
Miramides A–D: Identification of Detoxin-like Depsipeptides after Heterologous Expression of a Hybrid NRPS-PKS Gene Cluster from Streptomyces mirabilis Lu17588

Abstract
:1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. Cloning and Heterologous Expression of NRPS-PKS Gene Cluster
2.3. Cultivation, Metabolite Extraction and Dereplication
2.4. Genome Mining of S. Mirabilis Lu175588
2.5. Isolation and Purification of Metabolites Identified after Heterologus Expression
2.6. Nuclear Magnetic Resonance (NMR) Spectroscopy
3. Results and Discussion
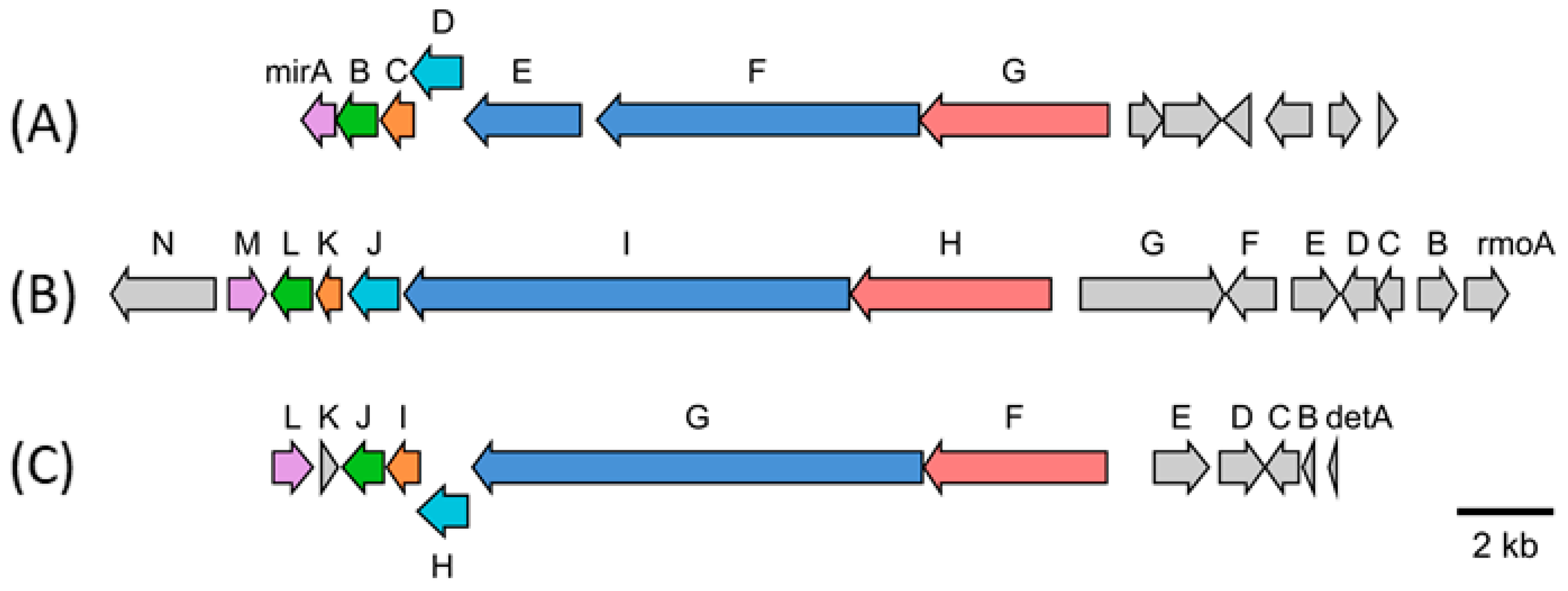
3.1. Identification and Heterologous Expression of the Hybrid NRPS-PKS Gene Cluster from S. mirabilis Lu17588 into S. albus Del14
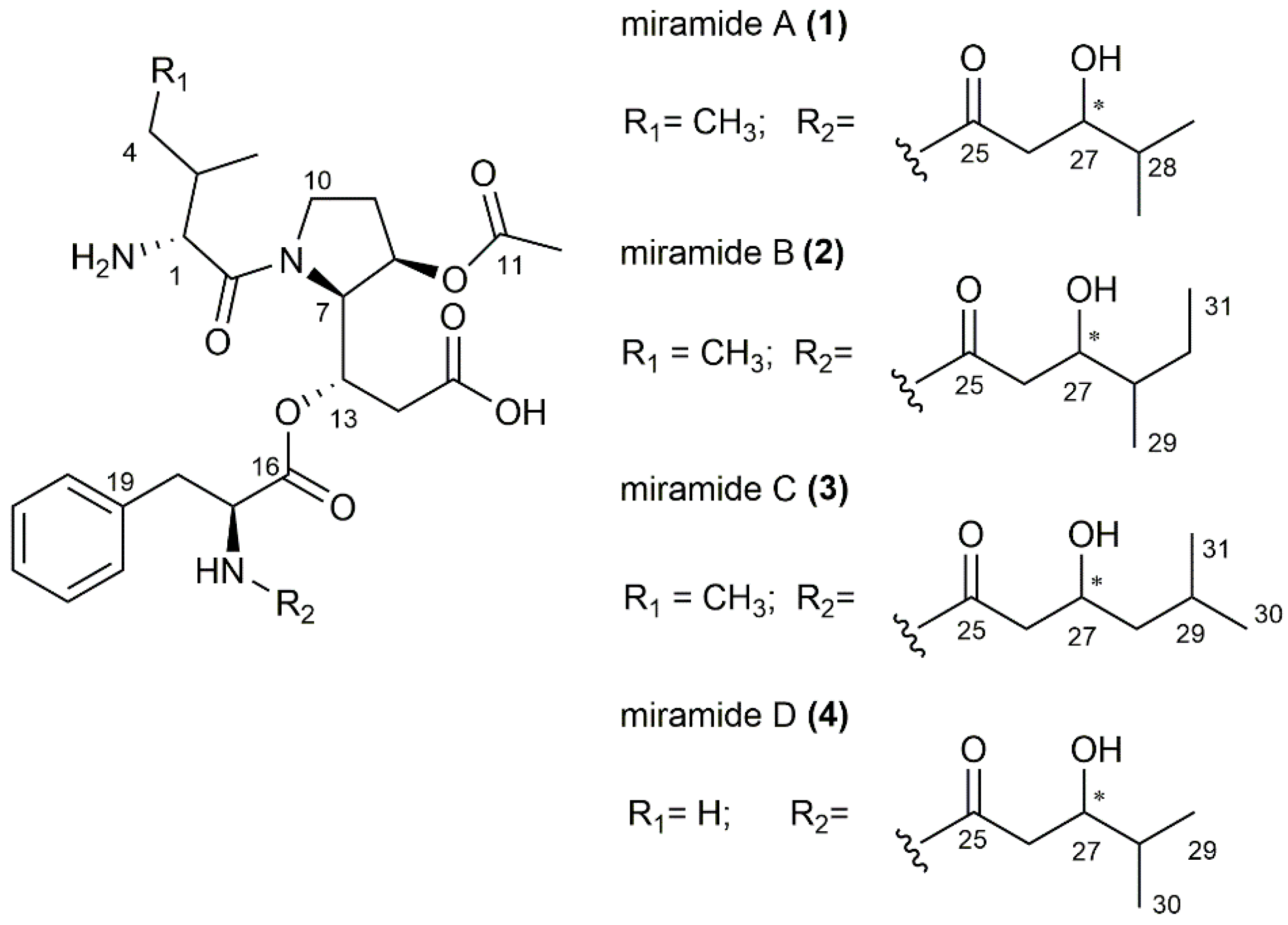
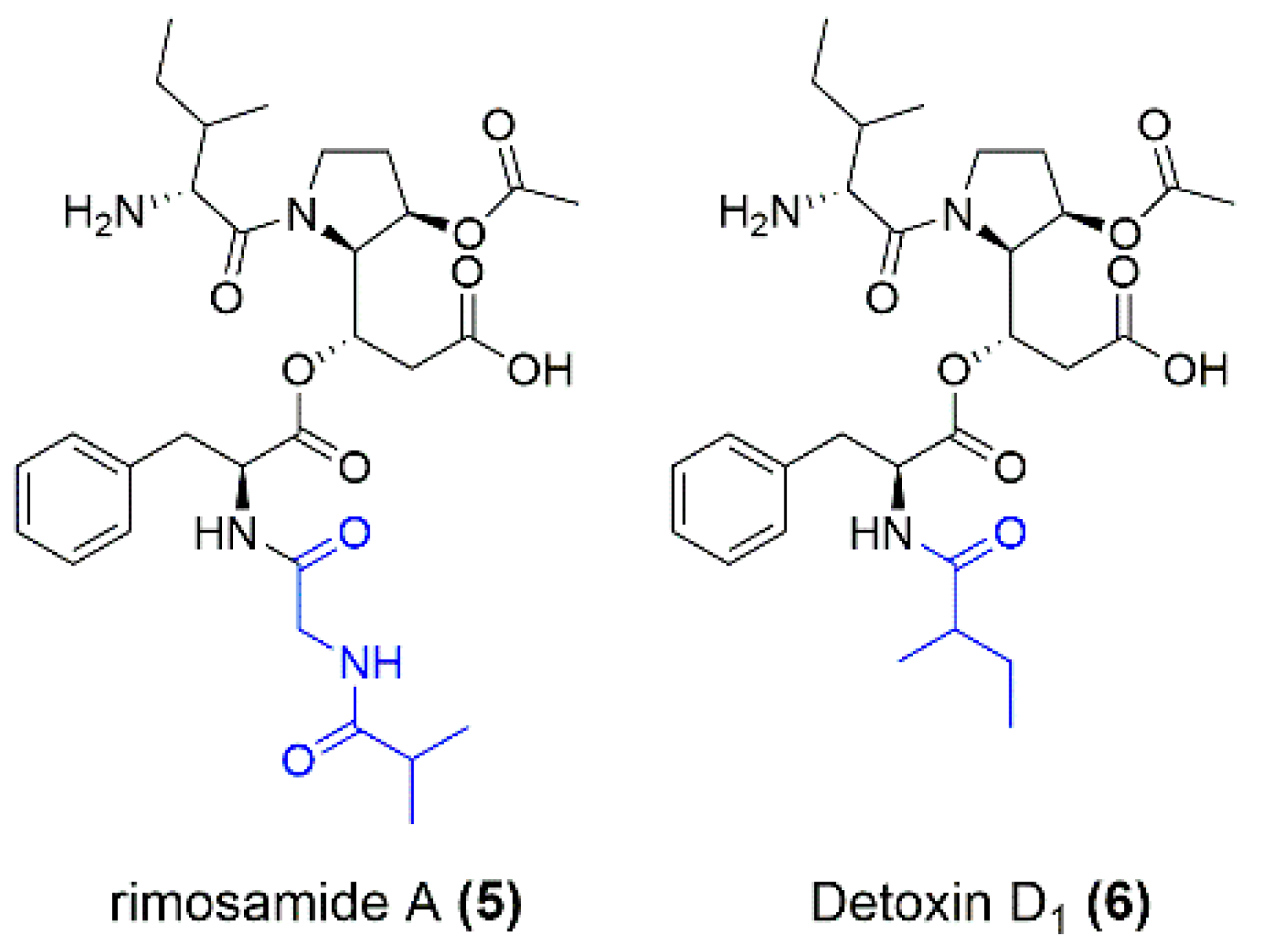
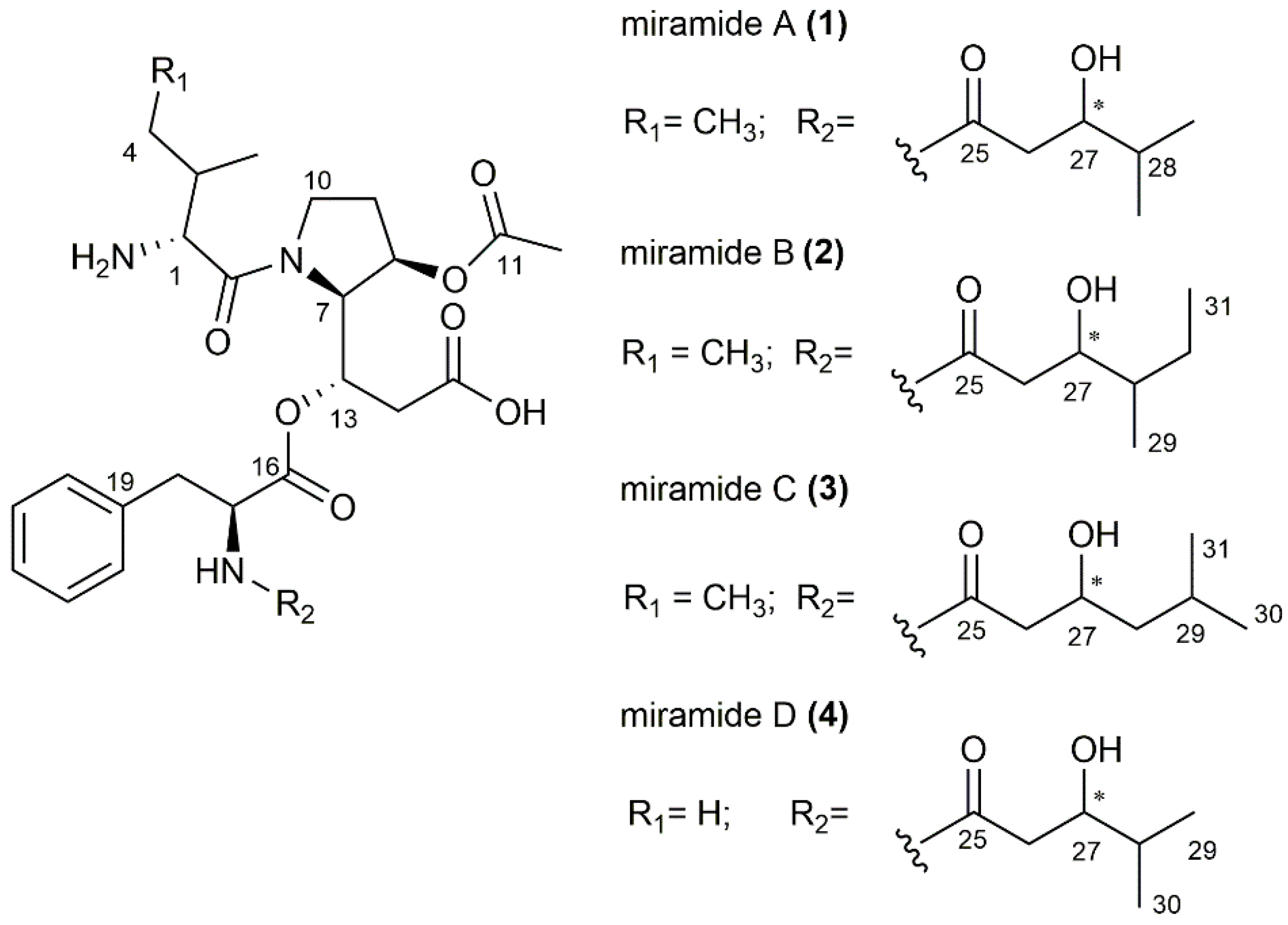
3.2. Purification and Structure Elucidation of the New Metabolites
3.3. Biosynthetic Gene Cluster and Derivation of Biosynthesis of Miramides A–D
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Reen, F.J.; Romano, S.; Dobson, A.D.W.; O’Gara, F. The Sound of Silence: Activating Silent Biosynthetic Gene Clusters in Marine Microorganisms. Mar. Drugs 2015, 13, 4754–4783. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Maclntyre, L.W.; Brady, S.F. Refactoring biosynthetic gene clusters for heterologous production of microbial natural products. Curr. Opin. Biotechnol. 2021, 69, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Hwang, S.; Kim, J.; Cho, S.; Palsson, B.; Cho, B.-K. Mini review: Genome mining approaches for the identification of secondary metabolite biosynthetic gene clusters in Streptomyces. Comput. Struct. Biotechnol. J. 2020, 18, 1548–1556. [Google Scholar] [CrossRef]
- Zhou, Z.; Xu, Q.; Bu, Q.; Guo, Y.; Liu, S.; Liu, Y.; Du, Y.; Li, Y. Genome mining-directed activation of a silent angucycline biosynthetic gene cluster in Streptomyces chattanoogensis. Chembiochem 2015, 16, 496–502. [Google Scholar] [CrossRef]
- Zhu, S.; Duan, Y.; Huang, Y. The Application of Ribosome Engineering to Natural Product Discovery and Yield Improvement in Streptomyces. Antibiotics 2019, 8, 133. [Google Scholar] [CrossRef]
- Li, Z.-Y.; Bu, Q.-T.; Wang, J.; Liu, Y.; Chen, X.-A.; Mao, X.-M.; Li, Y.-Q. Activation of anthrachamycin biosynthesis in Streptomyces chattanoogensis L10 by site-directed mutagenesis of rpoB. J. Zhejiang Univ. Sci. B 2019, 20, 983–994. [Google Scholar] [CrossRef]
- Almeida, E.L.; Kaur, N.; Jennings, L.K.; Carrillo Rincón, A.F.; Jackson, S.A.; Thomas, O.P.; Dobson, A.D.W. Genome Mining Coupled with OSMAC-Based Cultivation Reveal Differential Production of Surugamide A by the Marine Sponge Isolate Streptomyces sp. SM17 When Compared to Its Terrestrial Relative S. albidoflavus J1074. Microorganisms 2019, 7, 394. [Google Scholar] [CrossRef]
- Moon, K.; Xu, F.; Seyedsayamdost, M.R. Cebulantin, a Cryptic Lanthipeptide Antibiotic Uncovered Using Bioactivity-Coupled HiTES. Angew. Chem. 2019, 131, 6034–6038. [Google Scholar] [CrossRef]
- Seyedsayamdost, M.R. High-throughput platform for the discovery of elicitors of silent bacterial gene clusters. Proc. Natl. Acad. Sci. USA 2014, 111, 7266–7271. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Seyedsayamdost, M.R. Discovery of a Cryptic Depsipeptide from Streptomyces ghanaensis via MALDI-MS-Guided High-Throughput Elicitor Screening. Angew. Chem. Int. Ed Engl. 2020, 59, 23005–23009. [Google Scholar] [CrossRef]
- Gomez-Escribano, J.P.; Castro, J.F.; Razmilic, V.; Jarmusch, S.A.; Saalbach, G.; Ebel, R.; Jaspars, M.; Andrews, B.; Asenjo, J.A.; Bibb, M.J. Heterologous Expression of a Cryptic Gene Cluster from Streptomyces leeuwenhoekii C34T Yields a Novel Lasso Peptide, Leepeptin. Appl. Environ. Microbiol. 2019, 85, e01752-19. [Google Scholar] [CrossRef]
- Ahmed, Y.; Rebets, Y.; Estévez, M.R.; Zapp, J.; Myronovskyi, M.; Luzhetskyy, A. Engineering of Streptomyces lividans for heterologous expression of secondary metabolite gene clusters. Microb. Cell Fact. 2020, 19, 5. [Google Scholar] [CrossRef]
- Komatsu, M.; Komatsu, K.; Koiwai, H.; Yamada, Y.; Kozone, I.; Izumikawa, M.; Hashimoto, J.; Takagi, M.; Omura, S.; Shin-ya, K.; et al. Engineered Streptomyces avermitilis host for heterologous expression of biosynthetic gene cluster for secondary metabolites. ACS Synth. Biol. 2013, 2, 384–396. [Google Scholar] [CrossRef]
- Gomez-Escribano, J.P.; Bibb, M.J. Engineering Streptomyces coelicolor for heterologous expression of secondary metabolite gene clusters. Microb. Biotechnol. 2011, 4, 207–215. [Google Scholar] [CrossRef]
- Myronovskyi, M.; Rosenkränzer, B.; Nadmid, S.; Pujic, P.; Normand, P.; Luzhetskyy, A. Generation of a cluster-free Streptomyces albus chassis strains for improved heterologous expression of secondary metabolite clusters. Metab. Eng. 2018, 49, 316–324. [Google Scholar] [CrossRef]
- Li, W.R.; Han, S.Y.; Jouillié, M. The Detoxin Complex-A Naturally Occurring Safener. Heterocycles 1993, 36, 359. [Google Scholar]
- McClure, R.A.; Goering, A.W.; Ju, K.-S.; Baccile, J.A.; Schroeder, F.C.; Metcalf, W.W.; Thomson, R.J.; Kelleher, N.L. Elucidating the Rimosamide-Detoxin Natural Product Families and Their Biosynthesis Using Metabolite/Gene Cluster Correlations. ACS Chem. Biol. 2016, 11, 3452–3460. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.E.; Mehrhof, W.H.; Ernst, R.R. Limitations of thioglycolate broth as a sterility test medium for materials exposed to gaseous ethylene oxide. Appl. Microbiol. 1968, 16, 1742–1744. [Google Scholar] [CrossRef]
- Bertani, G. Lysogeny at mid-twentieth century: P1, P2, and other experimental systems. J. Bacteriol. 2004, 186, 595–600. [Google Scholar] [CrossRef]
- Larionov, V.; Kouprina, N.; Graves, J.; Chen, X.N.; Korenberg, J.R.; Resnick, M.A. Specific cloning of human DNA as yeast artificial chromosomes by transformation-associated recombination. Proc. Natl. Acad. Sci. USA 1996, 93, 491–496. [Google Scholar] [CrossRef] [Green Version]
- Kouprina, N.; Larionov, V. Exploiting the yeast Saccharomyces cerevisiae for the study of the organization and evolution of complex genomes. FEMS Microbiol. Rev. 2003, 27, 629–649. [Google Scholar] [CrossRef]
- Amberg, D.C.; Burke, D.J.; Strathern, J.N. Methods in Yeast Genetics: A Cold Spring Harbor Laboratory Course Manual, 2005 ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2005; ISBN 9780879697280. [Google Scholar]
- Synthetic Complete (SC) Medium. Cold Spring Harb. Protoc. 2016, 2016, pdb.rec090589. [CrossRef]
- Kieser, T. Practical Streptomyces Genetics; Innes: Norwich, UK, 2000; ISBN 0708406238. [Google Scholar]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef]
- Dale, J.A.; Mosher, H.S. Nuclear magnetic resonance enantiomer regents. Configurational correlations via nuclear magnetic resonance chemical shifts of diastereomeric mandelate, O-methylmandelate, and α-methoxy-α-trifluoromethylphenylacetate (MTPA) esters. J. Am. Chem. Soc. 1973, 95, 512–519. [Google Scholar] [CrossRef]
- Hoye, T.R.; Jeffrey, C.S.; Shao, F. Mosher ester analysis for the determination of absolute configuration of stereogenic (chiral) carbinol carbons. Nat. Protoc. 2007, 2, 2451–2458. [Google Scholar] [CrossRef]
- Flett, F.; Mersinias, V.; Smith, C.P. High efficiency intergeneric conjugal transfer of plasmid DNA from Escherichia coli to methyl DNA-restricting streptomycetes. FEMS Microbiol. Lett. 1997, 155, 223–229. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene; Locus Tag | Putative Product (Homologue Accession Number) | Homologue in Rmo/Det Pathway |
|---|---|---|
| mirA; LU17588_063290 | SpcZ, hypothetical protein (WP_004571773.1) | rmoM, detL |
| mirB; LU17588_063300 | Alpha-ketoglutarate-dependent taurine dioxygenase (WP_004571774.1) | rmoL, detJ |
| mirC; LU17588_063310 | Putative protein (WP_032920603.1) | rmoK, detI |
| mirD; LU17588_063320 | GSCFA family protein (WP_030179607.1) | rmoJ, detH |
| mirE; LU17588_063330 | Polyketide synthase; KR-T-TE (WP_004571777.1) | rmoI, detG |
| mirF; LU17588_063340 | Hybrid nonribosomal peptide synthetase-polyketide synthase; A-T-C-A-T-KS (WP_004571777.1) | rmoI, detG |
| mirG; LU17588_063350 | Nonribosomal peptide synthetase C-A-T-TE (WP_078586793.1) | rmoH, detF |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paulus, C.; Myronovskyi, M.; Zapp, J.; Rodríguez Estévez, M.; Lopatniuk, M.; Rosenkränzer, B.; Palusczak, A.; Luzhetskyy, A. Miramides A–D: Identification of Detoxin-like Depsipeptides after Heterologous Expression of a Hybrid NRPS-PKS Gene Cluster from Streptomyces mirabilis Lu17588. Microorganisms 2022, 10, 1752. https://doi.org/10.3390/microorganisms10091752
Paulus C, Myronovskyi M, Zapp J, Rodríguez Estévez M, Lopatniuk M, Rosenkränzer B, Palusczak A, Luzhetskyy A. Miramides A–D: Identification of Detoxin-like Depsipeptides after Heterologous Expression of a Hybrid NRPS-PKS Gene Cluster from Streptomyces mirabilis Lu17588. Microorganisms. 2022; 10(9):1752. https://doi.org/10.3390/microorganisms10091752
Chicago/Turabian StylePaulus, Constanze, Maksym Myronovskyi, Josef Zapp, Marta Rodríguez Estévez, Maria Lopatniuk, Birgit Rosenkränzer, Anja Palusczak, and Andriy Luzhetskyy. 2022. "Miramides A–D: Identification of Detoxin-like Depsipeptides after Heterologous Expression of a Hybrid NRPS-PKS Gene Cluster from Streptomyces mirabilis Lu17588" Microorganisms 10, no. 9: 1752. https://doi.org/10.3390/microorganisms10091752
APA StylePaulus, C., Myronovskyi, M., Zapp, J., Rodríguez Estévez, M., Lopatniuk, M., Rosenkränzer, B., Palusczak, A., & Luzhetskyy, A. (2022). Miramides A–D: Identification of Detoxin-like Depsipeptides after Heterologous Expression of a Hybrid NRPS-PKS Gene Cluster from Streptomyces mirabilis Lu17588. Microorganisms, 10(9), 1752. https://doi.org/10.3390/microorganisms10091752