
Probiotics and Their Bioproducts: A Promising Approach for Targeting Methicillin-Resistant Staphylococcus aureus and Vancomycin-Resistant Enterococcus



Abstract
:1. Introduction
2. Conventional Treatment of Multi-Drug Resistant Infections
2.1. Conventional Treatment of Methicillin-Resistant Staphylococcus aureus and Vancomycin-Resistant Enterococcal Infections
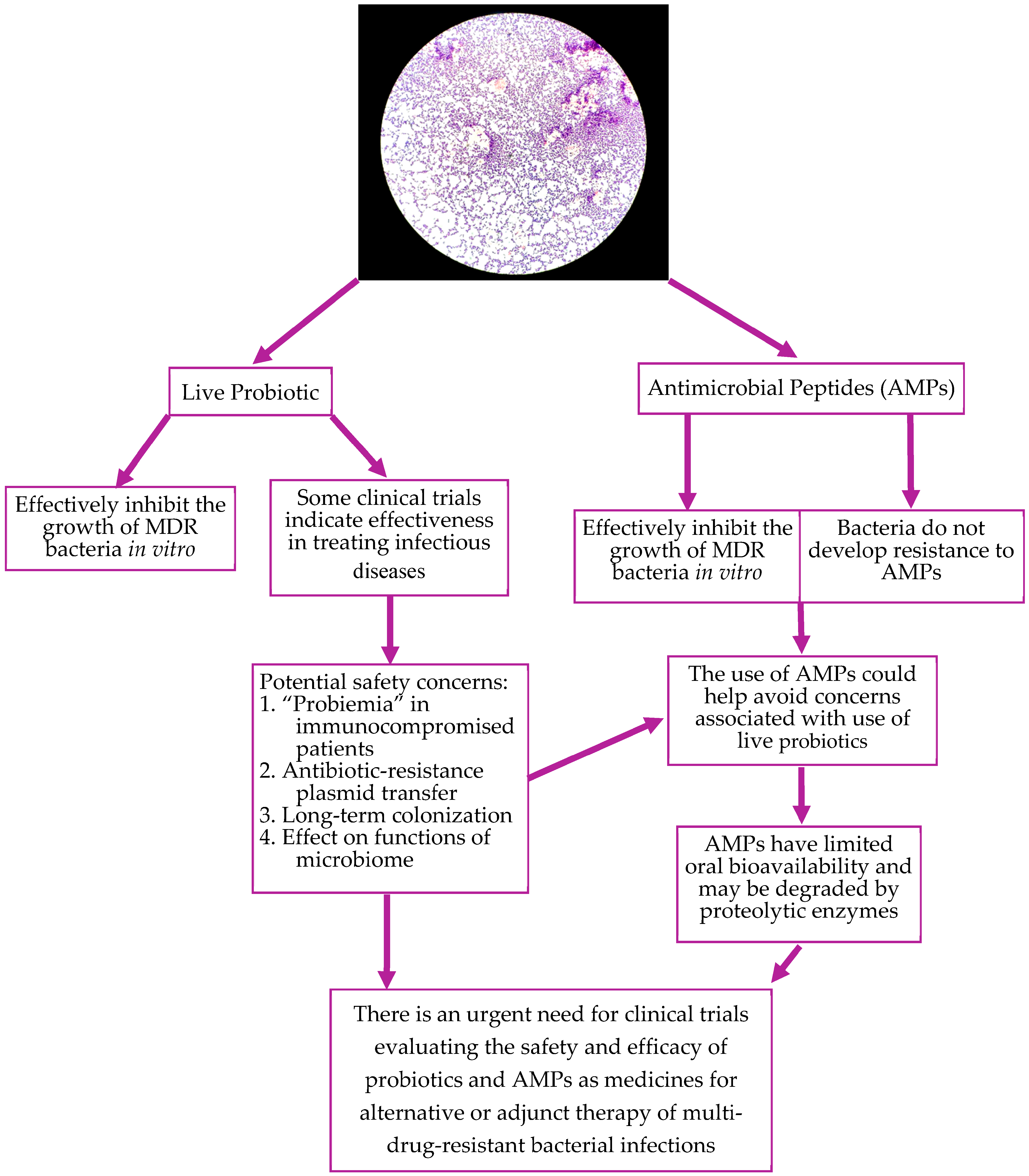
2.2. Challenges of Conventional Treatments of Multi-Drug Resistant Infection
3. Probiotics and Their Bioactive Compounds Interfering with the Growth of Pathogenic Microorganisms
3.1. Probiotics and Their Bioactive Compounds Interfering with the Growth of MRSA

{kind=link}
| Species/Strain | AMP | Activity against | Reference |
|---|---|---|---|
| Lactococcus garvieae | Garvicin KS | MRSA (along with micrococcin P1), Bacillus, Listeria, Enterococcus, along with polymyxin B or nisin, acts against A. baumannii and E. coli | [67,74,75] |
| Enterococcus faecalis 28 and 93 | Enterocin DD28 and DD93 | MRSA | [76] |
| L. plantarum | Plantaricin NC8 β Synthetic Linopeptide | MRSA Linopeptide acts against both Gram-positive and negative bacteria | [77] |
| M. canis | Micasin | MRSA, P. aeruginosa | [73] |
| Streptomyces actinomycinicus PJ85 | Actinomycin D and DGLA AMP | MRSA | [78] |
| Bacillus spp. T12 | Bacin A1, A2, A3, A4 | MRSA | [79] |
| Enterococcus faecalis 14 | Enterocin DD14 | Along with methicillin, acts against MRSA | [80] |
| L. lactis | Nisin Z | Along with methicillin, acts against MRSA | [81] |
| Staphylococcus capitis | Nisin J | MRSA | [82] |
| Bacillus spp. | Mersacidin | MRSA | [83] |
| Lactobacillus pentosus | Pentosin JL1 | MRSA | [84] |
| Bacillus subtilis | BAC-IB17 | MRSA | [85] |
| Enterococcus durans | Durancin 61A | MRSA, C. difficile | [86] |
| Bacillus subtilis subsp. spizizenii | Ba-49 | MRSA | [87] |
| Bacillus spp. MMA | Subtilomycin | L. monocytogenes, Clostridioides sporogenes, MRSA, and heterogeneous vancomycin-intermediate-level-resistant S. aureus. Also active against some Gram-negative bacteria, including Aeromonas hydrophila, Vibrio anguillarum, Alteromonas spp., and P. aeruginosa | [88,89] |
| Bacillus licheniformis | Lichenicidin | B. subtilis, B. pumilus, Bacillus megaterium, S. aureus (methicillin-sensitive and methicillin-resistant strains) | [89] |
| B. amyloliquefaciens GA1 | Amylolysin | MRSA, Bacillus spp. Enterococcus spp., Listeria monocytogenes | [89] |
| Staphylococcus epidermidis strain 224 | Epidermicin NI01 | MRSA | [90] |
3.2. Probiotics and Their Bioactive Compounds Interfering with the Growth of VRE
| Species/Strain | AMP/Antimicrobial Substance | Activity against | Reference |
|---|---|---|---|
| Vagococcus fluvialis | Vagococcocin T | Broad Gram-positive coverage except S. aureus | [91] |
| Pediococcus acidilactici MM33 | Pediocin PA-1/AcH | Enterococcus sp. Lactobacilli, Listeria | [92,101] |
| E. faecalis | EF478 | VRE | [94] |
| L. lactis | Enterococcus K1 | VRE | [95] |
3.3. Probiotics and Their Bioproducts Interfering with the Growth of MRSA and VRE
| Species/Strain | AMP/Antimicrobial Substance | Activity against | Reference |
|---|---|---|---|
| Streptomyces cacaoi | Cacaoidin | MRSA, VRE, C. difficile | [102] |
| L. casei | Lactomodulin | MRSA, VRE, C. difficile, B. cereus, L. monocytogenes, E. coli | [103] |
| Streptomyces antibioticus M7 | Actinomycin V, X2, D | MRSA. VRE, B. subtilis, K. pneumoniae, S. epidermidis, S. typhi | [104] |
| Bacillus pumilus | Pumilicin 4 | MRSA, VRE, B. subtilis, B. licheniformis, B. thurigenesis | [105] |
| Psuedoplectania nigerella | Plectasin | MRSA, VRE, S. pneumoniae, C. diphtherium, C. jeikeium | [106] |
| Streptomyces spp. | Neoactinomycin A and B | MRSA, VRE | [109] |
| L. lactis | Nisin Z (along with lacticin 3145) | E. faecium, MRSA, Lactobacillus sp., Gram-negative bacteria | [92,101,110] |
| Enterococcus faecalis 11 | KT11 | MRSA, VRE. Broad antimicrobial spectrum | [111] |
| Scedosporium apiospermum | Scedosporisin | MRSA, VRE | [112] |
| Bacillus amyloliquefaciens | YD1 | MRSA, VRE | [113] |
| B. subtilis subsp. spizizenii DSM 15029 | Entianin | MRSA, VRE | [89] |
| Bacillus cereus as-1.1846 | Cerecidins | MRSA, VRE | [114] |
4. Clinical Evidence for the Effectiveness of Probiotics for the Treatment of MRSA Infection
| Study Type | Study Population | Intervention | Key Findings | Reference |
|---|---|---|---|---|
| Case report | Adult | Oral vancomycin 250 mg twice daily for 10 days, Saccharomyces boulardi 250 mg twice daily for 15 days, topical mupirocin 2% for 5 days. | Negative MRSA culture after 11 days and 5 months of therapy. | [115] |
| Prospective randomized control study | Adult | Fifty adult patients undergoing live-donor liver transplant between September 2005 and June 2009 randomized into a group receiving 2 days of preoperative and 2 weeks of postoperative synbiotic therapy (Bifidobacterium breve, Lactobacillus casei, and galactooligosaccharides (synbiotic group) and a group without synbiotic therapy (control group). | 4% vs. 25% post-transplant infection in synbiotic vs. control group. | [116] |
| Prospective study | Adult | 7 patients given 4 species of Lactobacillus via oral and intranasal routs (5 × 109 CFU/mL). Treatment given till first negative MRSA culture. | 5 patients had throat clearance, 3 of which also had nasal clearance. Culture negative from 10–37 months after therapy. | [117] |
| Randomized double-blind, placebo-controlled trial | Adult | 48 subjects randomized either to probiotic group (L. rhamnosus HN001 1010 CFU/capsule, one capsule per day) for 4 weeks or control. | At the four-week time point, MRSA was detected in 67% and 50% of probiotic group subjects with nares and the gastrointestinal tract colonization, respectively. Three subjects who initially tested positive for VRE were negative after four weeks of probiotic exposure. | [118] |
| Randomized double-blind, placebo-controlled trial | Adults | 113 subjects once daily oral probiotic or placebo capsule administered for four weeks. The probiotic capsule contained 1 × 1010 colony forming units of L. rhamnosus HN001. | Odds of MRSA presence in probiotic vs. placebo group was 0.27 (0.07–0.98) in any sample and 0.17 (0.04–0.73) in stool samples. | [119] |
| Randomized double-blind, placebo-controlled trial | Adults | 115 participants randomly assigned and received 250 mg of probiotic B subtilis MB40 or placebo once per day for 30 days. | Oral administration of probiotic B. subtilis resulted in significant reduction of S. aureus in the stool (96.8%; p < 0.0001) and the nose (65.4%; p = 0.0002), whereas there were no significant differences in the placebo groups. | [120] |
Clinical Evidence for the Effectiveness of Probiotics for the Eradication of VRE Colonization
| Study Type | Study Population | Intervention | Key Findings | Reference |
|---|---|---|---|---|
| Double-blind, randomized, placebo-controlled trial | Adult | Twenty-seven patients randomly assigned to either a treatment group (receiving 100 g daily of yoghurt containing L. rhamnosus G.G. for 4 weeks) or a control group (receiving standard pasteurized yoghurt). | A total of 11/11 treatment participants cleared VRE and 8 remained VRE negative after 4 weeks of trial completion, 1/12 control participants cleared VRE. After cross over to L. rhamnosus G.G. containing yogurt group, all controls cleared VRE. | [121] |
| Case series | Adult | A total of 4 patients, 2 with end-stage liver disease, 1 with post-liver transplant infection, and 1 with complicated infective endocarditis were given bowel preparation, linezolid, and daptomycin followed by L. rhamnosus G.G. (80 mg). | Three out of four showed VRE clearance. | [122] |
| Open-label prospective follow-up | Newborn | Forty-five patients were randomly assigned to either a treatment group receiving L. rhamnosus G.G. 106 CFU or control group until three consecutive negative cultures or for maximum 6 months. | A total of 21/22 in treatment group vs. 12/23 patients in control group achieved eradication at the end of therapy. | [123] |
| Randomized, single blind | Children | Sixty-one patients were randomly assigned to either a treatment group receiving L. rhamnosus G.G. 3 billion CFU or placebo for 21 days with no follow-up until the end of therapy. | A total of 20 out of 32 patients in treatment group vs. 7 out of 29 in control group were VRE negative at the end of treatment. | [124] |
| Randomized, double blind | Adult | Nine patients were randomly assigned to either a treatment group receiving L. rhamnosus Lcr35 for 5 weeks (109 CFU) or placebo with no follow-up until the end of therapy. | VRE negative culture achieved in 2/6 patients in treatment group vs. ½ patients in placebo group. | [125] |
| Double-blind, randomized, placebo control | Adult | L. rhamnosus G.G., 1010 CFU twice daily for 14 days vs. placebo with 7 days of a follow-up period. | No difference in VRE counts at any time point. No difference in CFU in subjects who received L. rhamnosus GG. | [126] |
| Double-blind, randomized, placebo control | Adult | L. rhamnosus G.G. 1010 CFU vs. placebo until hospital discharge. | Odds of antibiotic resistance organism (ARO) acquisition for VRE in treatment vs. placebo 1.32. The odds of ARO loss for VRE in treatment vs. placebo 0.31 (0.02–4.41). | [127] |
| Double-blind, randomized, placebo control | Adult | L. rhamnosus G.G. 60 × 109 CFU vs. placebo. | A total of 12/21 vs. 15/27 patients cleared VRE in the treatment vs. placebo group. | [128] |
| Prospective cohort with cross-over | Adult | L. rhamnosus G.G. 109 CFU was given to patients with >48 h of hospital stay twice daily until discharge. | Probiotics did not prevent VRE acquisition. | [129] |
5. What Is Next?
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antimicrobial Resistance, C. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States, 2019; Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019. [CrossRef]
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M.; et al. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: A population-level modelling analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Palma, E.; Tilocca, B.; Roncada, P. Antimicrobial Resistance in Veterinary Medicine: An Overview. Int. J. Mol. Sci. 2020, 21, 1914. [Google Scholar] [CrossRef] [PubMed]
- Pomba, C.; Rantala, M.; Greko, C.; Baptiste, K.E.; Catry, B.; van Duijkeren, E.; Mateus, A.; Moreno, M.A.; Pyörälä, S.; Ružauskas, M.; et al. Public health risk of antimicrobial resistance transfer from companion animals. J. Antimicrob. Chemother. 2016, 72, 957–968. [Google Scholar] [CrossRef]
- Tamma, P.D.; Aitken, S.L.; Bonomo, R.A.; Mathers, A.J.; van Duin, D.; Clancy, C.J. Infectious Diseases Society of America 2023 Guidance on the Treatment of Antimicrobial Resistant Gram-Negative Infections. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2023, ciad428. [Google Scholar] [CrossRef]
- Rybak, M.J.; Le, J.; Lodise, T.P.; Levine, D.P.; Bradley, J.S.; Liu, C.; Mueller, B.A.; Pai, M.P.; Wong-Beringer, A.; Rotschafer, J.C.; et al. Therapeutic Monitoring of Vancomycin for Serious Methicillin-resistant Staphylococcus aureus Infections: A Revised Consensus Guideline and Review by the American Society of Health-system Pharmacists, the Infectious Diseases Society of America, the Pediatric Infectious Diseases Society, and the Society of Infectious Diseases Pharmacists. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2020, 71, 1361–1364. [Google Scholar] [CrossRef]
- Liu, C.; Bayer, A.; Cosgrove, S.E.; Daum, R.S.; Fridkin, S.K.; Gorwitz, R.J.; Kaplan, S.L.; Karchmer, A.W.; Levine, D.P.; Murray, B.E.; et al. Clinical Practice Guidelines by the Infectious Diseases Society ofAmerica for the Treatment of Methicillin-Resistant Staphylococcus aureus Infections in Adults and Children. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2011, 52, e18–e55. [Google Scholar] [CrossRef]
- Paul, M.; Carrara, E.; Retamar, P.; Tängdén, T.; Bitterman, R.; Bonomo, R.A.; De Waele, J.; Daikos, G.L.; Akova, M.; Harbarth, S.; et al. European Society of Clinical Microbiology and Infectious Diseases (ESCMID) guidelines for the treatment of infections caused by multidrug-resistant Gram-negative bacilli (endorsed by European society of intensive care medicine). Clin. Microbiol. Infect. 2022, 28, 521–547. [Google Scholar] [CrossRef]
- FAO; WHO. Evaluation of Health and Nutritional Properties of Powder Milk and Live Lactic Acid Bacteria; Food and Agriculture Organization of the United Nations: Rome, Italy; World Health Organization: Geneva, Switzerland, 2001; pp. 1–34.
- Liu, Y.; Tran, D.Q.; Rhoads, J.M. Probiotics in Disease Prevention and Treatment. J. Clin. Pharmacol. 2018, 58 (Suppl. S10), S164–S179. [Google Scholar] [CrossRef]
- Doron, S.; Snydman, D.R. Risk and safety of probiotics. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2015, 60 (Suppl. S2), S129–S134. [Google Scholar] [CrossRef]
- Rondanelli, M.; Faliva, M.A.; Perna, S.; Giacosa, A.; Peroni, G.; Castellazzi, A.M. Using probiotics in clinical practice: Where are we now? A review of existing meta-analyses. Gut Microbes 2017, 8, 521–543. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.Q.; Li, L. The Potential Role of Probiotics in Cancer Prevention and Treatment. Nutr. Cancer 2016, 68, 535–544. [Google Scholar] [CrossRef] [PubMed]
- So, S.S.; Wan, M.L.; El-Nezami, H. Probiotics-mediated suppression of cancer. Curr. Opin. Oncol. 2017, 29, 62–72. [Google Scholar] [CrossRef]
- Devaraj, N.K.; Suppiah, S.; Veettil, S.K.; Ching, S.M.; Lee, K.W.; Menon, R.K.; Soo, M.J.; Deuraseh, I.; Hoo, F.K.; Sivaratnam, D. The Effects of Probiotic Supplementation on the Incidence of Diarrhea in Cancer Patients Receiving Radiation Therapy: A Systematic Review with Meta-Analysis and Trial Sequential Analysis of Randomized Controlled Trials. Nutrients 2019, 11, 2886. [Google Scholar] [CrossRef] [PubMed]
- Casas-Solís, J.; Huizar-López, M.D.R.; Irecta-Nájera, C.A.; Pita-López, M.L.; Santerre, A. Immunomodulatory Effect of Lactobacillus casei in a Murine Model of Colon Carcinogenesis. Probiotics Antimicrob. Proteins 2020, 12, 1012–1024. [Google Scholar] [CrossRef]
- Umborowati, M.A.; Damayanti, D.; Anggraeni, S.; Endaryanto, A.; Surono, I.S.; Effendy, I.; Prakoeswa, C.R.S. The role of probiotics in the treatment of adult atopic dermatitis: A meta-analysis of randomized controlled trials. J. Health Popul. Nutr. 2022, 41, 37. [Google Scholar] [CrossRef]
- Nath, A.; Molnár, M.A.; Csighy, A.; Kőszegi, K.; Galambos, I.; Huszár, K.P.; Koris, A.; Vatai, G. Biological Activities of Lactose-Based Prebiotics and Symbiosis with Probiotics on Controlling Osteoporosis, Blood-Lipid and Glucose Levels. Medicina 2018, 54, 98. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C.J. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar] [CrossRef]
- Raheem, A.; Liang, L.; Zhang, G.; Cui, S. Modulatory Effects of Probiotics During Pathogenic Infections With Emphasis on Immune Regulation. Front. Immunol. 2021, 12, 616713. [Google Scholar] [CrossRef]
- Sassone-Corsi, M.; Raffatellu, M. No vacancy: How beneficial microbes cooperate with immunity to provide colonization resistance to pathogens. J. Immunol. 2015, 194, 4081–4087. [Google Scholar] [CrossRef]
- Żółkiewicz, J.; Marzec, A.; Ruszczyński, M.; Feleszko, W. Postbiotics—A Step Beyond Pre- and Probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Lu, T.K. Development and Challenges of Antimicrobial Peptides for Therapeutic Applications. Antibiotics 2020, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Zimina, M.; Babich, O.; Prosekov, A.; Sukhikh, S.; Ivanova, S.; Shevchenko, M.; Noskova, S. Overview of Global Trends in Classification, Methods of Preparation and Application of Bacteriocins. Antibiotics 2020, 9, 553. [Google Scholar] [CrossRef] [PubMed]
- Bin Hafeez, A.; Jiang, X.; Bergen, P.J.; Zhu, Y. Antimicrobial Peptides: An Update on Classifications and Databases. Int. J. Mol. Sci. 2021, 22, 1691. [Google Scholar] [CrossRef]
- Boparai, J.K.; Sharma, P.K. Mini Review on Antimicrobial Peptides, Sources, Mechanism and Recent Applications. Protein Pept. Lett. 2020, 27, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Jernigan, J.A.; Hatfield, K.M.; Wolford, H.; Nelson, R.E.; Olubajo, B.; Reddy, S.C.; McCarthy, N.; Paul, P.; McDonald, L.C.; Kallen, A.; et al. Multidrug-Resistant Bacterial Infections in U.S. Hospitalized Patients, 2012–2017. N. Engl. J. Med. 2020, 382, 1309–1319. [Google Scholar] [CrossRef]
- Cairns, K.A.; Udy, A.A.; Peel, T.N.; Abbot, I.J.; Dooley, M.J.; Peleg, A.Y. Therapeutics for Vancomycin-Resistant Enterococcal Bloodstream Infections. Clin. Microbiol. Rev. 2023, 36, e0005922. [Google Scholar] [CrossRef]
- Baddour, L.M.; Wilson, W.R.; Bayer, A.S.; Fowler, V.G.; Tleyjeh, I.M.; Rybak, M.J.; Barsic, B.; Lockhart, P.B.; Gewitz, M.H.; Levison, M.E.; et al. Infective Endocarditis in Adults: Diagnosis, Antimicrobial Therapy, and Management of Complications. Circulation 2015, 132, 1435–1486. [Google Scholar] [CrossRef]
- Jahanbakhsh, S.; Singh, N.; Yim, J.; Kebriaei, R.; Smith, J.; Lev, K.; Tran, T.; Rose, W.; Arias, C.; Rybak, M. Impact of Daptomycin Dose Exposure Alone or in Combination with β-Lactams or Rifampin against VancomycinResistant Enterococci in an In Vitro Biofilm Model. Antimicrob. Agents Chemother. 2020, 64, 10-1128. [Google Scholar] [CrossRef]
- Roberts, J.A.; Paul, S.K.; Akova, M.; Bassetti, M.; De Waele, J.J.; Dimopoulos, G.; Kaukonen, K.-M.; Koulenti, D.; Martin, C.; Montravers, P.; et al. DALI: Defining Antibiotic Levels in Intensive Care Unit Patients: Are Current -Lactam Antibiotic Doses Sufficient for Critically Ill Patients? Clin. Infect. Dis. 2014, 58, 1072–1083. [Google Scholar] [CrossRef]
- Legg, A.; Carmichael, S.; Chai, M.G.; Roberts, J.A.; Cotta, M.O. Beta-Lactam Dose Optimisation in the Intensive Care Unit: Targets, Therapeutic Drug Monitoring and Toxicity. Antibiotics 2023, 12, 870. [Google Scholar] [CrossRef] [PubMed]
- CDC. COVID-19: U.S. Impact on Antimicrobial Resistance; Special Report 2022; Department of Health and Human Services, CDC: Atlanta, GA, USA, 2022. Available online: https://www.cdc.gov/drugresistance/covid19.html (accessed on 1 May 2023). [CrossRef]
- Ozma, M.A.; Abbasi, A.; Asgharzadeh, M.; Pagliano, P.; Guarino, A.; Köse, Ş.; Samadi Kafil, H. Antibiotic therapy for pan-drug-resistant infections. Infez. Med. 2022, 30, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Lahtinen, S.J.; Jalonen, L.; Ouwehand, A.C.; Salminen, S.J. Specific Bifidobacterium strains isolated from elderly subjects inhibit growth of Staphylococcus aureus. Int. J. Food Microbiol. 2007, 117, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Lazarenko, L.; Babenko, L.; Sichel, L.S.; Pidgorskyi, V.; Mokrozub, V.; Voronkova, O.; Spivak, M. Antagonistic Action of Lactobacilli and Bifidobacteria in Relation to Staphylococcus aureus and Their Influence on the Immune Response in Cases of Intravaginal Staphylococcosis in Mice. Probiotics Antimicrob. Proteins 2012, 4, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Piewngam, P.; Zheng, Y.; Nguyen, T.H.; Dickey, S.W.; Joo, H.S.; Villaruz, A.E.; Glose, K.A.; Fisher, E.L.; Hunt, R.L.; Li, B.; et al. Pathogen elimination by probiotic Bacillus via signalling interference. Nature 2018, 562, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Kim, B.S.; Kang, S.S. Bacteriocin of Pediococcus acidilactici HW01 Inhibits Biofilm Formation and Virulence Factor Production by Pseudomonas aeruginosa. Probiotics Antimicrob. Proteins 2020, 12, 73–81. [Google Scholar] [CrossRef]
- Zagari, R.M.; Frazzoni, L.; Marasco, G.; Fuccio, L.; Bazzoli, F. Treatment of Helicobacter pylori infection: A clinical practice update. Minerva Med. 2021, 112, 281–287. [Google Scholar] [CrossRef]
- Aiba, Y.; Ishikawa, H.; Tokunaga, M.; Komatsu, Y. Anti-Helicobacter pylori activity of non-living, heat-killed form of lactobacilli including Lactobacillus johnsonii No.1088. FEMS Microbiol. Lett. 2017, 364, fnx102. [Google Scholar] [CrossRef]
- Lagrafeuille, R.; Miquel, S.; Balestrino, D.; Vareille-Delarbre, M.; Chain, F.; Langella, P.; Forestier, C. Opposing effect of Lactobacillus on in vitro Klebsiella pneumoniae in biofilm and in an in vivo intestinal colonisation model. Benef. Microbes 2018, 9, 87–100. [Google Scholar] [CrossRef]
- Vieira, A.T.; Rocha, V.M.; Tavares, L.; Garcia, C.C.; Teixeira, M.M.; Oliveira, S.C.; Cassali, G.D.; Gamba, C.; Martins, F.S.; Nicoli, J.R. Control of Klebsiella pneumoniae pulmonary infection and immunomodulation by oral treatment with the commensal probiotic Bifidobacterium longum 51A. Microbes Infect. 2016, 18, 180–189. [Google Scholar] [CrossRef]
- Nyanzi, R.; Awouafack, M.D.; Steenkamp, P.; Jooste, P.J.; Eloff, J.N. Anticandidal activity of cell extracts from 13 probiotic Lactobacillus strains and characterisation of lactic acid and a novel fatty acid derivative from one strain. Food Chem. 2014, 164, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Wannun, P.; Piwat, S.; Teanpaisan, R. Purification, Characterization, and Optimum Conditions of Fermencin SD11, a Bacteriocin Produced by Human Orally Lactobacillus fermentum SD11. Appl. Biochem. Biotechnol. 2016, 179, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Orsi, C.F.; Sabia, C.; Ardizzoni, A.; Colombari, B.; Neglia, R.G.; Peppoloni, S.; Morace, G.; Blasi, E. Inhibitory effects of different lactobacilli on Candida albicans hyphal formation and biofilm development. J. Biol. Regul. Homeost. Agents 2014, 28, 743–752. [Google Scholar]
- Itapary Dos Santos, C.; Ramos França, Y.; Duarte Lima Campos, C.; Quaresma Bomfim, M.R.; Oliveira Melo, B.; Assunção Holanda, R.; Santos, V.L.; Gomes Monteiro, S.; Buozzi Moffa, E.; Souza Monteiro, A.; et al. Antifungal and Antivirulence Activity of Vaginal Lactobacillus Spp. Products against Candida Vaginal Isolates. Pathogens 2019, 8, 150. [Google Scholar] [CrossRef]
- Ribeiro, F.C.; de Barros, P.P.; Rossoni, R.D.; Junqueira, J.C.; Jorge, A.O. Lactobacillus rhamnosus inhibits Candida albicans virulence factors in vitro and modulates immune system in Galleria mellonella. J. Appl. Microbiol. 2017, 122, 201–211. [Google Scholar] [CrossRef]
- Chew, S.Y.; Cheah, Y.K.; Seow, H.F.; Sandai, D.; Than, L.T. In vitro modulation of probiotic bacteria on the biofilm of Candida glabrata. Anaerobe 2015, 34, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Krzyściak, W.; Kościelniak, D.; Papież, M.; Vyhouskaya, P.; Zagórska-Świeży, K.; Kołodziej, I.; Bystrowska, B.; Jurczak, A. Effect of a Lactobacillus Salivarius Probiotic on a Double-Species Streptococcus Mutans and Candida Albicans Caries Biofilm. Nutrients 2017, 9, 1242. [Google Scholar] [CrossRef]
- Rossoni, R.D.; Fuchs, B.B.; de Barros, P.P.; Velloso, M.D.; Jorge, A.O.; Junqueira, J.C.; Mylonakis, E. Lactobacillus paracasei modulates the immune system of Galleria mellonella and protects against Candida albicans infection. PLoS ONE 2017, 12, e0173332. [Google Scholar] [CrossRef]
- Liao, H.; Liu, S.; Wang, H.; Su, H.; Liu, Z. Enhanced antifungal activity of bovine lactoferrin-producing probiotic Lactobacillus casei in the murine model of vulvovaginal candidiasis. BMC Microbiol. 2019, 19, 7. [Google Scholar] [CrossRef]
- Kober, M.M.; Bowe, W.P. The effect of probiotics on immune regulation, acne, and photoaging. Int. J. Women’s Dermatol. 2015, 1, 85–89. [Google Scholar] [CrossRef]
- Rosignoli, C.; Thibaut de Ménonville, S.; Orfila, D.; Béal, M.; Bertino, B.; Aubert, J.; Mercenier, A.; Piwnica, D. A topical treatment containing heat-treated Lactobacillus johnsonii NCC 533 reduces Staphylococcus aureus adhesion and induces antimicrobial peptide expression in an in vitro reconstructed human epidermis model. Exp. Dermatol. 2018, 27, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Navarro-López, V.; Ramírez-Boscá, A.; Ramón-Vidal, D.; Ruzafa-Costas, B.; Genovés-Martínez, S.; Chenoll-Cuadros, E.; Carrión-Gutiérrez, M.; Horga de la Parte, J.; Prieto-Merino, D.; Codoñer-Cortés, F.M. Effect of Oral Administration of a Mixture of Probiotic Strains on SCORAD Index and Use of Topical Steroids in Young Patients With Moderate Atopic Dermatitis: A Randomized Clinical Trial. JAMA Dermatol. 2018, 154, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Di Marzio, L.; Centi, C.; Cinque, B.; Masci, S.; Giuliani, M.; Arcieri, A.; Zicari, L.; De Simone, C.; Cifone, M.G. Effect of the lactic acid bacterium Streptococcus thermophilus on stratum corneum ceramide levels and signs and symptoms of atopic dermatitis patients. Exp. Dermatol. 2003, 12, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Blanchet-Réthoré, S.; Bourdès, V.; Mercenier, A.; Haddar, C.H.; Verhoeven, P.O.; Andres, P. Effect of a lotion containing the heat-treated probiotic strain Lactobacillus johnsonii NCC 533 on Staphylococcus aureus colonization in atopic dermatitis. Clin. Cosmet. Investig. Dermatol. 2017, 10, 249–257. [Google Scholar] [CrossRef]
- Knackstedt, R.; Knackstedt, T.; Gatherwright, J. The role of topical probiotics in skin conditions: A systematic review of animal and human studies and implications for future therapies. Exp. Dermatol. 2020, 29, 15–21. [Google Scholar] [CrossRef]
- Karska-Wysocki, B.; Bazo, M.; Smoragiewicz, W. Antibacterial activity of Lactobacillus acidophilus and Lactobacillus casei against methicillin-resistant Staphylococcus aureus (MRSA). Microbiol. Res. 2010, 165, 674–686. [Google Scholar] [CrossRef]
- Saravanakumari, P.; Mani, K. Structural characterization of a novel xylolipid biosurfactant from Lactococcus lactis and analysis of antibacterial activity against multi-drug resistant pathogens. Bioresour. Technol. 2010, 101, 8851–8854. [Google Scholar] [CrossRef]
- Bhola, J.; Bhadekar, R. Invitro synergistic activity of lactic acid bacteria against multi-drug resistant staphylococci. BMC Complement. Altern. Med. 2019, 19, 70. [Google Scholar] [CrossRef]
- Bendjeddou, K.; Fons, M.; Strocker, P.; Sadoun, D. Characterization and purification of a bacteriocin from Lactobacillus paracasei subsp. paracasei BMK2005, an intestinal isolate active against multidrug-resistant pathogens. World J. Microbiol. Biotechnol. 2012, 28, 1543–1552. [Google Scholar] [CrossRef]
- Chen, P.W.; Jheng, T.T.; Shyu, C.L.; Mao, F.C. Synergistic antibacterial efficacies of the combination of bovine lactoferrin or its hydrolysate with probiotic secretion in curbing the growth of meticillin-resistant Staphylococcus aureus. J. Med. Microbiol. 2013, 62, 1845–1851. [Google Scholar] [CrossRef]
- Moormeier, D.E.; Bayles, K.W. Staphylococcus aureus biofilm: A complex developmental organism. Mol. Microbiol. 2017, 104, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Walencka, E.; Różalska, S.; Sadowska, B.; Różalska, B. The influence of Lactobacillus acidophilus-derived surfactants on staphylococcal adhesion and biofilm formation. Folia Microbiol. 2008, 53, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Kumar, L.M.; Saad, W.Z.; Mohamad, R.; Rahim, R.A. Influence of biofilm-forming lactic acid bacteria against methicillin-resistant Staphylococcus aureus (MRSA S547). Asian Pac. J. Trop. Biomed. 2017, 7, 1107–1115. [Google Scholar] [CrossRef]
- Kranjec, C.; Ovchinnikov, K.V.; Grønseth, T.; Ebineshan, K.; Srikantam, A.; Diep, D.B. A bacteriocin-based antimicrobial formulation to effectively disrupt the cell viability of methicillin-resistant Staphylococcus aureus (MRSA) biofilms. Npj Biofilms Microbiomes 2020, 6, 58. [Google Scholar] [CrossRef]
- Salman, M.K.; Abuqwider, J.; Mauriello, G. Anti-Quorum Sensing Activity of Probiotics: The Mechanism and Role in Food and Gut Health. Microorganisms 2023, 11, 793. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wang, B.; Liu, S.; Chen, Y.; Lin, Y.; Liu, Z.; Zhang, X.; Yu, B. Bacillus subtilis revives conventional antibiotics against Staphylococcus aureus osteomyelitis. Microb. Cell Factories 2021, 20, 102. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, M.; Dolatabadi, S.; Ghorbannezhad, G.; Sharifi, F.; Rahimi, H.R. The role of probiotics in inhibition mechanism of methicillin-resistant Staphylococcus aureus. McGill J. Med. 2020, 18, 2194. [Google Scholar] [CrossRef]
- Jayashree, S.; Karthikeyan, R.; Nithyalakshmi, S.; Ranjani, J.; Gunasekaran, P.; Rajendhran, J. Anti-adhesion Property of the Potential Probiotic Strain Lactobacillus fermentum 8711 against Methicillin-Resistant Staphylococcus aureus (MRSA). Front. Microbiol. 2018, 9, 411. [Google Scholar] [CrossRef]
- Mokoena, M.P. Lactic Acid Bacteria and Their Bacteriocins: Classification, Biosynthesis and Applications against Uropathogens: A Mini-Review. Molecules 2017, 22, 1255. [Google Scholar] [CrossRef]
- Zhu, S.; Gao, B.; Harvey, P.J.; Craik, D.J. Dermatophytic defensin with antiinfective potential. Proc. Natl. Acad. Sci. USA 2012, 109, 8495–8500. [Google Scholar] [CrossRef]
- Chi, H.; Holo, H. Synergistic Antimicrobial Activity Between the Broad Spectrum Bacteriocin Garvicin KS and Nisin, Farnesol and Polymyxin B against Gram-Positive and Gram-Negative Bacteria. Curr. Microbiol. 2018, 75, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikov, K.V.; Chi, H.; Mehmeti, I.; Holo, H.; Nes, I.F.; Diep, D.B. Novel Group of Leaderless Multipeptide Bacteriocins from Gram-Positive Bacteria. Appl. Environ. Microbiol. 2016, 82, 5216–5224. [Google Scholar] [CrossRef] [PubMed]
- Al Atya, A.K.; Belguesmia, Y.; Chataigne, G.; Ravallec, R.; Vachée, A.; Szunerits, S.; Boukherroub, R.; Drider, D. Anti-MRSA Activities of Enterocins DD28 and DD93 and Evidences on Their Role in the Inhibition of Biofilm Formation. Front. Microbiol. 2016, 7, 817. [Google Scholar] [CrossRef]
- Wiman, E.; Zattarin, E.; Aili, D.; Bengtsson, T.; Selegård, R.; Khalaf, H. Development of novel broad-spectrum antimicrobial lipopeptides derived from plantaricin NC8 β. Sci. Rep. 2023, 13, 4104. [Google Scholar] [CrossRef] [PubMed]
- Chanthasena, P.; Hua, Y.; Rosyidah, A.; Pathom-Aree, W.; Limphirat, W.; Nantapong, N. Isolation and Identification of Bioactive Compounds from Streptomyces actinomycinicus PJ85 and Their In Vitro Antimicrobial Activities against Methicillin-Resistant Staphylococcus aureus. Antibiotics 2022, 11, 1797. [Google Scholar] [CrossRef]
- Liu, S.; Deng, S.; Liu, H.; Tang, L.; Wang, M.; Xin, B.; Li, F. Four Novel Leaderless Bacteriocins, Bacin A1, A2, A3, and A4 Exhibit Potent Antimicrobial and Antibiofilm Activities against Methicillin-Resistant Staphylococcus aureus. Microbiol. Spectr. 2022, 10, e00945-22. [Google Scholar] [CrossRef]
- Teiar, R.; Pérez-Ramos, A.; Zgheib, H.; Cudennec, B.; Belguesmia, Y.; Drider, D. Anti-adhesion and Anti-inflammatory Potential of the Leaderless Class IIb Bacteriocin Enterocin DD14. Probiotics Antimicrob. Proteins 2022, 14, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Mulders, J.W.; Boerrigter, I.J.; Rollema, H.S.; Siezen, R.J.; de Vos, W.M. Identification and characterization of the lantibiotic nisin Z, a natural nisin variant. Eur. J. Biochem. 1991, 201, 581–584. [Google Scholar] [CrossRef]
- O’Sullivan, J.N.; O’Connor, P.M.; Rea, M.C.; O’Sullivan, O.; Walsh, C.J.; Healy, B.; Mathur, H.; Field, D.; Hill, C.; Ross, R.P. Nisin J, a Novel Natural Nisin Variant, Is Produced by Staphylococcus capitis Sourced from the Human Skin Microbiota. J. Bacteriol. 2020, 202, e00639-19. [Google Scholar] [CrossRef]
- Kruszewska, D.; Sahl, H.G.; Bierbaum, G.; Pag, U.; Hynes, S.O.; Ljungh, A. Mersacidin eradicates methicillin-resistant Staphylococcus aureus (MRSA) in a mouse rhinitis model. J. Antimicrob. Chemother. 2004, 54, 648–653. [Google Scholar] [CrossRef]
- Jiang, H.; Zou, J.; Cheng, H.; Fang, J.; Huang, G. Purification, Characterization, and Mode of Action of Pentocin JL-1, a Novel Bacteriocin Isolated from Lactobacillus pentosus, against Drug-Resistant Staphylococcus aureus. Biomed Res. Int. 2017, 2017, 7657190. [Google Scholar] [CrossRef] [PubMed]
- Ansari, A.; Zohra, R.R.; Tarar, O.M.; Qader, S.A.U.; Aman, A. Screening, purification and characterization of thermostable, protease resistant Bacteriocin active against methicillin resistant Staphylococcus aureus (MRSA). BMC Microbiol. 2018, 18, 192. [Google Scholar] [CrossRef] [PubMed]
- Hanchi, H.; Hammami, R.; Gingras, H.; Kourda, R.; Bergeron, M.G.; Ben Hamida, J.; Ouellette, M.; Fliss, I. Inhibition of MRSA and of Clostridium difficile by durancin 61A: Synergy with bacteriocins and antibiotics. Future Microbiol. 2017, 12, 205–212. [Google Scholar] [CrossRef]
- Taggar, R.; Singh, S.; Bhalla, V.; Bhattacharyya, M.S.; Sahoo, D.K. Deciphering the Antibacterial Role of Peptide From Bacillus subtilis subsp. spizizenii Ba49 against Staphylococcus aureus. Front. Microbiol. 2021, 12, 708712. [Google Scholar] [CrossRef] [PubMed]
- Phelan, R.W.; Barret, M.; Cotter, P.D.; Connor, P.M.; Chen, R.; Morrissey, J.P.; Dobson, A.D.W.; Gara, F.; Barbosa, T.M. Subtilomycin: A New Lantibiotic from Bacillus subtilis Strain MMA7 Isolated from the Marine Sponge Haliclona simulans. Mar. Drugs 2013, 11, 1878–1898. [Google Scholar] [CrossRef]
- Barbosa, J.; Caetano, T.; Mendo, S. Class I and Class II Lanthipeptides Produced by Bacillus spp. J. Nat. Prod. 2015, 78, 2850–2866. [Google Scholar] [CrossRef]
- Sandiford, S.; Upton, M. Identification, characterization, and recombinant expression of epidermicin NI01, a novel unmodified bacteriocin produced by Staphylococcus epidermidis that displays potent activity against Staphylococci. Antimicrob. Agents Chemother. 2012, 56, 1539–1547. [Google Scholar] [CrossRef]
- Rosenbergová, Z.; Oftedal Thomas, F.; Ovchinnikov Kirill, V.; Thiyagarajah, T.; Rebroš, M.; Diep Dzung, B. Identification of a Novel Two-Peptide Lantibiotic from Vagococcus fluvialis. Microbiol. Spectr. 2022, 10, e00954-22. [Google Scholar] [CrossRef]
- Millette, M.; Cornut, G.; Dupont, C.; Shareck, F.; Archambault, D.; Lacroix, M. Capacity of Human Nisin- and Pediocin-Producing Lactic Acid Bacteria To Reduce Intestinal Colonization by Vancomycin-Resistant Enterococci. Appl. Environ. Microbiol. 2008, 74, 1997–2003. [Google Scholar] [CrossRef]
- Li, X.; Song, L.; Zhu, S.; Xiao, Y.; Huang, Y.; Hua, Y.; Chu, Q.; Ren, Z. Two Strains of Lactobacilli Effectively Decrease the Colonization of VRE in a Mouse Model. Front. Cell. Infect. Microbiol. 2019, 9, 6. [Google Scholar] [CrossRef]
- Phumisantiphong, U.; Siripanichgon, K.; Reamtong, O.; Diraphat, P. A novel bacteriocin from Enterococcus faecalis 478 exhibits a potent activity against vancomycin-resistant enterococci. PLoS ONE 2017, 12, e0186415. [Google Scholar] [CrossRef]
- Ovchinnikov, K.V.; Kristiansen, P.E.; Straume, D.; Jensen, M.S.; Aleksandrzak-Piekarczyk, T.; Nes, I.F.; Diep, D.B. The Leaderless Bacteriocin Enterocin K1 Is Highly Potent against Enterococcus faecium: A Study on Structure, Target Spectrum and Receptor. Front. Microbiol. 2017, 8, 774. [Google Scholar] [CrossRef] [PubMed]
- Plupjeen, S.N.; Chawjiraphan, W.; Charoensiddhi, S.; Nitisinprasert, S.; Nakphaichit, M. Lactococcus lactis KA-FF 1-4 reduces vancomycin-resistant enterococci and impacts the human gut microbiome. 3 Biotech 2020, 10, 295. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Lee, K.; Hsu, M.; Nau, G.; Mylonakis, E.; Ramratnam, B. Lactobacillus-derived extracellular vesicles enhance host immune responses against vancomycin-resistant enterococci. BMC Microbiol. 2017, 17, 66. [Google Scholar] [CrossRef] [PubMed]
- Bucheli, J.E.V.; Fugaban, J.I.I.; Holzapfel, W.H.; Todorov, S.D. Combined Action of Antibiotics and Bacteriocins against Vancomycin-Resistant Enterococci. Microorganisms 2022, 10, 1423. [Google Scholar] [CrossRef]
- El-Kazzaz, S.S.; Abou El-Khier, N.T. Effect of the lantibiotic nisin on inhibitory and bactericidal activities of antibiotics used against vancomycin-resistant enterococci. J. Glob. Antimicrob. Resist. 2020, 22, 263–269. [Google Scholar] [CrossRef]
- Sun, W.S.; Lee, Y.J.; Tsai, K.N.; Ho, Y.H.; Fang, S.B. Probiotic Cocktail Identified by Microbial Network Analysis Inhibits Growth, Virulence Gene Expression, and Host Cell Colonization of Vancomycin-Resistant Enterococci. Microorganisms 2020, 8, 816. [Google Scholar] [CrossRef]
- Millette, M.; Dupont, C.; Archambault, D.; Lacroix, M. Partial characterization of bacteriocins produced by human Lactococcus lactis and Pediococccus acidilactici isolates. J. Appl. Microbiol. 2007, 102, 274–282. [Google Scholar] [CrossRef]
- Deisinger, J.P.; Arts, M.; Kotsogianni, I.; Puls, J.S.; Grein, F.; Ortiz-López, F.J.; Martin, N.I.; Müller, A.; Genilloud, O.; Schneider, T. Dual targeting of the class V lanthipeptide antibiotic cacaoidin. iScience 2023, 26, 106394. [Google Scholar] [CrossRef]
- Mousa, W.K.; Ghemrawi, R.; Abu-Izneid, T.; Ramadan, A.; Al-Marzooq, F. Discovery of Lactomodulin, a Unique Microbiome-Derived Peptide That Exhibits Dual Anti-Inflammatory and Antimicrobial Activity against Multidrug-Resistant Pathogens. Int. J. Mol. Sci. 2023, 24, 6901. [Google Scholar] [CrossRef]
- Sharma, M.; Manhas, R.K. Purification and characterization of actinomycins from Streptomyces strain M7 active against methicillin resistant Staphylococcus aureus and vancomycin resistant Enterococcus. BMC Microbiol. 2019, 19, 44. [Google Scholar] [CrossRef] [PubMed]
- Aunpad, R.; Na-Bangchang, K. Pumilicin 4, a novel bacteriocin with anti-MRSA and anti-VRE activity produced by newly isolated bacteria Bacillus pumilus strain WAPB4. Curr. Microbiol. 2007, 55, 308–313. [Google Scholar] [CrossRef]
- Mygind, P.H.; Fischer, R.L.; Schnorr, K.M.; Hansen, M.T.; Sönksen, C.P.; Ludvigsen, S.; Raventós, D.; Buskov, S.; Christensen, B.; De Maria, L.; et al. Plectasin is a peptide antibiotic with therapeutic potential from a saprophytic fungus. Nature 2005, 437, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Breidenstein, E.B.; Courvalin, P.; Meziane-Cherif, D. Antimicrobial Activity of Plectasin NZ2114 in Combination with Cell Wall Targeting Antibiotics against VanA-Type Enterococcus faecalis. Microb. Drug Resist. 2015, 21, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.Q.; Hady, W.A.; Deslandes, A.; Rey, A.; Fraisse, L.; Kristensen, H.H.; Yeaman, M.R.; Bayer, A.S. Efficacy of NZ2114, a novel plectasin-derived cationic antimicrobial peptide antibiotic, in experimental endocarditis due to methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 5325–5330. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, Y.; Wang, M.; Tan, Y.; Hu, X.; He, H.; Xiao, C.; You, X.; Wang, Y.; Gan, M. Neo-actinomycins A and B, natural actinomycins bearing the 5H-oxazolo[4,5-b]phenoxazine chromophore, from the marine-derived Streptomyces sp. IMB094. Sci. Rep. 2017, 7, 3591. [Google Scholar] [CrossRef]
- Ellis, J.-C.; Ross, R.P.; Hill, C. Nisin Z and lacticin 3147 improve efficacy of antibiotics against clinically significant bacteria. Future Microbiol. 2019, 14, 1573–1587. [Google Scholar] [CrossRef] [PubMed]
- Abanoz, H.S.; Kunduhoglu, B. Antimicrobial Activity of a Bacteriocin Produced by Enterococcus faecalis KT11 against Some Pathogens and Antibiotic-Resistant Bacteria. Korean J. Food Sci. Anim. Resour. 2018, 38, 1064–1079. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Liu, S.; Wang, H. Invasive fungi-derived defensins kill drug-resistant bacterial pathogens. Peptides 2018, 99, 82–91. [Google Scholar] [CrossRef]
- Rahman, M.S.; Choi, Y.H.; Choi, Y.S.; Yoo, J.C. Glycin-rich antimicrobial peptide YD1 from B. amyloliquefaciens, induced morphological alteration in and showed affinity for plasmid DNA of E. coli. AMB Express 2017, 7, 8. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, L.; Teng, K.; Sun, S.; Sun, Z.; Zhong, J. Cerecidins, novel lantibiotics from Bacillus cereus with potent antimicrobial activity. Appl. Environ. Microbiol. 2014, 80, 2633–2643. [Google Scholar] [CrossRef]
- Sizemore, E.N.; Rivas, K.M.; Valdes, J.; Caballero, J. Enteral vancomycin and probiotic use for methicillin-resistant Staphylococcus aureus antibiotic-associated diarrhoea. BMJ Case Rep. 2012, 2012, bcr2012006366. [Google Scholar] [CrossRef]
- Eguchi, S.; Takatsuki, M.; Hidaka, M.; Soyama, A.; Ichikawa, T.; Kanematsu, T. Perioperative synbiotic treatment to prevent infectious complications in patients after elective living donor liver transplantation: A prospective randomized study. Am. J. Surg. 2011, 201, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Roos, K.; Simark-Mattsson, C.; Grahn Håkansson, E.; Larsson, L.; Sandberg, T.; Ahrén, C. Can probiotic lactobacilli eradicate persistent carriage of meticillin-resistant Staphylococcus aureus? J. Hosp. Infect. 2011, 78, 77–78. [Google Scholar] [CrossRef] [PubMed]
- Warrack, S.; Panjikar, P.; Duster, M.; Safdar, N. Tolerability of a probiotic in subjects with a history of methicillin-resistant Staphylococcus aureus colonisation. Benef. Microbes 2014, 5, 389–395. [Google Scholar] [CrossRef]
- Eggers, S.; Barker, A.K.; Valentine, S.; Hess, T.; Duster, M.; Safdar, N. Effect of Lactobacillus rhamnosus HN001 on carriage of Staphylococcus aureus: Results of the impact of probiotics for reducing infections in veterans (IMPROVE) study. BMC Infect. Dis. 2018, 18, 129. [Google Scholar] [CrossRef] [PubMed]
- Piewngam, P.; Khongthong, S.; Roekngam, N.; Theapparat, Y.; Sunpaweravong, S.; Faroongsarng, D.; Otto, M. Probiotic for pathogen-specific Staphylococcus aureus decolonisation in Thailand: A phase 2, double-blind, randomised, placebo-controlled trial. Lancet Microbe 2023, 4, e75–e83. [Google Scholar] [CrossRef] [PubMed]
- Manley, K.J.; Fraenkel, M.B.; Mayall, B.C.; Power, D.A. Probiotic treatment of vancomycin-resistant enterococci: A randomised controlled trial. Med. J. Aust. 2007, 186, 454–457. [Google Scholar] [CrossRef] [PubMed]
- Cheng, V.C.; Chen, J.H.; Tai, J.W.; Wong, S.C.; Poon, R.W.; Hung, I.F.; To, K.K.; Chan, J.F.; Ho, P.L.; Lo, C.M.; et al. Decolonization of gastrointestinal carriage of vancomycin-resistant Enterococcus faecium: Case series and review of literature. BMC Infect. Dis. 2014, 14, 514. [Google Scholar] [CrossRef]
- Buyukeren, M.; Yigit, S.; Buyukcam, A.; Kara, A.; Celik, H.T.; Yurdakok, M. A new use of Lactobacillus rhamnosus GG administration in the NICU: Colonized vancomycin-resistant enterococcus eradication in the gastrointestinal system. J. Matern. Fetal Neonatal Med. 2022, 35, 1192–1198. [Google Scholar] [CrossRef]
- Szachta, P.; Ignys, I.; Cichy, W. An Evaluation of the Ability of the Probiotic Strain Lactobacillus rhamnosus GG to Eliminate the Gastrointestinal Carrier State of Vancomycin-resistant Enterococci in Colonized Children. J. Clin. Gastroenterol. 2011, 45, 872–877. [Google Scholar] [CrossRef]
- Vidal, M.; Forestier, C.; Charbonnel, N.; Henard, S.; Rabaud, C.; Lesens, O. Probiotics and intestinal colonization by vancomycin-resistant enterococci in mice and humans. J. Clin. Microbiol. 2010, 48, 2595–2598. [Google Scholar] [CrossRef]
- Doron, S.; Hibberd Patricia, L.; Goldin, B.; Thorpe, C.; McDermott, L.; Snydman David, R. Effect of Lactobacillus rhamnosus GG Administration on Vancomycin-Resistant Enterococcus Colonization in Adults with Comorbidities. Antimicrob. Agents Chemother. 2015, 59, 4593–4599. [Google Scholar] [CrossRef] [PubMed]
- Rauseo, A.M.; Hink, T.; Reske, K.A.; Seiler, S.M.; Bommarito, K.M.; Fraser, V.J.; Burnham, C.D.; Dubberke, E.R. A randomized controlled trial of Lactobacillus rhamnosus GG on antimicrobial-resistant organism colonization. Infect. Control. Hosp. Epidemiol. 2022, 43, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Rubin, I.M.C.; Mollerup, S.; Broholm, C.; Knudsen, S.B.; Baker, A.; Helms, M.; Holm, M.K.A.; Kallemose, T.; Westh, H.; Dahl Knudsen, J.; et al. No Effect of Lactobacillus rhamnosus GG on Eradication of Colonization by Vancomycin-Resistant Enterococcus faecium or Microbiome Diversity in Hospitalized Adult Patients. Microbiol. Spectr. 2022, 10, e0234821. [Google Scholar] [CrossRef]
- de Regt, M.J.; Willems, R.J.; Hené, R.J.; Siersema, P.D.; Verhaar, H.J.; Hopmans, T.E.; Bonten, M.J. Effects of probiotics on acquisition and spread of multiresistant enterococci. Antimicrob. Agents Chemother. 2010, 54, 2801–2805. [Google Scholar] [CrossRef]
- Borgmann, S.; Rieß, B.; Siegmund, R.; Werner, G.; Klare, I. Treatment with Saccharomyces boulardii and Escherichia coli Nissle is safe and associated with reduced nosocomial transmission of van B vancomycin-resistant Enterococcus faecium on an early rehabilitation ward in Germany: A retrospective analysis. Ther. Clin. Risk Manag. 2019, 15, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Vanderhoof, J.A.; Young, R. Probiotics in the United States. Clin. Infect. Dis.Off. Publ. Infect. Dis. Soc. Am. 2008, 46 (Suppl. S2), S67–S72; discussion S144–S151. [Google Scholar] [CrossRef]
- Suez, J.; Zmora, N.; Segal, E.; Elinav, E. The pros, cons, and many unknowns of probiotics. Nat. Med. 2019, 25, 716–729. [Google Scholar] [CrossRef]
- Domínguez Rubio, A.P.; D’Antoni, C.L.; Piuri, M.; Pérez, O.E. Probiotics, Their Extracellular Vesicles and Infectious Diseases. Front. Microbiol. 2022, 13, 864720. [Google Scholar] [CrossRef]
- Ñahui Palomino, R.A.; Vanpouille, C.; Costantini, P.E.; Margolis, L. Microbiota–host communications: Bacterial extracellular vesicles as a common language. PLOS Pathog. 2021, 17, e1009508. [Google Scholar] [CrossRef] [PubMed]
- Merenstein, D.; Pot, B.; Leyer, G.; Ouwehand, A.C.; Preidis, G.A.; Elkins, C.A.; Hill, C.; Lewis, Z.T.; Shane, A.L.; Zmora, N.; et al. Emerging issues in probiotic safety: 2023 perspectives. Gut Microbes 2023, 15, 2185034. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.F.; Fung, Z.F.; Wu, C.L.; Chung, T.C. Molecular characterization of a plasmid-borne (pTC82) chloramphenicol resistance determinant (cat-TC) from Lactobacillus reuteriG4. Plasmid 1996, 36, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Tannock, G.W.; Luchansky, J.B.; Miller, L.; Connell, H.; Thode-Andersen, S.; Mercer, A.A.; Klaenhammer, T.R. Molecular characterization of a plasmid-borne (pGT633) erythromycin resistance determinant (ermGT) from Lactobacillus reuteri 100-63. Plasmid 1994, 31, 60–71. [Google Scholar] [CrossRef]
- van der Poll, T.; Shankar-Hari, M.; Wiersinga, W.J. The immunology of sepsis. Immunity 2021, 54, 2450–2464. [Google Scholar] [CrossRef]
- Liu, D.; Huang, S.Y.; Sun, J.H.; Zhang, H.C.; Cai, Q.L.; Gao, C.; Li, L.; Cao, J.; Xu, F.; Zhou, Y.; et al. Sepsis-induced immunosuppression: Mechanisms, diagnosis and current treatment options. Mil. Med. Res. 2022, 9, 56. [Google Scholar] [CrossRef]
- Roberts, M.B.; Fishman, J.A. Immunosuppressive Agents and Infectious Risk in Transplantation: Managing the “Net State of Immunosuppression”. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2021, 73, e1302–e1317. [Google Scholar] [CrossRef]
- Rodrigues, L.P.; Teixeira, V.R.; Alencar-Silva, T.; Simonassi-Paiva, B.; Pereira, R.W.; Pogue, R.; Carvalho, J.L. Hallmarks of aging and immunosenescence: Connecting the dots. Cytokine Growth Factor Rev. 2021, 59, 9–21. [Google Scholar] [CrossRef]
- Simons, A.; Alhanout, K.; Duval, R.E. Bacteriocins, Antimicrobial Peptides from Bacterial Origin: Overview of Their Biology and Their Impact against Multidrug-Resistant Bacteria. Microorganisms 2020, 8, 639. [Google Scholar] [CrossRef]
- Bierbaum, G.; Sahl, H.G. Lantibiotics: Mode of action, biosynthesis and bioengineering. Curr. Pharm. Biotechnol. 2009, 10, 2–18. [Google Scholar] [CrossRef]
- Sass, P.; Jansen, A.; Szekat, C.; Sass, V.; Sahl, H.-G.; Bierbaum, G. The lantibiotic mersacidin is a strong inducer of the cell wall stress response of Staphylococcus aureus. BMC Microbiol. 2008, 8, 186. [Google Scholar] [CrossRef] [PubMed]
- Gillor, O.; Kirkup, B.C.; Riley, M.A. Colicins and microcins: The next generation antimicrobials. Adv. Appl. Microbiol. 2004, 54, 129–146. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Björn, C.; Ekblom, J. Antimicrobial peptides as therapeutic agents: Opportunities and challenges. Crit. Rev. Biotechnol. 2020, 40, 978–992. [Google Scholar] [CrossRef] [PubMed]
| Organism | Clinical Scenario | Conventional Treatment Options |
|---|---|---|
| Extended-spectrum β-lactamase-producing Enterobacterales (ESBL-E) | Uncomplicated cystitis | Nitrofurantoin TMP-SMX * Ciprofloxacin Levofloxacin Carbapenem Single-dose aminoglycoside Fosfomycin (E. coli only) |
| Non-urinary tract | Meropenem Imipenem–cilastatin Ertapenem TMP-SMX (oral step-down) Ciprofloxacin (oral step-down) Levofloxacin (oral step-down) | |
| Carbapenem-resistant ESBL-E | Ceftazidime–avibactam Meropenem–vaborbactam Imipenem–cilastatin–relebactam Cefiderocol | |
| Amp-C β-lactamase-producing Enterobacterales | Cefepime MIC ** < 4 µg/mL | Cefepime |
| Cefepime MIC ≥ 4 µg/mL | Carbapenem (with demonstrated activity) | |
| Carbapenem-resistant Enterobacterales (CRE) | Uncomplicated cystitis | Nitrofurantoin TMP-SMX Ciprofloxacin Levofloxacin Ceftazidime–avibactam Meropenem–vaborbactam Imipenem–cilastatin–relebactam |
| Pyelonephritis/Complicated urinary tract infection | Nitrofurantoin TMP-SMX Ciprofloxacin Levofloxacin Ceftazidime–avibactam Meropenem–vaborbactam Imipenem–cilastatin–relebactam | |
| Non-urinary tract | Meropenem–vaborbactam Ceftazidime–avibactam Imipenem–cilastatin–relebactam Cefiderocol | |
| Multi-drug-resistant Pseudomonas aeruginosa | Traditional β-lactam susceptible | Fluoroquinolone Piperacillin–tazobactam Cefepime |
| Traditional β-lactam resistant | Ceftolozane–tazobactam Ceftazidime–avibactam Imipenem–cilastatin–relebactam Cefiderocol | |
| Carbapenem-resistant A. baumannii (CRAB) | All | High-dose ampicillin–sulbactam PLUS one of the following: Minocycline Tigecycline Polymyxin B Cefiderocol |
| Stenotrophomonas maltophilia | All | Ceftazidime–avibactam PLUS aztreonam OR any two of the following: TMP-SMX Minocycline Tigecycline Cefiderocol Levofloxacin |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jain, M.; Stitt, G.; Son, L.; Enioutina, E.Y. Probiotics and Their Bioproducts: A Promising Approach for Targeting Methicillin-Resistant Staphylococcus aureus and Vancomycin-Resistant Enterococcus. Microorganisms 2023, 11, 2393. https://doi.org/10.3390/microorganisms11102393
Jain M, Stitt G, Son L, Enioutina EY. Probiotics and Their Bioproducts: A Promising Approach for Targeting Methicillin-Resistant Staphylococcus aureus and Vancomycin-Resistant Enterococcus. Microorganisms. 2023; 11(10):2393. https://doi.org/10.3390/microorganisms11102393
Chicago/Turabian StyleJain, Manav, Gideon Stitt, Luke Son, and Elena Y. Enioutina. 2023. "Probiotics and Their Bioproducts: A Promising Approach for Targeting Methicillin-Resistant Staphylococcus aureus and Vancomycin-Resistant Enterococcus" Microorganisms 11, no. 10: 2393. https://doi.org/10.3390/microorganisms11102393
APA StyleJain, M., Stitt, G., Son, L., & Enioutina, E. Y. (2023). Probiotics and Their Bioproducts: A Promising Approach for Targeting Methicillin-Resistant Staphylococcus aureus and Vancomycin-Resistant Enterococcus. Microorganisms, 11(10), 2393. https://doi.org/10.3390/microorganisms11102393