
A Qualitative PCR Assay for the Discrimination of Bubaline Herpesvirus 1, Bovine Herpesvirus 1 and Bovine Herpesvirus 5

 ,
, 
 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Selection of Virus Strains
2.2. Virus Propagation
2.3. DNA Extraction
2.4. Discrimination of BuHV-1, BoHV-1, and BoHV-5
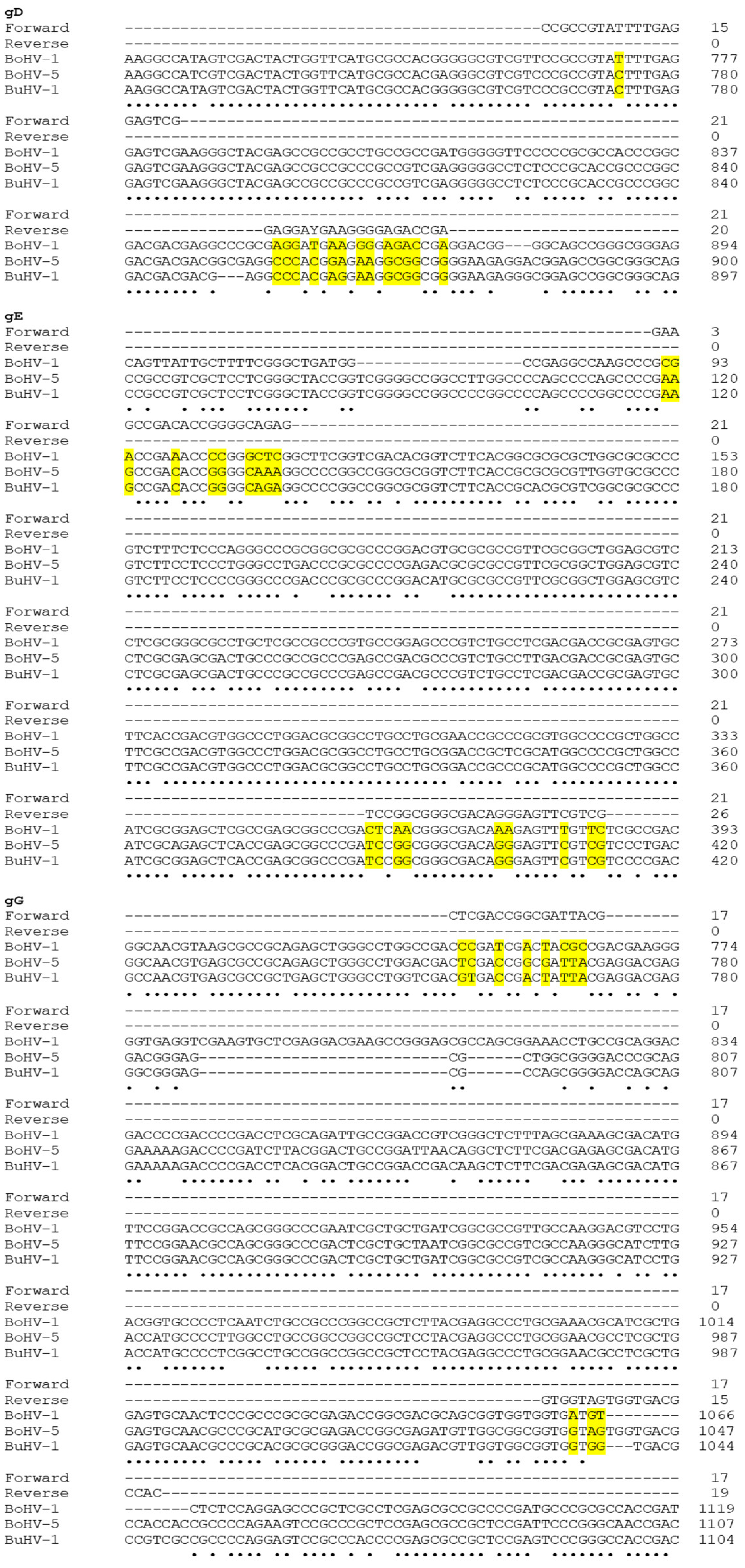
2.4.1. PCR of a Portion of gD Gene for Detecting BoHV-1
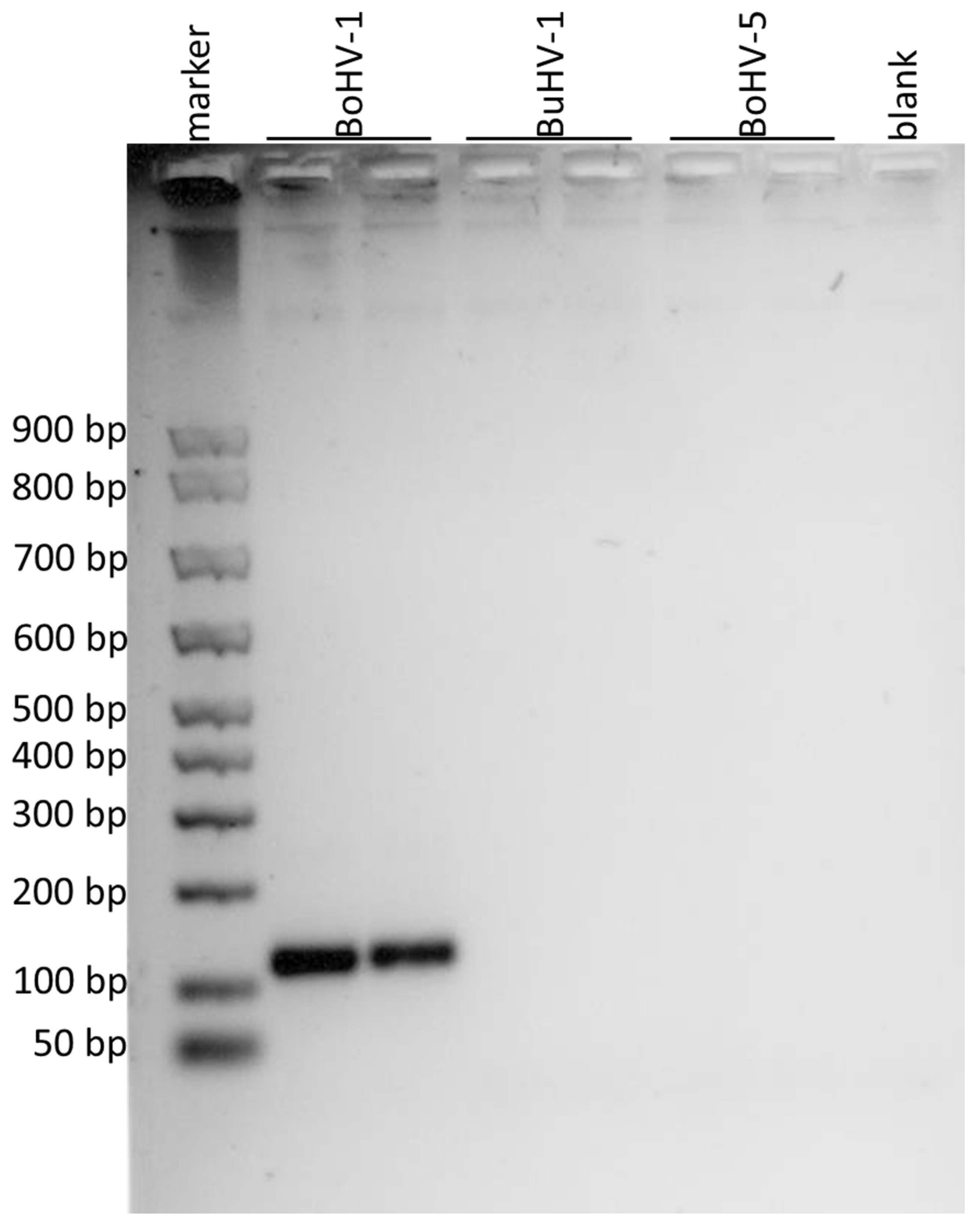
2.4.2. PCR of a Portion of gE Gene for Detecting BuHV-1 and BoHV-5
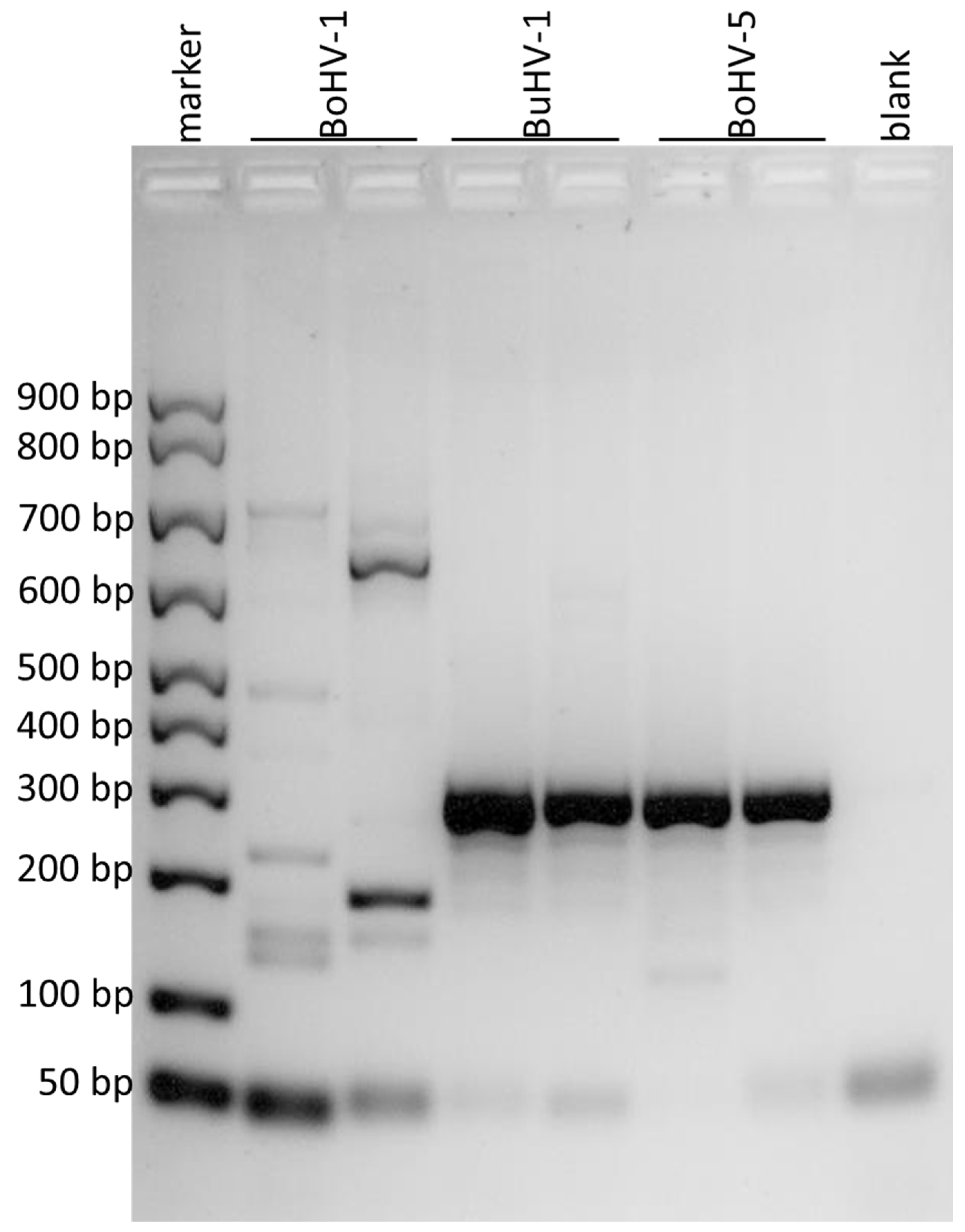
2.4.3. PCR of a Portion of gG Gene for Detecting BoHV-5
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Current ICTV Taxonomy Release. Available online: https://ictv.global/taxonomy/taxondetails?taxnode_id=202101440 (accessed on 19 January 2023).
- Kalman, D.; Egyed, L. PCR detection of bovine herpesvirus from nonbovine ruminants in hungary. J. Wildl. Dis. 2005, 41, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Intisar, K.S.; Ali, Y.H.; Khalafalla, A.I.; Rahman Mahasin, E.A.; Amin, A.S. Natural exposure of Dromedary camels in Sudan to infectious bovine rhinotracheitis virus (bovine herpes virus-1). Acta Trop. 2009, 111, 243–246. [Google Scholar] [CrossRef]
- Muylkens, B.; Thiry, J.; Kirten, P.; Schynts, F.; Thiry, E. Bovine herpesvirus 1 infection and infectious bovine rhinotracheitis. Vet. Res. 2007, 38, 181–209. [Google Scholar] [CrossRef] [PubMed]
- Thiry, J.; Widén, F.; Grégoire, F.; Linden, A.; Belák, S.; Thiry, E. Isolation and characterisation of a ruminant alphaherpesvirus closely related to bovine herpesvirus 1 in a free-ranging red deer. BMC Vet. Res. 2007, 3, 1–11. [Google Scholar] [CrossRef]
- Yatsentyuk, S.P.; Pchelnikov, A.V.; Safina, E.R.; Krasnikova, M.S. The first study on the occurrence of bovine herpesviruses in the wild fauna of the Moscow region, Russia. Vet. World 2022, 15, 2052–2058. [Google Scholar] [CrossRef] [PubMed]
- Boelaert, F.; Biront, P.; Soumare, B.; Dispas, M.; Vanopdenbosch, E.; Vermeersch, J.P.; Raskin, A.; Dufey, J.; Berkvens, D.; Kerhofs, P. Prevalence of bovine herpesvirus-1 in the Belgian cattle population. Prev. Vet. Med. 2000, 45, 285–295. [Google Scholar] [CrossRef]
- Esposito, C.; Fiorito, F.; Miletti, G.; Serra, F.; Balestrieri, A.; Cioffi, B.; Cerracchio, C.; Galiero, G.; De Carlo, E.; Amoroso, M.G.; et al. Involvement of herpesviruses in cases of abortion among water buffaloes in southern Italy. Vet. Res. Commun. 2022, 46, 719–729. [Google Scholar] [CrossRef]
- Marin, M.S.; Quintana, S.; Leunda, M.R.; Recavarren, M.; Pagnuco, I.; Späth, E.; Pérez, S.; Odeón, A. A new method for simultaneous detection and discrimination of Bovine herpesvirus types 1 (BoHV-1) and 5 (BoHV-5) using real time PCR with high resolution melting (HRM) analysis. J. Virol. Methods 2016, 227, 14–22. [Google Scholar] [CrossRef]
- Meyer, G.; Lemaire, M.; Ros, S.C.; Belak, K.; Gabriel, A.; Cassart, D.; Coignoul, F.; Belak, S.; Thiry, E. Comparative pathogenesis of acute and latent infections of calves with bovine herpesvirus types 1 and 5. Arch. Virol. 2001, 146, 633–652. [Google Scholar] [CrossRef]
- Thiry, J.; Keuser, V.; Muylkens, B.; Meurens, F.; Gogev, S.; Vanderplasschen, A.; Thiry, E. Ruminant alphaherpesviruses related to bovine herpesvirus 1. BMC Vet. Res. 2006, 37, 169–190. [Google Scholar] [CrossRef]
- De Carlo, E.; Letteriello, R.; Del Vecchio, V.; Giordanelli, M.R.; Magnino, S.; Fabbi, M.; Bazzocchi, C.; Bandi, C.; Galiero, G. Molecular characterisation of a field strain of bubaline herpesvirus isolated from buffaloes (Bubalus bubalis) after pharmacological reactivation. Vet. Rec. 2004, 154, 171–174. [Google Scholar] [CrossRef]
- Maidana, S.S.; Konrad, J.L.; Craig, M.I.; Zabal, O.; Mauroy, A.; Thiry, E.; Crudeli, G.; Romera, S.A. First report of isolation and molecular characterization of bubaline herpes virus 1 (BuHV-1) from Argentinean Water Buffaloes. Arch. Virol. 2014, 159, 2917–2923. [Google Scholar] [CrossRef]
- Scicluna, M.T.; Caprioli, A.; Saralli, G.; Manna, G.; Barone, A.; Cersini, A.; Cardeti, G.; Condoleo, R.U.; Autorino, G.L. Should the domestic buffalo (Bubalus bubalis) be considered in the epidemiology of Bovine Herpesvirus 1 infection. Vet. Microbiol. 2010, 143, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; O’Keefe, J.; Orr, D.; Loth, L.; Banks, M.; Wakeley, P.; West, D.; Card, R.; Ibata, G.; Van Maanen, K.; et al. Validation of a real-time PCR assay for the detection of bovine herpesvirus 1 in bovine semen. J. Virol. Methods 2007, 144, 103–108. [Google Scholar] [CrossRef]
- Diallo, I.S.; Corney, B.G.; Rodwell, B.J. Detection and differentiation of bovine herpesvirus 1 and 5 using a multiplex real time polymerase chain reaction. J. Virol. Methods 2011, 175, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Van Drunen Littel-van den Hurk, S. Rationale and perspectives on the success of vaccination against bovine herpesvirus-1. Vet. Microbiol. 2006, 113, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Romero-Salas, D.; Cruz-Romero, A.; Aguilar-Domínguez, M.; Ibarra-Priego, N.; Barradas-Piña, F.T.; Nogueira Domingues, L.; Castro-Arellano, I.; Lohmeyer, K.H.; Pérez de León, A.A. Seroepidemiology of Bovine Herpes Virus-1 Infection in Water Buffaloes from the state of Veracruz, Mexico. Trop. Biomed. 2018, 35, 541–552. [Google Scholar]
- Nogarol, C.; Bertolotti, L.; De Carlo, E.; Masoero, L.; Caruso, C.; Profiti, M.; Martucciello, A.; Galiero, G.; Cordioli, P.; Lelli, D.; et al. Expression and antigenic characterization of bubaline herpesvirus 1 (BuHV1) glycoprotein E and its potential application in the epidemiology and control of alphaherpesvirus infections in Mediterranean water buffalo. J. Virol. Methods 2014, 207, 16–21. [Google Scholar] [CrossRef]
- Kumar, N.; Chander, Y.; Riyesh, T.; Khandelwal, N.; Kumar, R.; Kumar, H.; Tripathi, B.N.; Barua, S. Isolation and characterization of bovine herpes virus 5 (BoHV5) from cattle in India. PLoS ONE 2020, 15, e0232093. [Google Scholar] [CrossRef]
- Ros, C.; Belak, S. Characterization of the glycoprotein B gene from ruminant alphaherpesviruses. Virus Genes 2002, 24, 99–105. [Google Scholar] [CrossRef]
- Campos, F.S.; Franco, A.C.; Hubner, S.O.; Oliveira, M.T.; Silva, A.D.; Esteves, P.A.; Roehe, P.M.; Rijsewijk, F.A.M. High prevalence of co-infections with bovine herpesvirus 1 and 5 found in cattle in southern Brazil. Vet. Microbiol. 2009, 139, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Claus, M.P.; Alfieri, A.F.; Folgueras-Flatschart, A.V.; Rezler Wosiacki, S.; Médici, K.C.; Alfieri, A.A. Rapid detection and differentiation of bovine herpesvirus 1 and 5 glycoprotein C gene in clinical specimens by multiplex-PCR. J. Virol. Methods 2005, 128, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Ros, C.; Riquelme, M.E.; Ohman Forslund, K.; Belàk, S. Improved detection of five closely related ruminant alphaherpesviruses by specific amplification of viral genomic sequences. J. Virol. Methods 1999, 83, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Ros, C.; Belàk, S. Studies of genetic relationships between bovine, caprine, cervine, and rangiferine alphaherpesviruses and improved molecular methods for virus detection and identification. J. Clin. Microbiol. 1999, 37, 1247–1253. [Google Scholar] [CrossRef] [PubMed]
- Caruso, C.; Prato, R.; Ingravalle, F.; Vecchio, D.; Sciarra, A.; Ternavasio, M.; Ceccarelli, L.; Martucciello, A.; Galiero, G.; De Carlo, E.; et al. Prevalence of antibodies against Bubaline herpesvirus BuHV 1 among Mediterranean water buffalo Bubalus bubalis with implications in buffalo trade. Vet. Q. 2016, 36, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Camero, M.; Lanave, M.S.; Lucente, M.; Losurdo, G.; Di Paola, G.; Lorusso, E.; Martella, V.; Buonavoglia, C.; Tempesta, M. Bubaline alphaherpesvirus 1 induces a latent/reactivable infection in goats. Comp. Immunol. Microbiol. Infect. Dis. 2019, 62, 54–57. [Google Scholar] [CrossRef]
- Camero, M.; Larocca, V.; Losurdo, M.; Lorusso, E.; Patruno, G.; Staffa, V.N.; Martella, V.; Buonavoglia, C.; Tempesta, M. Goats are susceptible to Bubaline alphaherpesvirus 1 infection Results of an experimental study. Comp. Immunol. Microbiol. Infect. Dis. 2017, 50, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Fusco, G.; Amoroso, M.G.; Aprea, G.; Veneziano, V.; Guarino, A.; Galiero, G.; Viscardi, M. First report of natural BoHV-1 infection in water buffalo. Vet. Rec. 2005, 177, 152. [Google Scholar] [CrossRef]
- Maidana, S.S.; Delgado, F.; Vagnoni, L.; Mauroy, A.; Thiry, E.; Romera, S. Cattle are a potential reservoir of bubaline herpesvirus 1 (BuHV1). Vet. Rec. Open 2016, 3, e000162. [Google Scholar] [CrossRef]
- Maresca, C.; Scoccia, E.; Dettori, A.; Felici, A.; Guarcini, R.; Petrini, S.; Quaglia, A.; Filippini, G. National surveillance plan for infectious bovine rhinotracheitis (IBR) in autochthonous Italian cattle breeds Results of first year of activity. Vet. Microbiol. 2018, 219, 150–153. [Google Scholar] [CrossRef]
- Rola, J.; Polak, M.; Zmudzinski, J. Amplification of DNA of BHV 1 isolated from semen of naturally infected bulls. Bull. Vet. Inst. 2003, 47, 71–75. [Google Scholar]
- Wernike, K.; Hoffmann, B.; Kalthoff, D.; König, P.; Beer, M. Development and validation of a triplex real-time PCR assay for the rapid detection and differentiation of wild-type and glycoprotein E-deleted vaccine strains of Bovine herpesvirus type. J. Virol. Methods 2011, 174, 77–84. [Google Scholar] [CrossRef]
- Keuser, V.; Schynts, F.; Detry, B.; Collard, A.; Robert, B.; Vanderplasschen, A.; Pastoret, P.P.; Thiry, E. Improved Antigenic Methods for Differential Diagnosis of Bovine, Caprine, and Cervine Alphaherpesviruses Related to Bovine Herpesvirus 1. J. Clin. Microbiol. 2004, 42, 1228–1235. [Google Scholar] [CrossRef] [PubMed]
- Maidana, S.S.; Ladelfa, M.F.; Pérez, S.E.; Lomónaco, P.M.; Del Médico Zajac, M.P.; Odeón, A.; Blanco Viera, J.; Combessies, G.; Fondevila, N.; Palacios, M.; et al. Characterization of BoHV-5 field strains circulation and report of transient specific subtype of bovine herpesvirus 5 in Argentina. BMC Vet. Res. 2011, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Strains | gD Gene | gE Gene | gG Gene |
|---|---|---|---|
| BuHV1 | 295 bp | ||
| BoHV1 | 110 bp | ||
| BoHV5 | 295 bp | 296 bp |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oberto, F.; Carella, E.; Caruso, C.; Acutis, P.L.; Lelli, D.; Bertolotti, L.; Masoero, L.; Peletto, S. A Qualitative PCR Assay for the Discrimination of Bubaline Herpesvirus 1, Bovine Herpesvirus 1 and Bovine Herpesvirus 5. Microorganisms 2023, 11, 577. https://doi.org/10.3390/microorganisms11030577
Oberto F, Carella E, Caruso C, Acutis PL, Lelli D, Bertolotti L, Masoero L, Peletto S. A Qualitative PCR Assay for the Discrimination of Bubaline Herpesvirus 1, Bovine Herpesvirus 1 and Bovine Herpesvirus 5. Microorganisms. 2023; 11(3):577. https://doi.org/10.3390/microorganisms11030577
Chicago/Turabian StyleOberto, Francesca, Emanuele Carella, Claudio Caruso, Pier Luigi Acutis, Davide Lelli, Luigi Bertolotti, Loretta Masoero, and Simone Peletto. 2023. "A Qualitative PCR Assay for the Discrimination of Bubaline Herpesvirus 1, Bovine Herpesvirus 1 and Bovine Herpesvirus 5" Microorganisms 11, no. 3: 577. https://doi.org/10.3390/microorganisms11030577
APA StyleOberto, F., Carella, E., Caruso, C., Acutis, P. L., Lelli, D., Bertolotti, L., Masoero, L., & Peletto, S. (2023). A Qualitative PCR Assay for the Discrimination of Bubaline Herpesvirus 1, Bovine Herpesvirus 1 and Bovine Herpesvirus 5. Microorganisms, 11(3), 577. https://doi.org/10.3390/microorganisms11030577