Jiao, D.; Wu, J.; Yang, F.; Ge, Y.; Shi, S.; Li, Y.; et al. High Mobility Group Box 1 Promotes the Epithelial-to-Mesenchymal Transition in Prostate Cancer PC3 Cells Via the RAGE/NF-kappaB Signaling Pathway. Int. J. Oncol. 2018, 53, 659–671. [Google Scholar] [CrossRef]
- Li, Z.; Wang, H.; Song, B.; Sun, Y.; Xu, Z.; Han, J. Silencing HMGB1 Expression by Lentivirus-Mediated Small Interfering RNA (siRNA) Inhibits the Proliferation and Invasion of Colorectal Cancer LoVo Cells in vitro and in vivo. Zhonghua Zhong Liu Za Zhi 2015, 37, 664–670. [Google Scholar]
- Bagatelli, F.F.M.; de Luna Vitorino, F.N.; da Cunha, J.P.C.; Oliveira, C.C. The Ribosome Assembly Factor Nop53 has a Structural Role in the Formation of Nuclear Pre-60S Intermediates, Affecting Late Maturation Events. Nucleic Acids Res. 2021, 49, 7053–7074. [Google Scholar] [CrossRef]
- Kim, H.K.; Fuchs, G.; Wang, S.; Wei, W.; Zhang, Y.; Park, H.; Roy-Chaudhuri, B.; Li, P.; Xu, J.; Chu, K.; et al. A Transfer-RNA-Derived Small RNA Regulates Ribosome Biogenesis. Nature 2017, 552, 57–62. [Google Scholar] [CrossRef]
- Barreiro-Alonso, A.; Lamas-Maceiras, M.; Lorenzo-Catoira, L.; Pardo, M.; Yu, L.; Choudhary, J.S.; Cerdan, M.E. HMGB1 Protein Interactions in Prostate and Ovary Cancer Models Reveal Links to RNA Processing and Ribosome Biogenesis through NuRD, THOC and Septin Complexes. Cancers 2021, 13, 4686. [Google Scholar] [CrossRef] [PubMed]
- Meraner, J.; Lechner, M.; Schwarze, F.; Gander, R.; Jesacher, F.; Loidl, P. Cell Cycle Dependent Role of HDAC1 for Proliferation Control through Modulating Ribosomal DNA Transcription. Cell Biol. Int. 2008, 32, 1073–1080. [Google Scholar] [CrossRef]
- Salamini-Montemurri, M.; Lamas-Maceiras, M.; Barreiro-Alonso, A.; Vizoso-Vazquez, A.; Rodriguez-Belmonte, E.; Quindos-Varela, M.; Cerdan, M.E. The Challenges and Opportunities of LncRNAs in Ovarian Cancer Research and Clinical use. Cancers 2020, 12, 1020. [Google Scholar] [CrossRef] [PubMed]
- Yong Joon Kim, J.; Sang, Z.; Xiang, Y.; Shen, Z.; Shi, Y. Nanobodies: Robust Miniprotein Binders in Biomedicine. Adv. Drug Deliv. Rev. 2023, 195, 114726. [Google Scholar] [CrossRef] [PubMed]
- Gast, V.; Sandegren, A.; Dunas, F.; Ekblad, S.; Guler, R.; Thoren, S.; Tous Mohedano, M.; Molin, M.; Engqvist, M.K.M.; Siewers, V. Engineering Saccharomyces Cerevisiae for the Production and Secretion of Affibody Molecules. Microb. Cell Fact. 2022, 21, 36. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, A.G.; Cenci, S. Autophagy and Longevity: Evolutionary Hints from Hyper-Longevous Mammals. Front. Endocrinol. 2022, 13, 1085522. [Google Scholar] [CrossRef]
- Kachroo, A.H.; Vandeloo, M.; Greco, B.M.; Abdullah, M. Humanized Yeast to Model Human Biology, Disease and Evolution. Dis. Model. Mech. 2022, 15, dmm049309. [Google Scholar] [CrossRef]
- Ball, D.A.; Jalloh, B.; Karpova, T.S. Impact of Saccharomyces Cerevisiae on the Field of Single-Molecule Biophysics. Int. J. Mol. Sci. 2022, 23, 15895. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lamas-Maceiras, M.; Vizoso-Vázquez, Á.; Barreiro-Alonso, A.; Cámara-Quílez, M.; Cerdán, M.E. Thanksgiving to Yeast, the HMGB Proteins History from Yeast to Cancer. Microorganisms 2023, 11, 993. https://doi.org/10.3390/microorganisms11040993
Lamas-Maceiras M, Vizoso-Vázquez Á, Barreiro-Alonso A, Cámara-Quílez M, Cerdán ME. Thanksgiving to Yeast, the HMGB Proteins History from Yeast to Cancer. Microorganisms. 2023; 11(4):993. https://doi.org/10.3390/microorganisms11040993
Chicago/Turabian StyleLamas-Maceiras, Mónica, Ángel Vizoso-Vázquez, Aida Barreiro-Alonso, María Cámara-Quílez, and María Esperanza Cerdán. 2023. "Thanksgiving to Yeast, the HMGB Proteins History from Yeast to Cancer" Microorganisms 11, no. 4: 993. https://doi.org/10.3390/microorganisms11040993
APA StyleLamas-Maceiras, M., Vizoso-Vázquez, Á., Barreiro-Alonso, A., Cámara-Quílez, M., & Cerdán, M. E. (2023). Thanksgiving to Yeast, the HMGB Proteins History from Yeast to Cancer. Microorganisms, 11(4), 993. https://doi.org/10.3390/microorganisms11040993