
Understanding the Effect of Different Glucose Concentrations in the Oligotrophic Bacterium Bacillus subtilis BS-G1 through Transcriptomics Analysis

Abstract
1. Introduction
2. Materials and Methods
2.1. Strain
2.2. Growth Curve and Determination of Residual Sugar Content
2.3. Construction of Transcriptome and Processing of Biological Information
2.4. Reading Mapping and Analysis
2.5. Bioinformatics Analysis
2.6. Quantitative Real-Time PCR (qRT-PCR)
2.7. Statistical Analysis
3. Results
3.1. Growth Curves and Glucose Utilization
3.2. Transcriptome Analysis of B. subtilis under Different Glucose Concentrations
3.3. Transcriptome Differential Gene Annotation
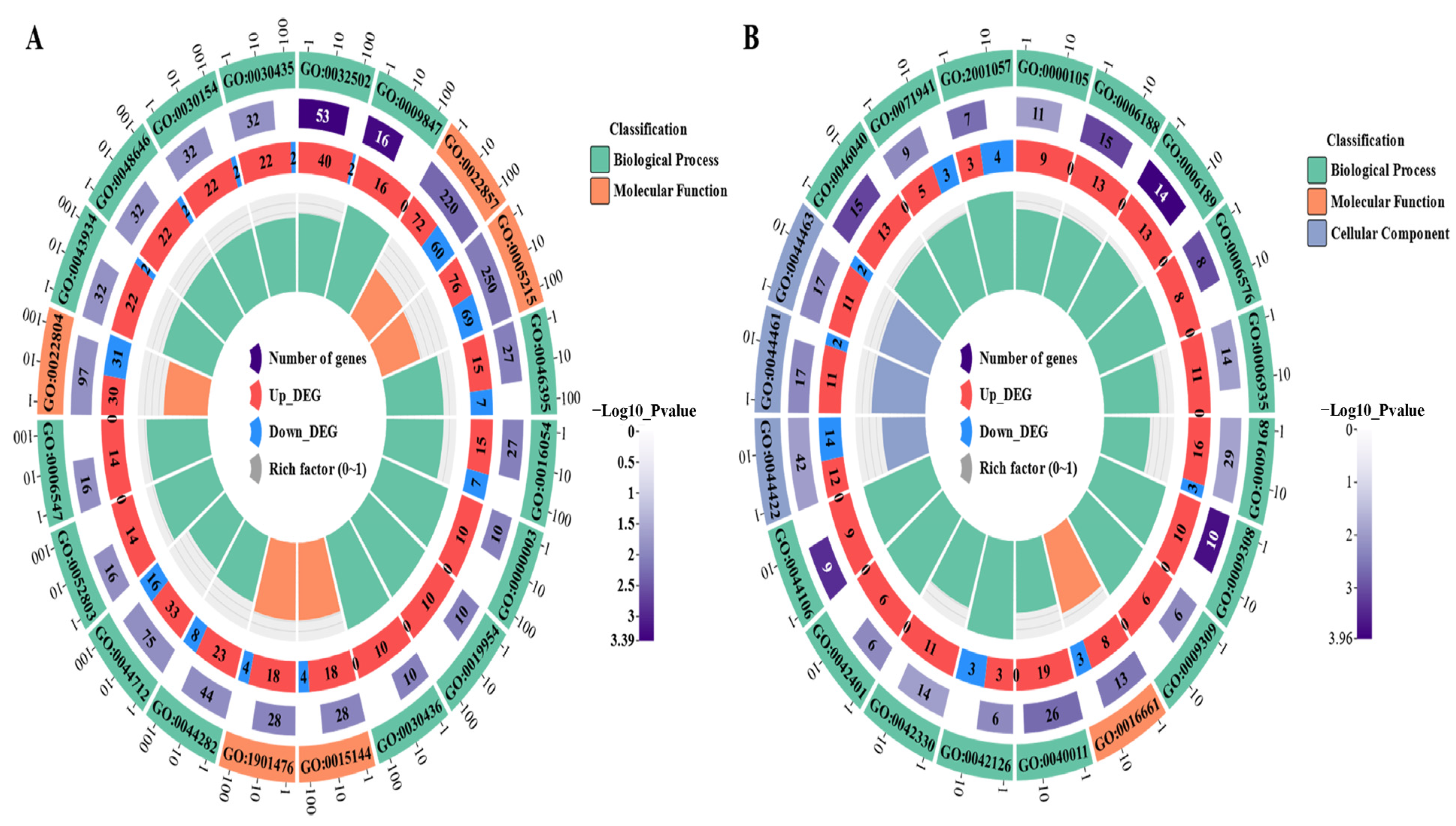
3.4. GO Functional Enrichment and KEGG Pathway Enrichment
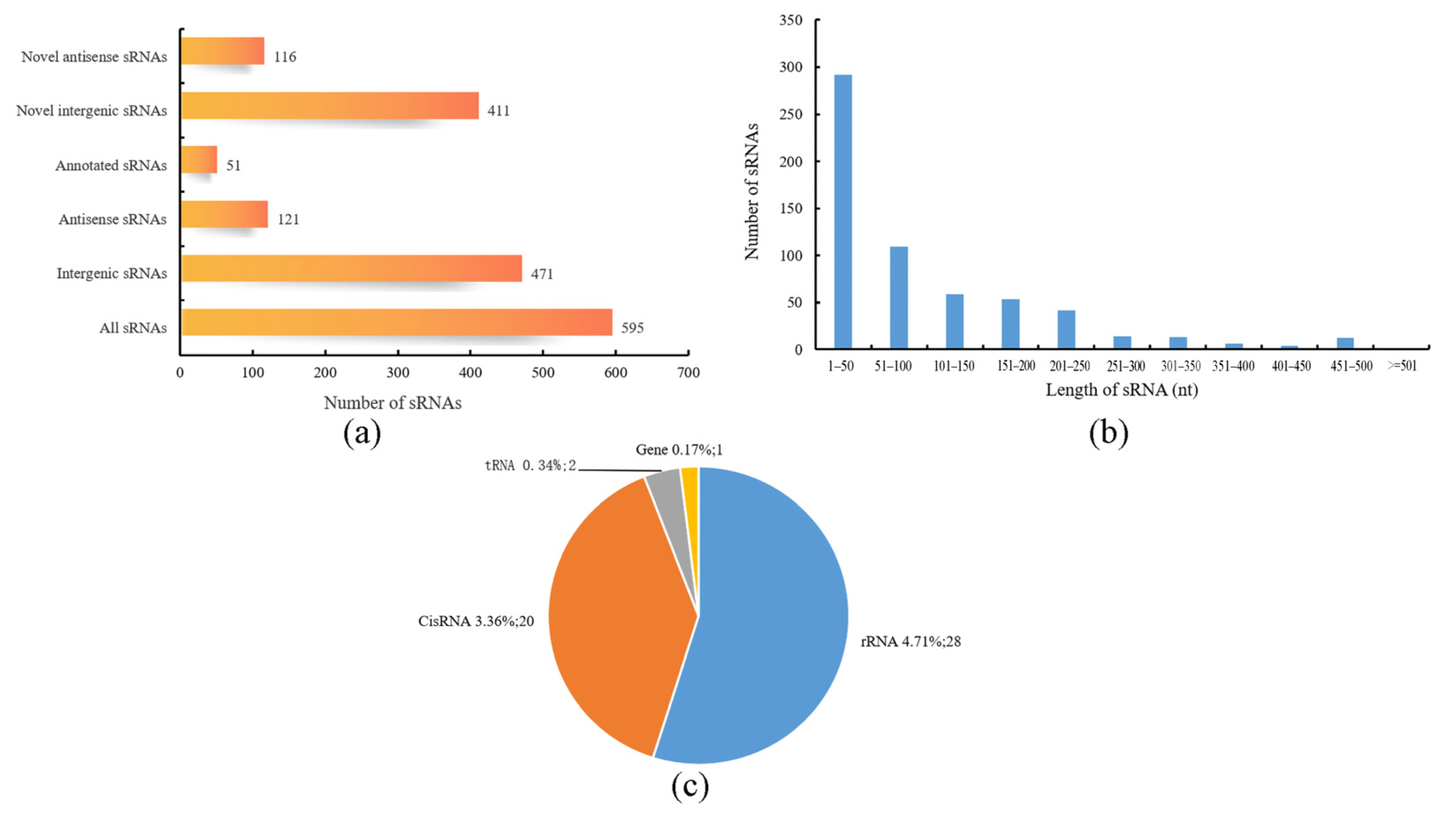
3.5. sRNA Analysis of B. subtilis BS-G1 under Different Glucose Concentrations
3.6. Annotation of Significantly Differentially Expressed sRNAs Target Genes
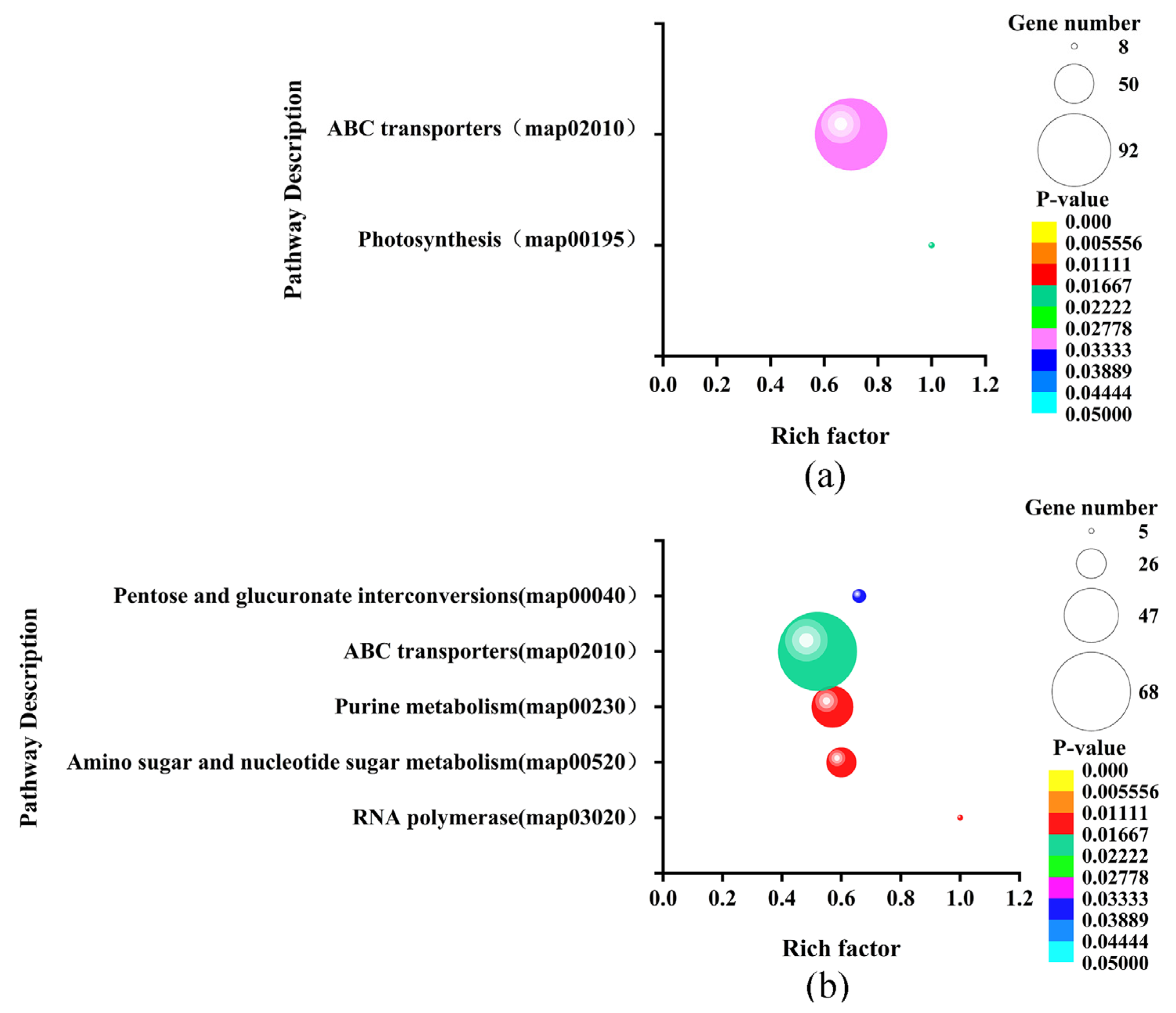
3.7. Enrichment Analysis of sRNA Target Genes
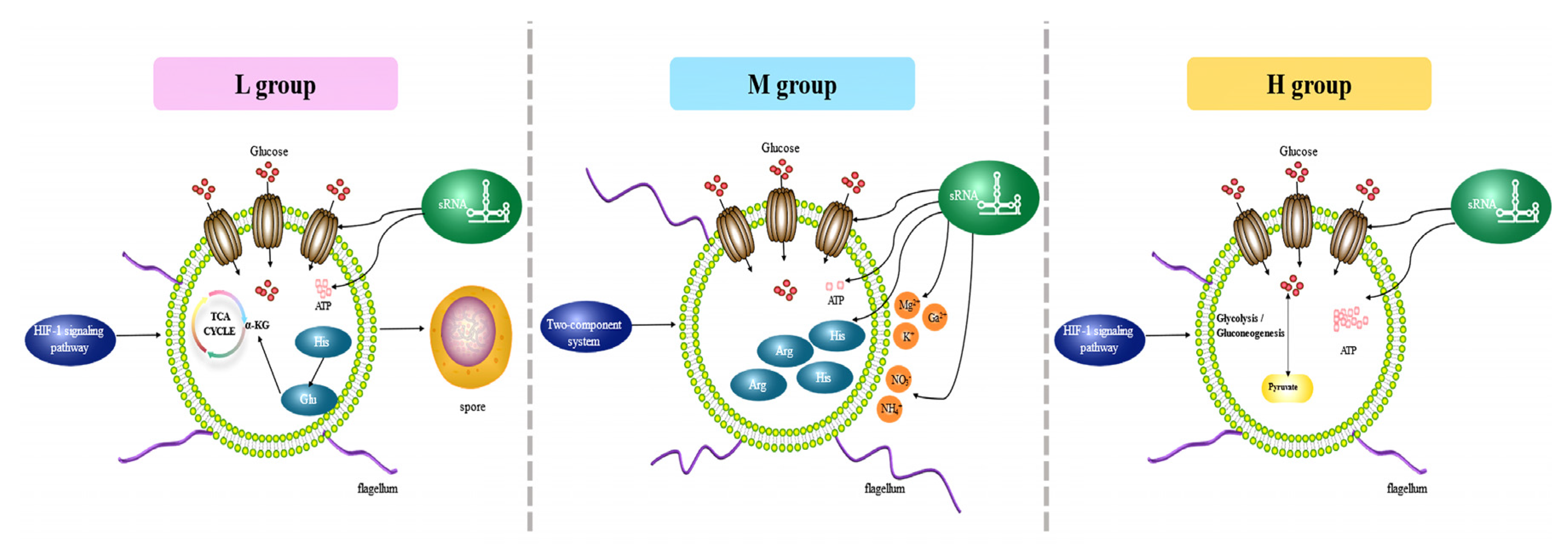
3.8. Adaptation Patterns and Key Genes in a Low-Glucose Environment
3.9. Critical Genes for Low-Sugar Adaptation
3.10. qRT-PCR Validation of DEGs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Props, R.; Monsieurs, P.; Vandamme, P.; Leys, N.; Denef, V.J.; Boon, N. Gene Expansion and Positive Selection as Bacterial Adaptations to Oligotrophic Conditions. mSphere 2019, 4, e00011-19. [Google Scholar] [CrossRef] [PubMed]
- Chiriac, M.C.; Haber, M.; Salcher, M.M. Adaptive Genetic Traits in Pelagic Freshwater Microbes. Environ. Microbiol. 2023, 25, 606–641. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Neilson, J.W.; Kushwaha, P.; Maier, R.M.; Barberán, A. Life-History Strategies of Soil Microbial Communities in an Arid Ecosystem. ISME J. 2021, 15, 649–657. [Google Scholar] [CrossRef]
- Cao, J.; Jiao, Y.; Che, R.; Holden, N.M.; Zhang, X.; Biswas, A.; Feng, Q. The Effects of Grazer Exclosure Duration on Soil Microbial Communities on the Qinghai-Tibetan Plateau. Sci. Total Environ. 2022, 839, 156238. [Google Scholar] [CrossRef]
- Dong, C.; Wei, L.; Wang, J.; Lai, Q.; Huang, Z.; Shao, Z. Genome-Based Taxonomic Rearrangement of Oceanobacter-Related Bacteria Including the Description of Thalassolituus Hydrocarbonoclasticus Sp. Nov. and Thalassolituus Pacificus Sp. Nov. and Emended Description of the Genus Thalassolituus. Front. Microbiol. 2022, 13, 1051202. [Google Scholar] [CrossRef] [PubMed]
- Ishida, Y.; Kadota, H. Growth Patterns and Substrate Requirements of Naturally Occurring Obligate Oligotrophs. Microb. Ecol. 1981, 7, 123–130. [Google Scholar] [CrossRef]
- de Araújo, H.L.; Martins, B.P.; Vicente, A.M.; Lorenzetti, A.P.R.; Koide, T.; Marques, M.V. Cold Regulation of Genes Encoding Ion Transport Systems in the Oligotrophic Bacterium Caulobacter Crescentus. Microbiol. Spectr. 2021, 9, e0071021. [Google Scholar] [CrossRef]
- Ishida, Y.; Imai, I.; Miyagaki, T.; Kadota, H. Growth and Uptake Kinetics of a Facultatively Oligotrophic Bacterium at Low Nutrient Concentrations. Microb. Ecol. 1982, 8, 23–32. [Google Scholar] [CrossRef]
- Ogura, M.; Sato, T.; Abe, K. Bacillus Subtilis YlxR, Which Is Involved in Glucose-Responsive Metabolic Changes, Regulates Expression of tsaD for Protein Quality Control of Pyruvate Dehydrogenase. Front. Microbiol. 2019, 10, 923. [Google Scholar] [CrossRef]
- Park, Y.S.; Kai, K.; Iijima, S.; Kobayashi, T. Enhanced Beta-Galactosidase Production by High Cell-Density Culture of Recombinant Bacillus Subtilis with Glucose Concentration Control. Biotechnol. Bioeng. 1992, 40, 686. [Google Scholar] [CrossRef]
- Dedysh, S.N. Cultivating Uncultured Bacteria from Northern Wetlands: Knowledge Gained and Remaining Gaps. Front. Microbiol. 2011, 2, 184. [Google Scholar] [CrossRef] [PubMed]
- Staley, J.T. Budding Bacteria of the Pasteuria-Blastobacter Group. Can. J. Microbiol. 1973, 19, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Strous, M.; Pelletier, E.; Mangenot, S.; Rattei, T.; Lehner, A.; Taylor, M.W.; Horn, M.; Daims, H.; Bartol-Mavel, D.; Wincker, P.; et al. Deciphering the Evolution and Metabolism of an Anammox Bacterium from a Community Genome. Nature 2006, 440, 790–794. [Google Scholar] [CrossRef]
- Kaboré, O.D.; Godreuil, S.; Drancourt, M. Planctomycetes as Host-Associated Bacteria: A Perspective That Holds Promise for Their Future Isolations, by Mimicking Their Native Environmental Niches in Clinical Microbiology Laboratories. Front. Cell Infect. Microbiol. 2020, 10, 519301. [Google Scholar] [CrossRef]
- Zhuang, H.; Wu, Z.; Xu, L.; Leu, S.-Y.; Lee, P.-H. Energy-Efficient Single-Stage Nitrite Shunt Denitrification with Saline Sewage through Concise Dissolved Oxygen (DO) Supply: Process Performance and Microbial Communities. Microorganisms 2020, 8, 919. [Google Scholar] [CrossRef] [PubMed]
- Koza, N.A.; Adedayo, A.A.; Babalola, O.O.; Kappo, A.P. Microorganisms in Plant Growth and Development: Roles in Abiotic Stress Tolerance and Secondary Metabolites Secretion. Microorganisms 2022, 10, 1528. [Google Scholar] [CrossRef]
- Carreón-Rodríguez, O.E.; Gosset, G.; Escalante, A.; Bolívar, F. Glucose Transport in Escherichia Coli: From Basics to Transport Engineering. Microorganisms 2023, 11, 1588. [Google Scholar] [CrossRef]
- Wen, J.; Zhao, X.; Si, F.; Qi, G. Surfactin, a Quorum Sensing Signal Molecule, Globally Affects the Carbon Metabolism in Bacillus Amyloliquefaciens. Metab. Eng. Commun. 2021, 12, e00174. [Google Scholar] [CrossRef]
- Chamoli, S.; Kumar, P.; Navani, N.K.; Verma, A.K. Secretory Expression, Characterization and Docking Study of Glucose-Tolerant β-Glucosidase from B. Subtilis. Int. J. Biol. Macromol. 2016, 85, 425–433. [Google Scholar] [CrossRef]
- Pan, S.; Chen, G.; Wu, R.; Cao, X.; Liang, Z. Non-Sterile Submerged Fermentation of Fibrinolytic Enzyme by Marine Bacillus Subtilis Harboring Antibacterial Activity with Starvation Strategy. Front. Microbiol. 2019, 10, 1025. [Google Scholar] [CrossRef]
- Norris, N.; Levine, N.M.; Fernandez, V.I.; Stocker, R. Mechanistic Model of Nutrient Uptake Explains Dichotomy between Marine Oligotrophic and Copiotrophic Bacteria. PLoS Comput. Biol. 2021, 17, e1009023. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hammes, F.; Boon, N.; Egli, T. Quantification of the Filterability of Freshwater Bacteria through 0.45, 0.22, and 0.1 Microm Pore Size Filters and Shape-Dependent Enrichment of Filterable Bacterial Communities. Environ. Sci. Technol. 2007, 41, 7080–7086. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Cañizares, C.; Prell, J.; Pini, F.; Rutten, P.; Kraxner, K.; Wynands, B.; Karunakaran, R.; Poole, P.S. Global Control of Bacterial Nitrogen and Carbon Metabolism by a PTSNtr-Regulated Switch. Proc. Natl. Acad. Sci. USA 2020, 117, 10234–10245. [Google Scholar] [CrossRef] [PubMed]
- Jeckelmann, J.M.; Erni, B. Carbohydrate Transport by Group Translocation: The Bacterial Phosphoenolpyruvate: Sugar Phosphotransferase System. Subcell. Biochem. 2019, 92, 223–274. [Google Scholar] [CrossRef]
- Fragoso-Jiménez, J.C.; Baert, J.; Nguyen, T.M.; Liu, W.; Sassi, H.; Goormaghtigh, F.; Van Melderen, L.; Gaytán, P.; Hernández-Chávez, G.; Martinez, A.; et al. Growth-Dependent Recombinant Product Formation Kinetics Can Be Reproduced through Engineering of Glucose Transport and Is Prone to Phenotypic Heterogeneity. Microb. Cell Fact. 2019, 18, 26. [Google Scholar] [CrossRef]
- Morabbi Heravi, K.; Altenbuchner, J. Cross Talk among Transporters of the Phosphoenolpyruvate-Dependent Phosphotransferase System in Bacillus Subtilis. J. Bacteriol. 2018, 200, e00213-18. [Google Scholar] [CrossRef]
- Storz, G.; Vogel, J.; Wassarman, K.M. Regulation by Small RNAs in Bacteria: Expanding Frontiers. Mol. Cell 2011, 43, 880–891. [Google Scholar] [CrossRef]
- Wagner, E.G.H.; Romby, P. Small RNAs in Bacteria and Archaea: Who They Are, What They Do, and How They Do It. Adv. Genet. 2015, 90, 133–208. [Google Scholar] [CrossRef]
- Papenfort, K.; Melamed, S. Small RNAs, Large Networks: Posttranscriptional Regulons in Gram-Negative Bacteria. Annu. Rev. Microbiol. 2023, 77, 36944261. [Google Scholar] [CrossRef]
- Taneja, S.; Dutta, T. On a Stake-out: Mycobacterial Small RNA Identification and Regulation. Noncoding RNA Res. 2019, 4, 86–95. [Google Scholar] [CrossRef]
- Charbonnier, M.; González-Espinoza, G.; Kehl-Fie, T.E.; Lalaouna, D. Battle for Metals: Regulatory RNAs at the Front Line. Front. Cell Infect. Microbiol. 2022, 12, 952948. [Google Scholar] [CrossRef] [PubMed]
- Parise, M.T.D.; Parise, D.; Aburjaile, F.F.; Pinto Gomide, A.C.; Kato, R.B.; Raden, M.; Backofen, R.; de Carvalho Azevedo, V.A.; Baumbach, J. An Integrated Database of Small RNAs and Their Interplay with Transcriptional Gene Regulatory Networks in Corynebacteria. Front. Microbiol. 2021, 12, 656435. [Google Scholar] [CrossRef] [PubMed]
- Gottesman, S.; Storz, G. Bacterial Small RNA Regulators: Versatile Roles and Rapidly Evolving Variations. Cold Spring Harb. Perspect. Biol. 2011, 3, a003798. [Google Scholar] [CrossRef]
- Fröhlich, K.S.; Papenfort, K. Regulation Outside the Box: New Mechanisms for Small RNAs. Mol. Microbiol. 2020, 114, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Romilly, C.; Deindl, S.; Wagner, E.G.H. The Ribosomal Protein S1-Dependent Standby Site in tisB mRNA Consists of a Single-Stranded Region and a 5′ Structure Element. Proc. Natl. Acad. Sci. USA 2019, 116, 15901–15906. [Google Scholar] [CrossRef] [PubMed]
- Melson, E.M.; Kendall, M.M. The sRNA DicF Integrates Oxygen Sensing to Enhance Enterohemorrhagic Escherichia Coli Virulence via Distinctive RNA Control Mechanisms. Proc. Natl. Acad. Sci. USA 2019, 116, 14210–14215. [Google Scholar] [CrossRef]
- Balasubramanian, D.; Vanderpool, C.K. New Developments in Post-Transcriptional Regulation of Operons by Small RNAs. RNA Biol. 2013, 10, 337–341. [Google Scholar] [CrossRef][Green Version]
- Sheehan, L.M.; Budnick, J.A.; Fyffe-Blair, J.; King, K.A.; Settlage, R.E.; Caswell, C.C. The Endoribonuclease RNase E Coordinates Expression of mRNAs and Small Regulatory RNAs and Is Critical for the Virulence of Brucella Abortus. J. Bacteriol. 2020, 202, e00240-20. [Google Scholar] [CrossRef]
- Quendera, A.P.; Seixas, A.F.; dos Santos, R.F.; Santos, I.; Silva, J.P.N.; Arraiano, C.M.; Andrade, J.M. RNA-Binding Proteins Driving the Regulatory Activity of Small Non-Coding RNAs in Bacteria. Front. Mol. Biosci. 2020, 7, 78. [Google Scholar] [CrossRef]
- McQuail, J.; Carpousis, A.J.; Wigneshweraraj, S. The Association between Hfq and RNase E in Long-Term Nitrogen-Starved Escherichia coli. Mol. Microbiol. 2022, 117, 54–66. [Google Scholar] [CrossRef]
- De Lay, N.; Schu, D.J.; Gottesman, S. Bacterial Small RNA-Based Negative Regulation: Hfq and Its Accomplices. J. Biol. Chem. 2013, 288, 7996–8003. [Google Scholar] [CrossRef] [PubMed]
- Murina, V.N.; Nikulin, A.D. Bacterial Small Regulatory RNAs and Hfq Protein. Biochemistry 2015, 80, 1647–1654. [Google Scholar] [CrossRef] [PubMed]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved Seed Pairing, Often Flanked by Adenosines, Indicates That Thousands of Human Genes Are microRNA Targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Melamed, S.; Peer, A.; Faigenbaum-Romm, R.; Gatt, Y.E.; Reiss, N.; Bar, A.; Altuvia, Y.; Argaman, L.; Margalit, H. Global Mapping of Small RNA-Target Interactions in Bacteria. Mol. Cell 2016, 63, 884–897. [Google Scholar] [CrossRef] [PubMed]
- Bobrovskyy, M.; Vanderpool, C.K. Regulation of Bacterial Metabolism by Small RNAs Using Diverse Mechanisms. Annu. Rev. Genet. 2013, 47, 209–232. [Google Scholar] [CrossRef]
- Roth-Rosenberg, D.; Aharonovich, D.; Luzzatto-Knaan, T.; Vogts, A.; Zoccarato, L.; Eigemann, F.; Nago, N.; Grossart, H.P.; Voss, M.; Sher, D. Prochlorococcus Cells Rely on Microbial Interactions Rather than on Chlorotic Resting Stages to Survive Long-Term Nutrient Starvation. mBio 2020, 11, e01846-20. [Google Scholar] [CrossRef]
- Szul, M.J.; Dearth, S.P.; Campagna, S.R.; Zinser, E.R. Carbon Fate and Flux in Prochlorococcus under Nitrogen Limitation. mSystems 2019, 4, e00254-18. [Google Scholar] [CrossRef]
- Santander, R.D.; Oliver, J.D.; Biosca, E.G. Cellular, Physiological, and Molecular Adaptive Responses of Erwinia Amylovora to Starvation. FEMS Microbiol. Ecol. 2014, 88, 258–271. [Google Scholar] [CrossRef]
- Read, R.W.; Berube, P.M.; Biller, S.J.; Neveux, I.; Cubillos-Ruiz, A.; Chisholm, S.W.; Grzymski, J.J. Nitrogen Cost Minimization Is Promoted by Structural Changes in the Transcriptome of N-Deprived Prochlorococcus Cells. ISME J. 2017, 11, 2267–2278. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Raihan, M.R.H.; Nowroz, F.; Fujita, M. Insight into the Mechanism of Salt-Induced Oxidative Stress Tolerance in Soybean by the Application of Bacillus Subtilis: Coordinated Actions of Osmoregulation, Ion Homeostasis, Antioxidant Defense, and Methylglyoxal Detoxification. Antioxidants 2022, 11, 1856. [Google Scholar] [CrossRef]
- Tibocha-Bonilla, J.D.; Zuñiga, C.; Lekbua, A.; Lloyd, C.; Rychel, K.; Short, K.; Zengler, K. Predicting Stress Response and Improved Protein Overproduction in Bacillus Subtilis. NPJ Syst. Biol. Appl. 2022, 8, 50. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Shang, C.; Zhang, H.; Sun, C.; Zhang, G.; Liu, L.; Li, C.; Li, A.; Du, P. Effects of Alkali Stress on the Growth and Menaquinone-7 Metabolism of Bacillus Subtilis Natto. Front. Microbiol. 2022, 13, 899802. [Google Scholar] [CrossRef] [PubMed]
- Gray, D.A.; Dugar, G.; Gamba, P.; Strahl, H.; Jonker, M.J.; Hamoen, L.W. Extreme Slow Growth as Alternative Strategy to Survive Deep Starvation in Bacteria. Nat. Commun. 2019, 10, 890. [Google Scholar] [CrossRef]
- Ford, B.A.; Sullivan, G.J.; Moore, L.; Varkey, D.; Zhu, H.; Ostrowski, M.; Mabbutt, B.C.; Paulsen, I.T.; Shah, B.S. Functional Characterisation of Substrate-Binding Proteins to Address Nutrient Uptake in Marine Picocyanobacteria. Biochem. Soc. Trans. 2021, 49, 2465–2481. [Google Scholar] [CrossRef] [PubMed]
- Lauro, F.M.; McDougald, D.; Thomas, T.; Williams, T.J.; Egan, S.; Rice, S.; DeMaere, M.Z.; Ting, L.; Ertan, H.; Johnson, J.; et al. The Genomic Basis of Trophic Strategy in Marine Bacteria. Proc. Natl. Acad. Sci. USA 2009, 106, 15527–15533. [Google Scholar] [CrossRef]
- Schwalbach, M.S.; Tripp, H.J.; Steindler, L.; Smith, D.P.; Giovannoni, S.J. The Presence of the Glycolysis Operon in SAR11 Genomes Is Positively Correlated with Ocean Productivity. Environ. Microbiol. 2010, 12, 490–500. [Google Scholar] [CrossRef]
- Tam, R.; Saier, M.H. Structural, Functional, and Evolutionary Relationships among Extracellular Solute-Binding Receptors of Bacteria. Microbiol. Rev. 1993, 57, 320–346. [Google Scholar] [CrossRef]
- Jaskulak, M.; Grobelak, A.; Vandenbulcke, F. Effects of Sewage Sludge Supplementation on Heavy Metal Accumulation and the Expression of ABC Transporters in Sinapis alba L. during Assisted Phytoremediation of Contaminated Sites. Ecotoxicol. Environ. Saf. 2020, 197, 110606. [Google Scholar] [CrossRef]
- Giovannoni, S.J.; Tripp, H.J.; Givan, S.; Podar, M.; Vergin, K.L.; Baptista, D.; Bibbs, L.; Eads, J.; Richardson, T.H.; Noordewier, M.; et al. Genome Streamlining in a Cosmopolitan Oceanic Bacterium. Science 2005, 309, 1242–1245. [Google Scholar] [CrossRef]
- Williams, T.J.; Ertan, H.; Ting, L.; Cavicchioli, R. Carbon and Nitrogen Substrate Utilization in the Marine Bacterium Sphingopyxis Alaskensis Strain RB2256. ISME J. 2009, 3, 1036–1052. [Google Scholar] [CrossRef]
- Orsi, W.D.; Smith, J.M.; Liu, S.; Liu, Z.; Sakamoto, C.M.; Wilken, S.; Poirier, C.; Richards, T.A.; Keeling, P.J.; Worden, A.Z.; et al. Diverse, Uncultivated Bacteria and Archaea Underlying the Cycling of Dissolved Protein in the Ocean. ISME J. 2016, 10, 2158–2173. [Google Scholar] [CrossRef] [PubMed]
- Lofton, M.E.; Brentrup, J.A.; Beck, W.S.; Zwart, J.A.; Bhattacharya, R.; Brighenti, L.S.; Burnet, S.H.; McCullough, I.M.; Steele, B.G.; Carey, C.C.; et al. Using Near-Term Forecasts and Uncertainty Partitioning to Inform Prediction of Oligotrophic Lake Cyanobacterial Density. Ecol. Appl. 2022, 32, e2590. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Allen, J.; Lu, J. Community Structures of Phytoplankton with Emphasis on Toxic Cyanobacteria in an Ohio Inland Lake during Bloom Season. J. Water Resour. Prot. 2017, 9, 11. [Google Scholar] [CrossRef] [PubMed]
- Rinta-Kanto, J.M.; Konopko, E.A.; Debruyn, J.M.; Bourbonniere, R.A.; Boyer, G.L.; Wilhelm, S.W. Lake Erie Microcystis: Relationship between Microcystin Production, Dynamics of Genotypes and Environmental Parameters in a Large Lake. Harmful Algae 2009, 8, 665–673. [Google Scholar] [CrossRef]
- Guo, S.; Wang, Y.; Huang, J.; Dong, J.; Zhang, J. Decoupling and Decomposition Analysis of Land Natural Capital Utilization and Economic Growth: A Case Study in Ningxia Hui Autonomous Region, China. Int. J. Environ. Res. Public Health 2021, 18, 646. [Google Scholar] [CrossRef]
- Peng, L.; Zhao, K.; Chen, S.; Ren, Z.; Wei, H.; Wan, C. Whole Genome and Acid Stress Comparative Transcriptome Analysis of Lactiplantibacillus Plantarum ZDY2013. Arch. Microbiol. 2021, 203, 2795–2807. [Google Scholar] [CrossRef]
- Zhang, L.; Song, D.; Wu, Z. Transcriptome Analysis of Cyclocarya Paliurus Flavonoids Regulation of Differently Expressed Genes in Enterococcus Faecalis under Low pH Stress. Arch. Microbiol. 2021, 203, 2147–2155. [Google Scholar] [CrossRef]
- Petrov, K.; Arsov, A.; Petrova, P. Butanol Tolerance of Lactiplantibacillus Plantarum: A Transcriptome Study. Genes 2021, 12, 181. [Google Scholar] [CrossRef]
- Cheng, C.; Han, X.; Xu, J.; Sun, J.; Li, K.; Han, Y.; Chen, M.; Song, H. YjbH Mediates the Oxidative Stress Response and Infection by Regulating SpxA1 and the Phosphoenolpyruvate-Carbohydrate Phosphotransferase System (PTS) in Listeria Monocytogenes. Gut Microbes 2021, 13, 1884517. [Google Scholar] [CrossRef]
- Vasylkivska, M.; Jureckova, K.; Branska, B.; Sedlar, K.; Kolek, J.; Provaznik, I.; Patakova, P. Transcriptional Analysis of Amino Acid, Metal Ion, Vitamin and Carbohydrate Uptake in Butanol-Producing Clostridium Beijerinckii NRRL B-598. PLoS ONE 2019, 14, e0224560. [Google Scholar] [CrossRef]
- Stautz, J.; Hellmich, Y.; Fuss, M.F.; Silberberg, J.M.; Devlin, J.R.; Stockbridge, R.B.; Hänelt, I. Molecular Mechanisms for Bacterial Potassium Homeostasis. J. Mol. Biol. 2021, 433, 166968. [Google Scholar] [CrossRef]
- Stülke, J.; Krüger, L. Cyclic Di-AMP Signaling in Bacteria. Annu. Rev. Microbiol. 2020, 74, 159–179. [Google Scholar] [CrossRef]
- Fang, H.; Qin, X.Y.; Zhang, K.D.; Nie, Y.; Wu, X.L. Role of the Group 2 Mrp Sodium/Proton Antiporter in Rapid Response to High Alkaline Shock in the Alkaline- and Salt-Tolerant Dietzia Sp. DQ12-45-1b. Appl. Microbiol. Biotechnol. 2018, 102, 3765–3777. [Google Scholar] [CrossRef] [PubMed]
- Dürre, P. Physiology and Sporulation in Clostridium. Microbiol. Spectr. 2014, 2, 4. [Google Scholar] [CrossRef] [PubMed]
- Eisenstadt, E. Potassium Content during Growth and Sporulation in Bacillus Subtilis. J. Bacteriol. 1972, 112, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Vohradsky, J.; Schwarz, M.; Ramaniuk, O.; Ruiz-Larrabeiti, O.; Vaňková Hausnerová, V.; Šanderová, H.; Krásný, L. Kinetic Modeling and Meta-Analysis of the Bacillus Subtilis SigB Regulon during Spore Germination and Outgrowth. Microorganisms 2021, 9, 112. [Google Scholar] [CrossRef]
- Stetsenko, A.; Guskov, A. Cation Permeability in CorA Family of Proteins. Sci. Rep. 2020, 10, 840. [Google Scholar] [CrossRef]
- Payandeh, J.; Pfoh, R.; Pai, E.F. The Structure and Regulation of Magnesium Selective Ion Channels. Biochim. Biophys. Acta 2013, 1828, 2778–2792. [Google Scholar] [CrossRef]
- Pohland, A.C.; Schneider, D. Mg2+ Homeostasis and Transport in Cyanobacteria—At the Crossroads of Bacterial and Chloroplast Mg2+ Import. Biol. Chem. 2019, 400, 1289–1301. [Google Scholar] [CrossRef]
- Trachsel, E.; Redder, P.; Linder, P.; Armitano, J. Genetic Screens Reveal Novel Major and Minor Players in Magnesium Homeostasis of Staphylococcus Aureus. PLoS Genet. 2019, 15, e1008336. [Google Scholar] [CrossRef]
- Saha, J.; Dey, S.; Pal, A. Whole Genome Sequencing and Comparative Genomic Analyses of Pseudomonas Aeruginosa Strain Isolated from Arable Soil Reveal Novel Insights into Heavy Metal Resistance and Codon Biology. Curr. Genet. 2022, 68, 481–503. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; He, Q. PfsR Is a Key Regulator of Iron Homeostasis in Synechocystis PCC 6803. PLoS ONE 2014, 9, e101743. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhang, T.; Cao, Y.; Wang, L.; Chen, W. New Insights into the Function of the Proteins IsiC and IsiD from Synechocystis Sp. PCC 6803 under Iron Limitation. Appl. Microbiol. Biotechnol. 2021, 105, 4693–4707. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.W.; Kim, D.; Latif, H.; O’Brien, E.J.; Szubin, R.; Palsson, B.O. Deciphering Fur Transcriptional Regulatory Network Highlights Its Complex Role beyond Iron Metabolism in Escherichia coli. Nat. Commun. 2014, 5, 4910. [Google Scholar] [CrossRef]
- Plante, S.; Labbé, S. Spore Germination Requires Ferrichrome Biosynthesis and the Siderophore Transporter Str1 in Schizosaccharomyces Pombe. Genetics 2019, 211, 893–911. [Google Scholar] [CrossRef]
- Pivato, M.; Ballottari, M. Chlamydomonas Reinhardtii Cellular Compartments and Their Contribution to Intracellular Calcium Signalling. J. Exp. Bot. 2021, 72, 5312–5335. [Google Scholar] [CrossRef]
- Cao, R.; Qin, P.; Li, W.; Shang, C.; Chai, Y.; Jin, D.; Chen, A. Hydrogen Sulfide and Calcium Effects on Cadmium Removal and Resistance in the White-Rot Fungus Phanerochaete Chrysosporium. Appl. Microbiol. Biotechnol. 2021, 105, 6451–6462. [Google Scholar] [CrossRef]
- Wan, Y.; Wang, M.; Chan, E.W.C.; Chen, S. Membrane Transporters of the Major Facilitator Superfamily Are Essential for Long-Term Maintenance of Phenotypic Tolerance to Multiple Antibiotics in E. coli. Microbiol. Spectr. 2021, 9, e0184621. [Google Scholar] [CrossRef]
- Wani, A.K.; Akhtar, N.; Sher, F.; Navarrete, A.A.; Américo-Pinheiro, J.H.P. Microbial Adaptation to Different Environmental Conditions: Molecular Perspective of Evolved Genetic and Cellular Systems. Arch. Microbiol. 2022, 204, 144. [Google Scholar] [CrossRef]
- Coloma-Rivero, R.F.; Flores-Concha, M.; Molina, R.E.; Soto-Shara, R.; Cartes, Á.; Oñate, Á.A. Brucella and Its Hidden Flagellar System. Microorganisms 2021, 10, 83. [Google Scholar] [CrossRef]
- Zhou, M.; Liu, Z.; Wang, J.; Zhao, Y.; Hu, B. Sphingomonas Relies on Chemotaxis to Degrade Polycyclic Aromatic Hydrocarbons and Maintain Dominance in Coking Sites. Microorganisms 2022, 10, 1109. [Google Scholar] [CrossRef] [PubMed]
- Solar Venero, E.C.; Ricardi, M.M.; Gomez-Lozano, M.; Molin, S.; Tribelli, P.M.; López, N.I. Oxidative Stress under Low Oxygen Conditions Triggers Hyperflagellation and Motility in the Antarctic Bacterium Pseudomonas Extremaustralis. Extremophiles 2019, 23, 587–597. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, J.; Gayathri, M. Transcriptome Based Identification of Silver Stress Responsive sRNAs from Bacillus Cereus ATCC14579. Bioinformation 2019, 15, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Amin, S.V.; Roberts, J.T.; Patterson, D.G.; Coley, A.B.; Allred, J.A.; Denner, J.M.; Johnson, J.P.; Mullen, G.E.; O’Neal, T.K.; Smith, J.T.; et al. Novel Small RNA (sRNA) Landscape of the Starvation-Stress Response Transcriptome of Salmonella Enterica Serovar Typhimurium. RNA Biol. 2016, 13, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Landt, S.G.; Lesley, J.A.; Britos, L.; Shapiro, L. CrfA, a Small Noncoding RNA Regulator of Adaptation to Carbon Starvation in Caulobacter Crescentus. J. Bacteriol. 2010, 192, 4763–4775. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Babitzke, P.; Lai, Y.-J.; Renda, A.J.; Romeo, T. Posttranscription Initiation Control of Gene Expression Mediated by Bacterial RNA-Binding Proteins. Annu. Rev. Microbiol. 2019, 73, 43–67. [Google Scholar] [CrossRef]
- Chioccioli, S.; Del Duca, S.; Vassallo, A.; Castronovo, L.M.; Fani, R. Exploring the Role of the Histidine Biosynthetic hisF Gene in Cellular Metabolism and in the Evolution of (Ancestral) Genes: From LUCA to the Extant (Micro)Organisms. Microbiol. Res. 2020, 240, 126555. [Google Scholar] [CrossRef]
- Fani, R.; Brilli, M.; Fondi, M.; Lió, P. The Role of Gene Fusions in the Evolution of Metabolic Pathways: The Histidine Biosynthesis Case. BMC Evol. Biol. 2007, 7 (Suppl. S2), S4. [Google Scholar] [CrossRef]
- Martínez-Guitián, M.; Vázquez-Ucha, J.C.; Álvarez-Fraga, L.; Conde-Pérez, K.; Lasarte-Monterrubio, C.; Vallejo, J.A.; Bou, G.; Poza, M.; Beceiro, A. Involvement of HisF in the Persistence of Acinetobacter Baumannii During a Pneumonia Infection. Front. Cell Infect. Microbiol. 2019, 9, 310. [Google Scholar] [CrossRef]
- Kuba, M.; Neha, N.; De Souza, D.P.; Dayalan, S.; Newson, J.P.M.; Tull, D.; McConville, M.J.; Sansom, F.M.; Newton, H.J. Coxiella Burnetii Utilizes Both Glutamate and Glucose during Infection with Glucose Uptake Mediated by Multiple Transporters. Biochem. J. 2019, 476, 2851–2867. [Google Scholar] [CrossRef]
- Byer, T.; Wang, J.; Zhang, M.G.; Vather, N.; Blachman, A.; Visser, B.; Liu, J.M. MtlR Negatively Regulates Mannitol Utilization by Vibrio Cholerae. Microbiology 2017, 163, 1902–1911. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Jiang, T.; Zou, L.; Ouyang, S.; Zhou, J.; Lin, X.; He, Q.; Wang, L.; Yu, B.; Xu, H.; et al. Simultaneous Consumption of Cellobiose and Xylose by Bacillus Coagulans to Circumvent Glucose Repression and Identification of Its Cellobiose-Assimilating Operons. Biotechnol. Biofuels 2018, 11, 320. [Google Scholar] [CrossRef] [PubMed]
- Aboulwafa, M.; Zhang, Z.; Saier, M.H. Protein-Protein Interactions in the Cytoplasmic Membrane of Escherichia coli: Influence of the Overexpression of Diverse Transporter-Encoding Genes on the Activities of PTS Sugar Uptake Systems. Microb. Physiol. 2020, 30, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Li, F.; Zuo, X.; Chen, J.; Qin, P.; Wang, C.; Xu, J.; Yang, D.; Xing, B.; Liu, Y.; et al. Reversible Domain Closure Modulates GlnBP Ligand Binding Affinity. PLoS ONE 2022, 17, e0263102. [Google Scholar] [CrossRef]
- Zhang, M.G.; Liu, J.M. Transcription of Cis Antisense Small RNA MtlS in Vibrio Cholerae Is Regulated by Transcription of Its Target Gene, mtlA. J. Bacteriol. 2019, 201, e00178-19. [Google Scholar] [CrossRef]
- Poorinmohammad, N.; Hamedi, J.; Masoudi-Nejad, A. Genome-Scale Exploration of Transcriptional Regulation in the Nisin Z Producer Lactococcus Lactis Subsp. Lactis IO-1. Sci. Rep. 2020, 10, 3787. [Google Scholar] [CrossRef]
- Jang, H.; Kim, S.T.; Sang, M.K. Suppressive Effect of Bioactive Extracts of Bacillus Sp. H8-1 and Bacillus Sp. K203 on Tomato Wilt Caused by Clavibacter Michiganensis Subsp. Michiganensis. Microorganisms 2022, 10, 403. [Google Scholar] [CrossRef]
- Cao, T.N.; Joyet, P.; Aké, F.M.D.; Milohanic, E.; Deutscher, J. Studies of the Listeria Monocytogenes Cellobiose Transport Components and Their Impact on Virulence Gene Repression. J. Mol. Microbiol. Biotechnol. 2019, 29, 10–26. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Key Gene ID | Description | Gene Name | Log2FC (LvsH) |
|---|---|---|---|
| gene2652 | amino acid ABC transporter ATPase | / | 7.448 |
| gene2985 | ABC transporter permease | / | 10.238 |
| gene3860 | S | celB | 9.383 |
| gene0435 | PTS mannitol transporter subunit IIB | mtlA | 7.98 |
| gene3876 | oligo-beta-mannoside permease IIC protein | celB | 7.362 |
| gene2653 | glutamine ABC transporter substrate-binding protein | / | 7.496 |
| gene2655 | glutamine ABC transporter permease | / | 5.051 |
| sRNA_id | Log2FC (LvsH) | Target Gene ID |
|---|---|---|
| sRNA0200 | 11.71890433 | gene0435; gene3876 |
| sRNA0113 | 11.533039 | gene3860 |
| sRNA0084 | 11.40169888 | gene3876 |
| sRNA0085 | 11.40169888 | gene3876 |
| sRNA0148 | 11.40169888 | gene3876 |
| sRNA0207 | 10.90597229 | gene0435 |
| sRNA0205 | 9.776851885 | gene3876 |
| sRNA0366 | 8.533373277 | gene0435 |
| sRNA0336 | 6.681651624 | gene0435 |
| sRNA0362 | 6.434679786 | gene0435; gene2655 |
| sRNA0116 | 6.373911204 | gene0435 |
| sRNA0360 | 6.357940855 | gene2653 |
| sRNA0542 | 6.212579528 | gene2652 |
| sRNA0557 | 6.026353075 | gene2985 |
| sRNA0199 | 5.159987109 | gene2655 |
| sRNA0504 | 5.048301125 | gene2985; gene0435 |
| sRNA0015 | 4.964673587 | gene3876 |
| sRNA0533 | 4.896095911 | gene0435 |
| sRNA0419 | 4.801798083 | gene2985; gene3876 |
| sRNA0239 | 4.618899947 | gene3876 |
| sRNA0249 | 4.578804747 | gene0435 |
| sRNA0495 | 3.975578933 | gene0435 |
| sRNA0411 | 3.854669068 | gene3876 |
| sRNA0509 | 3.764795398 | gene2985 |
| sRNA0221 | 3.520639327 | gene0435 |
| sRNA0414 | 3.022561069 | gene3876 |
| sRNA0481 | 2.967066947 | gene0435; gene2985 |
| sRNA0417 | 1.934699587 | gene3876 |
| sRNA0364 | 1.373518955 | gene3876 |
| sRNA0011 | 1.103524326 | gene3876 |
| sRNA0157 | −1.52566018 | gene3876 |
| sRNA0404 | −1.81557978 | gene0435 |
| sRNA0375 | −2.395421117 | gene2985; gene0435 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Wang, C.; Su, J. Understanding the Effect of Different Glucose Concentrations in the Oligotrophic Bacterium Bacillus subtilis BS-G1 through Transcriptomics Analysis. Microorganisms 2023, 11, 2401. https://doi.org/10.3390/microorganisms11102401
Chen L, Wang C, Su J. Understanding the Effect of Different Glucose Concentrations in the Oligotrophic Bacterium Bacillus subtilis BS-G1 through Transcriptomics Analysis. Microorganisms. 2023; 11(10):2401. https://doi.org/10.3390/microorganisms11102401
Chicago/Turabian StyleChen, Liping, Chenglong Wang, and Jianyu Su. 2023. "Understanding the Effect of Different Glucose Concentrations in the Oligotrophic Bacterium Bacillus subtilis BS-G1 through Transcriptomics Analysis" Microorganisms 11, no. 10: 2401. https://doi.org/10.3390/microorganisms11102401
APA StyleChen, L., Wang, C., & Su, J. (2023). Understanding the Effect of Different Glucose Concentrations in the Oligotrophic Bacterium Bacillus subtilis BS-G1 through Transcriptomics Analysis. Microorganisms, 11(10), 2401. https://doi.org/10.3390/microorganisms11102401