

Journey of the Probiotic Bacteria: Survival of the Fittest
 , , and
, , and
Abstract
1. Probiotics: The Current State of an Ancient Practice
1.1. The Origin: Evolution of Probiotic Meaning

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Definition | References |
|---|---|---|
| Paraprobiotic | “Dead bacterial cells or cell components to denote health benefits beyond the inherent viability of probiotics.” | [20] |
| Postbiotic | Any substance released by or produced through the metabolic activity of the microorganism, which exerts a beneficial effect on the host, directly or indirectly. | [21] |
| Prebiotics | “Substrate that is selectively utilized by host microorganisms conferring a health benefit.” | [16] |
| Synbiotic | Mixture of prebiotics and probiotics for the improvement of human or animal health. | [22] |
1.2. Screening and Selection: The Search for the Best
1.3. Applications in Contemporary Times: The Search for “Live Long and Prosper”
1.4. The Market: Opportunities beyond the Big Players
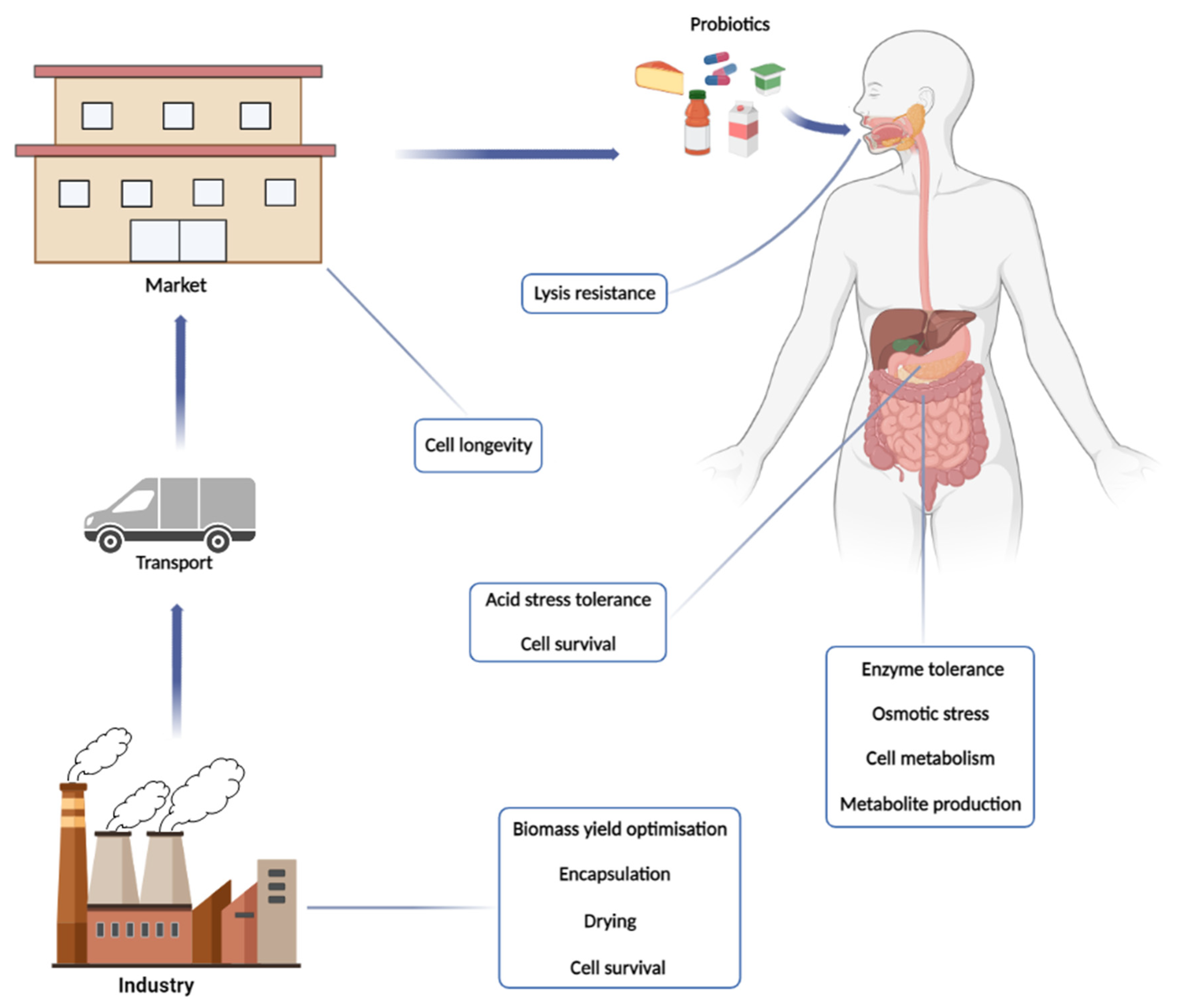
2. From the Physiology of Production to the Processing of the Product
2.1. Uptake of Sugars: The Sweety Diversity
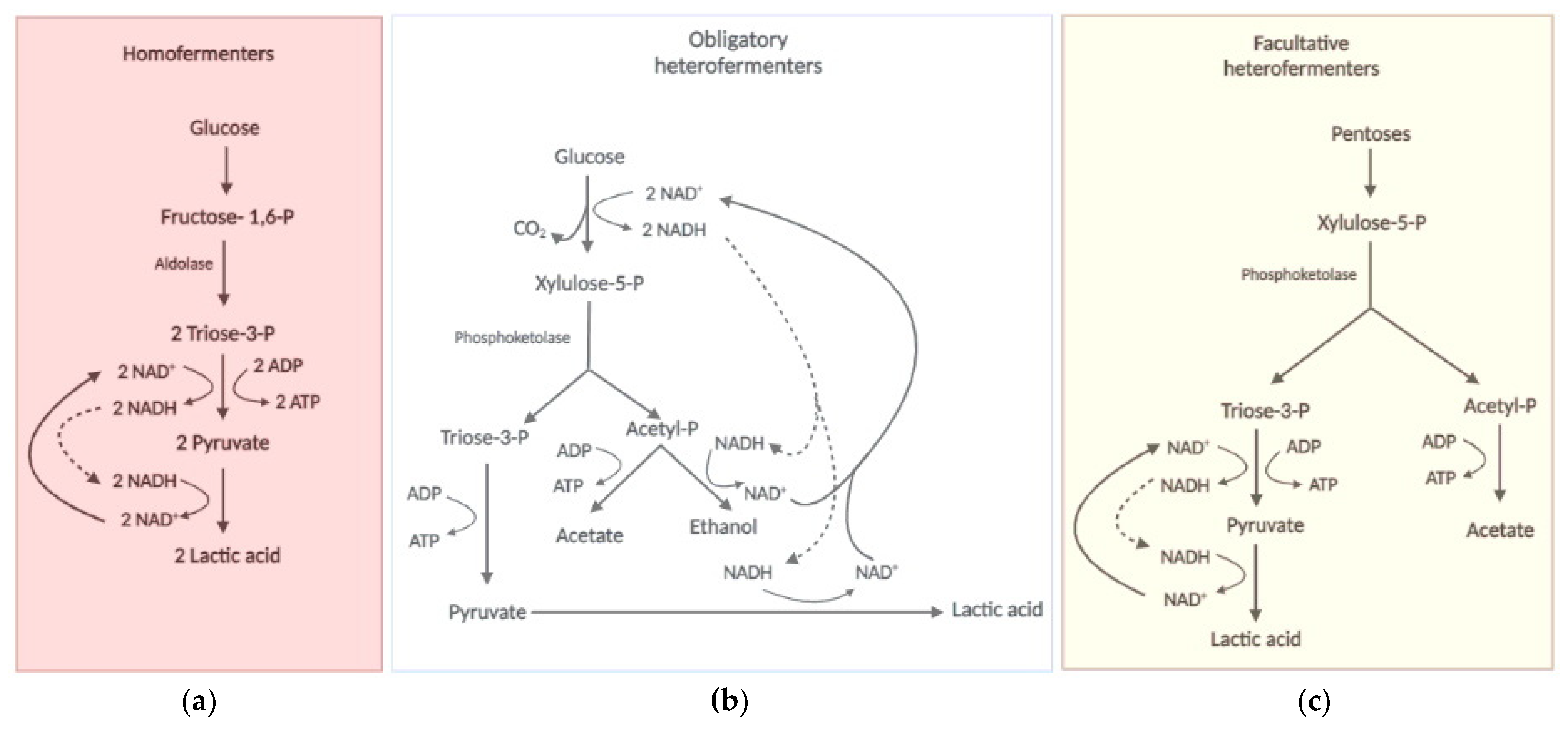
2.2. The LAB Biochemistry: Always Lactic, Sometimes Ethanoacetic
2.3. Processing Probiotic Cells: Dried but Not Dead
- (a)
- Salar-Behzadi et al. [181] microencapsulated B. bifidum BB-12 using Arabic gum, gelatine and pectin as protective agents, and observed partial protection of the microorganism from membrane damage caused by the drying process. In addition, the product maintained greater viability after one month’s storage.
- (b)
- Bora et al. [182] performed microencapsulation by lyophilization of L. acidophilus using whey and fructooligosaccharides (FOS) as protectors to enrich lyophilized banana powder. Besides the better protection of the cells, it did not interfere with the sensory characteristics of the banana powder.
- (c)
- Nunes et al. [183] used the spray-dryer in association with protective agents such as inulin, hi-maize® and trehalose to microencapsulate L. acidophilus LA-5. Trehalose protected the cells from GIT simulated conditions and extended the product stability beyond 120 days of storage. In addition, hi-maize® turned the cells more resistant to heat treatment.
- (d)
- Rosolen et al. [184] used spray-dryer, whey and inulin to microencapsulate L. lactis subsp. lactis R7 and observed a high level of cell protection against simulated GIT conditions and heat treatments.
- (e)
- Leylak et al. [185] carried out spray-dryer microencapsulation of Lactobacillus acidophilus LA-5 using powdered whey and Arabic gum as protective agents. The results obtained after exposure to simulated GIT conditions showed better survival capacity of the encapsulated microorganisms.
- (f)
- Minami et al. [186] used a method based on interfacial tension to produce a three-layered capsule composed by gelatin and pectin in the outmost layer and vegetable fats and oil in the middle layer to encapsulate B. brevis and B. longum cells. The results showed that B. brevis cells not only survived as they also grew when in the GIT of children.
- (g)
- Costa et al. [187] showed that encapsulated Bifidobacterium longum 51A for bacterial protection during atomisation. In addition, the cells were protected from the acidity of acerola pulp. The study concluded that acerola pulp is an interesting matrix for the production of probiotic fruit juice.
2.4. Probiotics Added to Food Matrices: A Tasty Way to Stay Healthy
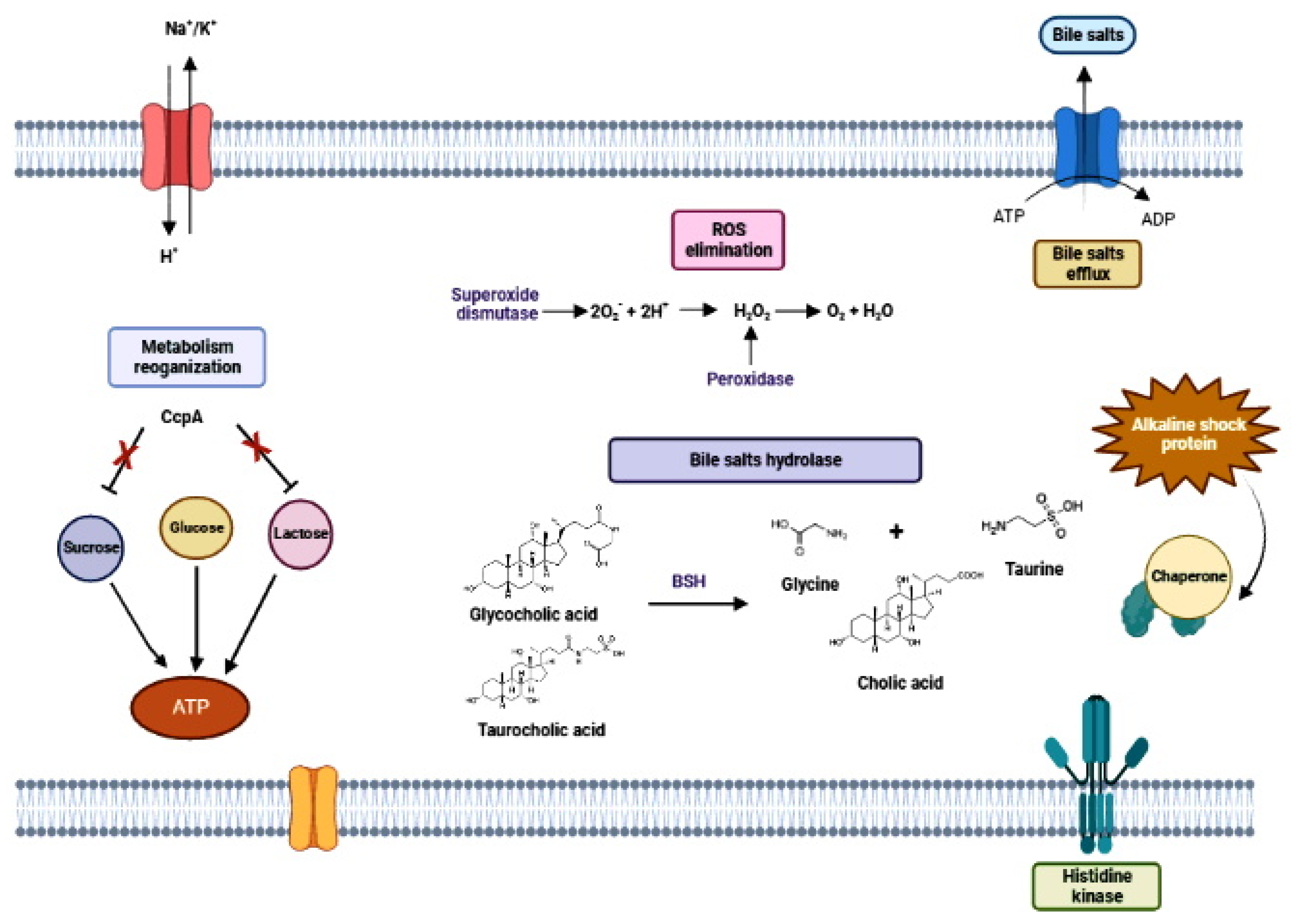
3. The Probiotic’s Passage through GIT: To the Mouth and Beyond
3.1. Overviewing the Metabolic Challenges of Probiotic Cells in the GIT
3.2. Oral Mucosa: The First Challenge at the Front Door
3.3. Stomach: The Acidic Battle Field
3.4. Intestine at Last: The Destination, but Still Full of Pitfalls
4. The End of the Journey and the Beginning of the Job
5. Microbiome and Faecal Transplant: The Future
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ozen, M.; Dinleyici, E.C. The History of Probiotics: The Untold Story. Benef. Microbes. 2015, 6, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Godoy, A.; Herrera, T.; Ulloa, M. Más Allá Del Pulque y El Tepache: Las Bebidas Alcohólicas No Destiladas Indígenas de México; 1a; Universidad Nacional Autonoma de México: Ciudad de México, Mexico, 2003. [Google Scholar]
- Nakazawa, Y.; Hosono, A. (Eds.) Functions of Fermented Milk: Challenges for the Health Sciences. In Fermented Milk in the Orient; Elsevier Applied Science: London, UK, 1992; pp. 61–78. [Google Scholar]
- Abou-Donia, S.A. Origin, History and Manufacturing Process of Egyptian Dairy Products: An Overview. Alex. J. Food Sci. Technol. 2008, 5, 51–62. [Google Scholar] [CrossRef]
- Tissier, H. Recherches Sur La Flore Intestinale Des Nourrissons: (État Normal et Pathologique); Méd.—Paris: Paris, France, 1900. [Google Scholar]
- Shokryazdan, P.; Faseleh Jahromi, M.; Liang, J.B.; Ho, Y.W. Probiotics: From Isolation to Application. J. Am. Coll. Nutr. 2017, 36, 666–676. [Google Scholar] [CrossRef]
- Lilly, D.M.; Stillwell, R.H. Probiotics: Growth-Promoting Factors Produced by Microorganisms. Science 1965, 147, 747–748. [Google Scholar] [CrossRef]
- Parker, R.B. Probiotics, the Other Half of the Antibiotic Story. Anim. Nutr. Health 1974, 29, 4–8. [Google Scholar]
- Fuller, R. Probiotics in Man and Animals. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar] [CrossRef]
- Gismondo, M.R.; Drago, L.; Lombardi, A. Review of Probiotics Available to Modify Gastrointestinal Flora. Int. J. Antimicrob. Agents 1999, 12, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Havenaar, R.; Huis In’t Veld, J.H.J. Probiotics: A General View. In The Lactic Acid Bacteria Volume 1; Springer US: Boston, MA, USA, 1992; pp. 151–170. [Google Scholar]
- Guarner, F. Probiotics. Int. J. Food Microbiol. 1998, 39, 237–238. [Google Scholar] [CrossRef]
- FAO; WHO. Guidelines for the Evaluation of Probiotics in Food; FAO: London, ON, Canada, 2002. [Google Scholar]
- NIH Probiotics: What You Need to Know. Available online: https://www.nccih.nih.gov/health/probiotics-what-you-need-to-know (accessed on 8 November 2022).
- Zendeboodi, F.; Khorshidian, N.; Mortazavian, A.M.; da Cruz, A.G. Probiotic: Conceptualization from a New Approach. Curr. Opin. Food Sci. 2020, 32, 103–123. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef]
- Silva, D.R.; Sardi, J.D.C.O.; Pitangui, N.D.S.; Roque, S.M.; da Silva, A.C.B.; Rosalen, P.L. Probiotics as an Alternative Antimicrobial Therapy: Current Reality and Future Directions. J. Funct. Foods 2020, 73, 104080. [Google Scholar] [CrossRef]
- Yadav, M.K.; Kumari, I.; Singh, B.; Sharma, K.K.; Tiwari, S.K. Probiotics, Prebiotics and Synbiotics: Safe Options for next-Generation Therapeutics. Appl. Microbiol. Biotechnol. 2022, 106, 505–521. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Lyu, Y.; Liu, X.; Jia, X.; Cui, F.; Wu, X.; Deng, S.; Yue, C. Engineered Probiotics. Microb. Cell Fact. 2022, 21, 72. [Google Scholar] [CrossRef]
- Cuevas-González, P.F.; Liceaga, A.M.; Aguilar-Toalá, J.E. Postbiotics and Paraprobiotics: From Concepts to Applications. Food Res. Int. 2020, 136, 109502. [Google Scholar] [CrossRef]
- Tsilingiri, K.; Rescigno, M. Postbiotics: What Else? Benef. Microbes 2013, 4, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Markowiak, P.; Śliżewska, K. Effects of Probiotics, Prebiotics, and Synbiotics on Human Health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.D.; Chu, M.; Oza, U.; Rajgarhia, V. The Value of Natural Products to Future Pharmaceutical Discovery. Nat. Prod. Rep. 2007, 24, 1225. [Google Scholar] [CrossRef]
- Bernardini, S.; Tiezzi, A.; Laghezza Masci, V.; Ovidi, E. Natural Products for Human Health: An Historical Overview of the Drug Discovery Approaches. Nat. Prod. Res. 2018, 32, 1926–1950. [Google Scholar] [CrossRef]
- Arnold, C. The New Danger of Synthetic Drugs. Lancet 2013, 382, 15–16. [Google Scholar] [CrossRef]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial Resistance: A Global Multifaceted Phenomenon. Pathog Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef]
- Trasande, L.; Shaffer, R.M.; Sathyanarayana, S.; Lowry, J.A.; Ahdoot, S.; Baum, C.R.; Bernstein, A.S.; Bole, A.; Campbell, C.C.; Landrigan, P.J.; et al. Food Additives and Child Health. Pediatrics 2018, 142, e20181410. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.; Holdbrooks, H.; Mishra, P.; Abrantes, M.A.; Eskew, S.; Garma, M.; Oca, C.-G.; McGuckin, C.; Hein, C.B.; Mitchell, R.D.; et al. Impact of Probiotic B. Infantis EVC001 Feeding in Premature Infants on the Gut Microbiome, Nosocomially Acquired Antibiotic Resistance, and Enteric Inflammation. Front. Pediatr. 2021, 9, 618009. [Google Scholar] [CrossRef] [PubMed]
- Sowden, M.; van Niekerk, E.; Bulabula, A.N.H.; Dramowski, A.; Whitelaw, A.; Twisk, J.; van Weissenbruch, M.M. Impact of a Multi-Strain Probiotic Administration on Peri-Rectal Colonization with Drug-Resistant Gram-Negative Bacteria in Preterm Neonates. Front. Pediatr. 2022, 10, 1002762. [Google Scholar] [CrossRef] [PubMed]
- Damaceno, Q.S.; Gallotti, B.; Reis, I.M.M.; Totte, Y.C.P.; Assis, G.B.; Figueiredo, H.C.; Silva, T.F.; Azevedo, V.; Nicoli, J.R.; Martins, F.S. Isolation and Identification of Potential Probiotic Bacteria from Human Milk. Probiotics Antimicrob. Proteins 2021. [Google Scholar] [CrossRef]
- Santos, D.D.S.; Calaça, P.R.D.A.; Porto, A.L.F.; de Souza, P.R.E.; de Freitas, N.S.A.; Soares, M.T.C.V. What Differentiates Probiotic from Pathogenic Bacteria? The Genetic Mobility of Enterococcus Faecium Offers New Molecular Insights. Omics 2020, 24, 706–713. [Google Scholar] [CrossRef]
- Bank, N.C.; Singh, V.; Rodriguez-Palacios, A. Classification of Parabacteroides Distasonis and Other Bacteroidetes Using O- Antigen Virulence Gene: RfbA -Typing and Hypothesis for Pathogenic vs. Probiotic Strain Differentiation. Gut Microbes 2022, 14, 1997293. [Google Scholar] [CrossRef]
- Asadi, A.; Lohrasbi, V.; Abdi, M.; Mirkalantari, S.; Esghaei, M.; Kashanian, M.; Oshaghi, M.; Talebi, M. The Probiotic Properties and Potential of Vaginal Lactobacillus Spp. Isolated from Healthy Women against Some Vaginal Pathogens. Lett. Appl. Microbiol. 2022, 74, 752–764. [Google Scholar] [CrossRef] [PubMed]
- Ouwehand, A.C.; Kirjavainen, P.v.; Shortt, C.; Salminen, S. Probiotics: Mechanisms and Established Effects. Int. Dairy J. 1999, 9, 43–52. [Google Scholar] [CrossRef]
- Lin, M.Y.; Chang, F.J. Antioxidative Effect of Intestinal Bacteria Bifidobacterium Longum ATCC 15708 and Lactobacillus Acidophilus ATCC 4356. Dig. Dis. Sci. 2000, 45, 1617–1622. [Google Scholar] [CrossRef]
- Pereira, D.I.A.; Gibson, G.R. Cholesterol Assimilation by Lactic Acid Bacteria and Bifidobacteria Isolated from the Human Gut. Appl. Environ. Microbiol. 2002, 68, 4689–4693. [Google Scholar] [CrossRef]
- Thirabunyanon, M.; Boonprasom, P.; Niamsup, P. Probiotic Potential of Lactic Acid Bacteria Isolated from Fermented Dairy Milks on Antiproliferation of Colon Cancer Cells. Biotechnol. Lett. 2009, 31, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Suez, J.; Zmora, N.; Segal, E.; Elinav, E. The Pros, Cons, and Many Unknowns of Probiotics. Nat. Med. 2019, 25, 716–729. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A Taxonomic Note on the Genus Lactobacillus: Description of 23 Novel Genera, Emended Description of the Genus Lactobacillus Beijerinck 1901, and Union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Probiotics Market Size, Share & Trends Analysis Report By Product (Probiotic Food & Beverages, Probiotic Dietary Supplements), By Ingredient (Bacteria, Yeast), By End Use, By Distribution Channel, And Segment Forecasts, 2021–2030. Available online: https://www.grandviewresearch.com/industry-analysis/probiotics-market/toc (accessed on 8 November 2022).
- Westermann, C.; Gleinser, M.; Corr, S.C.; Riedel, C.U. A Critical Evaluation of Bifidobacterial Adhesion to the Host Tissue. Front Microbiol. 2016, 7, 1220. [Google Scholar] [CrossRef]
- Ranadheera, C.; Vidanarachchi, J.; Rocha, R.; Cruz, A.; Ajlouni, S. Probiotic Delivery through Fermentation: Dairy vs. Non-Dairy Beverages. Fermentation 2017, 3, 67. [Google Scholar] [CrossRef]
- Souza, H.F.; Carosia, M.F.; Pinheiro, C.; de Carvalho, M.V.; de Oliveira, C.A.F.; Kamimura, E.S. On Probiotic Yeasts in Food Development: Saccharomyces Boularrdii, a Trend. Food Sci. Technol. 2022, 42. [Google Scholar] [CrossRef]
- George Kerry, R.; Patra, J.K.; Gouda, S.; Park, Y.; Shin, H.-S.; Das, G. Benefaction of Probiotics for Human Health: A Review. J. Food Drug Anal. 2018, 26, 927–939. [Google Scholar] [CrossRef]
- de Araújo Henriques Ferreira, G.; Magnani, M.; Cabral, L.; Brandão, L.R.; Noronha, M.F.; de Campos Cruz, J.; de Souza, E.L.; de Brito Alves, J.L. Potentially Probiotic Limosilactobacillus Fermentum Fruit-Derived Strains Alleviate Cardiometabolic Disorders and Gut Microbiota Impairment in Male Rats Fed a High-Fat Diet. Probiotics Antimicrob. Proteins 2022, 14, 349–359. [Google Scholar] [CrossRef]
- Moreira, C.F.; Cassini-Vieira, P.; Canesso, M.C.C.; Felipetto, M.; Ranfley, H.; Teixeira, M.M.; Nicoli, J.R.; Martins, F.S.; Barcelos, L.S. Lactobacillus Rhamnosus CGMCC 1.3724 (LPR) Improves Skin Wound Healing and Reduces Scar Formation in Mice. Probiotics Antimicrob. Proteins 2021, 13, 709–719. [Google Scholar] [CrossRef]
- Li, P.; Ji, B.; Luo, H.; Sundh, D.; Lorentzon, M.; Nielsen, J. One-Year Supplementation with Lactobacillus Reuteri ATCC PTA 6475 Counteracts a Degradation of Gut Microbiota in Older Women with Low Bone Mineral Density. NPJ Biofilms Microbiomes 2022, 8, 84. [Google Scholar] [CrossRef]
- Da Silva, T.F.; Casarotti, S.N.; De Oliveira, G.L.V.; Penna, A.L.V. The impact of probiotics, prebiotics, and synbiotics on the biochemical, clinical, and immunological markers, as well as on the gut microbiota of obese hosts. Crit. Rev. Food Sci. Nutr. 2021, 61, 337–355. [Google Scholar] [CrossRef] [PubMed]
- Dinkçi, N.; Akdeniz, V.; Akalin, A.S. Survival of Probiotics in Functional Foods during Shelf Life. In Food Quality and Shelf Life; Elsevier: Amsterdam, The Netherlands, 2019; pp. 201–233. [Google Scholar]
- Mushtaq, M.; Gani, A.; Masoodi, F.A. Himalayan Cheese (Kalari/Kradi) Fermented with Different Probiotic Strains: In Vitro Investigation of Nutraceutical Properties. LWT 2019, 104, 53–60. [Google Scholar] [CrossRef]
- Rolim, F.R.L.; Freitas Neto, O.C.; Oliveira, M.E.G.; Oliveira, C.J.B.; Queiroga, R.C.R.E. Cheeses as Food Matrixes for Probiotics: In Vitro and in Vivo Tests. Trends Food Sci. Technol. 2020, 100, 138–154. [Google Scholar] [CrossRef]
- Nematollahi, A.; Sohrabvandi, S.; Mortazavian, A.M.; Jazaeri, S. Viability of Probiotic Bacteria and Some Chemical and Sensory Characteristics in Cornelian Cherry Juice during Cold Storage. Electron. J. Biotechnol. 2016, 21, 49–53. [Google Scholar] [CrossRef]
- dos Santos Nascimento, D.; Sampaio, K.B.; do Nascimento, Y.M.; de Souza, T.A.; de Souza, F.S.; Júnior, J.V.C.; Tavares, J.F.; da Silva, M.S.; de Brito Alves, J.L.; de Souza, E.L. Evaluating the Stability of a Novel Nutraceutical Formulation Combining Probiotic Limosilactobacillus Fermentum 296, Quercetin, and Resveratrol Under Different Storage Conditions. Probiotics Antimicrob. Proteins 2022. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.E.; Yoon, L.S.; Michels, K.B.; Tranfield, W.; Jacobs, J.P.; May, F.P. The Impact of Prebiotic, Probiotic, and Synbiotic Supplements and Yogurt Consumption on the Risk of Colorectal Neoplasia among Adults: A Systematic Review. Nutrients 2022, 14, 4937. [Google Scholar] [CrossRef]
- Thangaleela, S.; Sivamaruthi, B.S.; Kesika, P.; Chaiyasut, C. Role of Probiotics and Diet in the Management of Neurological Diseases and Mood States: A Review. Microorganisms 2022, 10, 2268. [Google Scholar] [CrossRef]
- Wilmink, J.M.; Ladefoged, S.; Jongbloets, A.; Vernooij, J.C.M. The Evaluation of the Effect of Probiotics on the Healing of Equine Distal Limb Wounds. PLoS ONE 2020, 15, e0236761. [Google Scholar] [CrossRef]
- Albrecht, E.; Zitnan, R.; Karaffova, V.; Revajova, V.; Čechová, M.; Levkut, M., Jr.; Röntgen, M. Effects of the Probiotic Enterococcus Faecium on Muscle Characteristics of Chickens. Life 2022, 12, 1695. [Google Scholar] [CrossRef]
- Várhidi, Z.; Máté, M.; Ózsvári, L. The Use of Probiotics in Nutrition and Herd Health Management in Large Hungarian Dairy Cattle Farms. Front. Vet. Sci. 2022, 9, 957935. [Google Scholar] [CrossRef]
- de Oliveira, K.Á.R.; Fernandes, K.F.D.; de Souza, E.L. Current Advances on the Development and Application of Probiotic-Loaded Edible Films and Coatings for the Bioprotection of Fresh and Minimally Processed Fruit and Vegetables. Foods 2021, 10, 2207. [Google Scholar] [CrossRef] [PubMed]
- Foligné, B.; Daniel, C.; Pot, B. Probiotics from Research to Market: The Possibilities, Risks and Challenges. Curr. Opin. Microbiol. 2013, 16, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, M.; Oomah, B.D.; Oliveira, W.P.; Burgos-Díaz, C.; Rubilar, M.; Shene, C. Probiotics and Prebiotics Potential for the Care of Skin, Female Urogenital Tract, and Respiratory Tract. Folia Microbiol. 2020, 65, 245–264. [Google Scholar] [CrossRef] [PubMed]
- Mordor Intelligence Probiotics Market–Growth, Trends, and Forecasts (2022–2027). Available online: https://www.mordorintelligence.com/industry-reports/probiotics-market (accessed on 9 November 2022).
- Granato, D.; Branco, G.F.; Nazzaro, F.; Cruz, A.G.; Faria, J.A.F. Functional Foods and Nondairy Probiotic Food Development: Trends, Concepts, and Products. Compr. Rev. Food Sci. Food Saf. 2010, 9, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Ranadheera, R.D.C.S.; Baines, S.K.; Adams, M.C. Importance of Food in Probiotic Efficacy. Food Res. Int. 2010, 43, 1–7. [Google Scholar] [CrossRef]
- Banić, M.; Uroić, K.; Leboš Pavunc, A.; Novak, J.; Zorić, K.; Durgo, K.; Petković, H.; Jamnik, P.; Kazazić, S.; Kazazić, S.; et al. Characterization of S-Layer Proteins of Potential Probiotic Starter Culture Lactobacillus Brevis SF9B Isolated from Sauerkraut. LWT 2018, 93, 257–267. [Google Scholar] [CrossRef]
- Sperry, M.F.; Silva, H.L.A.; Balthazar, C.F.; Esmerino, E.A.; Verruck, S.; Prudencio, E.S.; Neto, R.P.C.; Tavares, M.I.B.; Peixoto, J.C.; Nazzaro, F.; et al. Probiotic Minas Frescal Cheese Added with L. Casei 01: Physicochemical and Bioactivity Characterization and Effects on Hematological/Biochemical Parameters of Hypertensive Overweighted Women–A Randomized Double-Blind Pilot Trial. J. Funct. Foods 2018, 45, 435–443. [Google Scholar] [CrossRef]
- Télessy, I.G. Nutraceuticals. In The Role of Functional Food Security in Global Health; Singh, R.B., Watson, R.R., Takahashi, T., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 409–421. [Google Scholar]
- Bellikci-Koyu, E.; Sarer-Yurekli, B.P.; Karagozlu, C.; Aydin-Kose, F.; Ozgen, A.G.; Buyuktuncer, Z. Probiotic Kefir Consumption Improves Serum Apolipoprotein A1 Levels in Metabolic Syndrome Patients: A Randomized Controlled Clinical Trial. Nutr. Res. 2022, 102, 59–70. [Google Scholar] [CrossRef]
- Markets and Markets Probiotics Market by Product Type (Functional Food & Beverages, Dietary Supplements, and Feed), Ingredient (Bacteria and Yeast), End User (Human and Animal), Distribution Channel, and Region–Global Forecast to 2027. Available online: https://www.marketresearch.com/MarketsandMarkets-v3719/Probiotics-Product-Type-Functional-Food-32586440/ (accessed on 9 November 2022).
- Brazil Probiotics Market–Forecasts from 2020 to 2025. Available online: https://www.researchandmarkets.com/r/r1axqw (accessed on 9 November 2022).
- Grumet, L.; Tromp, Y.; Stiegelbauer, V. The Development of High-Quality Multispecies Probiotic Formulations: From Bench to Market. Nutrients 2020, 12, 2453. [Google Scholar] [CrossRef]
- Zawistowska-Rojek, A.; Zaręba, T.; Tyski, S. Microbiological Testing of Probiotic Preparations. Int. J. Environ. Res. Public Health 2022, 19, 5701. [Google Scholar] [CrossRef]
- Zotta, T.; Ricciardi, A.; Ianniello, R.G.; Parente, E.; Reale, A.; Rossi, F.; Iacumin, L.; Comi, G.; Coppola, R. Assessment of aerobic and respiratory growth in the Lactobacillus casei group. PLoS ONE 2014, 9, e99189. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. Where in the World Do Bacteria Experience Oxidative Stress? Env. Microbiol. 2019, 21, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Zotta, T.; Parente, E.; Ricciardi, A. Aerobic Metabolism in the Genus Lactobacillus: Impact on Stress Response and Potential Applications in the Food Industry. J. Appl. Microbiol. 2017, 122, 857–869. [Google Scholar] [CrossRef]
- Zotta, T.; Ricciardi, A.; Ianniello, R.G.; Storti, L.v.; Glibota, N.A.; Parente, E. Aerobic and Respirative Growth of Heterofermentative Lactic Acid Bacteria: A Screening Study. Food Microbiol. 2018, 76, 117–127. [Google Scholar] [CrossRef]
- Schiraldi, C.; Adduci, V.; Valli, V.; Maresca, C.; Giuliano, M.; Lamberti, M.; Cartenì, M.; de Rosa, M. High Cell Density Cultivation of Probiotics and Lactic Acid Production. Biotechnol. Bioeng. 2003, 82, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, M.B.; Gaudu, P.; Lechardeur, D.; Petit, M.-A.; Gruss, A. Aerobic Respiration Metabolism in Lactic Acid Bacteria and Uses in Biotechnology. Annu. Rev. Food Sci. Technol. 2012, 3, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, M.; Mayo, B.; Fernández, M.; Aleksandrzak-Piekarczyk, T. Updates on Metabolism in Lactic Acid Bacteria in Light of “Omic” Technologies. In Biotechnology of Lactic Acid Bacteria; Mozzi, F., Raya, R.R., Vignolo, G.M., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2015; pp. 1–24. ISBN 9781118868386. [Google Scholar]
- Zúñiga, M.; Monedero, V.; Yebra, M.J. Utilization of Host-Derived Glycans by Intestinal Lactobacillus and Bifidobacterium Species. Front. Microbiol. 2018, 9, 1917. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Calasso, M.; Cavallo, N.; Di Cagno, R.; Gobbetti, M. Functional Proteomics within the Genus Lactobacillus. Proteomics 2016, 16, 946–962. [Google Scholar] [CrossRef]
- Tsujikawa, Y.; Ishikawa, S.; Sakane, I.; Yoshida, K.; Osawa, R. Identification of Genes Encoding a Novel ABC Transporter in Lactobacillus Delbrueckii for Inulin Polymers Uptake. Sci. Rep. 2021, 11, 16007. [Google Scholar] [CrossRef]
- Jeckelmann, J.-M.; Erni, B. Transporters of Glucose and Other Carbohydrates in Bacteria. Pflug. Arch. 2020, 472, 1129–1153. [Google Scholar] [CrossRef]
- Andersen, J.M.; Barrangou, R.; Hachem, M.A.; Lahtinen, S.J.; Goh, Y.J.; Svensson, B.; Klaenhammer, T.R. Transcriptional Analysis of Oligosaccharide Utilization by Bifidobacterium LactisBl-04. BMC Genom. 2013, 14, 312. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Ma, Q.; Chen, X.; Wang, H.; Zhu, J.; Lee, Y.; Zhang, H.; Zhao, J.; Lu, W.; Chen, W. An α Type Gluco -Oligosaccharide from Brown Algae Laminaria Japonica Stimulated the Growth of Lactic Acid Bacteria Encoding Specific ABC Transport System Components. Food Funct. 2022, 13, 11153–11168. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, M.; Bardowski, J. Regulation of Sugar Catabolism in Lactococcus lactis. Crit. Rev. Microbiol. 2007, 33, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Duar, R.M.; Casaburi, G.; Mitchell, R.D.; Scofield, L.N.C.; Ortega Ramirez, C.A.; Barile, D.; Henrick, B.M.; Frese, S.A. Comparative Genome Analysis of Bifidobacterium Longum Subsp. Infantis Strains Reveals Variation in Human Milk Oligosaccharide Utilization Genes among Commercial Probiotics. Nutrients 2020, 12, 3247. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Gänzle, M.G. Genetic and Phenotypic Analysis of Carbohydrate Metabolism and Transport in Lactobacillus Reuteri. Int. J. Food Microbiol. 2018, 272, 12–21. [Google Scholar] [CrossRef]
- Pfeiler, E.A.; Klaenhammer, T.R. Role of Transporter Proteins in Bile Tolerance of Lactobacillus acidophilus. Appl. Env. Microbiol. 2009, 75, 6013–6016. [Google Scholar] [CrossRef]
- Zhang, W.; Sun, Z.; Wu, R.; Meng, H.; Zhang, H. Comparative Genome Analysis of a New Probiotic Strain Lactobacillus Casei Zhang. In Genomics II-Bacteria, Viruses and Metabolic Pathways; Iconcept Press, Ed.; Createspace: Scotts Valley, CA, USA, 2013; pp. 276–296. ISBN 978-1-4802-5414-5. [Google Scholar]
- Arsköld, E.; Lohmeier-Vogel, E.; Cao, R.; Roos, S.; Rådström, P.; van Niel, E.W.J. Phosphoketolase Pathway Dominates in Lactobacillus Reuteri ATCC 55730 Containing Dual Pathways for Glycolysis. J. Bacteriol. 2008, 190, 206–212. [Google Scholar] [CrossRef]
- Gänzle, M.G.; Follador, R. Metabolism of Oligosaccharides and Starch in Lactobacilli: A Review. Front. Microbiol. 2012, 3, 1–15. [Google Scholar] [CrossRef]
- Zúñiga, M.; Yebra, M.J.; Monedero, V. Complex Oligosaccharide Utilization Pathways in Lactobacillus. Curr. Issues Mol. Biol. 2021, 40, 49–80. [Google Scholar] [CrossRef]
- Nishimura, J. Review Probiotic Characteristics and Carbohydrate Metabolism of Lactobacillus Reuteri. Milk Sci. 2020, 69, 71–82. [Google Scholar] [CrossRef]
- Barrangou, R.; Azcarate-Peril, M.A.; Duong, T.; Conners, S.B.; Kelly, R.M.; Klaenhammer, T.R. Global Analysis of Carbohydrate Utilization by Lactobacillus Acidophilus Using CDNA Microarrays. Proc. Natl. Acad. Sci. USA 2006, 103, 3816–3821. [Google Scholar] [CrossRef]
- Gänzle, M.G. Lactic Metabolism Revisited: Metabolism of Lactic Acid Bacteria in Food Fermentations and Food Spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Li, R.; Luo, W.; Liu, Y.; Chen, C.; Chen, S.; Yang, J.; Wu, P.; Lv, X.; Liu, Z.; Ni, L.; et al. The Investigation on the Characteristic Metabolites of Lactobacillus Plantarum RLL68 during Fermentation of Beverage from By-Products of Black Tea Manufacture. Curr. Res. Food Sci. 2022, 5, 1320–1329. [Google Scholar] [CrossRef]
- Stincone, A.; Prigione, A.; Cramer, T.; Wamelink, M.M.C.; Campbell, K.; Cheung, E.; Olin-Sandoval, V.; Grüning, N.; Krüger, A.; Tauqeer Alam, M.; et al. The Return of Metabolism: Biochemistry and Physiology of the Pentose Phosphate Pathway. Biol. Rev. 2015, 90, 927–963. [Google Scholar] [CrossRef]
- Cui, Y.; Qu, X. Genetic Mechanisms of Prebiotic Carbohydrate Metabolism in Lactic Acid Bacteria: Emphasis on Lacticaseibacillus Casei and Lacticaseibacillus Paracasei as Flexible, Diverse and Outstanding Prebiotic Carbohydrate Starters. Trends Food Sci. Technol. 2021, 115, 486–499. [Google Scholar] [CrossRef]
- Hatti-Kaul, R.; Chen, L.; Dishisha, T.; Enshasy, H.E. El Lactic Acid Bacteria: From Starter Cultures to Producers of Chemicals. FEMS Microbiol. Lett. 2018, 365, 1–20. [Google Scholar] [CrossRef]
- Doi, Y. Glycerol Metabolism and Its Regulation in Lactic Acid Bacteria. Appl. Microbiol. Biotechnol. 2019, 103, 5079–5093. [Google Scholar] [CrossRef]
- Laëtitia, G.; Pascal, D.; Yann, D. The Citrate Metabolism in Homo- and Heterofermentative LAB: A Selective Means of Becoming Dominant over Other Microorganisms in Complex Ecosystems. Food Nutr. Sci. 2014, 5, 953–969. [Google Scholar] [CrossRef]
- Viana, R.; Pérez-Martínez, G.; Deutscher, J.; Monedero, V. The Glycolytic Genes Pfk and Pyk from Lactobacillus Casei Are Induced by Sugars Transported by the Phosphoenolpyruvate:Sugar Phosphotransferase System and Repressed by CcpA. Arch. Microbiol. 2005, 183, 385–393. [Google Scholar] [CrossRef]
- Tanaka, K.; Komiyama, A.; Sonomoto, K.; Ishizaki, A.; Hall, S.J.; Stanbury, P.F. Phosphoketolase Pathway Dominates in Lactobacillus Reuteri ATCC 55730 Containing Dual Pathways for Glycolysis. Appl. Microbiol. Biotechnol. 2002, 60, 160–167. [Google Scholar] [CrossRef]
- Okano, K.; Yoshida, S.; Yamada, R.; Tanaka, T.; Ogino, C.; Fukuda, H.; Kondo, A. Improved Production of Homo-D-Lactic Acid via Xylose Fermentation by Introduction of Xylose Assimilation Genes and Redirection of the Phosphoketolase Pathway to the Pentose Phosphate Pathway in L-Lactate Dehydrogenase Gene-Deficient Lactobacillus Plantaru. Appl. Environ. Microbiol. 2009, 75, 7858–7861. [Google Scholar] [CrossRef] [PubMed]
- Basen, M.; Kurrer, S.E. A Close Look at Pentose Metabolism of Gut Bacteria. FEBS J. 2021, 288, 1804–1808. [Google Scholar] [CrossRef]
- Lee, K.; Lee, H.-G.; Pi, K.; Choi, Y.-J. The Effect of Low PH on Protein Expression by the Probiotic BacteriumLactobacillus Reuteri. Proteomics 2008, 8, 1624–1630. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Wu, M.; Wang, X.; Xu, L.; Zheng, X. Effect of Different Polymerized Xylooligosaccharides on the Metabolic Pathway in Bifidobacterium Adolescentis. J. Food Qual. 2022, 2022, 4412324. [Google Scholar] [CrossRef]
- Zhao, J.; Cheung, P.C.K. Comparative Proteome Analysis of Bif Idobacterium Longum Subsp. Infantis Grown on β–Glucans from Di Ff Erent Sources and a Model for Their Utilization. J. Agric. Food Chem. 2013, 61, 4360–4370. [Google Scholar] [CrossRef]
- Gänzle, M.G.; Vermeulen, N.; Vogel, R.F. Carbohydrate, Peptide and Lipid Metabolism of Lactic Acid Bacteria in Sourdough. Food Microbiol. 2007, 24, 128–138. [Google Scholar] [CrossRef]
- da Silva, P.K.N.P.K.N.; Mendonça, A.A.A.A.; de Miranda, A.R.A.R.; Calazans, T.L.S.T.L.S.; de Souza, R.B.R.B.; de Morais, M.A.M.A. Nutritional Requirements for Lactobacillus Vini Growth in Sugarcane Derivative Substrate of Ethanol Fermentation. FEMS Microbiol. Lett. 2019, 366, 1–9. [Google Scholar] [CrossRef]
- Yadav, A.K.; Chaudhari, A.B.; Kothari, R.M. Bioconversion of Renewable Resources into Lactic Acid: An Industrial View. Crit Rev. Biotechnol. 2011, 31, 1–19. [Google Scholar] [CrossRef]
- Duar, R.M.; Lin, X.B.; Zheng, J.; Martino, M.E.; Grenier, T.; Pérez-Muñoz, M.E.; Leulier, F.; Gänzle, M.; Walter, J. Lifestyles in Transition: Evolution and Natural History of the Genus Lactobacillus. FEMS Microbiol. Rev. 2017, 41, S27–S48. [Google Scholar] [CrossRef]
- Rodas, A.M.; Chenoll, E.; Macián, M.C.; Ferrer, S.; Pardo, I.; Aznar, R. Lactobacillus Vini Sp. Nov., a Wine Lactic Acid Bacterium Homofermentative for Pentoses. Int. J. Syst. Evol. Microbiol. 2006, 56, 513–517. [Google Scholar] [CrossRef]
- Endo, A.; Salminen, S. Honeybees and Beehives Are Rich Sources for Fructophilic Lactic Acid Bacteria. Syst. Appl. Microbiol. 2013, 36, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Maeno, S.; Tanizawa, Y.; Kanesaki, Y.; Kubota, E.; Kumar, H.; Dicks, L.; Salminen, S.; Nakagawa, J.; Arita, M.; Endo, A. Genomic Characterization of a Fructophilic Bee Symbiont Lactobacillus Kunkeei Reveals Its Niche-Specific Adaptation. Syst. Appl. Microbiol. 2016, 39, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Goh, Y.J.; Klaenhammer, T.R. A Functional Glycogen Biosynthesis Pathway in L Actobacillus Acidophilus: Expression and Analysis of the Glg Operon. Mol. Microbiol. 2013, 89, 1187–1200. [Google Scholar] [CrossRef]
- Goh, Y.J.; Klaenhammer, T.R. Insights into Glycogen Metabolism in Lactobacillus Acidophilus: Impact on Carbohydrate Metabolism, Stress Tolerance and Gut Retention. Microb. Cell Fact. 2014, 13, 94. [Google Scholar] [CrossRef] [PubMed]
- Misra, G.; Rojas, E.R.; Gopinathan, A.; Huang, K.C. Mechanical Consequences of Cell-Wall Turnover in the Elongation of a Gram-Positive Bacterium. Biophys. J. 2013, 104, 2342–2352. [Google Scholar] [CrossRef]
- Chapot-Chartier, M.-P.; Kulakauskas, S. Cell Wall Structure and Function in Lactic Acid Bacteria. Microb. Cell Fact. 2014, 13 (Suppl. S1), S9. [Google Scholar] [CrossRef]
- Scheffers, D.-J.; Pinho, M.G. Bacterial Cell Wall Synthesis: New Insights from Localization Studies. Microbiol. Mol. Biol. Rev. 2005, 69, 585–607. [Google Scholar] [CrossRef]
- Jeanloz, R.W. The Chemical Structure of the Cell Wall of Gram-Positive Bacteria. Pure Appl. Chem. 1967, 14, 57–70. [Google Scholar] [CrossRef]
- Mendonça, A.A.; da Silva, P.K.N.; Calazans, T.L.S.; de Souza, R.B.; Pita, W.d.B.; Elsztein, C.; de Morais Junior, M.A. Lactobacillus Vini: Mechanistic Response to Stress by Medium Acidification. Microbiology 2019, 165, 26–36. [Google Scholar] [CrossRef]
- Vinderola, G.; Mocchiutti, P.; Reinheimer, J.A. Interactions among lactic acid starter and probiotic bacteria used for fermented dairy products. J. Dairy Sci. 2020, 85, 721–729. [Google Scholar] [CrossRef]
- van de Guchte, M.; Serror, P.; Chervaux, C.; Smokvina, T.; Ehrlich, S.D.; Maguin, E. Stress Responses in Lactic Acid Bacteria. Antonie Van Leeuwenhoek 2002, 82, 187–216. [Google Scholar] [CrossRef] [PubMed]
- Mendonça, A.A.; da Silva, P.K.N.; Calazans, T.L.S.; de Souza, R.B.; Elsztein, C.; de Morais Junior, M.A. Gene Regulation of the Lactobacillus Vini in Response to Industrial Stress in the Fuel Ethanol Production. Microbiol. Res. 2020, 236, 126450. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Yang, P.; Yang, J.; Zhang, J. Comparative Transcriptome Analysis Reveals the Contribution of Membrane Transporters to Acid Tolerance in Lactococcus Lactis. J. Biotechnol. 2022, 357, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Nicolai, B.M.; van Impe, J.F.; Verlinden, B.; Martens, T.; Vandewalle, J.; de Baerdemaeker, J. Predictive Modelling of Surface Growth of Lactic Acid Bacteria in Vacuum-Packed Meat. Food Microbiol. 1993, 10, 229–238. [Google Scholar] [CrossRef]
- Vereecken, K.M.; van Impe, J.F. Analysis and Practical Implementation of a Model for Combined Growth and Metabolite Production of Lactic Acid Bacteria. Int. J. Food Microbiol. 2002, 73, 239–250. [Google Scholar] [CrossRef]
- Hathi, Z.; Mettu, S.; Priya, A.; Athukoralalage, S.; Lam, T.N.; Choudhury, N.R.; Dutta, N.K.; El-Omar, E.M.; Gong, L.; Mohan, G.; et al. Methodological Advances and Challenges in Probiotic Bacteria Production: Ongoing Strategies and Future Perspectives. Biochem. Eng. J. 2021, 176, 108199. [Google Scholar] [CrossRef]
- Fu, N.; Chen, X.D. Towards a Maximal Cell Survival in Convective Thermal Drying Processes. Food Res. Int. 2011, 44, 1127–1149. [Google Scholar] [CrossRef]
- Bansal, T.; Garg, S. Probiotics: From Functional Foods to Pharmaceutical Products. Curr. Pharm. Biotechnol. 2008, 9, 267–287. [Google Scholar] [CrossRef]
- Brachkova, M.I.; Duarte, A.; Pinto, J.F. Evaluation of the Viability of Lactobacillus Spp. after the Production of Different Solid Dosage Forms. J. Pharm. Sci. 2009, 98, 3329–3339. [Google Scholar] [CrossRef]
- Govender, M.; Choonara, Y.E.; Kumar, P.; du Toit, L.C.; van Vuuren, S.; Pillay, V. A Review of the Advancements in Probiotic Delivery: Conventional vs. Non-Conventional Formulations for Intestinal Flora Supplementation. AAPS PharmSciTech 2014, 15, 29–43. [Google Scholar] [CrossRef]
- Coman, M.M.; Cecchini, C.; Verdenelli, M.C.; Silvi, S.; Orpianesi, C.; Cresci, A. Functional Foods as Carriers for SYNBIO®, a Probiotic Bacteria Combination. Int. J. Food Microbiol. 2012, 157, 346–352. [Google Scholar] [CrossRef]
- Dimitrellou, D.; Tsaousi, K.; Kourkoutas, Y.; Panas, P.; Kanellaki, M.; Koutinas, A.A. Fermentation Efficiency of Thermally Dried Immobilized Kefir on Casein as Starter Culture. Process Biochem. 2008, 43, 1323–1329. [Google Scholar] [CrossRef]
- Gardiner, G.E.; Bouchier, P.; O’Sullivan, E.; Kelly, J.; Kevin Collins, J.; Fitzgerald, G.; Paul Ross, R.; Stanton, C. A Spray-Dried Culture for Probiotic Cheddar Cheese Manufacture. Int. Dairy J. 2002, 12, 749–756. [Google Scholar] [CrossRef]
- Melin, P.; Sundh, I.; Håkansson, S.; Schnürer, J. Biological Preservation of Plant Derived Animal Feed with Antifungal Microorganisms: Safety and Formulation Aspects. Biotechnol. Lett. 2007, 29, 1147–1154. [Google Scholar] [CrossRef] [PubMed]
- Bommasamudram, J.; Muthu, A.; Devappa, S. Effect of Sub-Lethal Heat Stress on Viability of Lacticaseibacillus Casei N in Spray-Dried Powders. LWT 2022, 155, 112904. [Google Scholar] [CrossRef]
- Huang, S.; Vignolles, M.-L.; Chen, X.D.; le Loir, Y.; Jan, G.; Schuck, P.; Jeantet, R. Spray Drying of Probiotics and Other Food-Grade Bacteria: A Review. Trends Food Sci. Technol. 2017, 63, 1–17. [Google Scholar] [CrossRef]
- da Silva Guedes, J.; Pimentel, T.C.; Diniz-Silva, H.T.; Tayse da Cruz Almeida, E.; Tavares, J.F.; Leite de Souza, E.; Garcia, E.F.; Magnani, M. Protective Effects of β-Glucan Extracted from Spent Brewer Yeast during Freeze-Drying, Storage and Exposure to Simulated Gastrointestinal Conditions of Probiotic Lactobacilli. LWT 2019, 116, 108496. [Google Scholar] [CrossRef]
- Archacka, M.; Celińska, E.; Białas, W. Techno-Economic Analysis for Probiotics Preparation Production Using Optimized Corn Flour Medium and Spray-Drying Protective Blends. Food Bioprod. Process. 2020, 123, 354–366. [Google Scholar] [CrossRef]
- Depaz, R.A.; Pansare, S.; Patel, S.M. Freeze-Drying Above the Glass Transition Temperature in Amorphous Protein Formulations While Maintaining Product Quality and Improving Process Efficiency. J. Pharm. Sci. 2016, 105, 40–49. [Google Scholar] [CrossRef]
- Madhu, A.N.; Awasthi, S.P.; Bhasker, K.; Reddy, P.K.; Gurudutt Prapulla, S. Impact of Freeze and Spray Drying on the Retention of Probiotic Properties of Lactobacillus Fermentum: An in Vitro Evaluation Model. Int. J. Microbiol. Res. 2011, 2, 243–251. [Google Scholar]
- Bjelošević, M.; Seljak, K.B.; Trstenjak, U.; Logar, M.; Brus, B.; Ahlin Grabnar, P. Aggressive Conditions during Primary Drying as a Contemporary Approach to Optimise Freeze-Drying Cycles of Biopharmaceuticals. Eur. J. Pharm. Sci. 2018, 122, 292–302. [Google Scholar] [CrossRef]
- Wang, G.-Q.; Pu, J.; Yu, X.-Q.; Xia, Y.-J.; Ai, L.-Z. Influence of Freezing Temperature before Freeze-Drying on the Viability of Various Lactobacillus Plantarum Strains. J. Dairy Sci. 2020, 103, 3066–3075. [Google Scholar] [CrossRef]
- Akbarbaglu, Z.; Peighambardoust, S.H.; Sarabandi, K.; Jafari, S.M. Spray Drying Encapsulation of Bioactive Compounds within Protein-Based Carriers; Different Options and Applications. Food Chem. 2021, 359, 129965. [Google Scholar] [CrossRef]
- Ranadheera, C.S.; Evans, C.A.; Adams, M.C.; Baines, S.K. In Vitro Analysis of Gastrointestinal Tolerance and Intestinal Cell Adhesion of Probiotics in Goat’s Milk Ice Cream and Yogurt. Food Res. Int. 2012, 49, 619–625. [Google Scholar] [CrossRef]
- Béal, C.; Fonseca, F. Freezing of Probiotic Bacteria. In Advances in Probiotic Technology; CRC Press: Boca Raton, FL, USA, 2015; pp. 179–212. [Google Scholar]
- Tyutkov, N.; Zhernyakova, A.; Birchenko, A.; Eminova, E.; Nadtochii, L.; Baranenko, D. Probiotics Viability in Frozen Food Products. Food Biosci. 2022, 50, 101996. [Google Scholar] [CrossRef]
- Cayley, S.; Lewis, B.A.; Guttman, H.J.; Record, M.T. Characterization of the Cytoplasm of Escherichia Coli K-12 as a Function of External Osmolarity. J. Mol. Biol. 1991, 222, 281–300. [Google Scholar] [CrossRef]
- Beumer, R.R.; te Giffel, M.C.; Cox, L.J.; Rombouts, F.M.; Abee, T. Effect of Exogenous Proline, Betaine, and Carnitine on Growth of Listeria Monocytogenes in a Minimal Medium. Appl. Environ. Microbiol. 1994, 60, 1359–1363. [Google Scholar] [CrossRef] [PubMed]
- Galinski, E.A. Osmoadaptation in Bacteria. Adv. Microb. Physiol. 1995, 37, 272–328. [Google Scholar] [PubMed]
- Assadpour, E.; Jafari, S.M. Advances in Spray-Drying Encapsulation of Food Bioactive Ingredients: From Microcapsules to Nanocapsules. Annu. Rev. Food Sci. Technol. 2019, 10, 103–131. [Google Scholar] [CrossRef] [PubMed]
- Salama, A.H. Spray Drying as an Advantageous Strategy for Enhancing Pharmaceuticals Bioavailability. Drug Deliv. Transl. Res. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mendonsa, N.; Almutairy, B.; Kallakunta, V.R.; Sarabu, S.; Thipsay, P.; Bandari, S.; Repka, M.A. Manufacturing Strategies to Develop Amorphous Solid Dispersions: An Overview. J. Drug Deliv. Sci. Technol. 2020, 55, 101459. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, K.; Abdullah, M.; Wuled Lenggoro, I.; Iskandar, F. Preparation of Functional Nanostructured Particles by Spray Drying. Adv. Powder Technol. 2006, 17, 587–611. [Google Scholar] [CrossRef]
- Murugesan, R.; Orsat, V. Spray Drying for the Production of Nutraceutical Ingredients—A Review. Food Bioproc. Technol. 2012, 5, 3–14. [Google Scholar] [CrossRef]
- Hlaing, M.M.; Wood, B.R.; McNaughton, D.; Ying, D.; Dumsday, G.; Augustin, M.A. Effect of Drying Methods on Protein and DNA Conformation Changes in Lactobacillus Rhamnosus GG Cells by Fourier Transform Infrared Spectroscopy. J. Agric. Food Chem. 2017, 65, 1724–1731. [Google Scholar] [CrossRef] [PubMed]
- Rodklongtan, A.; Nitisinprasert, S.; Chitprasert, P. Antioxidant Activity and the Survival-Enhancing Effect of Ascorbic Acid on Limosilactobacillus Reuteri KUB-AC5 Microencapsulated with Lactose by Spray Drying. LWT 2022, 164, 113645. [Google Scholar] [CrossRef]
- Alpert, P. The Limits and Frontiers of Desiccation-Tolerant Life. Integr. Comp. Biol. 2005, 45, 685–695. [Google Scholar] [CrossRef]
- Kim, S.X.; Çamdere, G.; Hu, X.; Koshland, D.; Tapia, H. Synergy between the Small Intrinsically Disordered Protein Hsp12 and Trehalose Sustain Viability after Severe Desiccation. Elife 2018, 7, e38337. [Google Scholar] [CrossRef]
- Potts, M. Desiccation Tolerance: A Simple Process? Trends Microbiol. 2001, 9, 553–559. [Google Scholar] [CrossRef]
- Greffe, V.R.G.; Michiels, J. Desiccation-Induced Cell Damage in Bacteria and the Relevance for Inoculant Production. Appl. Microbiol. Biotechnol. 2020, 104, 3757–3770. [Google Scholar] [CrossRef]
- Laskowska, E.; Kuczyńska-Wiśnik, D. New Insight into the Mechanisms Protecting Bacteria during Desiccation. Curr. Genet. 2020, 66, 313–318. [Google Scholar] [CrossRef]
- Schimel, J.; Balser, T.C.; Wallenstein, M. Microbial stress-response physiology and its implications for ecosystem function. Ecology 2007, 88, 1386–1394. [Google Scholar] [CrossRef] [PubMed]
- Hecker, M.; Völker, U. General Stress Response of Bacillus Subtilis and Other Bacteria. Adv. Microb. Physiol. 2001, 44, 35–91. [Google Scholar] [PubMed]
- Winther, K.S.; Roghanian, M.; Gerdes, K. Activation of the Stringent Response by Loading of RelA-TRNA Complexes at the Ribosomal A-Site. Mol. Cell 2018, 70, 95–105.e4. [Google Scholar] [CrossRef] [PubMed]
- Burgain, J.; Gaiani, C.; Linder, M.; Scher, J. Encapsulation of Probiotic Living Cells: From Laboratory Scale to Industrial Applications. J. Food Eng. 2011, 104, 467–483. [Google Scholar] [CrossRef]
- Rama, G.R.; Führ, A.J.; da Silva, J.A.B.S.; Gennari, A.; Giroldi, M.; Goettert, M.I.; Volken de Souza, C.F. Encapsulation of Lactobacillus Spp. Using Bovine and Buffalo Cheese Whey and Their Application in Orange Juice. 3 Biotech 2020, 10, 263. [Google Scholar] [CrossRef] [PubMed]
- Rajam, R.; Subramanian, P. Encapsulation of Probiotics: Past, Present and Future. Beni Suef. Univ. J. Basic Appl. Sci. 2022, 11, 46. [Google Scholar] [CrossRef]
- Reque, P.M.; Brandelli, A. Encapsulation of Probiotics and Nutraceuticals: Applications in Functional Food Industry. Trends Food Sci. Technol. 2021, 114, 1–10. [Google Scholar] [CrossRef]
- Abd El-salam, M.H.; El-Shibiny, S. Formation and Potential Uses of Milk Proteins as Nano Delivery Vehicles for Nutraceuticals: A Review. Int. J. Dairy Technol. 2012, 65, 13–21. [Google Scholar] [CrossRef]
- Nualkaekul, S.; Cook, M.T.; Khutoryanskiy, V.v.; Charalampopoulos, D. Influence of Encapsulation and Coating Materials on the Survival of Lactobacillus Plantarum and Bifidobacterium Longum in Fruit Juices. Food Res. Int. 2013, 53, 304–311. [Google Scholar] [CrossRef]
- Yoha, K.S.; Moses, J.A.; Anandharamakrishnan, C. Effect of Encapsulation Methods on the Physicochemical Properties and the Stability of Lactobacillus Plantarum (NCIM 2083) in Synbiotic Powders and in-Vitro Digestion Conditions. J. Food Eng. 2020, 283, 110033. [Google Scholar] [CrossRef]
- Liu, H.; Gong, J.; Chabot, D.; Miller, S.S.; Cui, S.W.; Zhong, F.; Wang, Q. Improved Survival of Lactobacillus Zeae LB1 in a Spray Dried Alginate-Protein Matrix. Food Hydrocoll. 2018, 78, 100–108. [Google Scholar] [CrossRef]
- Tao, T.; Ding, Z.; Hou, D.; Prakash, S.; Zhao, Y.; Fan, Z.; Zhang, D.; Wang, Z.; Liu, M.; Han, J. Influence of Polysaccharide as Co-Encapsulant on Powder Characteristics, Survival and Viability of Microencapsulated Lactobacillus Paracasei Lpc-37 by Spray Drying. J. Food Eng. 2019, 252, 10–17. [Google Scholar] [CrossRef]
- Quirós-Sauceda, A.E.; Ayala-Zavala, J.F.; Olivas, G.I.; González-Aguilar, G.A. Edible Coatings as Encapsulating Matrices for Bioactive Compounds: A Review. J. Food Sci. Technol. 2014, 51, 1674–1685. [Google Scholar] [CrossRef]
- Kowalska, E.; Ziarno, M.; Ekielski, A.; Żelaziński, T. Materials Used for the Microencapsulation of Probiotic Bacteria in the Food Industry. Molecules 2022, 27, 3321. [Google Scholar] [CrossRef]
- Chen, M.-J.; Chen, K.-N. Applications of Probiotic Encapsulation in Dairy Products. In Encapsulation and Controlled Release Technologies in Food Systems; Blackwell Publishing: Ames, IA, USA, 2007; pp. 83–112. [Google Scholar]
- Salar-Behzadi, S.; Wu, S.; Toegel, S.; Hofrichter, M.; Altenburger, I.; Unger, F.M.; Wirth, M.; Viernstein, H. Impact of Heat Treatment and Spray Drying on Cellular Properties and Culturability of Bifidobacterium Bifidum BB-12. Food Res. Int. 2013, 54, 93–101. [Google Scholar] [CrossRef]
- Massounga Bora, A.F.; Li, X.; Zhu, Y.; Du, L. Improved Viability of Microencapsulated Probiotics in a Freeze-Dried Banana Powder During Storage and Under Simulated Gastrointestinal Tract. Probiotics Antimicrob. Proteins 2019, 11, 1330–1339. [Google Scholar] [CrossRef] [PubMed]
- Nunes, G.L.; Etchepare, M.d.A.; Cichoski, A.J.; Zepka, L.Q.; Jacob Lopes, E.; Barin, J.S.; Flores, É.M.d.M.; da Silva, C.d.B.; de Menezes, C.R. Inulin, Hi-Maize, and Trehalose as Thermal Protectants for Increasing Viability of Lactobacillus Acidophilus Encapsulated by Spray Drying. LWT 2018, 89, 128–133. [Google Scholar] [CrossRef]
- Rosolen, M.D.; Bordini, F.W.; de Oliveira, P.D.; Conceição, F.R.; Pohndorf, R.S.; Fiorentini, Â.M.; da Silva, W.P.; Pieniz, S. Symbiotic Microencapsulation of Lactococcus Lactis Subsp. Lactis R7 Using Whey and Inulin by Spray Drying. LWT 2019, 115, 108411. [Google Scholar] [CrossRef]
- Leylak, C.; Özdemir, K.S.; Gurakan, G.C.; Ogel, Z.B. Optimisation of Spray Drying Parameters for Lactobacillus Acidophilus Encapsulation in Whey and Gum Arabic: Its Application in Yoghurt. Int. Dairy J. 2021, 112, 104865. [Google Scholar] [CrossRef]
- Minami, M.; Tsuji, S.; Akagawa, S.; Akagawa, Y.; Yoshimoto, Y.; Kawakami, H.; Kohno, M.; Kaneko, K. Effect of a Bifidobacterium-Containing Acid-Resistant Microcapsule Formulation on Gut Microbiota: A Pilot Study. Nutrients 2022, 14, 4829. [Google Scholar] [CrossRef]
- Costa, K.; Silva, L.S.; Kobori, C.N.; Silva, A.M.; Nicoli, J.R. Microencapsulation of Bifidobacterium Longum 5 1A Cells by Spray Drying and Its Incorporation in Acerola (Malpighia emarginata) Pulp Powder. Int. J. Food Sci. Technol. 2022, 57, 323–329. [Google Scholar] [CrossRef]
- Boza-Mendez, E.; Lopez-Calvo, R.; Cortes-Munoz, M. Innovative Dairy Products Development Using Probiotics: Challenges and Limitations. In Probiotics; InTech: London, UK, 2012. [Google Scholar]
- Lollo, P.C.B.; Cruz, A.G.; Morato, P.N.; Moura, C.S.; Carvalho-Silva, L.B.; Oliveira, C.A.F.; Faria, J.A.F.; Amaya-Farfan, J. Probiotic Cheese Attenuates Exercise-Induced Immune Suppression in Wistar Rats. J. Dairy Sci. 2012, 95, 3549–3558. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, B.F.; Alves, J.L.; Belo, G.A.; Oliveira, E.R.; Braga, M.P.; da Silva, S.H.; Lemos, L.; Guimarães, J.T.; Silva, R.; Rocha, R.S.; et al. Therapeutic Effects of Probiotic Minas Frescal Cheese on the Attenuation of Ulcerative Colitis in a Murine Model. Front. Microbiol. 2021, 12, 623920. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gonzalez, C.; Gibson, T.; Jauregi, P. Novel Probiotic-Fermented Milk with Angiotensin I-Converting Enzyme Inhibitory Peptides Produced by Bifidobacterium Bifidum MF 20/5. Int. J. Food Microbiol. 2013, 167, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Moroti, C.; Souza Magri, L.F.; de Rezende Costa, M.; Cavallini, D.C.; Sivieri, K. Effect of the Consumption of a New Symbiotic Shake on Glycemia and Cholesterol Levels in Elderly People with Type 2 Diabetes Mellitus. Lipids Health Dis. 2012, 11, 29. [Google Scholar] [CrossRef]
- Vesterlund, S.; Salminen, K.; Salminen, S. Water Activity in Dry Foods Containing Live Probiotic Bacteria Should Be Carefully Considered: A Case Study with Lactobacillus Rhamnosus GG in Flaxseed. Int. J. Food Microbiol. 2012, 157, 319–321. [Google Scholar] [CrossRef]
- Terpou, A.; Papadaki, A.; Lappa, I.; Kachrimanidou, V.; Bosnea, L.; Kopsahelis, N. Probiotics in Food Systems: Significance and Emerging Strategies Towards Improved Viability and Delivery of Enhanced Beneficial Value. Nutrients 2019, 11, 1591. [Google Scholar] [CrossRef]
- Dantas, A.; Verruck, S.; Machado Canella, M.H.; Maran, B.M.; Murakami, F.S.; de Avila Junior, L.B.; de Campos, C.E.M.; Hernandez, E.; Prudencio, E.S. Current Knowledge about Physical Properties of Innovative Probiotic Spray-Dried Powders Produced with Lactose-Free Milk and Prebiotics. LWT 2021, 151, 112175. [Google Scholar] [CrossRef]
- Fenster, K.; Freeburg, B.; Hollard, C.; Wong, C.; Rønhave Laursen, R.; Ouwehand, A. The Production and Delivery of Probiotics: A Review of a Practical Approach. Microorganisms 2019, 7, 83. [Google Scholar] [CrossRef]
- Serrazanetti, D.I.; Guerzoni, M.E.; Corsetti, A.; Vogel, R. Metabolic Impact and Potential Exploitation of the Stress Reactions in Lactobacilli. Food Microbiol. 2009, 26, 700–711. [Google Scholar] [CrossRef]
- da Cruz, A.G.; Faria, J.d.A.F.; van Dender, A.G.F. Packaging System and Probiotic Dairy Foods. Food Res. Int. 2007, 40, 951–956. [Google Scholar] [CrossRef]
- Khosravi Zanjani, M.A.; Ghiassi Tarzi, B.; Sharifan, A.; Mohammadi, N. Microencapsulation of Probiotics by Calcium Alginate-Gelatinized Starch with Chitosan Coating and Evaluation of Survival in Simulated Human Gastro-Intestinal Condition. Iran J. Pharm. Res. 2014, 13, 843–852. [Google Scholar] [PubMed]
- Meng, J.; Wang, Y.-Y.; Hao, Y.-P. Protective Function of Surface Layer Protein from Lactobacillus Casei Fb05 against Intestinal Pathogens in Vitro. Biochem. Biophys. Res. Commun. 2021, 546, 15–20. [Google Scholar] [CrossRef]
- Hengge, R. Linking Bacterial Growth, Survival, and Multicellularity–Small Signaling Molecules as Triggers and Drivers. Curr. Opin. Microbiol. 2020, 55, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-K.; Guevarra, R.B.; Kim, Y.-T.; Kwon, J.; Kim, H.; Cho, J.H.; Kim, H.B.; Lee, J.-H. Role of Probiotics in Human Gut Microbiome-Associated Diseases. J. Microbiol. Biotechnol. 2019, 29, 1335–1340. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Vanderleyden, J.; de Keersmaecker, S.C.J. Genes and Molecules of Lactobacilli Supporting Probiotic Action. Microbiol. Mol. Biol. Rev. 2008, 72, 728–764. [Google Scholar] [CrossRef] [PubMed]
- Walter, J. Ecological Role of Lactobacilli in the Gastrointestinal Tract: Implications for Fundamental and Biomedical Research. Appl. Environ. Microbiol. 2008, 74, 4985–4996. [Google Scholar] [CrossRef]
- Remus, D.M.; Kleerebezem, M.; Bron, P. A An Intimate Tête-à-Tête–How Probiotic Lactobacilli Communicate with the Host. Eur. J. Pharm. 2011, 668, S33–S42. [Google Scholar] [CrossRef]
- Manzoor, S.; Wani, S.M.; Ahmad Mir, S.; Rizwan, D. Role of Probiotics and Prebiotics in Mitigation of Different Diseases. Nutrition 2022, 96, 111602. [Google Scholar] [CrossRef]
- Jana, U.K.; Suryawanshi, R.K.; Prajapati, B.P.; Kango, N. Prebiotic Mannooligosaccharides: Synthesis, Characterization and Bioactive Properties. Food Chem. 2021, 342, 128328. [Google Scholar] [CrossRef]
- Thongaram, T.; Hoeflinger, J.L.; Chow, J.; Miller, M.J. Prebiotic Galactooligosaccharide Metabolism by Probiotic Lactobacilli and Bifidobacteria. J. Agric. Food Chem. 2017, 65, 4184–4192. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Fratianni, F.; Nicolaus, B.; Poli, A.; Orlando, P. The Prebiotic Source Influences the Growth, Biochemical Features and Survival under Simulated Gastrointestinal Conditions of the Probiotic Lactobacillus Acidophilus. Anaerobe 2012, 18, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Pan, T.; Xiang, H.; Diao, T.; Ma, W.; Shi, C.; Xu, Y.; Xie, Q. Effects of Probiotics and Nutrients Addition on the Microbial Community and Fermentation Quality of Peanut Hull. Bioresour. Technol. 2019, 273, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Chen, Y.; Zou, X.; Li, Q.; Li, H.; Shu, Y.; Li, X.; Li, W.; Han, L.; Ge, C. Salivary Secretory Immunoglobulin (SIgA) and Lysozyme in Malignant Tumor Patients. Biomed. Res. Int. 2016, 2016, 8701423. [Google Scholar] [CrossRef]
- Callewaert, L.; Michiels, C.W. Lysozymes in the Animal Kingdom. J. Biosci. 2010, 35, 127–160. [Google Scholar] [CrossRef]
- Ogundele, M.O. Inhibitors of Complement Activity in Human Breast-Milk: A Proposed Hypothesis of Their Physiological Significance. Mediat. Inflamm. 1999, 8, 69–75. [Google Scholar] [CrossRef]
- Derde, M.; Lechevalier, V.; Guérin-Dubiard, C.; Cochet, M.-F.; Jan, S.; Baron, F.; Gautier, M.; Vié, V.; Nau, F. Hen Egg White Lysozyme Permeabilizes Escherichia Coli Outer and Inner Membranes. J. Agric. Food Chem. 2013, 61, 9922–9929. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, H.R.; Matsuzaki, T.; Aoki, T. Genetic Evidence That Antibacterial Activity of Lysozyme Is Independent of Its Catalytic Function. FEBS Lett. 2001, 506, 27–32. [Google Scholar] [CrossRef]
- Ferraboschi, P.; Ciceri, S.; Grisenti, P. Applications of Lysozyme, an Innate Immune Defense Factor, as an Alternative Antibiotic. Antibiotics 2021, 10, 1534. [Google Scholar] [CrossRef]
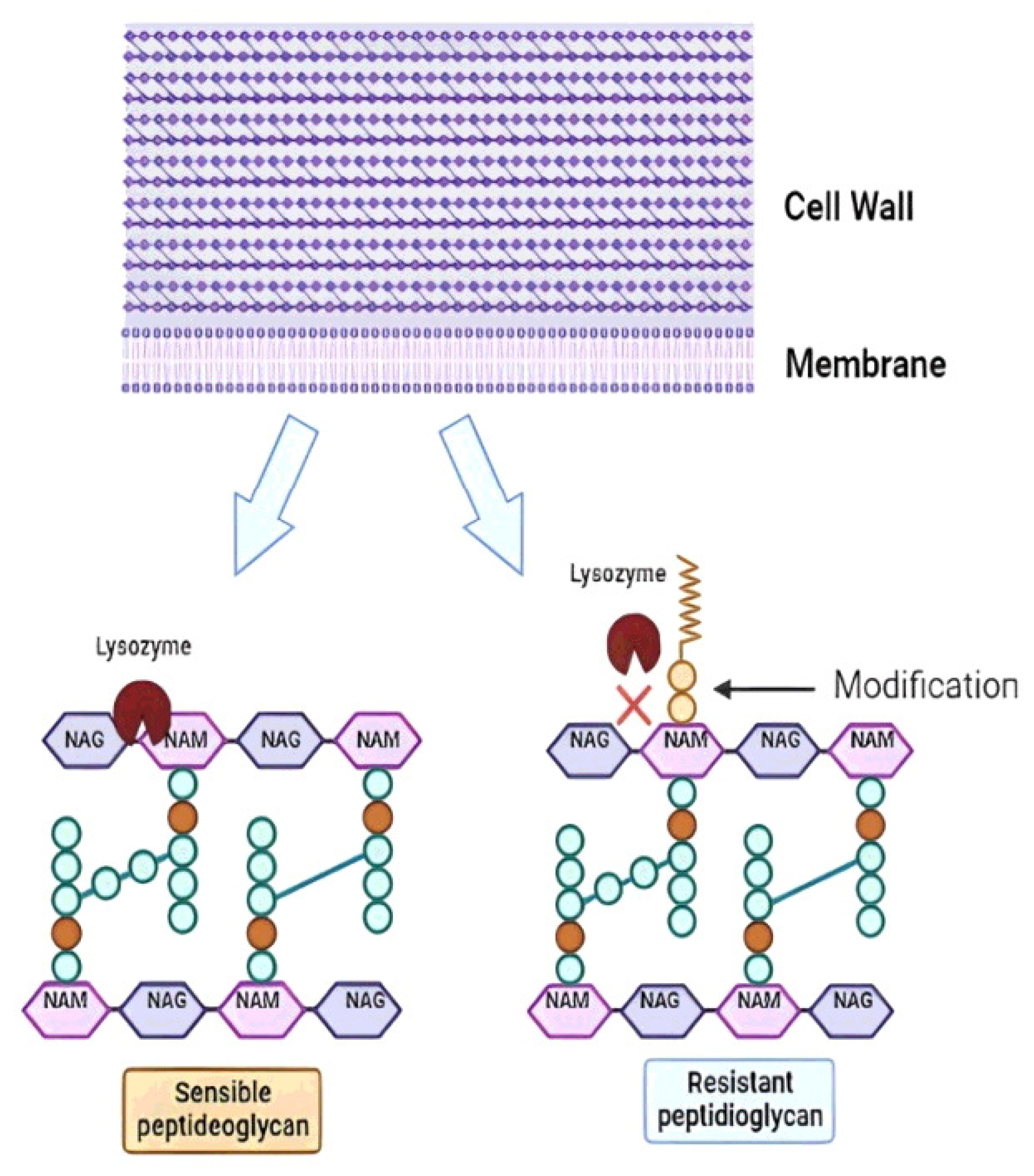
- Bera, A.; Biswas, R.; Herbert, S.; Kulauzovic, E.; Weidenmaier, C.; Peschel, A.; Götz, F. Influence of Wall Teichoic Acid on Lysozyme Resistance in Staphylococcus Aureus. J. Bacteriol. 2007, 189, 280–283. [Google Scholar] [CrossRef]
- Veiga, P.; Bulbarela-Sampieri, C.; Furlan, S.; Maisons, A.; Chapot-Chartier, M.P.; Erkelenz, M.; Mervelet, P.; Noirot, P.; Frees, D.; Kuipers, O.P.; et al. SpxB Regulates O-Acetylation-Dependent Resistance of Lactococcus Lactis Peptidoglycan to Hydrolysis. J. Biol. Chem. 2007, 282, 19342–19354. [Google Scholar] [CrossRef]
- Girardin, S.E.; Boneca, I.G.; Viala, J.; Chamaillard, M.; Labigne, A.; Thomas, G.; Philpott, D.J.; Sansonetti, P.J. Nod2 Is a General Sensor of Peptidoglycan through Muramyl Dipeptide (MDP) Detection. J. Biol. Chem. 2003, 278, 8869–8872. [Google Scholar] [CrossRef] [PubMed]
- Rada, V.; Splichal, I.; Rockova, S.; Grmanova, M.; Vlkova, E. Susceptibility of Bifidobacteria to Lysozyme as a Possible Selection Criterion for Probiotic Bifidobacterial Strains. Biotechnol. Lett. 2010, 32, 451–455. [Google Scholar] [CrossRef]
- Inayah, I.; Wibowo, M.S.; Julianti, E.; Suciati, T. Characterization of Lactobacillus Zeae as Probiotic and Starter Culture for Tamarillo Fermented Product. Food Sci. Technol. 2022, 42, e54021. [Google Scholar] [CrossRef]
- Papadimitriou, K.; Alegría, Á.; Bron, P.A.; de Angelis, M.; Gobbetti, M.; Kleerebezem, M.; Lemos, J.A.; Linares, D.M.; Ross, P.; Stanton, C.; et al. Stress Physiology of Lactic Acid Bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 837–890. [Google Scholar] [CrossRef] [PubMed]
- Piuri, M.; Sanchez-Rivas, C.; Ruzal, S.M. Cell Wall Modifications during Osmotic Stress in Lactobacillus Casei. J. Appl. Microbiol. 2005, 98, 84–95. [Google Scholar] [CrossRef]
- Freeman, H.J.; Kim, Y.S. Digestion and Absorption of Protein. Annu. Rev. Med. 1978, 29, 99–116. [Google Scholar] [CrossRef] [PubMed]
- Martinsen; Fossmark; Waldum The Phylogeny and Biological Function of Gastric Juice—Microbiological Consequences of Removing Gastric Acid. Int. J. Mol. Sci. 2019, 20, 6031. [CrossRef]
- Heda, R.; Toro, F.; Tombazzi, C.R. Physiology, Pepsin; StatPearls Publishing: Treasure Island, FL, USA, 2022; Available online: https://www.ncbi.nlm.nih.gov/books/NBK537005/ (accessed on 12 November 2022).
- Ajuebor, J.; McAuliffe, O.; O’Mahony, J.; Ross, R.P.; Hill, C.; Coffey, A. Bacteriophage Endolysins and Their Applications. Sci. Prog. 2016, 99, 183–199. [Google Scholar] [CrossRef]
- Tokatlı, M.; Gülgör, G.; Bağder Elmacı, S.; Arslankoz İşleyen, N.; Özçelik, F. In Vitro Properties of Potential Probiotic Indigenous Lactic Acid Bacteria Originating from Traditional Pickles. Biomed. Res. Int. 2015, 2015, 315819. [Google Scholar] [CrossRef]
- Perczyk, P.; Broniatowski, M. Simultaneous Action of Microbial Phospholipase C and Lipase on Model Bacterial Membranes–Modeling the Processes Crucial for Bioaugmentation. Biochim. Biophys. Acta (BBA)-Biomembr. 2021, 1863, 183620. [Google Scholar] [CrossRef] [PubMed]
- Šeme, H.; Gjuračić, K.; Kos, B.; Fujs, Š.; Štempelj, M.; Petković, H.; Šušković, J.; Bogovič Matijašić, B.; Kosec, G. Acid Resistance and Response to PH-Induced Stress in Two Lactobacillus plantarum Strains with Probiotic Potential. Benef. Microbes 2015, 6, 369–379. [Google Scholar] [CrossRef]
- Wang, R.-M.; Li, N.; Zheng, K.; Hao, J.-F. Enhancing Acid Tolerance of the Probiotic Bacterium Lactobacillus Acidophilus NCFM with Trehalose. FEMS Microbiol. Lett. 2018, 365, fny217. [Google Scholar] [CrossRef]
- Bernatek, M.; Żukiewicz-Sobczak, W.; Lachowicz-Wiśniewska, S.; Piątek, J. Factors Determining Effective Probiotic Activity: Evaluation of Survival and Antibacterial Activity of Selected Probiotic Products Using an “In Vitro” Study. Nutrients 2022, 14, 3323. [Google Scholar] [CrossRef]
- Wu, C.; Zhang, J.; Chen, W.; Wang, M.; Du, G.; Chen, J. A Combined Physiological and Proteomic Approach to Reveal Lactic-Acid-Induced Alterations in Lactobacillus Casei Zhang and Its Mutant with Enhanced Lactic Acid Tolerance. Appl. Microbiol. Biotechnol. 2012, 93, 707–722. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, B.M.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Survival of Probiotic Lactobacilli in Acidic Environments Is Enhanced in the Presence of Metabolizable Sugars. Appl. Environ. Microbiol. 2005, 71, 3060–3067. [Google Scholar] [CrossRef]
- Trček, J.; Mira, N.P.; Jarboe, L.R. Adaptation and Tolerance of Bacteria against Acetic Acid. Appl. Microbiol. Biotechnol. 2015, 99, 6215–6229. [Google Scholar] [CrossRef] [PubMed]
- Geng, P.; Zhang, L.; Shi, G.Y. Omics Analysis of Acetic Acid Tolerance in Saccharomyces Cerevisiae. World J. Microbiol. Biotechnol. 2017, 33, 94. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Dopson, M. Life in Acid: PH Homeostasis in Acidophiles. Trends Microbiol. 2007, 15, 165–171. [Google Scholar] [CrossRef]
- Jain, P.; Jain, V.; Singh, A.; Chauhan, A.; Sinha, S. Evaluation on the Responses of Succinate Dehydrogenase, Isocitrate Dehydrogenase, Malate Dehydrogenase and Glucose-6-Phosphate Dehydrogenase to Acid Shock Generated Acid Tolerance in Escherichia Coli. Adv. Biomed. Res. 2013, 2, 75. [Google Scholar] [CrossRef]
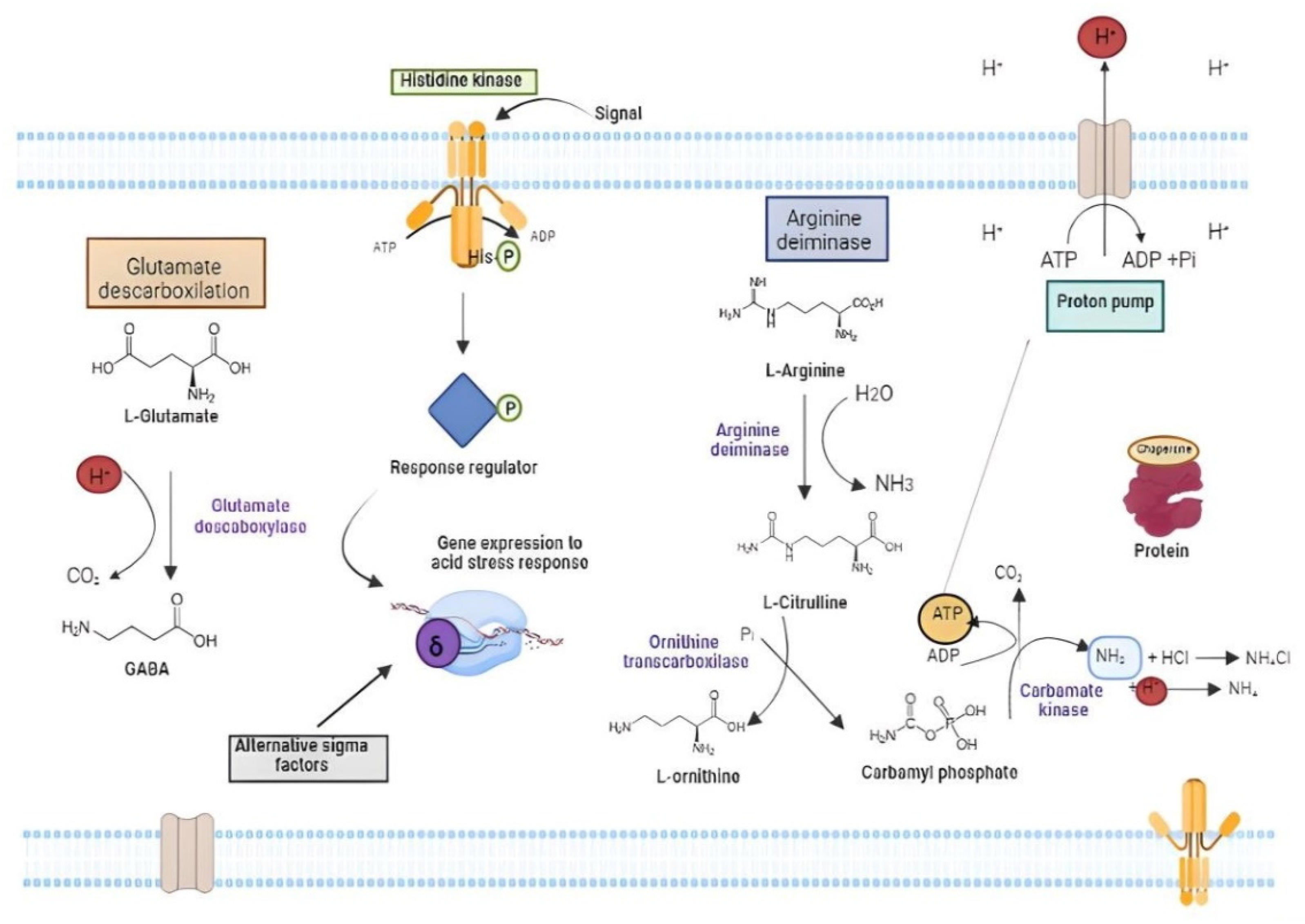
- Cotter, P.D.; Hill, C. Surviving the Acid Test: Responses of Gram-Positive Bacteria to Low PH. Microbiol. Mol. Biol. Rev. 2003, 67, 429–453. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Fukamachi, T.; Saito, H.; Kobayashi, H. Respiration and the F1Fo-ATPase Enhance Survival under Acidic Conditions in Escherichia Coli. PLoS ONE 2012, 7, e52577. [Google Scholar] [CrossRef] [PubMed]
- Senouci-Rezkallah, K.; Schmitt, P.; Jobin, M.P. Amino Acids Improve Acid Tolerance and Internal PH Maintenance in Bacillus Cereus ATCC14579 Strain. Food Microbiol. 2011, 28, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Mercade, M.; Lindley, N.D.; Loubière, P. Metabolism of Lactococcus lactis subsp. cremoris MG 1363 in acid stress conditions. Int. J. Food Microbiol. 2000, 55, 161–165. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, H.; Lin, Z.; Xu, P. Mechanisms of Acid Tolerance in Bacteria and Prospects in Biotechnology and Bioremediation. Biotechnol. Adv. 2015, 33, 1484–1492. [Google Scholar] [CrossRef]
- O’Byrne, C.P.; Karatzas, K.A.G. The Role of Sigma B (ΣB) in the Stress Adaptations of Listeria Monocytogenes: Overlaps Between Stress Adaptation and Virulence. Adv. Appl. Microbiol. 2008, 65, 115–140. [Google Scholar]
- Reeve, B.W.P.; Reid, S.J. Glutamate and Histidine Improve Both Solvent Yields and the Acid Tolerance Response of Clostridium beijerinckii NCP 260. J. Appl. Microbiol. 2016, 120, 1271–1281. [Google Scholar] [CrossRef]
- Wu, Q.; Tun, H.M.; Law, Y.-S.; Khafipour, E.; Shah, N.P. Common Distribution of Gad Operon in Lactobacillus Brevis and Its GadA Contributes to Efficient GABA Synthesis toward Cytosolic Near-Neutral PH. Front. Microbiol. 2017, 8, 206. [Google Scholar] [CrossRef]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-Aminobutyric Acid as a Bioactive Compound in Foods: A Review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Sohlenkamp, C. Membrane Homeostasis in Bacteria upon PH Challenge. In Biogenesis of Fatty Acids, Lipids and Membranes; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–13. [Google Scholar]
- Eguchi, Y.; Utsumi, R. Two-Component Systems in Sensing and Adapting to Acid Stress in Escherichia coli. In Stress and Environmental Regulation of Gene Expression and Adaptation in Bacteria; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; pp. 927–934. [Google Scholar]
- Klauck, E.; Typas, A.; Hengge, R. The σ S Subunit of RNA Polymerase as a Signal Integrator and Network Master Regulator in the General Stress Response in Escherichia coli. Sci. Prog. 2007, 90, 103–127. [Google Scholar] [CrossRef]
- Wu, R.; Zhang, W.; Sun, T.; Wu, J.; Yue, X.; Meng, H.; Zhang, H. Proteomic Analysis of Responses of a New Probiotic Bacterium Lactobacillus Casei Zhang to Low Acid Stress. Int. J. Food Microbiol. 2011, 147, 181–187. [Google Scholar] [CrossRef] [PubMed]
- O’Connell Motherway, M.; O’Brien, F.; O’Driscoll, T.; Casey, P.G.; Shanahan, F.; van Sinderen, D. Carbohydrate Syntrophy Enhances the Establishment of Bifidobacterium Breve UCC2003 in the Neonatal Gut. Sci. Rep. 2018, 8, 10627. [Google Scholar] [CrossRef]
- Derrien, M.; van Hylckama Vlieg, J.E.T. Fate, Activity, and Impact of Ingested Bacteria within the Human Gut Microbiota. Trends Microbiol. 2015, 23, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Urdaneta, V.; Casadesús, J. Interactions between Bacteria and Bile Salts in the Gastrointestinal and Hepatobiliary Tracts. Front. Med. 2017, 4. [Google Scholar] [CrossRef] [PubMed]
- Islam, K.B.M.S.; Fukiya, S.; Hagio, M.; Fujii, N.; Ishizuka, S.; Ooka, T.; Ogura, Y.; Hayashi, T.; Yokota, A. Bile Acid Is a Host Factor That Regulates the Composition of the Cecal Microbiota in Rats. Gastroenterology 2011, 141, 1773–1781. [Google Scholar] [CrossRef] [PubMed]
- Arnold, J.W.; Simpson, J.B.; Roach, J.; Kwintkiewicz, J.; Azcarate-Peril, M.A. Intra-Species Genomic and Physiological Variability Impact Stress Resistance in Strains of Probiotic Potential. Front. Microbiol. 2018, 9, 242. [Google Scholar] [CrossRef]
- Nyanga-Koumou, A.P.; Ouoba, L.I.I.; Kobawila, S.C.; Louembe, D. Response Mechanisms of Lactic Acid Bacteria to Alkaline Environments: A Review. Crit. Rev. Microbiol. 2012, 38, 185–190. [Google Scholar] [CrossRef]
- Kakinuma, Y.; Igarashi, K. Potassium/Proton Antiport System of Growing Enterococcus Hirae at High PH. J. Bacteriol. 1995, 177, 2227–2229. [Google Scholar] [CrossRef]
- Kobayashi, H.; Saito, H.; Kakegawa, T. Bacterial Strategies to Inhabit Acidic Environments. J. Gen. Appl. Microbiol. 2000, 46, 235–243. [Google Scholar] [CrossRef]
- Sawatari, Y.; Yokota, A. Diversity and Mechanisms of Alkali Tolerance in Lactobacilli. Appl. Env. Microbiol. 2007, 73, 3909–3915. [Google Scholar] [CrossRef]
- Begley, M.; Gahan, C.G.M.; Hill, C. The Interaction between Bacteria and Bile. FEMS Microbiol. Rev. 2005, 29, 625–651. [Google Scholar] [CrossRef] [PubMed]
- Foley, M.H.; O’Flaherty, S.; Allen, G.; Rivera, A.J.; Stewart, A.K.; Barrangou, R.; Theriot, C.M. Lactobacillus Bile Salt Hydrolase Substrate Specificity Governs Bacterial Fitness and Host Colonization. Proc. Natl. Acad. Sci. USA 2021, 118, e2017709118. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, L.; Margolles, A.; Sánchez, B. Bile Resistance Mechanisms in Lactobacillus and Bifidobacterium. Front. Microbiol. 2013, 4, 396. [Google Scholar] [CrossRef] [PubMed]
- de Boever, P.; Wouters, R.; Verschaeve, L.; Berckmans, P.; Schoeters, G.; Verstraete, W. Protective Effect of the Bile Salt Hydrolase-Active Lactobacillus Reuteri against Bile Salt Cytotoxicity. Appl. Microbiol. Biotechnol. 2000, 53, 709–714. [Google Scholar] [CrossRef]
- Fasnacht, M.; Polacek, N. Oxidative Stress in Bacteria and the Central Dogma of Molecular Biology. Front. Mol. Biosci. 2021, 8, 671037. [Google Scholar] [CrossRef]
- Imlay, J.A. Cellular Defenses against Superoxide and Hydrogen Peroxide. Annu. Rev. Biochem. 2008, 77, 755–776. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Sun, Z.; Wu, J.; Meng, H.; Zhang, H. Effect of Bile Salts Stress on Protein Synthesis of Lactobacillus Casei Zhang Revealed by 2-Dimensional Gel Electrophoresis. J. Dairy Sci. 2010, 93, 3858–3868. [Google Scholar] [CrossRef]
- Sanchez, B. Proteomics of Stress Response in Bifidobacterium. Front. Biosci. 2008, 13, 6905. [Google Scholar] [CrossRef][Green Version]
- Savijoki, K.; Suokko, A.; Palva, A.; Valmu, L.; Kalkkinen, N.; Varmanen, P. Effect of Heat-Shock and Bile Salts on Protein Synthesis of Bifidobacterium Longum Revealed by [35S]Methionine Labelling and Two-Dimensional Gel Electrophoresis. FEMS Microbiol. Lett. 2005, 248, 207–215. [Google Scholar] [CrossRef][Green Version]
- Bron, P.A.; van Baarlen, P.; Kleerebezem, M. Emerging Molecular Insights into the Interaction between Probiotics and the Host Intestinal Mucosa. Nat. Rev. Microbiol. 2012, 10, 66–78. [Google Scholar] [CrossRef]
- Haddaji, N.; Mahdhi, A.K.; Krifi, B.; Ismail, M.B.; Bakhrouf, A. Change in Cell Surface Properties of Lactobacillus Casei under Heat Shock Treatment. FEMS Microbiol. Lett. 2015, 362, fnv047. [Google Scholar] [CrossRef] [PubMed]
- Popowska, M.; Krawczyk-Balska, A.; Ostrowski, R.; Desvaux, M. InlL from Listeria Monocytogenes Is Involved in Biofilm Formation and Adhesion to Mucin. Front. Microbiol. 2017, 8, 660. [Google Scholar] [CrossRef]
- Irie, Y.; Borlee, B.R.; O’Connor, J.R.; Hill, P.J.; Harwood, C.S.; Wozniak, D.J.; Parsek, M.R. Self-Produced Exopolysaccharide Is a Signal That Stimulates Biofilm Formation in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2012, 109, 20632–20636. [Google Scholar] [CrossRef]
- Hooper, L.V.; Gordon, J.I. Commensal Host-Bacterial Relationships in the Gut. Science 2001, 292, 1115–1118. [Google Scholar] [CrossRef] [PubMed]
- Halfvarson, J.; Brislawn, C.J.; Lamendella, R.; Vázquez-Baeza, Y.; Walters, W.A.; Bramer, L.M.; D’Amato, M.; Bonfiglio, F.; McDonald, D.; Gonzalez, A.; et al. Dynamics of the Human Gut Microbiome in Inflammatory Bowel Disease. Nat. Microbiol. 2017, 2, 17004. [Google Scholar] [CrossRef] [PubMed]
- Kailasapathy, K.; Chin, J. Survival and Therapeutic Potential of Probiotic Organisms with Reference to Lactobacillus Acidophilus and Bifidobacterium Spp. Immunol. Cell Biol. 2000, 78, 80–88. [Google Scholar] [CrossRef]
- Huang, Y.; Adams, M.C. In Vitro Assessment of the Upper Gastrointestinal Tolerance of Potential Probiotic Dairy Propionibacteria. Int. J. Food Microbiol. 2004, 91, 253–260. [Google Scholar] [CrossRef]
- Naissinger da Silva, M.; Tagliapietra, B.L.; Flores, V.d.A.; Pereira dos Santos Richards, N.S. In Vitro Test to Evaluate Survival in the Gastrointestinal Tract of Commercial Probiotics. Curr. Res. Food Sci. 2021, 4, 320–325. [Google Scholar] [CrossRef]
- Guan, N.; Liu, L. Microbial Response to Acid Stress: Mechanisms and Applications. Appl. Microbiol. Biotechnol. 2020, 104, 51–65. [Google Scholar] [CrossRef]
- Guan, N.; Li, J.; Shin, H.; Du, G.; Chen, J.; Liu, L. Metabolic Engineering of Acid Resistance Elements to Improve Acid Resistance and Propionic Acid Production of Propionibacterium jensenii. Biotechnol. Bioeng. 2016, 113, 1294–1304. [Google Scholar] [CrossRef]
- Shi, S.; Qi, Z.; Sheng, T.; Tu, J.; Shao, Y.; Qi, K. Antagonistic Trait of Lactobacillus Reuteri S5 against Salmonella Enteritidis and Assessment of Its Potential Probiotic Characteristics. Microb. Pathog. 2019, 137, 103773. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Gong, L.; Yu, H.; He, G.; Zhang, J.; Han, Y.; Liu, Y.; Hu, J.; Dong, J.; Liu, J.; et al. Antagonistic Activity and Mechanism of Lactobacillus Rhamnosus SQ511 against Salmonella Enteritidis. 3 Biotech 2022, 12, 126. [Google Scholar] [CrossRef] [PubMed]
- Calaça, P.R.D.A.; Da Silva, E.C.; De Melo, F.P.; Santos, D.D.S.; Aragão, A.B.L.; Silva, P.E.D.C.E.; Barros, M.R.; Porto, A.L.F.; Soares, M.T.C.V. Enterococcus Faecium EF137V: Uma Nova Fonte Estratégica Para o Controle Da Saúde Humana e Animal Contra Espécies de Campylobacter. Res. Soc. Dev. 2020, 9, e529108853. [Google Scholar] [CrossRef]
- Balta, I.; Butucel, E.; Stef, L.; Pet, I.; Gradisteanu-Pircalabioru, G.; Chifiriuc, C.; Gundogdu, O.; McCleery, D.; Corcionivoschi, N. Anti- Campylobacter Probiotics: Latest Mechanistic Insights. Foodborne Pathog. Dis 2022, 19, 693–703. [Google Scholar] [CrossRef]
- Fonseca, J.F.; Alvim, L.B.; Nunes, Á.C.; Oliveira, F.M.S.; Amaral, R.S.; Caliari, M.V.; Nicoli, J.R.; Neumann, E.; Gomes, M.A. Probiotic Effect of Bifidobacterium longum 5 1A and Weissella paramesenteroides WpK4 on Gerbils Infected with Giardia lamblia. J. Appl. Microbiol. 2019, 127, 1184–1191. [Google Scholar] [CrossRef]
- Nistor-Cseppento, C.D.; Moga, T.D.; Bungau, A.F.; Tit, D.M.; Negrut, N.; Pasca, B.; Bochis, C.F.; Ghitea, T.C.; Jurcau, A.; Purza, A.L.; et al. The Contribution of Diet Therapy and Probiotics in the Treatment of Sarcopenia Induced by Prolonged Immobilization Caused by the COVID-19 Pandemic. Nutrients 2022, 14, 4701. [Google Scholar] [CrossRef]
- Ren, Z.; Hong, Y.; Huo, Y.; Peng, L.; Lv, H.; Chen, J.; Wu, Z.; Wan, C. Prospects of Probiotic Adjuvant Drugs in Clinical Treatment. Nutrients 2022, 14, 4723. [Google Scholar] [CrossRef]
- Lye, H.-S.; Rahmat-Ali, G.R.; Liong, M.-T. Mechanisms of Cholesterol Removal by Lactobacilli under Conditions That Mimic the Human Gastrointestinal Tract. Int. Dairy J. 2010, 20, 169–175. [Google Scholar] [CrossRef]
- Elkins, C.A.; Moser, S.A.; Savage, D.C. Genes Encoding Bile Salt Hydrolases and Conjugated Bile Salt Transporters in Lactobacillus Johnsonii 100-100 and Other Lactobacillus Species. Microbiology 2001, 147, 3403–3412. [Google Scholar] [CrossRef]
- Ishimwe, N.; Daliri, E.B.; Lee, B.H.; Fang, F.; Du, G. The Perspective on Cholesterol-Lowering Mechanisms of Probiotics. Mol Nutr. Food Res. 2015, 59, 94–105. [Google Scholar] [CrossRef]
- Grill, J.P.; Cayuela, C.; Antoine, J.M.; Schneider, F. Effects of Lactobacillus Amylovorus and Bifidobacterium Breve on Cholesterol. Lett. Appl. Microbiol. 2000, 31, 154–156. [Google Scholar] [CrossRef]
- Kumar, M.; Nagpal, R.; Kumar, R.; Hemalatha, R.; Verma, V.; Kumar, A.; Chakraborty, C.; Singh, B.; Marotta, F.; Jain, S.; et al. Cholesterol-Lowering Probiotics as Potential Biotherapeutics for Metabolic Diseases. Exp. Diabetes Res. 2012, 2012, 902917. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.H.; Kim, J.G.; Shin, Y.W.; Kim, H.S.; Kim, Y.-J.; Chun, T.; Kim, S.H.; Whang, K.Y. Effects of Lactobacillus acidophilus 43121 and a Mixture of Lactobacillus casei and Bifidobacterium longum on the Serum Cholesterol Level and Fecal Sterol Excretion in Hypercholesterolemia-Induced Pigs. Biosci. Biotechnol. Biochem. 2008, 72, 595–600. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jones, M.L.; Martoni, C.J.; Prakash, S. Cholesterol Lowering and Inhibition of Sterol Absorption by Lactobacillus Reuteri NCIMB 30242: A Randomized Controlled Trial. Eur. J. Clin. Nutr. 2012, 66, 1234–1241. [Google Scholar] [CrossRef]
- Choi, S.-B.; Lew, L.-C.; Yeo, S.-K.; Nair Parvathy, S.; Liong, M.-T. Probiotics and the BSH-Related Cholesterol Lowering Mechanism: A Jekyll and Hyde Scenario. Crit. Rev. Biotechnol. 2015, 35, 392–401. [Google Scholar] [CrossRef] [PubMed]
- Lenoir-Wijnkoop, I.; Sanders, M.E.; Cabana, M.D.; Caglar, E.; Corthier, G.; Rayes, N.; Sherman, P.M.; Timmerman, H.M.; Vaneechoutte, M.; van Loo, J.; et al. Probiotic and Prebiotic Influence Beyond the Intestinal Tract. Nutr. Rev. 2008, 65, 469–489. [Google Scholar] [CrossRef] [PubMed]
- Ivey, K.L.; Hodgson, J.M.; Kerr, D.A.; Lewis, J.R.; Thompson, P.L.; Prince, R.L. The Effects of Probiotic Bacteria on Glycaemic Control in Overweight Men and Women: A Randomised Controlled Trial. Eur. J. Clin. Nutr. 2014, 68, 447–452. [Google Scholar] [CrossRef]
- Yadav, H.; Lee, J.-H.; Lloyd, J.; Walter, P.; Rane, S.G. Beneficial Metabolic Effects of a Probiotic via Butyrate-Induced GLP-1 Hormone Secretion. J. Biol. Chem. 2013, 288, 25088–25097. [Google Scholar] [CrossRef]
- Drucker, D.J.; Nauck, M.A. The Incretin System: Glucagon-like Peptide-1 Receptor Agonists and Dipeptidyl Peptidase-4 Inhibitors in Type 2 Diabetes. Lancet 2006, 368, 1696–1705. [Google Scholar] [CrossRef]
- Soty, M.; Penhoat, A.; Amigo-Correig, M.; Vinera, J.; Sardella, A.; Vullin-Bouilloux, F.; Zitoun, C.; Houberdon, I.; Mithieux, G. A Gut–Brain Neural Circuit Controlled by Intestinal Gluconeogenesis Is Crucial in Metabolic Health. Mol. Metab. 2015, 4, 106–117. [Google Scholar] [CrossRef]
- Belenguer, A.; Duncan, S.H.; Calder, A.G.; Holtrop, G.; Louis, P.; Lobley, G.E.; Flint, H.J. Two Routes of Metabolic Cross-Feeding between Bifidobacterium Adolescentis and Butyrate-Producing Anaerobes from the Human Gut. Appl. Env. Microbiol. 2006, 72, 3593–3599. [Google Scholar] [CrossRef] [PubMed]
- Nikbakht, E.; Khalesi, S.; Singh, I.; Williams, L.T.; West, N.P.; Colson, N. Effect of Probiotics and Synbiotics on Blood Glucose: A Systematic Review and Meta-Analysis of Controlled Trials. Eur. J. Nutr. 2018, 57, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Bagarolli, R.A.; Tobar, N.; Oliveira, A.G.; Araújo, T.G.; Carvalho, B.M.; Rocha, G.Z.; Vecina, J.F.; Calisto, K.; Guadagnini, D.; Prada, P.O.; et al. Probiotics Modulate Gut Microbiota and Improve Insulin Sensitivity in DIO Mice. J. Nutr. Biochem. 2017, 50, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Yu, C.-H.; Li, X.-J.; Yao, J.-M.; Fang, Z.-Y.; Yoon, S.-H.; Yu, W.-Y. Gut Dysbiosis in Nonalcoholic Fatty Liver Disease: Pathogenesis, Diagnosis, and Therapeutic Implications. Front. Cell Infect. Microbiol. 2022, 12, 997018. [Google Scholar] [CrossRef]
- Hashim, H.M.; Makpol, S. A Review of the Preclinical and Clinical Studies on the Role of the Gut Microbiome in Aging and Neurodegenerative Diseases and Its Modulation. Front. Cell Neurosci. 2022, 16, 1007166. [Google Scholar] [CrossRef]
- Fang, X.; Miao, R.; Wei, J.; Wu, H.; Tian, J. Advances in Multi-Omics Study of Biomarkers of Glycolipid Metabolism Disorder. Comput. Struct. Biotechnol. J. 2022, 20, 5935–5951. [Google Scholar] [CrossRef]
- Cheng, Z.; Zhang, L.; Yang, L.; Chu, H. The Critical Role of Gut Microbiota in Obesity. Front. Endocrinol. 2022, 13, 1025706. [Google Scholar] [CrossRef]
- Xu, H.; Yang, F.; Bao, Z. Gut Microbiota and Myocardial Fibrosis. Eur. J. Pharm. 2022, 175355. [Google Scholar] [CrossRef]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the Gut Microbiota in Disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef]
- Wischmeyer, P.E.; McDonald, D.; Knight, R. Role of the Microbiome, Probiotics, and ‘Dysbiosis Therapy’ in Critical Illness. Curr. Opin. Crit. Care 2016, 22, 347–353. [Google Scholar] [CrossRef]
- Mahmoudi, H.; Hossainpour, H. Application and Development of Fecal Microbiota Transplantation in the Treatment of Gastrointestinal and Metabolic Diseases: A Review. Saudi J. Gastroenterol. 2022. [Google Scholar] [CrossRef] [PubMed]






| Definition | References |
|---|---|
| Substances produced by microorganisms that promote the growth of other microorganisms. | [7] |
| “Organisms and substances which contribute to intestinal microbial balance.” | [8] |
| “A live microbial feed supplement that beneficially affects the host animal by improving its intestinal microbial balance.” | [9] |
| “Living micro-organisms administered in a sufficient number to survive in the intestinal ecosystem. They must have a positive effect on the host.” | [10] |
| “A mono or mixed culture of living microorganisms that benefit humans or animals by improving the properties of the indigenous microflora.” | [11] |
| “Living micro-organisms, which upon ingestion in certain numbers, exert health benefits beyond inherent basic nutrition.” | [12] |
| “Live microorganisms which when administered in adequate amounts confer a health benefit on the host.” | [13] |
| “Live microorganisms that are intended to have health benefits when consumed or applied to the body.” | [14] |
| “Viable or inviable microbial cell (vegetative or spore; intact or broken) that is potentially healthful to the host.” | [15] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendonça, A.A.; Pinto-Neto, W.d.P.; da Paixão, G.A.; Santos, D.d.S.; De Morais, M.A., Jr.; De Souza, R.B. Journey of the Probiotic Bacteria: Survival of the Fittest. Microorganisms 2023, 11, 95. https://doi.org/10.3390/microorganisms11010095
Mendonça AA, Pinto-Neto WdP, da Paixão GA, Santos DdS, De Morais MA Jr., De Souza RB. Journey of the Probiotic Bacteria: Survival of the Fittest. Microorganisms. 2023; 11(1):95. https://doi.org/10.3390/microorganisms11010095
Chicago/Turabian StyleMendonça, Allyson Andrade, Walter de Paula Pinto-Neto, Giselle Alves da Paixão, Dayane da Silva Santos, Marcos Antonio De Morais, Jr., and Rafael Barros De Souza. 2023. "Journey of the Probiotic Bacteria: Survival of the Fittest" Microorganisms 11, no. 1: 95. https://doi.org/10.3390/microorganisms11010095
APA StyleMendonça, A. A., Pinto-Neto, W. d. P., da Paixão, G. A., Santos, D. d. S., De Morais, M. A., Jr., & De Souza, R. B. (2023). Journey of the Probiotic Bacteria: Survival of the Fittest. Microorganisms, 11(1), 95. https://doi.org/10.3390/microorganisms11010095