
Spring Viremia of Carp Virus Infection Induces Carp IL-10 Expression, Both In Vitro and In Vivo


Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses, Cells and Fish
2.2. Reagents
2.3. Stimulation of EPC and cHK Cells with SVCV Infection
2.4. SVCV Inoculation of Carp
2.5. Inhibition of Signal Transduction Pathways
2.6. Quantification of Carp IL-10 Gene Expression Using RT-qPCR
2.7. Quantification of IL-10 Protein Expression Using ELISA
2.8. Statistical Analysis
3. Results
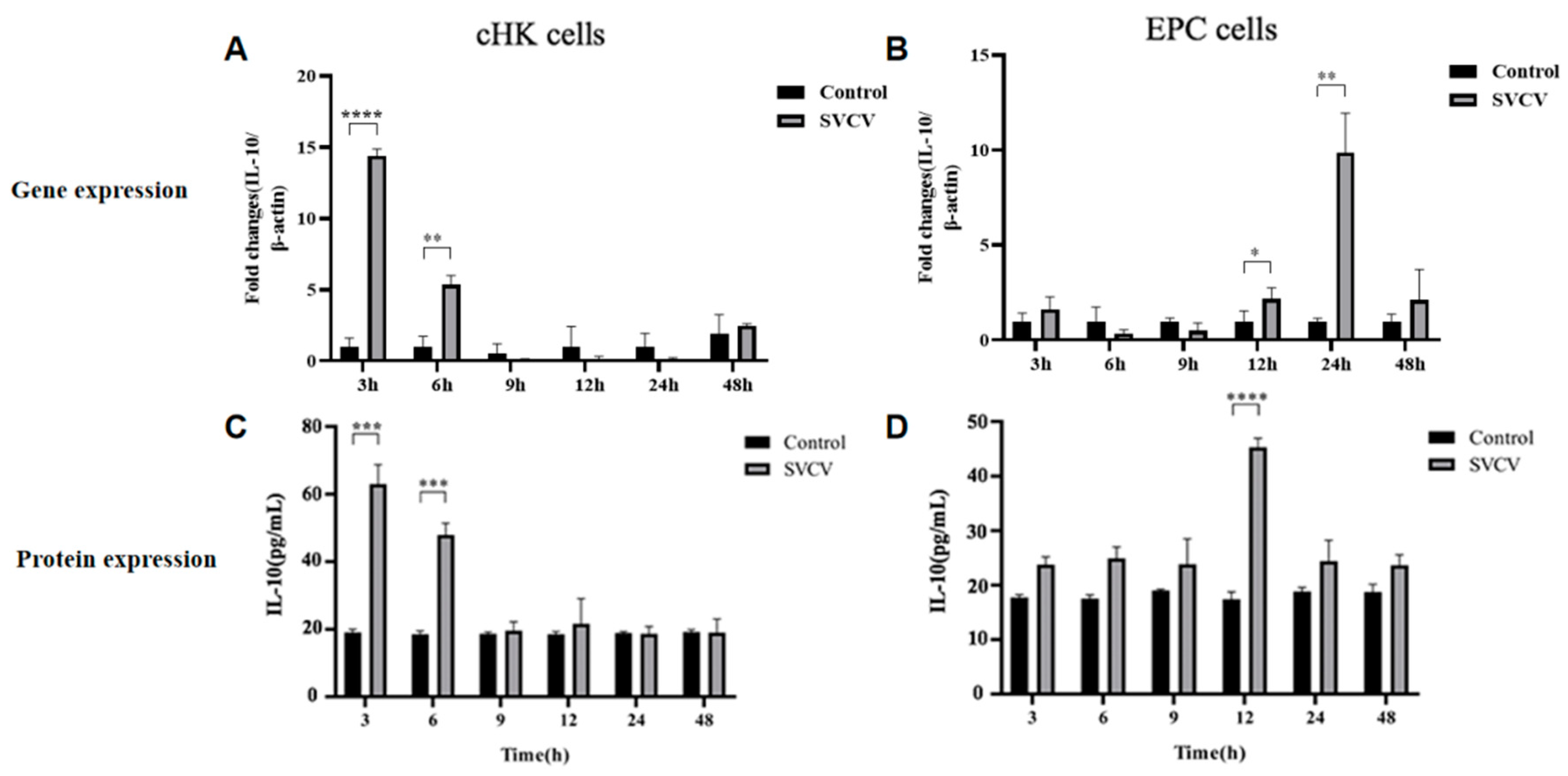
3.1. SVCV Infection Induced Carp IL-10 Production at mRNA and Protein Levels in EPC and cHK Cells
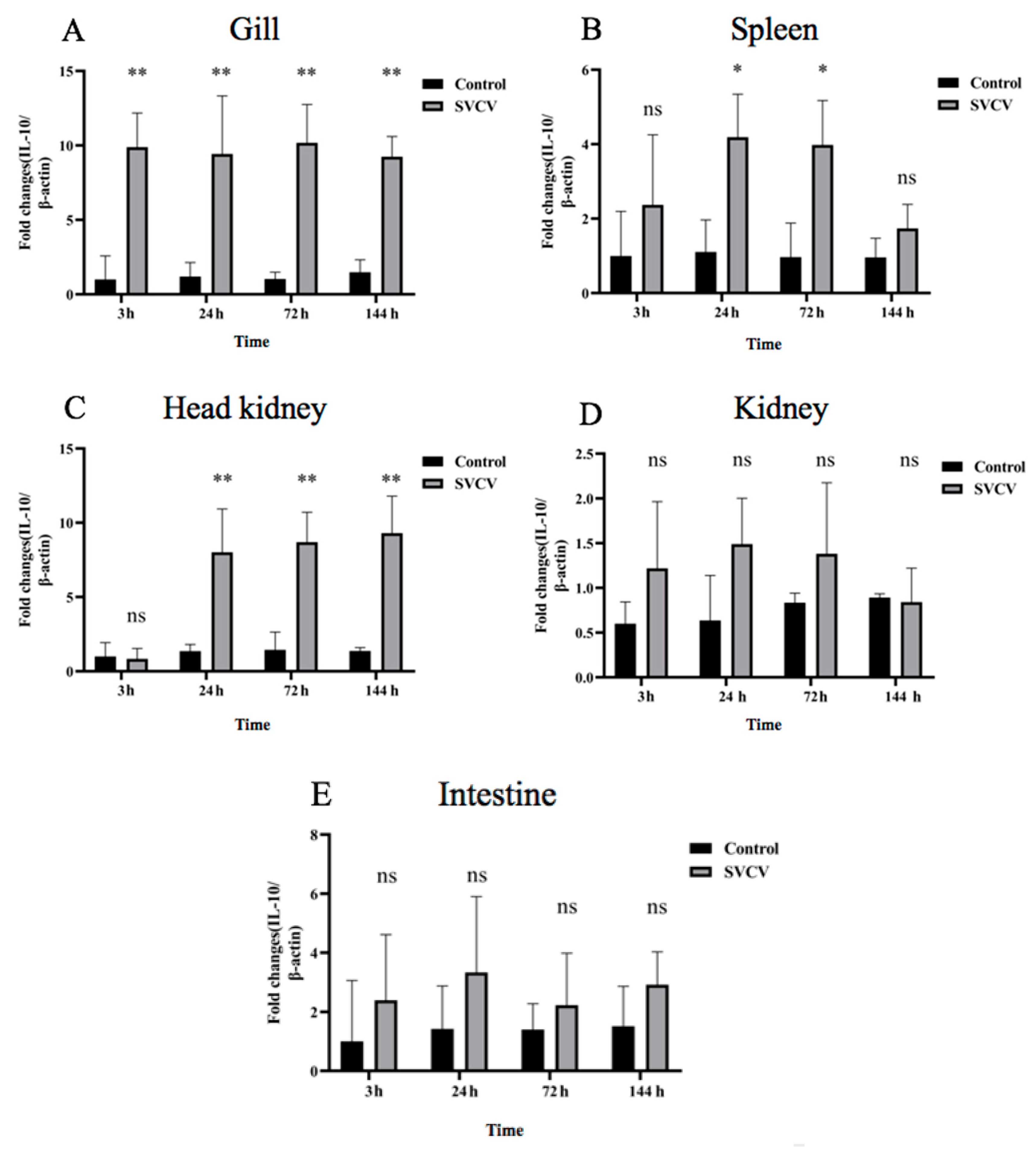
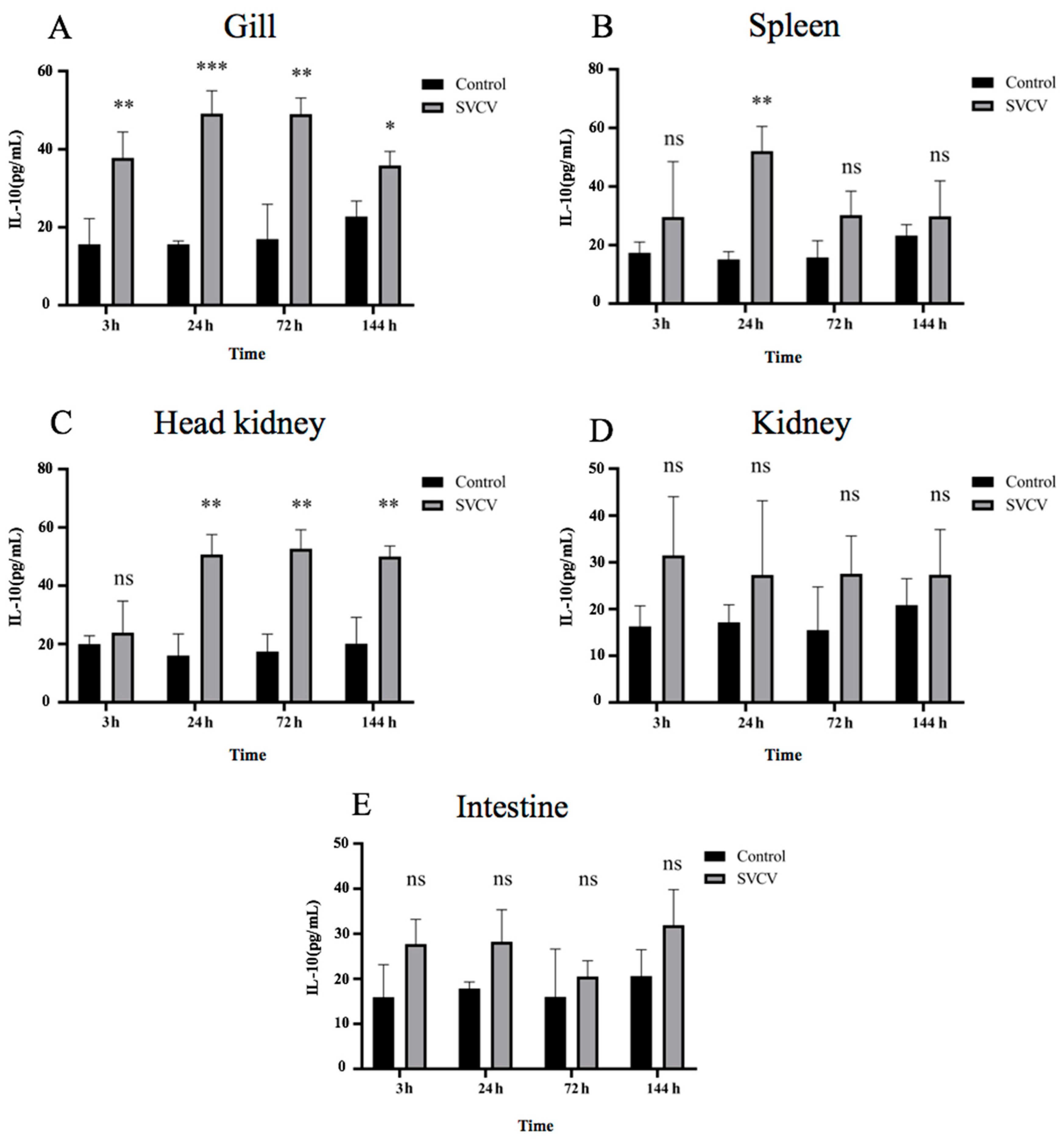
3.2. SVCV Infection Induced Carp IL-10 Production at mRNA and Protein Levels in Carp Tissues
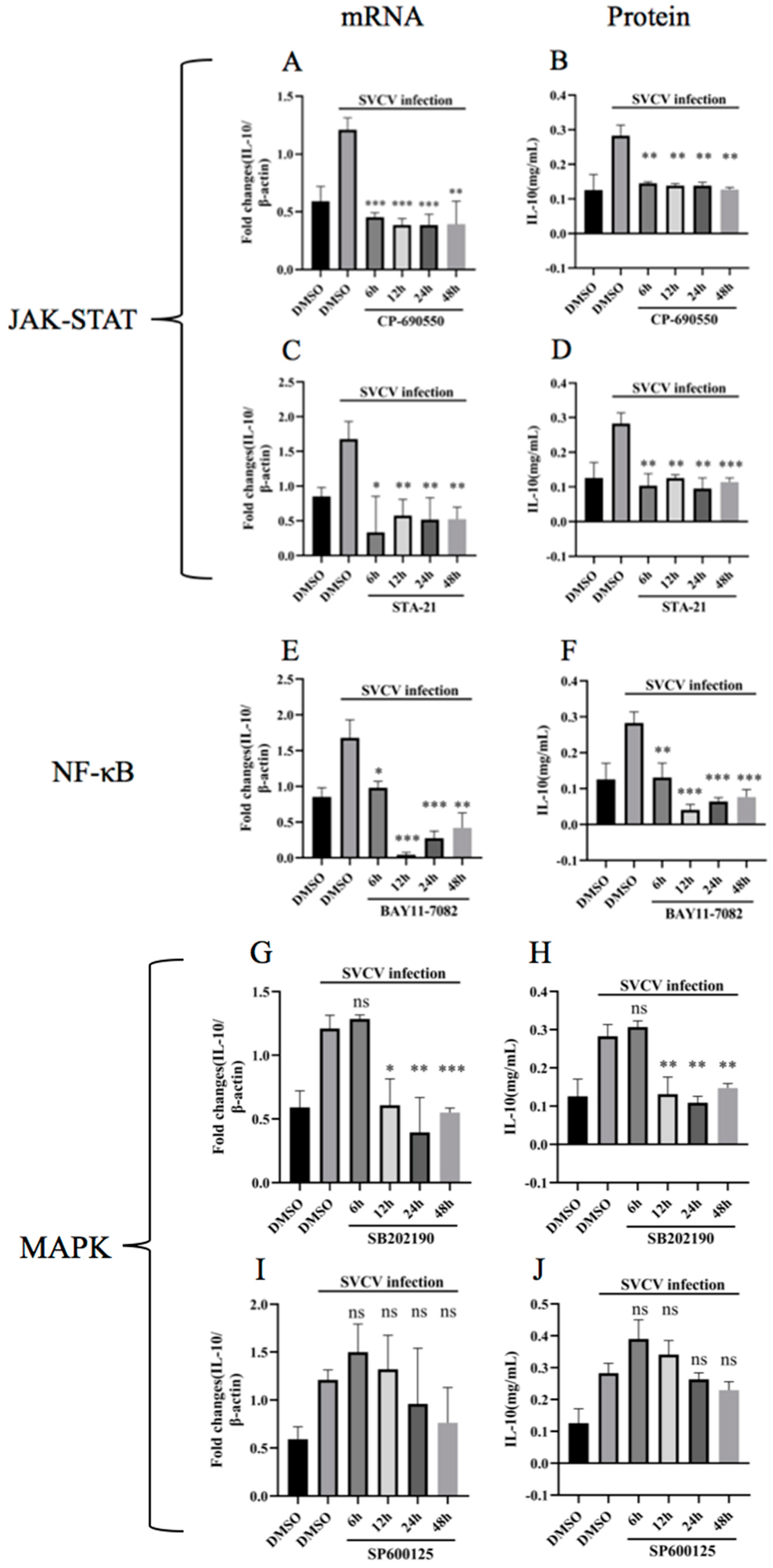
3.3. Inhibition of JAK-STAT, NF-κB and p38MAPK Pathways Hindered the Upregulation of Carp IL-10
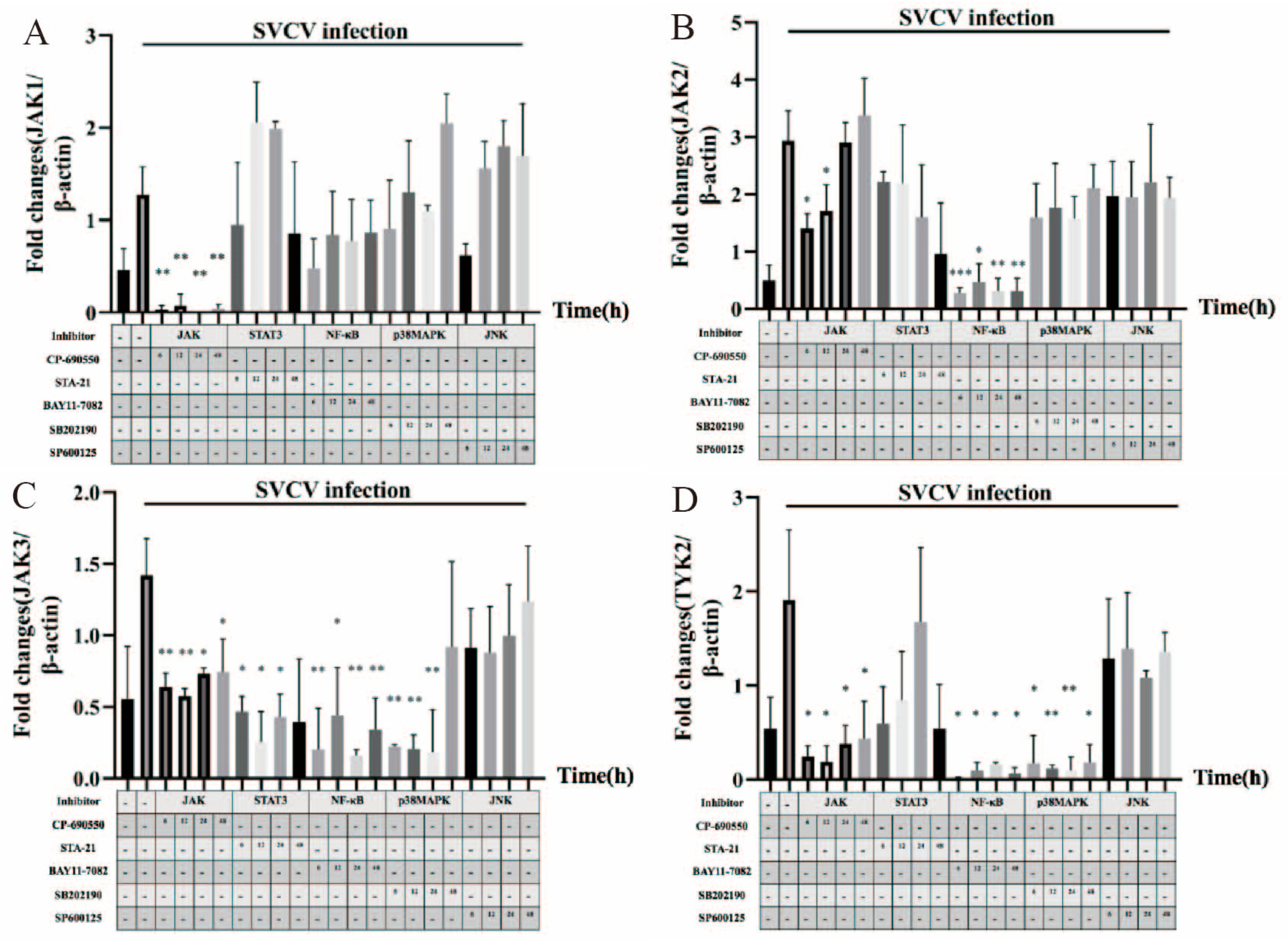
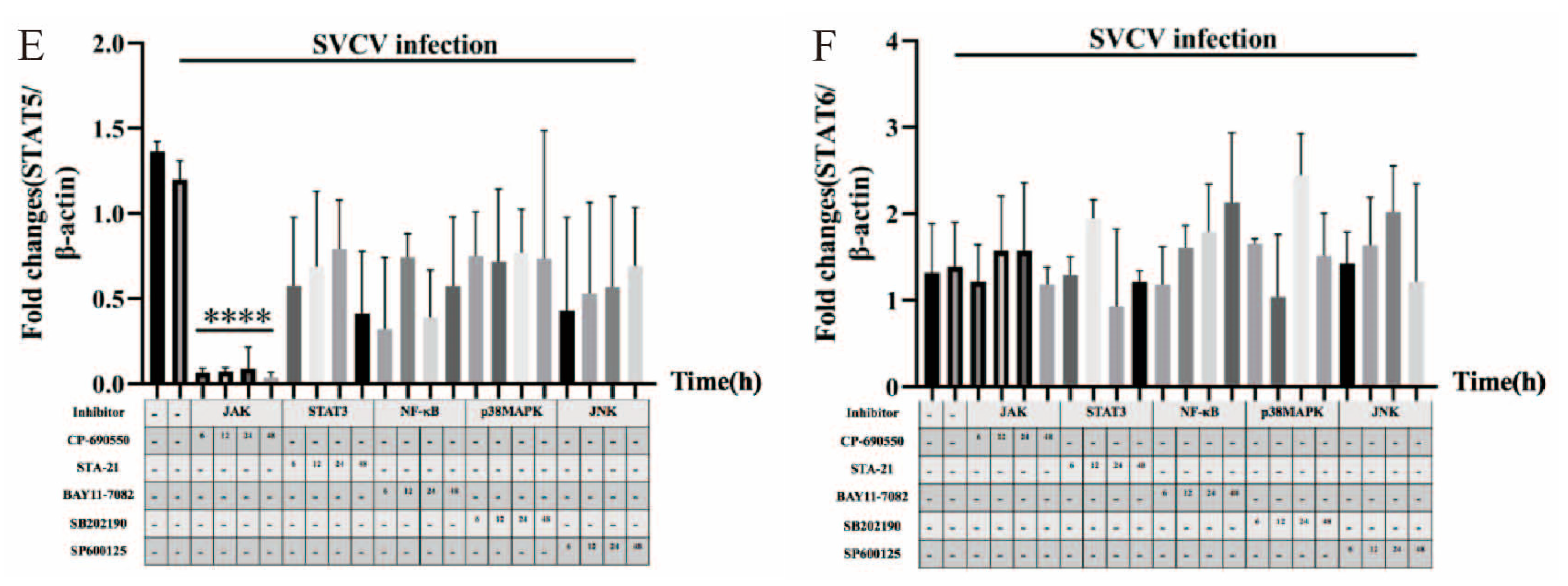
3.4. JAK1, JAK2, JAK3, TYK2, STAT5 and STAT6 Were Involved in SVCV-Induced Carp IL-10 Expression
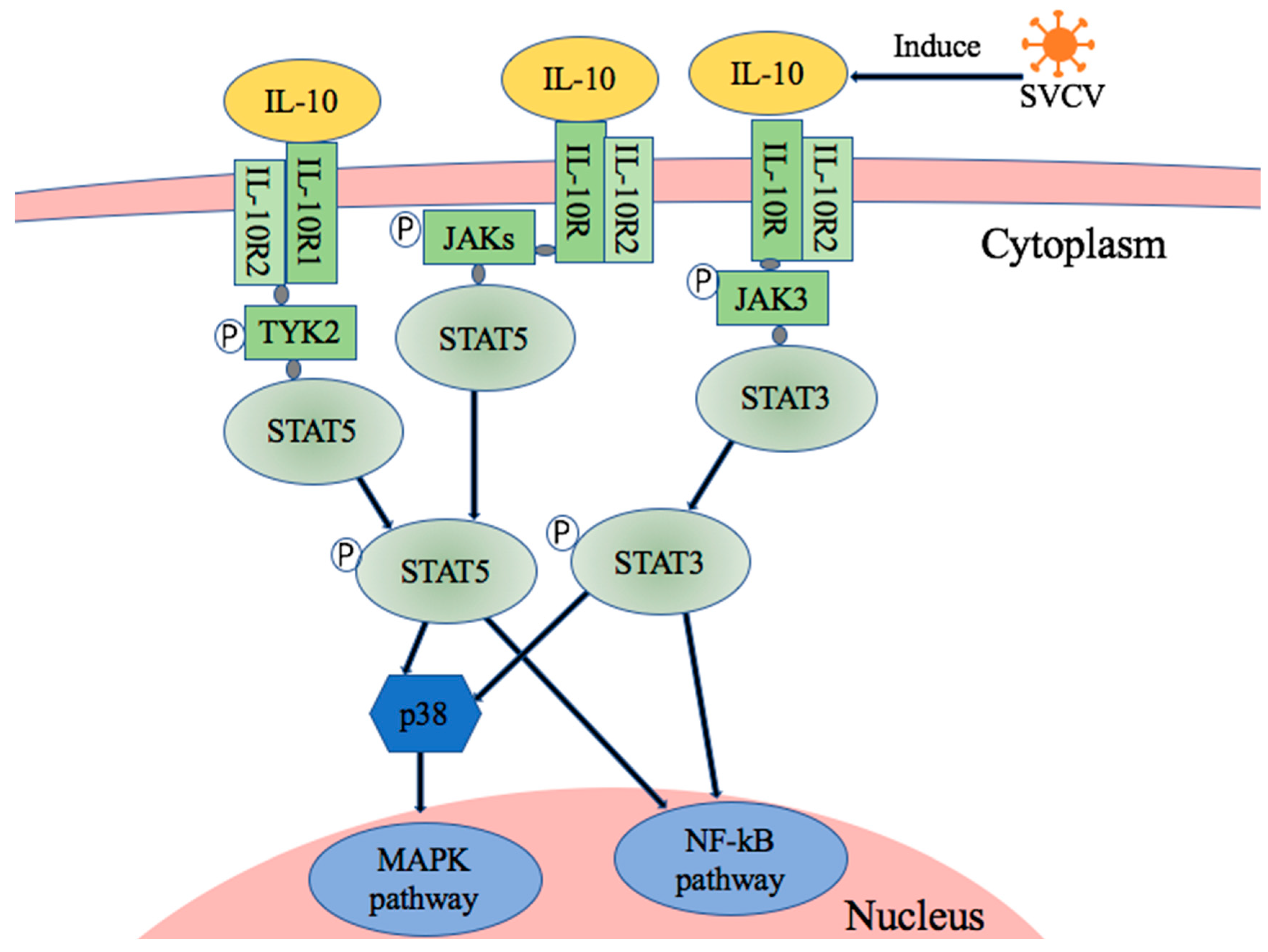
4. Discussions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, C.; Li, L.; Jin, L.; Yuan, J. Heme Oxygenase-1 inhibits spring viremia of carp virus replication through carbon monoxide mediated cyclic GMP/Protein kinase G signaling pathway. Fish Shellfish Immunol. 2018, 79, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Song, D.W.; Liu, G.L.; Shan, L.P.; Qiu, T.X.; Chen, J. Hydroxycoumarin efficiently inhibits spring viraemia of carp virus infection in vitro and in vivo. Zool. Res. 2020, 41, 395–409. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Yang, Y.; Nie, H.; Li, L.; Gu, W.; Lin, L.; Zou, M.; Liu, X.; Wang, M.; Gu, Z. Transcriptome analysis of epithelioma papulosum cyprini cells after SVCV infection. BMC Genom. 2014, 15, 935. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, D.F.; Bond, M.W.; Mosmann, T.R. Two types of mouse T helper cell. IV. Th2 clones secrete a factor that inhibits cytokine production by Th1 clones. J. Exp. Med. 1989, 170, 2081–2095. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Su, N.; Wang, M.; Xie, P.; Shi, Z.; Li, L. Down-regulation of heme oxygenase-1 by SVCV infection. Fish Shellfish Immunol. 2012, 32, 301–306. [Google Scholar] [CrossRef]
- Piazzon, M.C.; Savelkoul, H.S.; Pietretti, D.; Wiegertjes, G.F.; Forlenza, M. Carp Il10 Has Anti-Inflammatory Activities on Phagocytes, Promotes Proliferation of Memory T Cells, and Regulates B Cell Differentiation and Antibody Secretion. J. Immunol. 2015, 194, 187–199. [Google Scholar] [CrossRef]
- Aborsangaya, K.B.; Dembinski, I.; Khatkar, S.; Alphonse, M.P.; Nickerson, P.; Rempel, J.D. Impact of aboriginal ethnicity on HCV core-induced IL-10 synthesis: Interaction with IL-10 gene polymorphisms. Hepatology 2007, 45, 623–630. [Google Scholar] [CrossRef]
- Brockman, M.A.; Kwon, D.S.; Tighe, D.P.; Pavlik, D.F.; Rosato, P.C.; Sela, J.; Porichis, F.; Le Gall, S.; Waring, M.T.; Moss, K.; et al. IL-10 is up-regulated in multiple cell types during viremic HIV infection and reversibly inhibits virus-specific T cells. Blood 2009, 114, 346–356. [Google Scholar] [CrossRef]
- Lu, L.; Zhang, H.; Dauphars, D.J.; He, Y.W. A Potential Role of Interleukin 10 in COVID-19 Pathogenesis. Trends Immunol. 2021, 42, 3–5. [Google Scholar] [CrossRef]
- Ouyang, P.; Rakus, K.; van Beurden, S.J.; Westphal, A.H.; Davison, A.J.; Gatherer, D.; Vanderplasschen, A.F. IL-10 encoded by viruses: A remarkable example of independent acquisition of a cellular gene by viruses and its subsequent evolution in the viral genome. J. Gen. Virol. 2014, 95, 245–262. [Google Scholar] [CrossRef]
- Guo, C.J.; He, J.; He, J.G. The immune evasion strategies of fish viruses. Fish Shellfish Immunol. 2019, 86, 772–784. [Google Scholar] [CrossRef] [PubMed]
- Bjorgen, H.; Kumar, S.; Gunnes, G.; Press, C.M.; Rimstad, E.; Koppang, E.O. Immunopathological characterization of red focal changes in Atlantic salmon Salmo salar white muscle. Vet. Immunol. Immunopathol. 2020, 222, 9. [Google Scholar] [CrossRef] [PubMed]
- Collet, B.; Urquhart, K.; Monte, M.; Collins, C.; Perez, S.G.; Secombes, C.J.; Hall, M. Individual Monitoring of Immune Response in Atlantic Salmon Salmo salar following Experimental Infection with Infectious Salmon Anaemia Virus (ISAV). PLoS ONE 2015, 10, 14. [Google Scholar] [CrossRef] [PubMed]
- Buonocore, F.; Randelli, E.; Bird, S.; Secombes, C.J.; Facchiano, A.; Costantini, S.; Scapigliati, G. Interleukin-10 expression by real-time PCR and homology modelling analysis in the European sea bass (Dicentrarchus labrax L.). Aquaculture 2007, 270, 512–522. [Google Scholar] [CrossRef]
- Grayfer, L.; Hodgkinson, J.W.; Hitchen, S.J.; Belosevic, M. Characterization and functional analysis of goldfish (Carassius auratus L.) interleukin-10. Mol. Immunol. 2011, 48, 563–571. [Google Scholar] [CrossRef]
- Inoue, Y.; Kamota, S.; Ito, K.; Yoshiura, Y.; Ototake, M.; Moritomo, T.; Nakanishi, T. Molecular cloning and expression analysis of rainbow trout (Oncorhynchus mykiss) interleukin-10 cDNAs. Fish Shellfish Immunol. 2005, 18, 335–344. [Google Scholar] [CrossRef]
- Karan, S.; Dash, P.; Kaushik, H.; Sahoo, P.K.; Garg, L.C.; Dixit, A. Structural and Functional Characterization of Recombinant Interleukin-10 from Indian Major Carp Labeo rohita. J. Immunol. Res. 2016, 2016, 3962596. [Google Scholar] [CrossRef]
- Savan, R.; Igawa, D.; Sakai, M. Cloning, characterization and expression analysis of interleukin-10 from the common carp, Cyprinus carpio L. Eur. J. Biochem. 2003, 270, 4647–4654. [Google Scholar] [CrossRef]
- Wei, H.; Yang, M.; Zhao, T.; Wang, X.; Zhou, H. Functional expression and characterization of grass carp IL-10: An essential mediator of TGF-beta1 immune regulation in peripheral blood lymphocytes. Mol. Immunol. 2013, 53, 313–320. [Google Scholar] [CrossRef]
- Zou, J.; Clark, M.S.; Secombes, C.J. Characterisation, expression and promoter analysis of an interleukin 10 homologue in the puffer fish, Fugu rubripes. Immunogenetics 2003, 55, 325–335. [Google Scholar] [CrossRef]
- Zheng, L.P.; Geng, Y.; Yu, Z.H.; Wang, K.Y.; Ou, Y.P.; Chen, D.F.; Huang, X.L.; Deng, L.J.; Gan, W.X.; Fang, J.; et al. First report of spring viremia of carp virus in Percocypris pingi in China. Aquaculture 2018, 493, 214–218. [Google Scholar] [CrossRef]
- Dong, C.; Weng, S.; Shi, X.; Xu, X.; Shi, N.; He, J. Development of a mandarin fish Siniperca chuatsi fry cell line suitable for the study of infectious spleen and kidney necrosis virus (ISKNV). Virus Res. 2008, 135, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, P.; Yang, R.; Yin, L.; Geng, Y.; Lai, W.; Huang, X.; Chen, D.; Fang, J.; Chen, Z.; Tang, L.; et al. Molecular characterization of Cyprinid herpesvirus 3 encoded viral interleukin10. Fish Shellfish. Immunol. 2019, 89, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Wu, P. Cloning and Tissue Expression Pattern Analysis of JAK/STAT Molecule in Carp. Master’s Thesis, Central China Normal University, Wuhan, China, 2014. [Google Scholar]
- Sun, J.; Wang, J.; Li, L.; Wu, Z.; Chen, X.; Yuan, J. ROS induced by spring viraemia of carp virus activate the inflammatory response via the MAPK/AP-1 and PI3K signaling pathways. Fish Shellfish Immunol. 2020, 101, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Giri, S.S.; Sen, S.S.; Chi, C.; Kim, H.J.; Yun, S.; Park, S.C.; Sukumaran, V. Effect of guava leaves on the growth performance and cytokine gene expression of Labeo rohita and its susceptibility to Aeromonas hydrophila infection. Fish Shellfish Immunol. 2015, 46, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.C.; Shao, Y.Q.; Huang, Y.Q.; Jiang, S.G. Cloning, characterization and expression analysis of interleukin-10 from the zebrafish (Danio rerion). J. Biochem. Mol. Biol. 2005, 38, 571–576. [Google Scholar] [CrossRef]
- Ingerslev, H.C.; Ronneseth, A.; Pettersen, E.F.; Wergeland, H.I. Differential Expression of Immune Genes in Atlantic SalmonSalmo salar Challenged Intraperitoneally or by Cohabitation with IPNV. Scand. J. Immunol. 2009, 69, 90–98. [Google Scholar] [CrossRef]
- Huo, H.J.; Chen, S.N.; Li, L.; Nie, P. Functional characterization of IL-10 and its receptor subunits in a perciform fish, the mandarin fish, Siniperca chuatsi. Dev. Comp. Immunol. 2019, 97, 64–75. [Google Scholar] [CrossRef]
- Wang, J.; Chen, Z.F.; Li, M.; Zhang, Z.W.; Song, Y.; Xu, W.T.; Wang, L.; Chen, S.L. Genome-wide identification, immune response profile and functional characterization of IL-10 from spotted knifejaw Oplegnathus punctatus during host defense against bacterial and viral infection. Fish Shellfish Immunol. 2022, 124, 513–524. [Google Scholar] [CrossRef]
- Xin, P.; Xu, X.; Deng, C.; Liu, S.; Wang, Y.; Zhou, X.; Ma, H.; Wei, D.; Sun, S. The role of JAK/STAT signaling pathway and its inhibitors in diseases. Int. Immunopharmacol. 2020, 80, 106210. [Google Scholar] [CrossRef]
- Michalski, M.N.; Koh, A.J.; Weidner, S.; Roca, H.; McCauley, L.K. Modulation of Osteoblastic Cell Efferocytosis by Bone Marrow Macrophages. J. Cell. Biochem. 2016, 117, 2697–2706. [Google Scholar] [CrossRef]
- Schmetterer, K.G.; Pickl, W.F. The IL-10/STAT3 axis: Contributions to immune tolerance by thymus and peripherally derived regulatory T-cells. Eur. J. Immunol. 2017, 47, 1256–1265. [Google Scholar] [CrossRef] [PubMed]
- Islam, H.; Chamberlain, T.C.; Mui, A.L.; Little, J.P. Elevated Interleukin-10 Levels in COVID-19: Potentiation of Pro-Inflammatory Responses or Impaired Anti-Inflammatory Action? Front. Immunol. 2021, 12, 677008. [Google Scholar] [CrossRef] [PubMed]
- Piazzon, M.C.; Wentzel, A.S.; Wiegertjes, G.F.; Forlenza, M. Carp Il10a and Il10b exert identical biological activities in vitro, but are differentially regulated in vivo. Dev. Comp. Immunol. 2017, 67, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Carey, A.J.; Tan, C.K.; Ulett, G.C. Infection-induced IL-10 and JAK-STAT: A review of the molecular circuitry controlling immune hyperactivity in response to pathogenic microbes. JAKSTAT 2012, 1, 159–167. [Google Scholar] [CrossRef]
- Nascimento, D.S.; do Vale, A.; Tomas, A.M.; Zou, J.; Secombes, C.J.; dos Santos, N.M. Cloning, promoter analysis and expression in response to bacterial exposure of sea bass (Dicentrarchus labrax L.) interleukin-12 p40 and p35 subunits. Mol. Immunol. 2007, 44, 2277–2291. [Google Scholar] [CrossRef]
- Song, S.; Bi, J.; Wang, D.; Fang, L.; Zhang, L.; Li, F.; Chen, H.; Xiao, S. Porcine reproductive and respiratory syndrome virus infection activates IL-10 production through NF-kappaB and p38 MAPK pathways in porcine alveolar macrophages. Dev. Comp. Immunol. 2013, 39, 265–272. [Google Scholar] [CrossRef]
- Agrawal, S.; Gollapudi, S.; Su, H.; Gupta, S. Leptin activates human B cells to secrete TNF-alpha, IL-6, and IL-10 via JAK2/STAT3 and p38MAPK/ERK1/2 signaling pathway. J. Clin. Immunol. 2011, 31, 472–478. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5′–3′) | Accession No. |
|---|---|---|
| β-actin-F | GACCTGTATGCCAACACTGTAT | M24113 |
| β-actin-R | TCCTGCTTGCTAATCCACATC | |
| IL-10-F | AGTCCTTATGGCTGTCACGTCATG | AB110780 |
| IL-10-R | TTGAGTGCAAGTGGTCCTTCTGG | |
| JAK1-F | AGGGGACACCTCTACTGGATGC | AH004872 |
| JAK1-R | GTGTGAGAAGTTACGCTGCTTA | |
| JAK2-F | CGGAGTGTCACCAGTCTAC | KJ782027 |
| JAK2-R | GGTAAGACATAACACAGTCATCC | |
| JAK3-F | GGCATCAGAGGACCTTTCATAC | AF148993 |
| JAK3-R | CTGCCATTCCCAAGCATTCCTG | |
| TYK2-F | TGAGGGTGAGGTGACTGCTGAAG | KJ782031 |
| TYK2-R | AGCGGTTCCTTTTCACCTAATCC | |
| STAT5-F | CCTGGGATGGAATAGATTTAGAG | KJ782032 |
| STAT5-R | GACAGCGGTCATACGTGCTCTTTAG | |
| STAT6-F | AATGACCCCGCAGTTACAGTTC | KJ782033 |
| STAT6-R | GTTGTAGTTTGACCCTCTCCAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouyang, P.; Tao, Y.; Wei, W.; Li, Q.; Liu, S.; Ren, Y.; Huang, X.; Chen, D.; Geng, Y. Spring Viremia of Carp Virus Infection Induces Carp IL-10 Expression, Both In Vitro and In Vivo. Microorganisms 2023, 11, 2812. https://doi.org/10.3390/microorganisms11112812
Ouyang P, Tao Y, Wei W, Li Q, Liu S, Ren Y, Huang X, Chen D, Geng Y. Spring Viremia of Carp Virus Infection Induces Carp IL-10 Expression, Both In Vitro and In Vivo. Microorganisms. 2023; 11(11):2812. https://doi.org/10.3390/microorganisms11112812
Chicago/Turabian StyleOuyang, Ping, Yu Tao, Wenyan Wei, Qiunan Li, Shuya Liu, Yongqiang Ren, Xiaoli Huang, Defang Chen, and Yi Geng. 2023. "Spring Viremia of Carp Virus Infection Induces Carp IL-10 Expression, Both In Vitro and In Vivo" Microorganisms 11, no. 11: 2812. https://doi.org/10.3390/microorganisms11112812
APA StyleOuyang, P., Tao, Y., Wei, W., Li, Q., Liu, S., Ren, Y., Huang, X., Chen, D., & Geng, Y. (2023). Spring Viremia of Carp Virus Infection Induces Carp IL-10 Expression, Both In Vitro and In Vivo. Microorganisms, 11(11), 2812. https://doi.org/10.3390/microorganisms11112812