
Prevalence, Antibiotics Resistance and Plasmid Profiling of Vibrio spp. Isolated from Cultured Shrimp in Peninsular Malaysia

 , , , ,
, , , ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Shrimp Sampling and In-Situ Examination
2.2. Bacterial Isolation and Identification
2.3. Biochemical Tests
2.4. Bacterial Identification Using pyrH Gene
2.5. Sequence Editing and Analysis
2.6. Phylogenetic Analysis
2.7. Antibiotic Susceptibility Test
2.8. Plasmid Extraction
2.9. Plasmid Curing
3. Results
3.1. Clinical Examination
3.2. Isolation and Identification of Vibrio spp.
3.3. Biochemical Characterization of Vibrio spp.
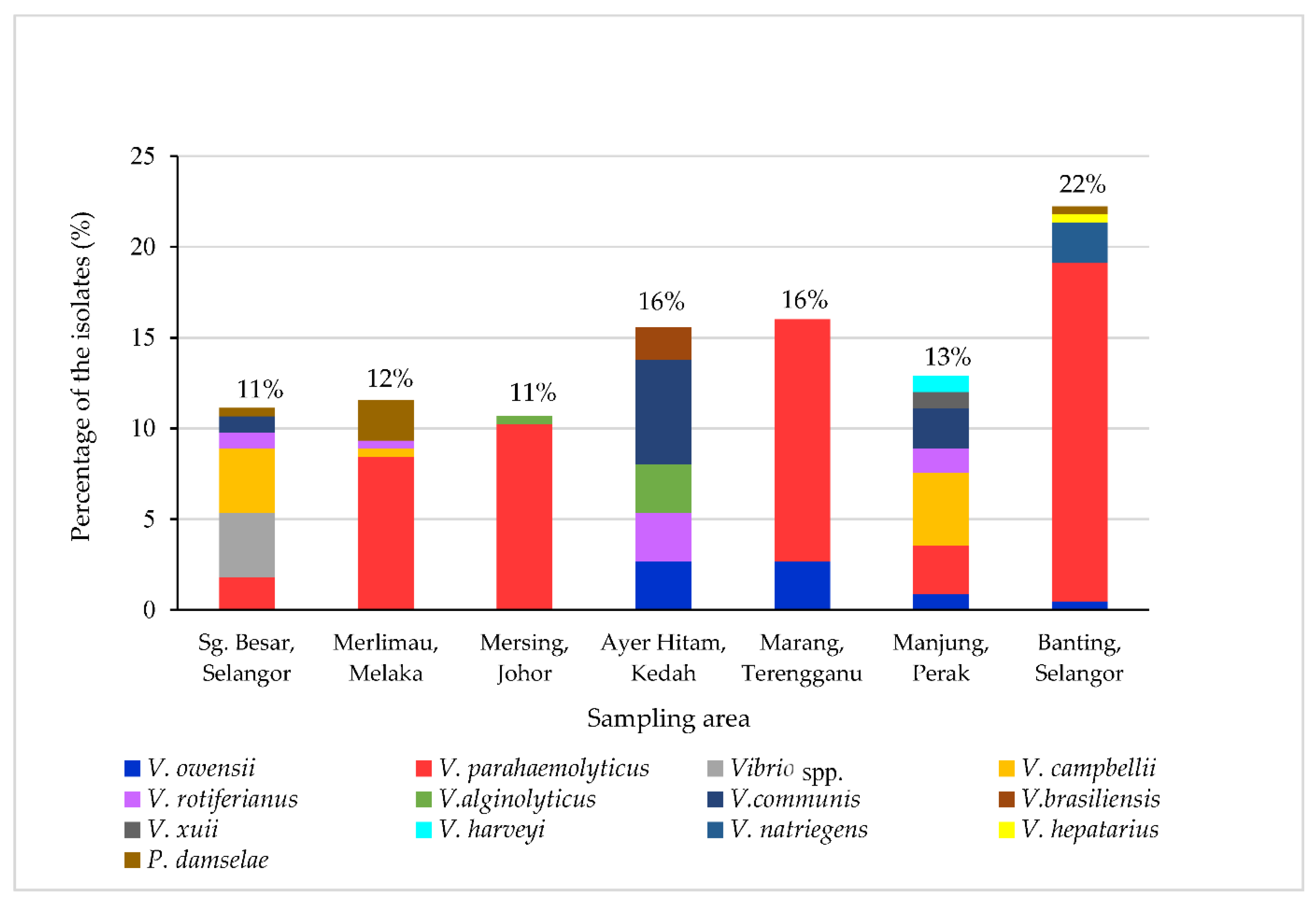
3.4. Prevalence of Vibrio spp.
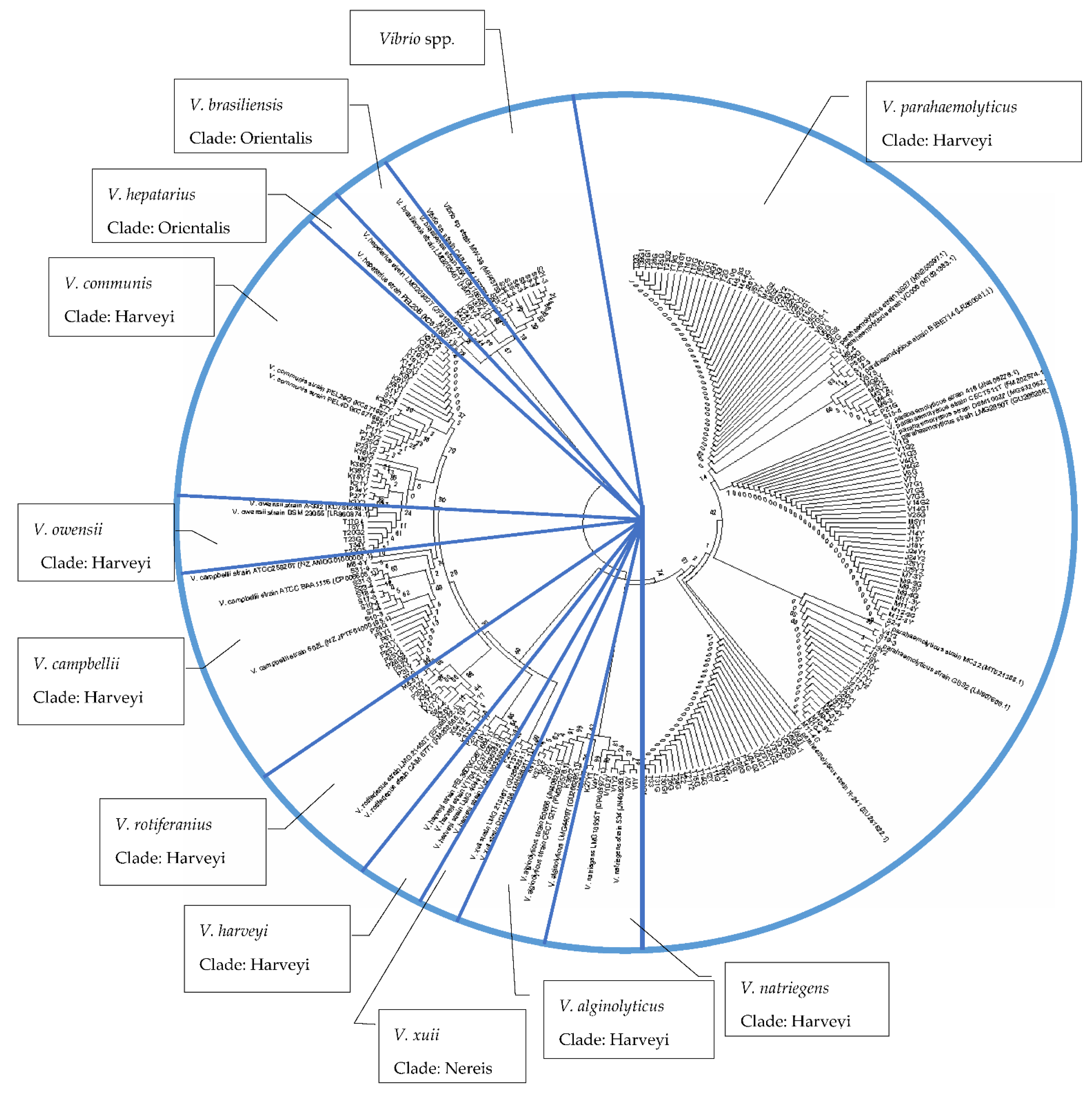
3.5. Phylogenetic Analyses of Vibrio spp.
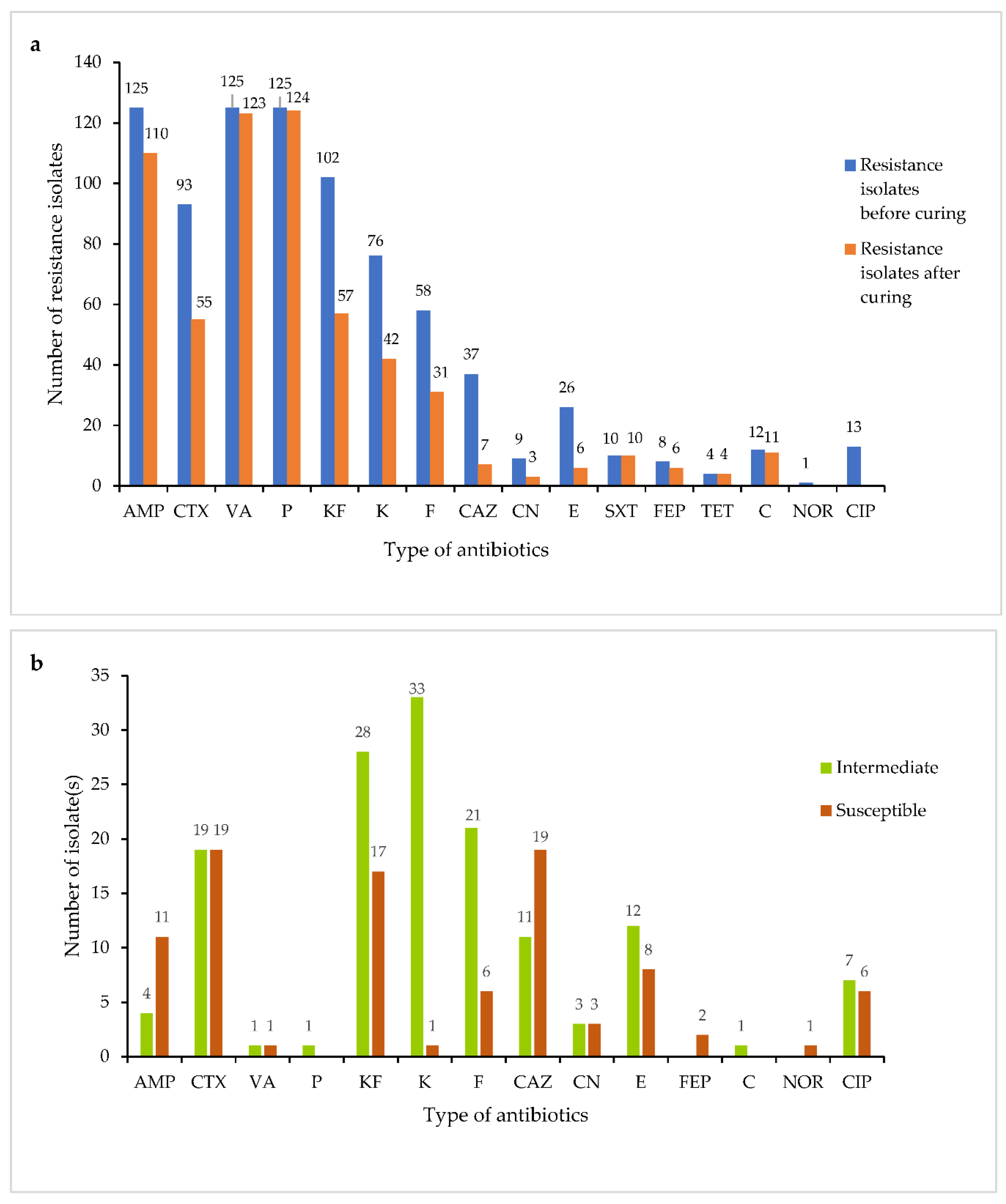
3.6. Antibiotics Susceptibility Test
3.7. Multiple Antibiotic Resistance (MAR) Index
3.8. Plasmid Profiling
3.9. Plasmid Curing
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Vibrio spp. | Accession Number |
|---|---|---|
| 1. | V. communis strain PEL26G | KC871657.1 |
| 2. | V. communis strain PEL4D | KC871668.1 |
| 3. | V. campbellii strain ATCC BAA1116 | CP006605.1 |
| 4. | V. campbellii strain 602L | NZJPTF01000195.1 |
| 5. | V. campbellii strain ATCC25920T | NZ_AMDG01000007.1 |
| 6. | V. rotiferianus strain CAIM 577T | FM202568.1 |
| 7. | V. rotiferianus strain LMG 21460T | EF596722.1 |
| 8. | V. brasiliensis strain LMG20546T | HM771374.1 |
| 9. | V. brasiliensis strain 45A | GU186250.1 |
| 10. | V. hepatarius strain LMG20362T | JF316674.1 |
| 11. | V. hepatarius strain PEL20B | KC871687.1 |
| 12. | V. xuii strain DSM 17185 | MH325211.1 |
| 13. | V. xuii strain LMG 21346T | GU266284.1 |
| 14. | V. parahaemolyticus strain 418 | JN408278.1 |
| 15. | V. parahaemolyticus strain B1BET14 | LR860681.1 |
| 16. | V. parahaemolyticus strain CECT511T | FM202574.1 |
| 17. | V. parahaemolyticus strain DSM10027 | MG932062.1 |
| 18. | V. parahaemolyticus strain GBS2 | LN907608.1 |
| 19. | V. parahaemolyticus strain LMG2850T | GU266286.1 |
| 20. | V. parahaemolyticus strain MC32 | MT521388.1 |
| 21. | V. parahaemolyticus strain NS27 | MN253397.1 |
| 22. | V. parahaemolyticus strain R-241 | EU251622.1 |
| 23. | V. parahaemolyticus strain VC006 | MT521383.1 |
| 24. | V. alginolyticus strain E0666 | JN408262.1 |
| 25. | V. alginolyticus strain CECT 521T | FM202578.1 |
| 26. | V. alginolyticus LMG4409T | GU266285.1 |
| 27. | V. natriegens strain 534 | JN408283.1 |
| 28. | V. natriegens LMG10935T | CP009977.1 |
| 29. | V. owensii strain DSM 23055 | LR860874.1 |
| 30. | V. owensii strain A-332 | KC751288.1 |
| 31. | V. harveyi strain PEL36D | KC871684.1 |
| 32. | V. harveyi strain V1704 | LC370201.1 |
| 33. | V. harveyi strain VJ2 | JN033883.1 |
| 34. | V. harveyi strain LMG 4044T | EF596355.1 |
| 35. | Vibrio sp. strain CAIM 954 | JF739417.1 |
| 36. | Vibrio sp. strain MW-38 | MH457531.1 |
References
- Yano, Y.; Hamano, K.; Satomi, M.; Tsutsui, I.; Ban, M.; Aue-Umneoy, D. Prevalence and antimicrobial susceptibility of Vibrio species related to food safety isolated from shrimp cultured at inland ponds in Thailand. Food Control 2014, 38, 30–36. [Google Scholar] [CrossRef]
- FAO. State of Fisheries and Aquaculture in the World; Food and Agriculture Organization of the United Nations: Rome, Italy, 2019; ISBN 9781424464968. [Google Scholar]
- Xu, X.; Cheng, J.; Wu, Q.; Zhang, J.; Xie, T. Prevalence, characterization, and antibiotic susceptibility of Vibrio parahaemolyticus isolated from retail aquatic products in North China. BMC Microbiol. 2016, 16, 32. [Google Scholar] [CrossRef]
- Verdegem, M.C.J. Nutrient discharge from aquaculture operations in function of system design and production environment. Rev. Aquac. 2013, 5, 158–171. [Google Scholar] [CrossRef]
- Allcock, S.; Young, E.H.; Holmes, M.; Gurdasani, D.; Dougan, G.; Sandhu, M.S.; Török, M.E. Antimicrobial resistance in human populations: Challenges and opportunities. Glob. Health Epidemiol. Genom. 2017, 2, 1–7. [Google Scholar]
- Retamales, R. Bacterial community composition in a semi-intensive shrimp culture of L. vannamei during El Niño 1997-98 and La Niña 1999. Investig. Mar. 2001, 30, 198–200. [Google Scholar] [CrossRef]
- Bermudez-Almada, M.C.; Espinosa-Plascencia, A. The use of antibiotics in shrimp farming. Health Environ. Aquac. 2012, 1, 199–214. [Google Scholar]
- Rico, A.; Oliveira, R.; McDonough, S.; Matser, A.; Khatikarn, J.; Satapornvanit, K.; Van den Brink, P.J. Use, fate and ecological risks of antibiotics applied in tilapia cage farming in Thailand. Environ. Pollut. 2014, 191, 8–16. [Google Scholar] [CrossRef]
- Nsofor, C.A.; Nwokenkwo, V.N.; Ohale, C.U. Prevalence and antibiotic susceptibility pattern of Staphylococcus aureus isolated from various clinical specimens in south-East Nigeria. MOJ Cell Sci. Rep. 2016, 3, 1–5. [Google Scholar] [CrossRef]
- Azzam-Sayuti, M.; Ina-Salwany, M.Y.; Zamri-Saad, M.; Yusof, M.T.; Annas, S.; Najihah, M.Y.; Liles, M.R.; Monir, M.S.; Zaidi, Z.; Amal, M.N.A. The prevalence, putative virulence genes, and antibiotic resistance profiles of Aeromonas spp. isolated from cultivated freshwater fish in peninsular Malaysia. Aquaculture 2021, 540, 736719. [Google Scholar] [CrossRef]
- Elmahdi, S.; DaSilva, L.V.; Parveen, S. Antibiotic resistance of Vibrio parahaemolyticus and Vibrio vulnificus in various countries: A review. Food Microbiol. 2016, 57, 128–134. [Google Scholar] [CrossRef]
- Sambhu, C.; Mamdouh, H.; Mohammad, B.; Khalid, K.; Osama, A.; Hazeem, F.; Abdulmohsen, S.; Nasser, A. Green water technology for the production of Pacific white shrimp Penaeus vannamei (Boone, 1931). Indian J. Fish. 2017, 64, 43–49. [Google Scholar]
- Hong, X.; Lu, L.; Xu, D. Progress in research on acute hepatopancreatic necrosis disease (AHPND). Aquac. Int. 2016, 24, 577–593. [Google Scholar] [CrossRef]
- Ng, W.; Koh, C. The utilization and mode of action of organic acids in the feeds of cultured aquatic animals. Rev. Aquac. 2016, 9, 342–368. [Google Scholar] [CrossRef]
- Chaweepack, T.; Muenthaisong, B.; Chaweepack, S.; Kamei, K. The potential of Galangal (Alpinia galanga Linn.) extract against the pathogens that cause white feces syndrome and acute hepatopancreatic necrosis disease (AHPND) in Pacific White Shrimp (Litopenaeus vannamei). Int. J. Biol. 2015, 7, 8–17. [Google Scholar] [CrossRef]
- De Schryver, P.; Defoirdt, T.; Sorgeloos, P. Early mortality syndrome outbreaks: A microbial management issue in shrimp farming? PLoS Pathog. 2014, 10, e1003919. [Google Scholar] [CrossRef] [PubMed]
- Zorriehzahra, M.J.; Banaederakhshan, R. Early mortality syndrome (EMS) as new emerging threat in shrimp industry. Adv. Amin. Vet. Sci. 2015, 3, 64–72. [Google Scholar] [CrossRef]
- Lesley, M.B.; Velnetti, L.; Cheah, Y.K.; Son, R.; Kasing, A.; Samuel, L. Antibiotic resistance and plasmid profiling of Vibrio parahaemolyticus isolated from cockles (Anadara granosa) at Tanjung Karang, Kuala Selangor. Int. Food Res. J. 2011, 18, 1183–1188. [Google Scholar]
- Letchumanan, V.; Chan, K.G.; Lee, L.H. An insight of traditional plasmid curing in Vibrio species. Front. Microbiol. 2015, 6, 753. [Google Scholar] [CrossRef]
- Amalina, N.Z.; Santha, S.; Zulperi, D.; Amal, M.N.A.; Yusof, M.T.; Zamri-Saad, M.; Ina-Salwany, M.Y. Prevalence, antimicrobial susceptibility and plasmid profiling of Vibrio spp. isolated from cultured groupers in Peninsular Malaysia. BMC Microbiol. 2019, 19, 251. [Google Scholar] [CrossRef]
- Liu, X.; Wang, D.; Wang, H.; Feng, E.; Zhu, L. Curing of plasmid pXO1 from Bacillus anthracis using plasmid incompatibility. PLoS ONE 2012, 7, e29875. [Google Scholar]
- Muthukrishnan, S.; Defoirdt, T.; Ina-Salwany, M.Y.; Yusoff, F.M.; Shariff, M.; Ismail, S.I.; Natrah, I. Vibrio parahaemolyticus and Vibrio harveyi causing Acute Hepatopancreatic Necrosis Disease (AHPND) in Penaeus vannamei (Boone, 1931) isolated from Malaysian shrimp ponds. Aquaculture 2019, 511, 734227. [Google Scholar] [CrossRef]
- Kua, B.C.; Mohd Fariduddin, O.; Marzukhi, O.; Ahmad Iftikhar, A.M. Mortality outbreaks in whiteleg shrimp (Penaeus vannamei Boone 1931) cultured in Peninsular Malaysia. Asian Fish. Sci. 2018, 31, 242–256. [Google Scholar] [CrossRef]
- Haldar, S.; Chatterjee, S.; Sugimoto, N.; Das, S.; Chowdhury, N.; Hinenoya, A.; Asakura, M.; Yamasaki, S. Identification of Vibrio campbellii isolated from diseased farm-shrimps from south India and establishment of its pathogenic potential in an Artemia model. Microbiology 2011, 157, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Jayasinghe, C.V.L.; Ahmed, S.B.N.; Kariyawasam, M.G.I.U. The isolation and identification of Vibrio species in marine shrimp of Sri Lanka. J. Food Agric. 2008, 1, 36–44. [Google Scholar] [CrossRef]
- Buller, N.B. Bacteria and Fungi from Fish and Other Aquatic Animals: A Practice Identification Manual, 2nd ed.; CABI Publishing: Wallingford, UK, 2015. [Google Scholar]
- Chimetto, T.L.; Brocchi, M.; Gondo, M.L.; Thompson, C.C.; Gomez-Gil, B.; Thompson, F. Genomic diversity of Vibrios associated with the Brazilian coral Mussismilia hispida and its sympatric zoanthids (Palythoa caribaeorum, Palythoa variabilis and Zoanthus solanderi). J. Appl. Microbiol. 2009, 106, 1818–1826. [Google Scholar] [CrossRef] [PubMed]
- Noorlis, A.; Ghazali, F.M.; Cheah, Y.K.; Tuan Zainazor, T.C.; Wong, W.C.; Tunung, R.; Nishibuchi, M.; Nakaguchi, Y.; Son, R.; Pui, C.F. Antibiotic resistance and biosafety of Vibrio cholera and Vibrio parahaemolyticus from freshwater fish at retail level. Int. Food Res. J. 2011, 18, 59–66. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Devi, R.; Surendran, P.; Chakraborty, K. Antibiotic resistance and plasmid profiling of Vibrio parahaemolyticus isolated from shrimp farms along the Southwest coast of India. World J. Microbiol. Biotechnol. 2009, 25, 2005–2012. [Google Scholar] [CrossRef]
- Gufe, C.; Canaan Hodobo, T.; Mbonjani, B.; Majonga, O.; Marumure, J.; Musari, S.; Jongi, G.; Makaya, P.V.; Machakwa, J. Antimicrobial profiling of bacteria isolated from fish sold at informal market in Mufakose, Zimbabwe. Int. J. Microbiol. 2019, 2019, 8759636. [Google Scholar] [CrossRef]
- Clinical Laboratory Standard Institute. Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Facteria; Approved Guideline, 2nd ed.; CLSI document M45-A2: Wayne, PA, USA, 2011. [Google Scholar]
- Bauer, A.W.; Kirby, W.M.M.; Shenis, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493. [Google Scholar] [CrossRef]
- Ottaviani, D.; Masini, L.; Bacchiocchi, S. A biochemical protocol for the isolation and identification of current species of Vibrio in seafood. J. Appl. Microbiol. 2003, 95, 1277–1284. [Google Scholar] [CrossRef] [PubMed]
- Kathleen, M.M.; Samuel, L.; Felecia, C.; Reagan, E.L.; Kasing, A.; Lesley, M.; Toh, S.C. Antibiotic resistance of diverse bacteria from aquaculture in Borneo. Int. J. Microbiol. 2016, 2016, 2164761. [Google Scholar] [CrossRef] [PubMed]
- Reboucas, R.H.; De Sousa, O.V.; Lima, A.S.; Vasconcelos, F.R.; de Carvalho, P.B. Antimicrobial resistance profile of Vibrio species isolated from marine shrimp farming environments (Litopenaeus vannamei) at Ceara, Brazil. Environ. Res. 2011, 111, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Sawabe, T.; Kumar, R.; Fukui, Y.; Satomi, M.; Matsushima, R.; Thompson, F.L.; Gomez-Gil, B.; Christen, R.; Maruyama, F.; Kurokawa, K.; et al. Updating the Vibrio clades defined by multilocus sequence phylogeny: Proposal of eight new clades, and the description of Vibrio tritonius sp. nov. Front. Microbiol. 2013, 4, 414. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Trinanes, J.; Gonzalez-Escalona, N.; Martinez-Urtaza, J. Non-Cholera vibrios: The microbial barometer of climate change. Trends Microbiol. 2017, 25, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Haldar, S. Vibrio related diseases in aquaculture and development of rapid and accurate identification methods. J. Mar. Sci. Res. Dev. S 2012, 1, 1–7. [Google Scholar] [CrossRef]
- Banerjee, S.; Ooi, M.C.; Shariff, M.; Khatoon, H. Antibiotic resistant Salmonella and Vibrio associated with farmed Litopenaeus vannamei. Sci. World J. 2012, 2021, 130136. [Google Scholar]
- Pruzzo, C.; Huq, A.; Colwell, R.R. Pathogenic vibrio species in the marine and estuarine environment. In Oceans and Health: Pathogens in the Marine Environment; Belkin, S., Colwell, R.R., Eds.; Springer: Boston, MA, USA, 2005; pp. 217–252. [Google Scholar]
- Baker-Austin, C.; Stockley, L.; Rangdale, R.; Martinez-Urtaza, J. Environmental occurrence and clinical impact of Vibrio vulnificus and Vibrio parahaemolyticus: A European perspective. Environ. Microbiol. Rep. 2010, 2, 7–18. [Google Scholar] [CrossRef]
- Hazen, T.H.; Lafon, P.C.; Garrett, N.M.; Lowe, T.M.; Silberger, D.J.; Rowe, L.A. Insights into the environmental reservoir of pathogenic Vibrio parahaemolyticus using comparative genomics. Front. Microbiol. 2015, 6, 204. [Google Scholar] [CrossRef]
- Kriem, M.R.; Banni, B.; El Bouchtaoui, H.; Hamama, A.; El Marrakchi, A.; Chaouqy, N.; Robert-Pillot, A.; Quilici, M.L. Prevalence of Vibrio spp. in raw shrimp (Parapenaeus longirostris) and performance of a chromogenic medium for the isolation of Vibrio strains. Lett. Appl. Microbiol. 2015, 61, 224–230. [Google Scholar] [CrossRef]
- O’Hara, C.M.; Sowers, E.G.; Bopp, C.A.; Duda, S.B.; Strockbine, N.A. Accuracy of six commercially available systems for identification of members of the family Vibrionaceae. J. Clin. Microbiol. 2003, 41, 5654–5659. [Google Scholar] [CrossRef] [PubMed]
- Garibyan, L.; Avashia, N. 2013. Research techniques made simple: Polymerase chain reaction (PCR). J. Investig. Dermatol. 2013, 133, e6. [Google Scholar]
- Amatul-Samahah, M.; Muthukrishnan, S.; Omar, W.H.H.; Ikhsan, N.; Md Yasin, I. Vibrio spp. associated with acute hepatopancreatic necrosis disease (AHPND) found in penaeid shrimp pond from the east coast of peninsular Malaysia. J. Environ. Biol. 2020, 41, 1160–1170. [Google Scholar] [CrossRef]
- Rajendhran, J.; Gunasekaran, P. Microbial phylogeny and diversity: Small subunit ribosomal RNA sequence analysis and beyond. Microbiol. Res. 2011, 166, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xu, X.; Pu, S.; Huang, S.; Sun, J.; Yang, S.; Zhang, L. Characterization of carbapenemases, extended spectrum β-lactamases, quinolone resistance and aminoglycoside resistance determinants in carbapenem-non-susceptible Escherichia coli from a teaching hospital in Chongqing, Southwest China. Infect. Genet. Evol. 2014, 27, 271–276. [Google Scholar] [CrossRef]
- Voet, D.; Voet, J.G. Biochemistry, 3rd ed.; Wiley: Hoboken, NJ, USA, 2004. [Google Scholar]
- Chimetto, L.A.; Cleenwerck, I.; Alves, N.J.; Silva, B.S.; Brocchi, M.; Willems, A.; De Vos, P.; Thompson, F.L. Vibrio communis sp. nov., isolated from the marine animals Mussismilia hispida, Phyllogorgia dilatata, Palythoa caribaeorum, Palythoa variabilis and Litopenaeus vannamei. Int. J. Syst. Evol. Microbiol. 2011, 61, 362–368. [Google Scholar] [CrossRef]
- Thompson, F.L.; Gomez-Gil, B.; Ribeiro-Vasconcelos, A.T.; Sawabe, T. Multilocus sequence analysis reveals that Vibrio harveyi and Vibrio campbellii form distinct species. Appl. Environ. Microbiol. 2007, 73, 4279–4285. [Google Scholar] [CrossRef]
- Pascual, J.; Macián, M.C.; Arahal, D.R.; Garay, E.; Pujalte, M.J. Multilocus sequence analysis of the central clade of the genus Vibrio by using the 16S rRNA, recA, pyrH, rpoD, gyrB, rctB and toxR genes. Int. J. Syst. Evol. Microbiol. 2010, 60, 154–165. [Google Scholar] [CrossRef]
- Thompson, F.L.; Gevers, D.; Thompson, C.C.; Dawyndt, P.; Naser, S.; Hoste, B.; Munn, C.B.; Swings, J. Phylogeny and molecular identification of vibrios on the basis of multilocus sequence analysis. Appl. Environ. Microbiol. 2005, 71, 5107–5115. [Google Scholar] [CrossRef]
- Zarei, M.; Borujeni, M.P.; Jamnejad, A.; Khezrzadeh, M. Seasonal prevalence of Vibrio species in retail shrimp with an emphasis on Vibrio parahaemolyticus. Food Control 2012, 25, 107–109. [Google Scholar] [CrossRef]
- Jacksic, S.; Suncica, U.; Petrak, T.; Bazulic, D.; Lada, G.K. Occurrence of Vibrio spp. in sea fish, shrimp and bivalve mollusks harvested from Adriatic Sea. Food Control 2002, 13, 491–493. [Google Scholar] [CrossRef]
- Letchumanan, V.; Yin, W.F.; Lee, L.H.; Chan, K.G. Prevalence and antimicrobial susceptibility of Vibrio parahaemolyticus isolated from retail shrimp in Malaysia. Front. Microbiol. 2015, 30, 6–33. [Google Scholar] [CrossRef] [PubMed]
- Koralage, M.S.; Alter, T.; Pichpol, D.; Strauch, E.; Zessin, K.H.; Huehn, S. Prevalence and molecular characteristics of Vibrio spp. isolated from preharvest shrimp of the Northwestern Province of Sri Lanka. J. Food Prot. 2012, 75, 1846–1850. [Google Scholar] [CrossRef] [PubMed]
- Sperling, L.; Alter, T.; Huehn, S. Prevalence and antimicrobial resistance of Vibrio spp. in retail and farm shrimp in Ecuador. J. Food Prot. 2015, 78, 2089–2092. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, H.A.; El Bayomi, R.M.; Hussein, M.A.; Khedr, M.H.E.; Abo Remela, E.M.; El-Ashram, A.M.M. Molecular characterization, antibiotic resistance pattern and biofilm formation of Vibrio parahaemolyticus and V. cholerae isolated from crustaceans and humans. Int. J. Food Microbiol. 2018, 274, 31–37. [Google Scholar] [CrossRef]
- Tanil, G.B.; Radu, S.; Nishibuchi, M.; Rahim, R.A.; Napis, S.; Maurice, L.; Gunsalam, J.W. Characterization of Vibrio parahaemolyticus isolated from coastal seawater in Peninsular Malaysia. Southeast Asian J. Trop. Med. Public Health 2005, 36, 940–945. [Google Scholar]
- Costa, R.A.; Colares, L.P.; Lima, R.A.; Vieira, R.H.S.D.F.; de Sousa, S.V. Effect of seawater on the activity of antibiotics against Vibrios isolated from the hemolymph of cultured pacific white shrimp. J. World Aqua. Soc. 2014, 43, 727–732. [Google Scholar] [CrossRef]
- Chitov, T.; Wongdao, S.; Thatum, W.; Puprae, T.; Sisuwan, P. Occurrence of potentially pathogenic Vibrio species in raw, processed, and ready-to-eat seafood and seafood products. Maejo Int. J. Sci. Technol. 2009, 3, 88–98. [Google Scholar]
- Baffone, W.; Pianetti, A.; Bruscolini, F.; Barbieri, E.; Citterio, B. Occurrence and expression of virulence-related properties of Vibrio species isolated from widely consumed seafood products. Int. J. Food Microbiol. 2000, 5, 9–18. [Google Scholar] [CrossRef]
- Abd-Elghany, S.M.; Sallam, K.I. Occurrence and molecular identification of Vibrio parahaemolyticus in retail shellfish in Mansoura, Egypt. Food Control 2013, 33, 399–405. [Google Scholar] [CrossRef]
- Caburlotto, G.; Suffredini, E.; Toson, M.; Fasolato, L.; Antonetti, P.; Zambon, M.; Manfrin, A. Occurrence and molecular characterisation of Vibrio parahaemolyticus in crustaceans commercialised in Venice area, Italy. Int. J. Food Microbiol. 2016, 220, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, T.J.; Neigel, J.E. Effects of temperature and salinity on prevalence and intensity of infection of blue crabs, Callinectes sapidus, by Vibrio cholerae, V. parahaemolyticus, and V. vulnificus in Louisiana. J. Invertebr. Pathol. 2018, 151, 82–90. [Google Scholar] [CrossRef]
- Smith, P. Antimicrobial resistance in aquaculture. Rev. Sci. Tech. 2008, 27, 243–264. [Google Scholar] [CrossRef] [PubMed]
- Sudha, S.; Mridula, C.; Silvester, R.; Abdulla, M.H. Prevalence and antibiotic resistance of pathogenic Vibrios in shellfishes from Cochin market. Indian J. Geo Mar. Sci. 2014, 43, 815–824. [Google Scholar]
- Vaseeharan, B.; Ramasamy, P.; Murugan, T.; Chen, J.C. In vitro susceptibility of antibiotics against Vibrio spp. and Aeromonas spp. isolated from Penaeus monodon hatcheries and ponds. Int. J. Antimicrob. Agents 2005, 26, 285–291. [Google Scholar] [CrossRef]
- Jayasree, L.; Janakiram, P.; Madhavi, R. Characterization of Vibrio spp. associated with diseased shrimp from culture ponds of Andhra Pradesh (India). J. World Aquac. Soc. 2006, 37, 523–532. [Google Scholar] [CrossRef]
- Srinivasan, P.; Ramasamy, P. Occurrence, distribution and antibiotic resistance patterns of Vibrio species associated with viral diseased shrimp of South Indian aquaculture environment. Int. J. Agric. Sci. 2009, 1, 1. [Google Scholar]
- Albuquerque, C.R.; Araújo, R.L.; Souza, O.V.; Vieira, R.H.S.D.F. Antibiotic-resistant Vibrios in farmed shrimp. BioMed Res. Int. 2015, 2015, 505914. [Google Scholar] [CrossRef]
- Ottaviani, D.; Bacchiocchi, I.; Masini, L.; Leoni, F.; Carraturo, A.; Giammarioli, M.; Sbaraglia, G. Antimicrobial susceptibility of potentially pathogenic halophilic vibrios isolated from seafood. Int. J. Antimicrob. Agents 2001, 18, 135–140. [Google Scholar] [CrossRef]
- Sahilah, A.M.; Laila, R.; Sallehuddin, H.; Hassan, O.; Abdullah, A.; Ariffin, A. Antibiotic resistance and molecular typing among cockle (Anadara granosa) strains of Vibrio parahaemolyticus by polymerase chain reaction (PCR)-based analysis. World J. Microbiol. Biotechnol. 2013, 30, 649–659. [Google Scholar] [CrossRef]
- Mohamad, N.; Amal, M.N.A.; Saad, M.Z.; Yasin, I.S.M.; Zulkiply, N.A.; Mustafa, M.; Nasruddin, N.S. Virulence-associated genes and antibiotic resistance patterns of Vibrio spp. isolated from cultured marine fishes in Malaysia. BMC Vet. Res. 2019, 15, 176. [Google Scholar] [CrossRef] [PubMed]
- Tendencia, E.A.; De La Pena, L.D. Antibiotic resistance of bacteria from shrimp ponds. Aquaculture 2001, 195, 193–204. [Google Scholar] [CrossRef]
- Zanetti, S.; Spanu, T.; Deriu, A.; Romano, L.; Sechi, L.A.; Fadda, G. In vitro susceptibility of Vibrio spp. isolated from the environment. Int. J. Antimicrob. Agents 2001, 17, 407–409. [Google Scholar] [CrossRef]
- You, K.G.; Bong, C.W.; Lee, C.W. Antibiotic resistance and plasmid profiling of Vibrio spp. in tropical waters of Peninsular Malaysia. Environ. Monit. Assess. 2016, 188, 171. [Google Scholar] [CrossRef] [PubMed]
- Thomas, B.T.; Agu, G.C.; Makanjuola, S.O.; Davies, A. Plasmid profiling and antibiotic resistance of extended spectrum beta lactamases producing Pseudomonas aeruginosa expressing Ampc beta-lactamase enzyme. Am. Eurasian J. Sci. Res. 2015, 10, 109–117. [Google Scholar]
- Carattoli, A. Plasmids and the spread of resistance. Int. J. Med. Microbiol. 2013, 303, 298–304. [Google Scholar] [CrossRef]
- Manjusha, S.; Sarita, G.B. Characterization of plasmids from multiple antibiotic resistant Vibrios isolated from molluscan and crustacean of Kerala. Int. Food Res. J. 2013, 20, 77–86. [Google Scholar]
- Zulkifli, Y.; Alitheen, N.B.; Raha, A.R.; Yeap, S.K.; Marlina, S.R.; Nishibuchi, M. Antibiotic resistance and plasmid profiling of Vibrio parahaemolyticus isolated from cockles in Padang, Indonesia. Int. Food Res. J. 2009, 16, 53–58. [Google Scholar]




| Sampling Site | State | Region | Types of Ponds | Number of Shrimp | Shrimp Species | Date | Coordinate |
|---|---|---|---|---|---|---|---|
| Sg. Besar | Selangor | Central | Earthen | 30 | P. monodon | March 2019 | 3.7726 N 100.9666 E |
| Merlimau | Melaka | West | Earthen | 30 | P. monodon | April 2019 | 2.0631 N 102.2910 E |
| Mersing | Johor | South | Earthen | 30 | P. monodon | January 2020 | 2.2941 N 103.4902 E |
| Ayer Hitam | Kedah | North | HDPE lined earth | 30 | P. monodon | March 2020 | 6.2438 N 100.2220 E |
| Marang | Terengganu | East | HDPE lined earth | 30 | P. vannamei | December 2020 | 5.1846 N 103.1991 E |
| Manjung | Perak | North | Earthen | 30 | P. vannamei | January 2021 | 4.3722 N 100.6089 E |
| Banting | Selangor | Central | Earthen | 30 | P. monodon | March 2021 | 2.8222 N 101.4179 E |
| Sampling Site | State | Total Number of Isolates, n (%) | Number of Isolates on TCBS, n (%) | |
|---|---|---|---|---|
| Green Colony | Yellow Colony | |||
| Sg. Besar | Selangor | 25 (11) | 13 (6) | 12 (5) |
| Banting | Selangor | 50 (22) | 43 (19) | 7 (3) |
| Merlimau | Melaka | 26 (12) | 25 (11) | 1 (0.4) |
| Mersing | Johor | 24 (11) | 23 (10) | 1 (0.4) |
| Ayer Hitam | Kedah | 35 (16) | 0 (0) | 35 (16) |
| Marang | Terengganu | 36 (16) | 30 (13) | 6 (3) |
| Manjung | Perak | 29 (13) | 15 (7) | 14 (6) |
| Total | 225 (100) | 149 (66) | 76 (34) | |
| Species | Number of Isolates | TCBS | Gram Staining | Oxidase | Catalase | ONPG | SIM | TSI | LDC | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sulphide Gas Production | Indole Production | Motility | A/A, K/A | Gas Production | H2S Production | ||||||||
| V. owensii | 15 | Y | negative | + | + | + | - | + | + | A/A | - | - | + |
| V. parahaemolyticus | 124 | G | negative | + | + | + | - | + | + | K/A | - | - | + |
| Vibrio spp. | 8 | Y | negative | + | + | - | - | + | + | A/A | - | - | - |
| V. campbellii | 18 | G | negative | + | + | - | - | + | + | A/A | - | - | - |
| V. rotiferianus | 12 | Y | negative | + | + | - | - | + | + | K/A | - | - | + |
| V. alginolyticus | 7 | Y | negative | + | + | - | - | + | + | A/A | - | - | + |
| V. communis | 20 | Y | negative | + | + | - | - | + | + | A/A | - | - | + |
| V. brasiliensis | 4 | Y | negative | + | + | + | - | + | + | A/A | - | - | - |
| V. xuii | 2 | Y | negative | + | + | - | - | + | + | A/A | - | - | - |
| V. harveyi | 2 | Y | negative | + | + | - | - | + | + | A/A | - | - | + |
| V. natriegens | 5 | Y | negative | + | + | - | - | - | + | A/A | - | - | - |
| V. hepatarius | 1 | Y | negative | + | + | - | - | + | + | K/A | - | - | - |
| P. damselae | 7 | G | negative | + | + | + | - | - | + | A/A | - | - | - |
| Vibrio spp. | No of Isolates | Percentage of Resistance Isolates (%) | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TET | SXT | E | AMP | C | CN | NOR | K | FEP | CTX | CAZ | KF | CIP | F | VA | P | ||
| V. parahaemolyticus | 124 | 1 | 14 | 19 | 85 | 9 | 7 | 1 | 40 | 2 | 73 | 24 | 86 | 6 | 53 | 97 | 97 |
| V. campbellii | 18 | 6 | 0 | 44 | 100 | 0 | 0 | 0 | 6 | 0 | 50 | 0 | 56 | 11 | 0 | 50 | 56 |
| V. rotiferianus | 12 | 50 | 17 | 33 | 83 | 0 | 0 | 0 | 8 | 25 | 50 | 33 | 25 | 0 | 0 | 67 | 83 |
| V. owensii | 15 | 0 | 0 | 7 | 100 | 0 | 0 | 0 | 80 | 40 | 80 | 13 | 73 | 27 | 33 | 100 | 87 |
| V. alginolyticus | 7 | 14 | 0 | 14 | 86 | 0 | 14 | 0 | 57 | 0 | 57 | 14 | 100 | 0 | 43 | 100 | 100 |
| V. xuii | 2 | 0 | 0 | 0 | 50 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 50 | 0 | 0 | 100 | 100 |
| V. harveyi | 2 | 0 | 0 | 0 | 100 | 0 | 0 | 0 | 0 | 0 | 50 | 0 | 50 | 0 | 50 | 100 | 100 |
| V. natriegens | 5 | 0 | 20 | 20 | 0 | 0 | 20 | 0 | 80 | 0 | 100 | 100 | 100 | 0 | 100 | 100 | 100 |
| V. hepatarius | 1 | 0 | 0 | 0 | 100 | 0 | 0 | 0 | 100 | 0 | 100 | 0 | 100 | 0 | 100 | 100 | 100 |
| V. communis | 20 | 5 | 0 | 10 | 95 | 5 | 5 | 0 | 35 | 0 | 70 | 0 | 70 | 5 | 5 | 90 | 90 |
| V. brasiliensis | 4 | 0 | 0 | 25 | 75 | 0 | 0 | 0 | 50 | 0 | 0 | 0 | 100 | 0 | 0 | 100 | 100 |
| Vibrio spp. | 8 | 0 | 0 | 100 | 100 | 0 | 0 | 0 | 0 | 0 | 25 | 0 | 13 | 13 | 0 | 100 | 100 |
| P. damselae | 7 | 0 | 0 | 43 | 100 | 0 | 0 | 0 | 57 | 0 | 0 | 0 | 0 | 0 | 0 | 86 | 71 |
| AR Index Value | Group of MAR Index | Number of Resistant Antibiotics | Percentage of Isolates (%) |
|---|---|---|---|
| 0.06 | <0.2 | 1 | 0.4 |
| 0.13 | <0.2 | 2 | 4 |
| 0.19 | <0.2 | 3 | 12 |
| 0.25 | >0.2 | 4 | 15 |
| 0.31 | >0.2 | 5 | 16 |
| 0.38 | >0.2 | 6 | 22 |
| 0.44 | >0.2 | 7 | 13 |
| 0.50 | >0.2 | 8 | 6 |
| 0.56 | >0.2 | 9 | 6 |
| 0.63 | >0.2 | 10 | 2 |
| 0.69 | >0.2 | 11 | 3 |
| 0.75 | >0.2 | 12 | 0.4 |
| Strain ID | Species | Antibiograms | Presence of Plasmid | No of Resistance Antibiotics |
|---|---|---|---|---|
| V22G1 | V. parahaemolyticus | AMP, C, CN, K, CTX, CAZ, KF, F, SXT, E, VA, P | + | 12 |
| V22Y2 | V. parahaemolyticus | AMP, C, CN, K, CTX, CAZ, KF, F, SXT, VA, P | + | 11 |
| V5G | V. parahaemolyticus | AMP, K, CTX, CAZ, KF, CN, F, SXT, E, VA, P | + | 11 |
| V7Y | V. parahaemolyticus | AMP, CN, NOR, CTX, CAZ, KF, CIP, F, E, VA, P | + | 11 |
| V22Y1 | V. parahaemolyticus | AMP, C, K, CTX, CAZ, KF, F, SXT, E, VA, P | + | 11 |
| V23Y2 | V. parahaemolyticus | AMP, C, K, CTX, CAZ, KF, F, SXT, E, VA, P | + | 11 |
| T26G | V.parahaemolyticus | AMP, K, CTX, CAZ, KF, CIP, F, TET, SXT, VA, P | + | 11 |
| V3G2 | V. parahaemolyticus | AMP, K, CTX, CAZ, KF, F, E, C, VA, P | + | 10 |
| V4G1 | V. parahaemolyticus | AMP, K, CTX, CAZ, KF, CIP, CN, F, VA, P | + | 10 |
| V7G1 | V. parahaemolyticus | AMP, K, CTX, CAZ, KF, CIP, F, E, VA, P | + | 10 |
| V8G1 | V. parahaemolyticus | AMP, CN, K, CTX, CAZ, KF, F, SXT, VA, P | + | 10 |
| V5Y1 | V. parahaemolyticus | AMP, C, K, CTX, CAZ, KF, F, SXT, VA, P | + | 10 |
| V7G3 | V. parahaemolyticus | AMP, C, K, CTX, CAZ, KF, F, SXT, VA, P | + | 10 |
| V15G | V. parahaemolyticus | AMP, C, K, CTX, CAZ, KF, F, SXT, VA, P | + | 10 |
| V23G1 | V. parahaemolyticus | AMP, C, K, CTX, K, F, SXT, E, VA, P | - | 10 |
| P10G | V.parahaemolyticus | AMP, K, CTX, CAZ, KF, F, SXT, VA, P | + | 9 |
| T31G | V.parahaemolyticus | AMP, K, CTX, CAZ, KF, F, FEP, VA, P | + | 9 |
| V2G | V. parahaemolyticus | AMP, K, CTX, CAZ, KF, F, SXT, VA, P | + | 9 |
| V3G3-1 | V. parahaemolyticus | AMP, K, CTX, CAZ, KF, F, E, VA, P | + | 9 |
| V4G2 | V. parahaemolyticus | AMP, K, CTX, CAZ, KF, CIP, F, VA, P | + | 9 |
| M5Y1 | V. parahaemolyticus | AMP, K, CTX, KF, F, E, CN, VA, P | - | 9 |
| M5G2 | V. parahaemolyticus | AMP, CTX, KF, CIP, F, SXT, C, VA, P | + | 9 |
| V7G3-1 | V. parahaemolyticus | AMP, K, CTX, CAZ, KF, F, E, VA, P | + | 9 |
| T23G2 | V.parahaemolyticus | AMP, K, CTX, CAZ, KF, F, VA, P | + | 8 |
| T38G1 | V.parahaemolyticus | AMP, K, CTX, KF, CIP, F, VA, P | + | 8 |
| M5Y2 | V. parahaemolyticus | AMP, K, CTX, KF, F, CN, VA, P | - | 8 |
| V1G3 | V. parahaemolyticus | AMP, K CTX, CAZ, KF, F, VA, P | + | 8 |
| V7G2 | V. parahaemolyticus | AMP, K, CTX, CAZ, KF, F, VA, P | + | 8 |
| T39G1 | V.parahaemolyticus | AMP, K, CTX, CAZ, KF, F, VA, P | + | 8 |
| MG2 | V. parahaemolyticus | AMP, C, CTX, KF, SXT, E, VA, P | - | 8 |
| M12-3G | V. parahaemolyticus | AMP, K, CTX, KF, F, VA, P | + | 7 |
| J11Y | V. parahaemolyticus | AMP, K, CTX, KF, F, VA, P | - | 7 |
| T1G | V. parahaemolyticus | AMP, K, CTX, KF, F, VA, P | + | 7 |
| T2G | V. parahaemolyticus | AMP, K, CTX, KF, F, VA, P | + | 7 |
| T3G | V. parahaemolyticus | AMP, K, CTX, KF, F, VA, P | + | 7 |
| T7G1 | V. parahaemolyticus | AMP, K, CTX, KF, F, VA, P | + | 7 |
| T11G | V. parahaemolyticus | AMP, K, CTX, KF, F, VA, P | + | 7 |
| T12Y | V. parahaemolyticus | AMP, K, CTX, KF, F, VA, P | + | 7 |
| T15G | V. parahaemolyticus | AMP, K, CTX, KF, F, VA, P | + | 7 |
| T22 | V. parahaemolyticus | AMP, K, CTX, KF, F, VA, P | + | 7 |
| T24G | V. parahaemolyticus | AMP, K, CTX, KF, F, VA, P | + | 7 |
| V6G | V. parahaemolyticus | AMP, K, CTX, CAZ, KF, VA, P | + | 7 |
| V22G2 | V. parahaemolyticus | AMP, CTX CAZ, KF, E, VA, P | + | 7 |
| V23Y1 | V. parahaemolyticus | AMP, K, CTX, KF, F, VA, P | + | 7 |
| V3Y3 | V. parahaemolyticus | AMP, K, KF, F, E, VA, P | + | 7 |
| V3Y2G | V. parahaemolyticus | AMP, K, KF, F, E, VA, P | + | 7 |
| V3Y3G | V. parahaemolyticus | AMP, K, KF, F, E, VA, P | + | 7 |
| T10Y1 | V. parahaemolyticus | AMP, K, KF, F, VA, P | + | 6 |
| T18G1 | V. parahaemolyticus | AMP, K, CTX, CAZ, VA, P | + | 6 |
| T30G1 | V. parahaemolyticus | AMP, K, CTX, F, VA, P | + | 6 |
| T32G | V. parahaemolyticus | AMP, K, CTX, KF, VA, P | + | 6 |
| T14G | V. parahaemolyticus | AMP, K, CTX, KF, VA, P | + | 6 |
| T25G | V. parahaemolyticus | AMP, K, CTX, KF, VA, P | + | 6 |
| M6-3 | V. parahaemolyticus | AMP, K, CTX, KF, VA, P | - | 6 |
| M6-4 | V. parahaemolyticus | AMP, K, CTX, KF, VA, P | - | 6 |
| M7-3Y | V. parahaemolyticus | AMP, K, CTX, KF, VA, P | - | 6 |
| V24G1 | V. parahaemolyticus | AMP, K, KF, E, VA, P | + | 6 |
| V1G | V. parahaemolyticus | AMP, CTX, F, E, VA, P | + | 6 |
| V1G2 | V. parahaemolyticus | AMP, CTX, KF, F, VA, P | + | 6 |
| T21Y2 | V. parahaemolyticus | AMP, CTX, FEP, F, VA, P | + | 6 |
| V4Y3 | V. parahaemolyticus | AMP, CTX, KF, E, VA, P | + | 6 |
| P14G | V. parahaemolyticus | AMP, CTX, CAZ, KF, VA, P | - | 6 |
| M12-3Y | V. parahaemolyticus | AMP, CTX, KF, F, VA, P | + | 6 |
| J26Y | V. parahaemolyticus | AMP, CTX, KF, F, VA, P | + | 6 |
| T4G1 | V. parahaemolyticus | AMP, CTX, KF, F, VA, P | + | 6 |
| T28G | V. parahaemolyticus | AMP, CTX, KF, F, VA, P | + | 6 |
| T29G1 | V. parahaemolyticus | AMP, CTX, KF, F, VA, P | + | 6 |
| M11-4Y | V. parahaemolyticus | AMP, CAZ, KF, FEP, VA, P | + | 6 |
| J29Y2 | V. parahaemolyticus | K, CTX, KF, F, VA, P | - | 6 |
| J21Y | V. parahaemolyticus | K, CTX, KF, F, VA, P | - | 6 |
| J31Y | V. parahaemolyticus | K, CTX, KF, CIP, VA, P | - | 6 |
| J29Y1 | V. parahaemolyticus | CTX, CAZ, KF, F, VA, P | - | 6 |
| V14G2 | V. parahaemolyticus | AMP, K, CTX, F, VA, P | + | 6 |
| MG2Y | V. parahaemolyticus | AMP, K, CTX, F, VA, P | + | 6 |
| M2Y2-1 | V. parahaemolyticus | AMP, K, CTX, KF, VA, P | - | 6 |
| V14G1 | V. parahaemolyticus | AMP, CTX, KF, SXT, VA, P | + | 6 |
| V25G | V. parahaemolyticus | AMP, CTX, KF, E, VA, P | + | 6 |
| V3G1 | V. parahaemolyticus | AMP, KF, E, VA, P | + | 5 |
| V3Y2Y | V. parahaemolyticus | AMP, KF, E, VA, P | + | 5 |
| V24G2 | V. parahaemolyticus | AMP, KF, E, VA, P | + | 5 |
| P33G | V. parahaemolyticus | AMP, K, KF, VA, P | - | 5 |
| T19G | V. parahaemolyticus | AMP, K, KF, VA, P | + | 5 |
| M4-4G | V. parahaemolyticus | AMP, K, CTX, VA, P | + | 5 |
| T33 | V. parahaemolyticus | AMP, CTX, F, VA, P | + | 5 |
| M9-4Y | V. parahaemolyticus | AMP, CTX, KF, VA, P | - | 5 |
| M10-3Y | V. parahaemolyticus | AMP, CTX, KF, VA, P | + | 5 |
| M10-4 | V. parahaemolyticus | AMP, CTX, KF, VA, P | - | 5 |
| M11-3Y | V. parahaemolyticus | AMP, CTX, KF, VA, P | - | 5 |
| M11-4G | V. parahaemolyticus | AMP, CTX, KF, VA, P | - | 5 |
| T8Y2 | V. parahaemolyticus | AMP, CTX, KF, VA, P | + | 5 |
| T13G | V. parahaemolyticus | AMP, CTX, KF, VA, P | + | 5 |
| T16G | V. parahaemolyticus | AMP, CTX, KF, VA, P | + | 5 |
| J1Y | V. parahaemolyticus | K, CTX, KF, VA, P | - | 5 |
| J10Y | V. parahaemolyticus | K, CTX, KF, VA, P | - | 5 |
| J17Y1 | V. parahaemolyticus | CTX, KF, F, VA, P | - | 5 |
| J23 | V. parahaemolyticus | CTX, KF, F, VA, P | - | 5 |
| P21G | V. parahaemolyticus | AMP, E, VA, P | + | 4 |
| T6G | V. parahaemolyticus | AMP, F, VA, P | + | 4 |
| M4-3Y | V. parahaemolyticus | AMP, K, VA, P | + | 4 |
| J14Y | V. parahaemolyticus | AMP, KF, VA, P | - | 4 |
| J15Y | V. parahaemolyticus | AMP, KF, VA, P | - | 4 |
| M9-3G | V. parahaemolyticus | AMP, KF, VA, P | - | 4 |
| M9-3Y | V. parahaemolyticus | AMP, KF, VA, P | - | 4 |
| M9-4G | V. parahaemolyticus | AMP, KF, VA, P | - | 4 |
| J25Y2 | V. parahaemolyticus | AMP, KF, VA, P | - | 4 |
| P29G | V. parahaemolyticus | AMP, KF, VA, P | - | 4 |
| M2-4Y | V. parahaemolyticus | AMP, K, VA, P | - | 4 |
| J3Y2 | V. parahaemolyticus | KF, F, VA, P | - | 4 |
| J6 | V. parahaemolyticus | KF, F, VA, P | - | 4 |
| J17Y2 | V. parahaemolyticus | KF, F, VA, P | - | 4 |
| J18Y | V. parahaemolyticus | KF, F, VA, P | - | 4 |
| J9Y | V. parahaemolyticus | K, KF, VA, P | - | 4 |
| J4Y | V. parahaemolyticus | CTX, KF, VA, P | - | 4 |
| J24Y1 | V. parahaemolyticus | CTX, KF, VA, P | - | 4 |
| S2-4 | V. parahaemolyticus | AMP, VA, P | + | 3 |
| S12-3 | V. parahaemolyticus | AMP, VA, P | + | 3 |
| S15-4 | V. parahaemolyticus | AMP, VA, P | - | 3 |
| S19-3 | V. parahaemolyticus | AMP, VA, P | + | 3 |
| M4-4Y | V. parahaemolyticus | AMP, VA, P | - | 3 |
| M5-3G | V. parahaemolyticus | AMP, VA, P | + | 3 |
| P22G | V. parahaemolyticus | AMP, VA, P | - | 3 |
| J5Y | V. parahaemolyticus | KF, VA, P | - | 3 |
| J24Y2 | V. parahaemolyticus | KF, VA, P | - | 3 |
| J25Y1 | V. parahaemolyticus | KF, VA, P | - | 3 |
| K36Y1 | V. communis | AMP, K, CTX, KF, F, VA, P | + | 7 |
| K8Y1 | V. communis | AMP, K, CTX, KF, VA, P | - | 6 |
| K14Y1 | V. communis | AMP, K, CTX, KF, VA, P | - | 6 |
| K18Y1 | V. communis | AMP, K, CTX, KF, VA, P | - | 6 |
| K19Y1 | V. communis | AMP, K, CTX, KF, VA, P | - | 6 |
| K22Y | V. communis | AMP, K, CTX, KF, VA, P | + | 6 |
| K33Y2 | V. communis | AMP, K, CTX, KF, VA, P | + | 6 |
| K28Y1 | V. communis | AMP, CN, CTX, KF, VA, P | + | 6 |
| K9Y | V. communis | AMP, CTX, KF, E, VA, P | - | 6 |
| P11Y | V. communis | AMP, CTX, KF, TET, VA, P | - | 6 |
| S1-3 | V. communis | AMP, CTX, CIP, VA, P | + | 5 |
| K4Y1 | V. communis | AMP, C, CTX, VA, P | - | 5 |
| K7Y | V. communis | AMP, K, KF, VA, P | + | 5 |
| K13Y | V. communis | E, CTX, KF, VA, P | - | 5 |
| S1-4 | V. communis | AMP, CTX, VA, P | + | 4 |
| K1Y1 | V. communis | AMP, KF, VA, P | - | 4 |
| P13Y2 | V. communis | AMP, KF, VA, P | - | 4 |
| P1Y | V. communis | AMP, VA, P | - | 3 |
| P23Y2 | V. communis | AMP, VA, P | - | 3 |
| P25G | V. communis | AMP, VA, P | + | 3 |
| S7-4 | V. campbellii | AMP, CTX, KF, CIP, VA, P | + | 6 |
| S23-4 | V. campbellii | AMP, CTX, KF, CIP, VA, P | + | 6 |
| S10-4 | V. campbellii | AMP, CTX, KF, VA, P | + | 5 |
| S17-3 | V. campbellii | AMP, CTX, KF, VA, P | + | 5 |
| S18-4 | V. campbellii | AMP, CTX, KF, VA, P | + | 5 |
| S31-3 | V. campbellii | AMP, CTX, KF, VA, P | + | 5 |
| S31-4 | V. campbellii | AMP, CTX, KF, VA, P | + | 5 |
| M8-4Y | V. campbellii | AMP, CTX, CAZ, VA, P | - | 5 |
| P24G | V. campbelliii | AMP, CTX, VA, P | - | 4 |
| P28G2 | V. campbelliii | AMP, TET, VA, P, | - | 4 |
| P31Y | V. campbellii | AMP, KF, VA, P | - | 4 |
| S10-3 | V. campbellii | AMP, KF, VA, P | + | 4 |
| P21Y | V. campbellii | AMP, VA, P | - | 3 |
| P25Y1 | V. campbellii | AMP, VA, P | - | 3 |
| P32Y | V. campbellii | AMP, VA, P | - | 3 |
| P8Y | V. campbellii | AMP, K, P | - | 3 |
| P25Y2 | V. campbellii | AMP, P | - | 2 |
| P3Y1 | V. campbellii | AMP, P | - | 2 |
| T17G4 | V. owensii | AMP, FEP, CTX, CAZ, KF, K, CIP, VA, P | + | 9 |
| T34Y | V. owensii | AMP, FEP, CTX, CAZ, KF, F, E, VA, P | + | 9 |
| T20G2 | V. owensii | AMP, K, FEP, CTX, KF, CIP, VA, P | + | 8 |
| T23G1 | V. owensii | AMP, K, FEP, CTX, KF, CIP, VA, P | + | 8 |
| T23G5 | V. owensii | AMP, K, FEP, CTX, CIP, VA, P | + | 7 |
| K16Y2 | V. owensii | AMP, K, CTX, KF, F, VA, P | - | 7 |
| K38Y2 | V. owensii | AMP, K, CTX, KF, F, VA, P | + | 7 |
| M6Y | V. owensii | AMP, K, CTX, KF, F, VA, P | + | 7 |
| T5Y1 | V. owensii | AMP, FEP, CTX, KF, VA, P | + | 7 |
| K3Y2 | V. owensii | AMP, K, CTX, KF, VA, P | - | 6 |
| K16Y1 | V. owensii | AMP, K, CTX, KF, VA, P | - | 6 |
| K21Y | V. owensii | AMP, K, CTX, KF, VA, P | + | 6 |
| K38Y1 | V. owensii | AMP, CTX, F, E, VA, P | - | 6 |
| P27G | V. owensii | AMP, KF, VA, P | - | 4 |
| P34Y | V. owensii | AMP, VA, P | - | 3 |
| K34Y2 | V. rotiferianus | AMP, FEP, CTX, CAZ, TET, SXT, E, VA, P | + | 9 |
| K37Y1 | V. rotiferianus | AMP, FEP, CTX, KF, F, TET, E, VA, P | + | 9 |
| P20Y1 | V. rotiferianus | AMP, FEP, CTX, CAZ, KF, TET, VA, P | - | 8 |
| K26Y | V. rotiferianus | AMP, CTX, CAZ, TET, SXT, E, VA, P | + | 8 |
| M5-3Y | V. rotiferianus | AMP, K, CTX, KF, F, VA, P | - | 7 |
| S15-3 | V. rotiferianus | AMP, K, KF, E, VA, P | + | 6 |
| K17Y1 | V. rotiferianus | AMP, CTX, TET, VA, P | - | 5 |
| S24-4 | V. rotiferianus | AMP, VA, P | - | 3 |
| K30Y | V. rotiferianus | AMP, VA, P | + | 3 |
| P12Y | V. rotiferianus | AMP, CTX, P | - | 3 |
| K17Y2 | V. rotiferianus | TET, VA, P | - | 3 |
| P7G | V. rotiferianus | P | - | 1 |
| V1Y | V. natriegens | E, CN, K, CTX, CAZ, KF, F, VA, P | - | 9 |
| V1G2Y | V. natriegens | SXT, CTX, CAZ, KF, F, VA, P | + | 7 |
| V1Y2 | V. natriegens | K, CTX, CAZ, KF, F, VA, P | + | 7 |
| V2Y | V. natriegens | K, CTX, CAZ, KF, F, VA, P | - | 7 |
| V4Y1 | V. natriegens | K, CTX, CAZ, KF, F, VA, P | - | 7 |
| K8Y2 | V.brasiliensis | AMP, K, KF, VA, P | - | 5 |
| K40Y | V. brasiliensis | AMP, KF, E, VA, P | + | 5 |
| K6Y | V. brasiliensis | AMP, KF, VA, P | + | 4 |
| K24Y | V. brasiliensis | K, KF, VA, P | - | 4 |
| P19Y | V. harveyi | AMP, CTX, KF, F, VA, P | - | 6 |
| P23Y1 | V. harveyi | AMP, VA, P | - | 3 |
| P29Y | V. xuii | AMP, KF, VA, P | - | 4 |
| P13Y1 | V. xuii | KF, VA, P | - | 3 |
| K37Y2 | V. alginolyticus | AMP, CN, K, CTX, KF, F, TET, VA, P | + | 9 |
| K27Y1 | V. alginolyticus | AMP, K, CTX, CAZ, KF, VA, P | + | 7 |
| K36Y2 | V. alginolyticus | AMP, K, KF, F, E, VA, P | - | 7 |
| K5Y | V. alginolyticus | AMP, K, CTX, KF, VA, P | + | 6 |
| K41Y | V. alginolyticus | AMP, CTX, KF, VA, P | + | 5 |
| J3Y1 | V. alginolyticus | KF, F, VA, P | - | 4 |
| K27Y2 | V. alginolyticus | AMP, VA, P | + | 3 |
| M13Y | V. hepatarius | AMP, K, CTX, KF, F, VA, P | - | 7 |
| S6-3 | Vibrio spp. | AMP, KF, CIP, VA, P | + | 5 |
| S3-4 | Vibrio spp. | CTX, VA, P | - | 3 |
| S3-3 | Vibrio spp. | VA, P | - | 2 |
| S5-4 | Vibrio spp. | VA, P | - | 2 |
| S21-3 | Vibrio spp. | VA, P | + | 2 |
| S25-3 | Vibrio spp. | VA, P | + | 2 |
| S34-4 | Vibrio spp. | VA, P | - | 2 |
| S35-4 | Vibrio spp. | VA, P | - | 2 |
| M14-4Y | P. damselae | AMP, CN, CTX, KF, VA, P | - | 6 |
| M8-4G | P. damselae | AMP, K, CTX, KF, VA, P | - | 6 |
| S27-3 | P. damselea | AMP, K, VA, P | + | 4 |
| M2-4G | P. damselae | AMP, K, VA, P | + | 4 |
| M1-4Y | P. damselae | AMP, E, VA, P | - | 4 |
| M2-3Y | P. damselae | AMP, E, VA, P | - | 4 |
| M4Y1G | P. damselea | AMP, VA | + | 2 |
| No of Plasmid | Plasmid Profiles | Plasmid Size (kb) | No of Isolates (%) |
|---|---|---|---|
| 1 | 1 | 1.3 | 4 (3.2) |
| 2 | 3.0 | 1 (0.8) | |
| 3 | 5.0 | 2 (1.6) | |
| 4 | above 10.0 | 65 (52.0) | |
| 2 | 5 | 1.0, 3.0 | 1 (0.8) |
| 6 | 1.0, above 10.0 | 2 (1.6) | |
| 7 | 1.5, 3.0 | 1 (0.8) | |
| 8 | 2.5, above 10.0 | 2 (1.6) | |
| 9 | 3.0, 5.0 | 1 (0.8) | |
| 10 | 3.0, above 10.0 | 1 (0.8) | |
| 11 | 4.0, above 10.0 | 1 (0.8) | |
| 12 | 5.0, above 10.0 | 9 (7.2) | |
| 13 | 6.0, above 10.0 | 1 (0.8) | |
| 3 | 14 | 1.3, 3.0, above 10.0 | 2 (1.6) |
| 15 | 1.5, 2.5, above 10.0 | 1 (0.8) | |
| 16 | 2.0, 5.0, above 10.0 | 2 (1.6) | |
| 17 | 2.5, 5.0, above10.0 | 5 (4.0) | |
| 18 | 3.0, 5.0, above10.0 | 13 (10.4) | |
| 4 | 19 | 1.0, 3.0, 5.0, above 10.0 | 2 (1.6) |
| 20 | 1.5, 3.0, 5.0, above 10.0 | 2 (1.6) | |
| 5 | 21 | 1.0, 1.5, 2.0, 3.0, above 10.0 | 1 (0.8) |
| 22 | 3.0, 4.0, 5.0, 6.0, above 10.0 | 2 (1.6) | |
| 6 | 23 | 1.0, 1.5, 2.0, 3.0, 5.0, above 10.0 | 4 (3.2) |
| Total | 125 (100) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haifa-Haryani, W.O.; Amatul-Samahah, M.A.; Azzam-Sayuti, M.; Chin, Y.K.; Zamri-Saad, M.; Natrah, I.; Amal, M.N.A.; Satyantini, W.H.; Ina-Salwany, M.Y. Prevalence, Antibiotics Resistance and Plasmid Profiling of Vibrio spp. Isolated from Cultured Shrimp in Peninsular Malaysia. Microorganisms 2022, 10, 1851. https://doi.org/10.3390/microorganisms10091851
Haifa-Haryani WO, Amatul-Samahah MA, Azzam-Sayuti M, Chin YK, Zamri-Saad M, Natrah I, Amal MNA, Satyantini WH, Ina-Salwany MY. Prevalence, Antibiotics Resistance and Plasmid Profiling of Vibrio spp. Isolated from Cultured Shrimp in Peninsular Malaysia. Microorganisms. 2022; 10(9):1851. https://doi.org/10.3390/microorganisms10091851
Chicago/Turabian StyleHaifa-Haryani, Wan Omar, Md. Ali Amatul-Samahah, Mohamad Azzam-Sayuti, Yong Kit Chin, Mohd Zamri-Saad, I. Natrah, Mohammad Noor Azmai Amal, Woro Hastuti Satyantini, and Md Yasin Ina-Salwany. 2022. "Prevalence, Antibiotics Resistance and Plasmid Profiling of Vibrio spp. Isolated from Cultured Shrimp in Peninsular Malaysia" Microorganisms 10, no. 9: 1851. https://doi.org/10.3390/microorganisms10091851
APA StyleHaifa-Haryani, W. O., Amatul-Samahah, M. A., Azzam-Sayuti, M., Chin, Y. K., Zamri-Saad, M., Natrah, I., Amal, M. N. A., Satyantini, W. H., & Ina-Salwany, M. Y. (2022). Prevalence, Antibiotics Resistance and Plasmid Profiling of Vibrio spp. Isolated from Cultured Shrimp in Peninsular Malaysia. Microorganisms, 10(9), 1851. https://doi.org/10.3390/microorganisms10091851