
Ferulic Acid Esterase Producing Lactobacillus johnsonii from Goat Feces as Corn Silage Inoculants

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of FAE+ LAB from Goat Feces
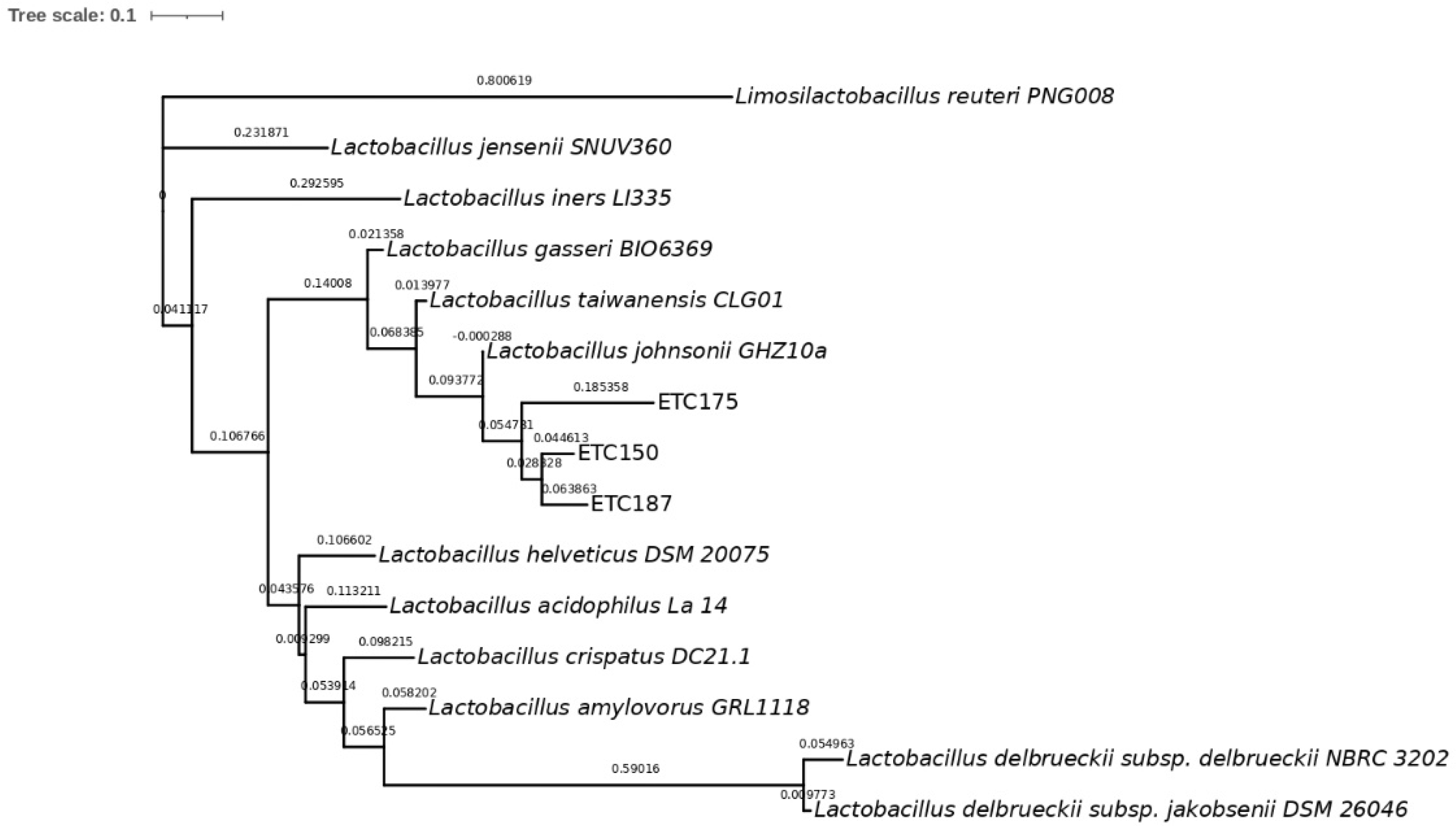
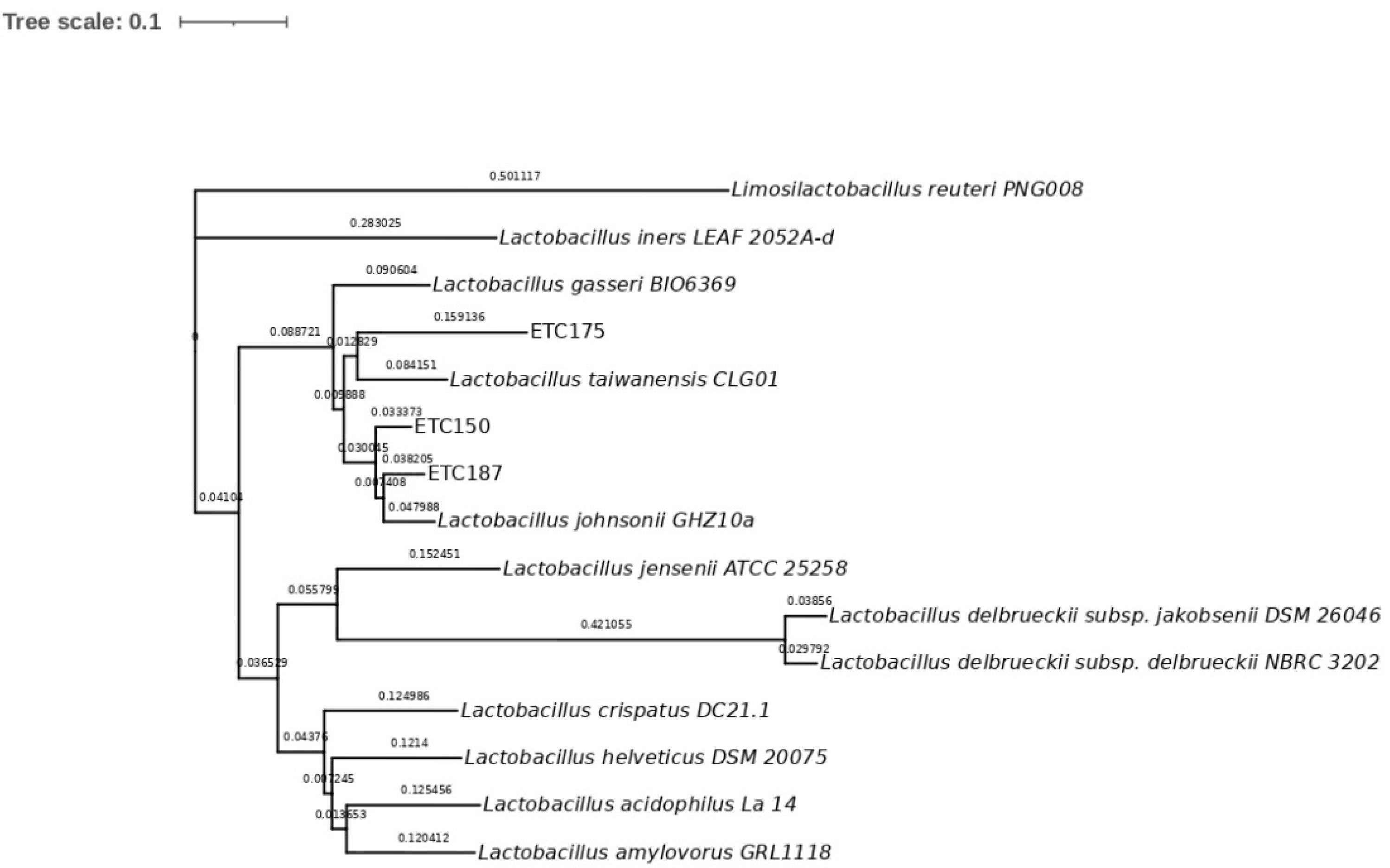
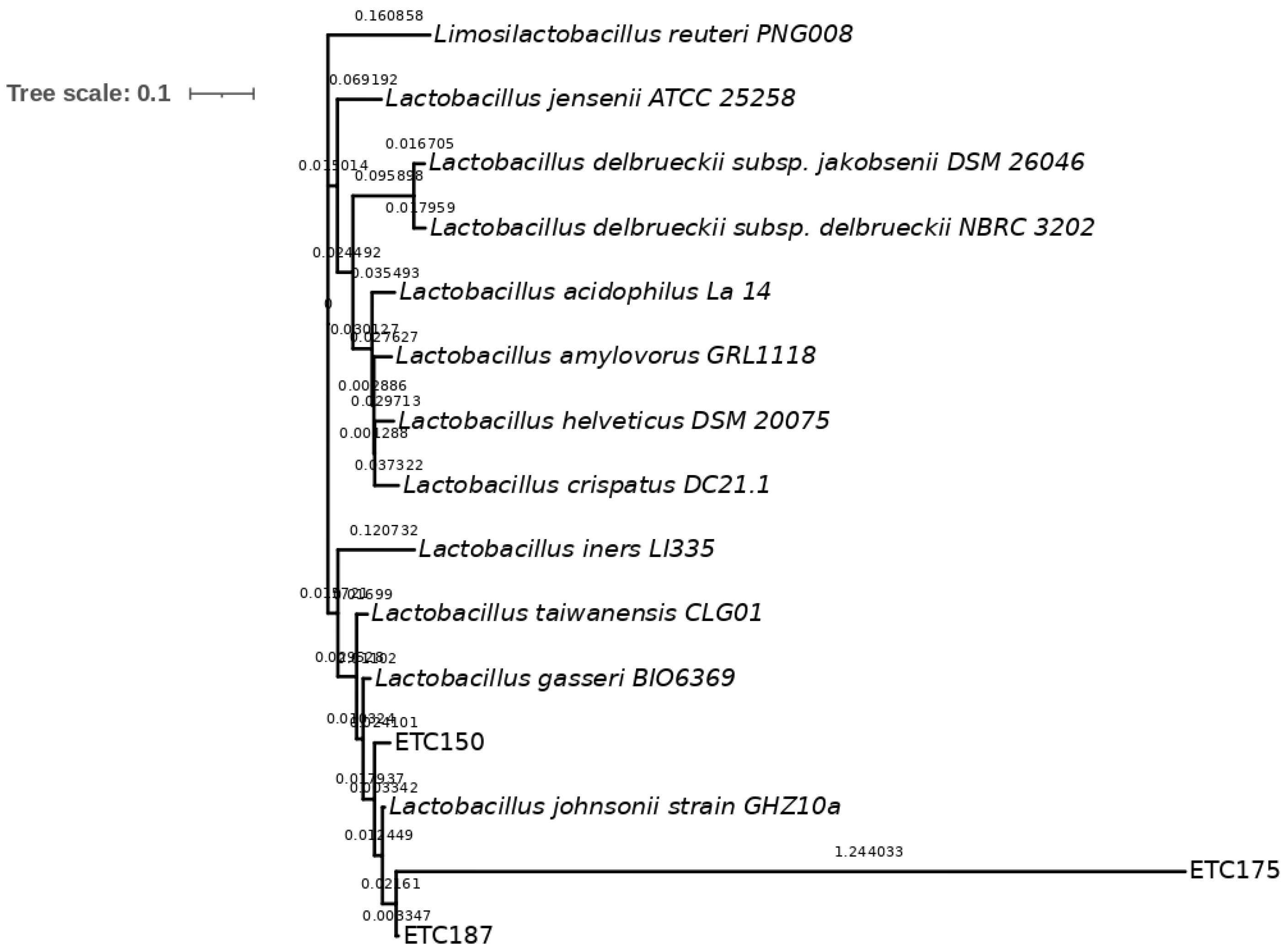
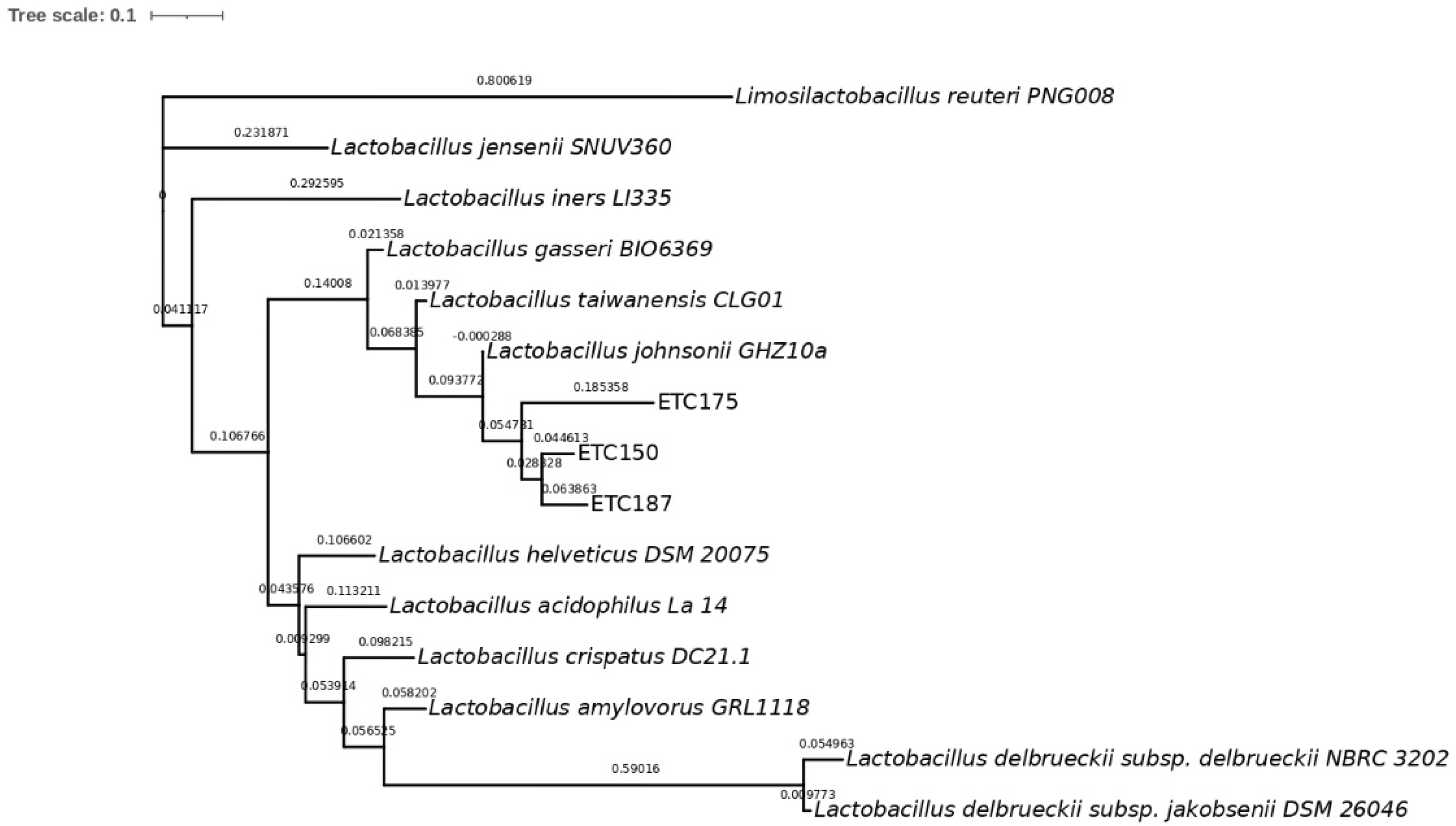
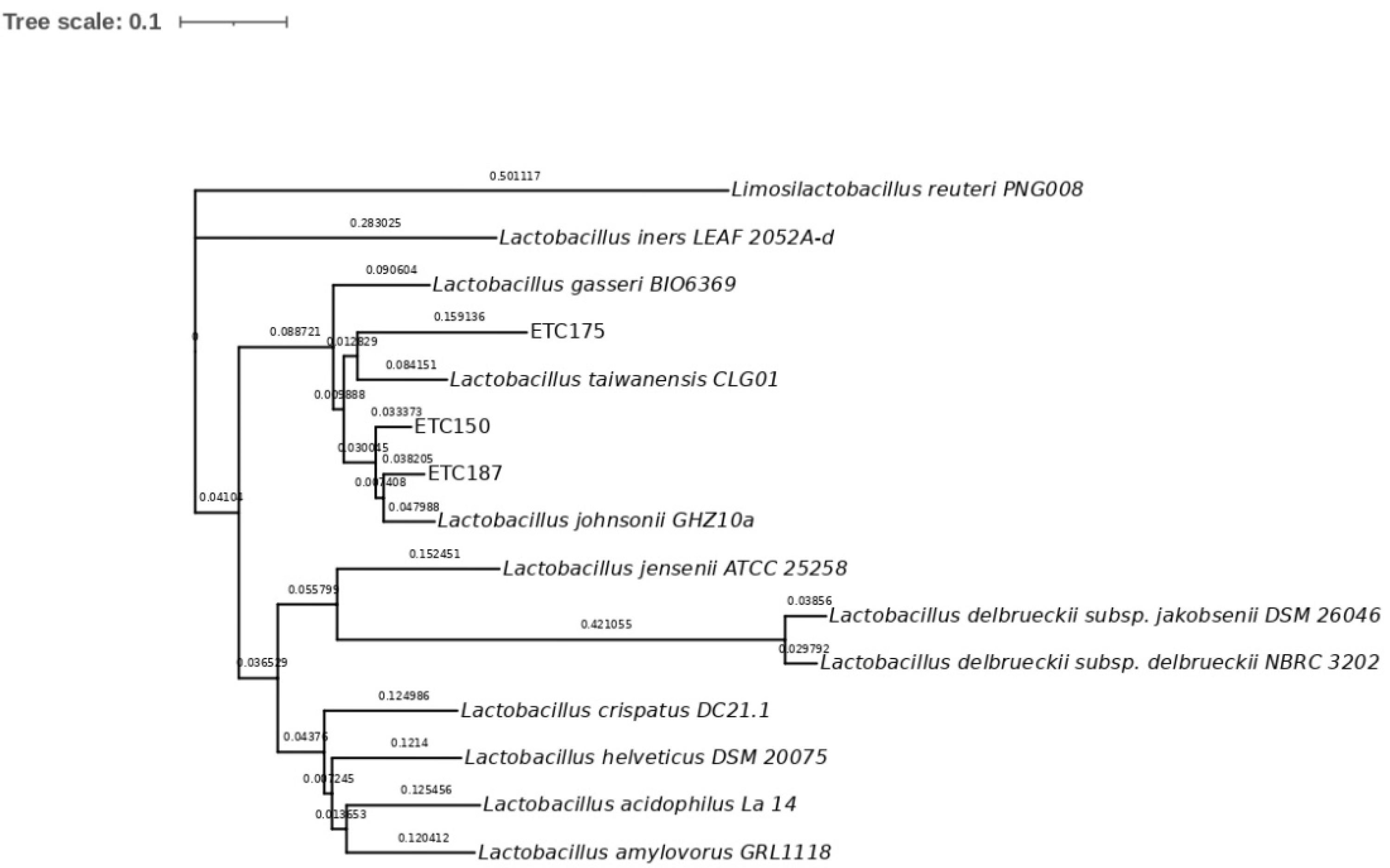
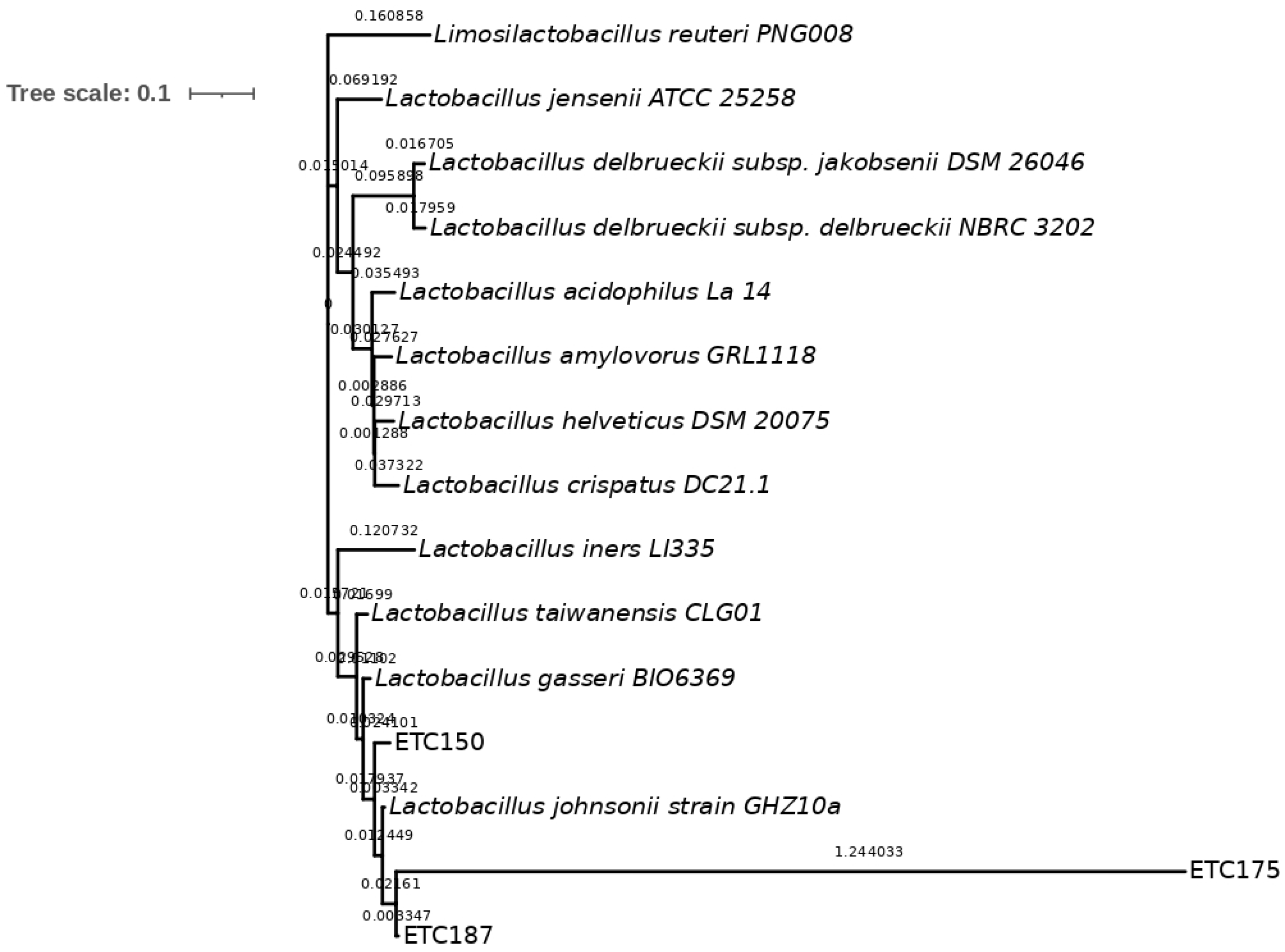
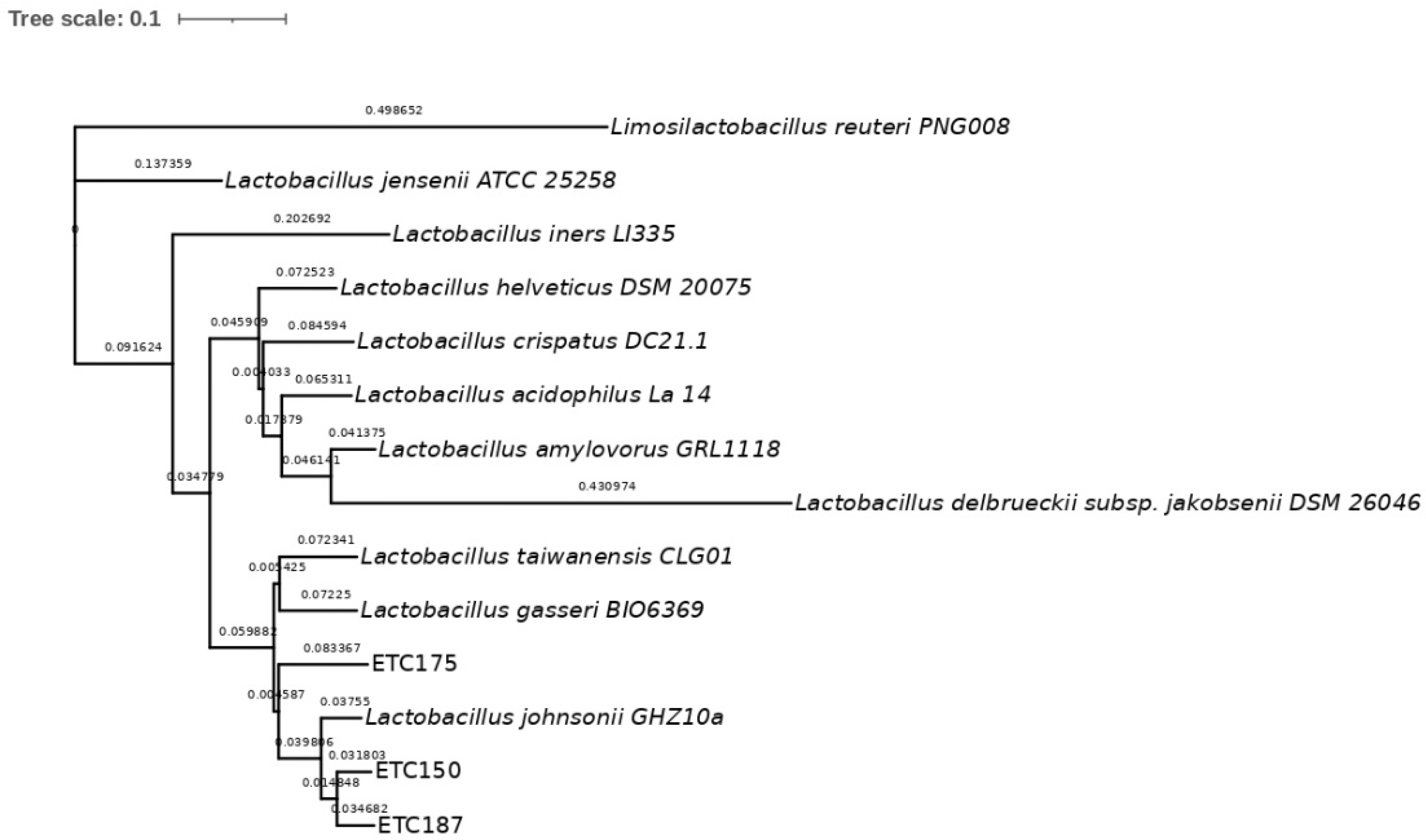
2.2. Molecular Identification and Phylogenetic Analysis of Strains
2.3. General Probiotic Characterization
2.4. FAE Activity Quantification at 37 °C
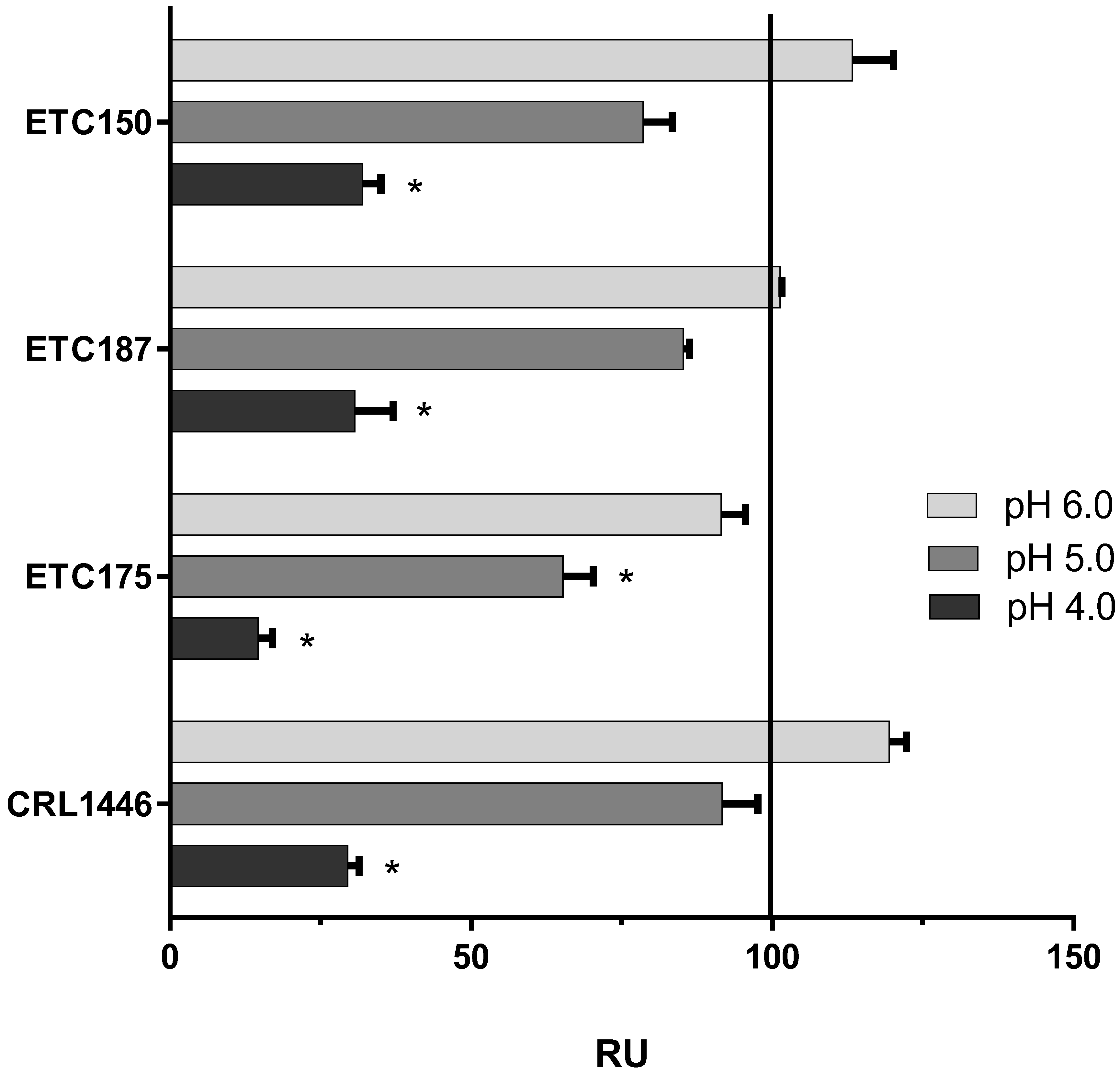
2.5. Effect of pH and Temperature on FAE Activity
2.6. Growth in Maize Soluble Fraction Medium (MSFM)
2.7. Maize Mini Silo Experiment
2.7.1. Inoculants, Forage, and Silage Preparation
2.7.2. Microbial Analysis
2.7.3. pH and Chemical Analysis
2.7.4. Compositional and Digestibility Analysis
2.8. Statistical Calculations
3. Results
3.1. Isolation of FAE+ LAB from Goat Feces
3.2. Molecular Identification of FAE+ Strains and Phylogenetic Analysis
3.3. General Probiotic Characterization
3.4. FAE Activity Quantification at 37 °C
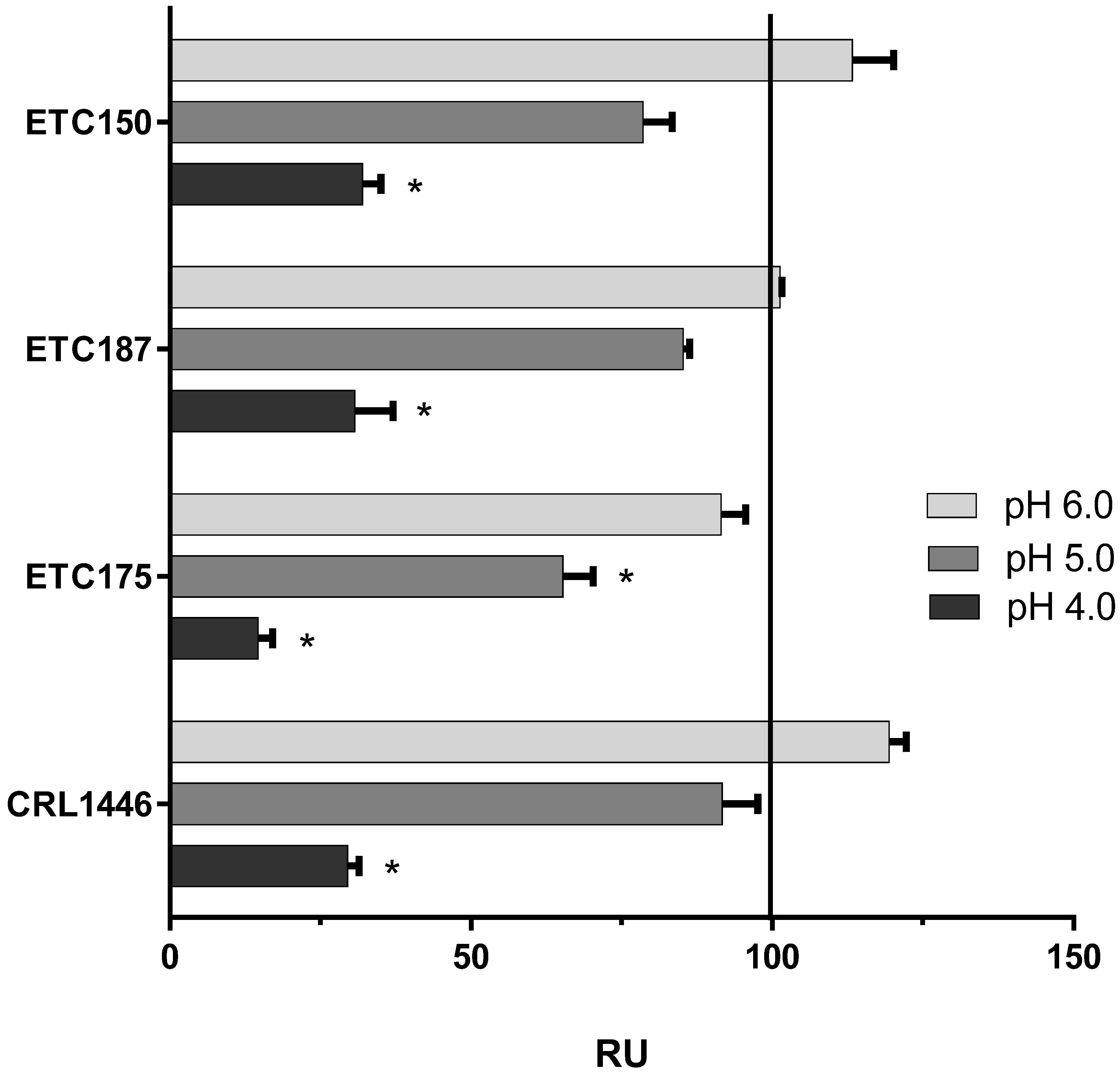
3.5. Effect of pH and Temperature on FAE Activity
3.6. Growth on MSFM
3.7. Maize Mini Silo Experiment
4. Discussion
4.1. Isolation of FAE+ LAB from Goat Feces
4.2. Molecular Identification of FAE+ Strains and Phylogenetic Analysis
4.3. General Probiotic Characterization
4.4. FAE Activity Quantification and Effect of pH and Temperature
4.5. Growth on MSFM
4.6. Maize Mini Silo Experiment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Muck, R.E.; Nadeau, E.M.G.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L., Jr. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P.; Prema, D.; Van Hamme, J.D.; Church, J.S.; Beauchemin, K.A. Fiber degradability, chemical composition and conservation characteristics of alfalfa haylage ensiled with exogenous fibrolytic enzymes and a ferulic acid esterase-producing inoculant. Can. J. Anim. Sci. 2014, 94, 697–704. [Google Scholar] [CrossRef]
- Jin, L.; Duniere, L.; Lynch, J.P.; McAllister, T.A.; Baah, J.; Wang, Y. Impact of ferulic acid esterase producing lactobacilli and fibrolytic enzymes on conservation characteristics, aerobic stability and fiber degradability of barley silage. Anim. Feed Sci. Technol. 2015, 207, 62–74. [Google Scholar] [CrossRef]
- Adesogan, A.T.; Arriola, K.G.; Jiang, Y.; Oyebade, A.; Paula, E.M.; Pech-Cervantes, A.A.; Romero, J.J.; Ferraretto, L.F.; Vyas, D. Symposium review: Technologies for improving fiber utilization. J. Dairy Sci. 2019, 102, 5726–5755. [Google Scholar] [CrossRef] [PubMed]
- Adesogan, A.; Queiroz, O.; Queiroz, M. Effects of homolactic inoculants on silage nutrient digestibility. In Proceedings of the Pre-Conference Symposium Tri-State Dairy Nutrition Conference, Fort Wayne, IN, USA, 21 April 2009. [Google Scholar]
- Nsereko, V.L.; Smiley, B.K.; Rutherford, W.M.; Spielbauer, A.; Forrester, K.J.; Hettinger, G.H.; Harman, E.K.; Harman, B.R. Influence of inoculating forage with lactic acid bacterial strains that produce ferulate esterase on ensilage and ruminal degradation of fiber. Anim. Feed Sci. Technol. 2008, 145, 122–135. [Google Scholar] [CrossRef]
- Kang, T.W.; Adesogan, A.T.; Kim, S.C.; Lee, S.S. Effects of an esterase-producing inoculant on fermentation, aerobic stability, and neutral detergent fiber digestibility of corn silage. J. Dairy Sci. 2009, 92, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Krueger, N.A.; Adesogan, A.T.; Staples, C.R.; Krueger, W.K.; Dean, D.B.; Littell, R.C. The potential to increase digestibility of tropical grasses with a fungal, ferulic acid esterase enzyme preparation. Anim. Feed Sci. Technol. 2008, 145, 95–108. [Google Scholar] [CrossRef]
- Hatfield, R.D.; Jung, H.; Marita, J.M.; Kim, H. Cell Wall Characteristics of a Maize Mutant Selected for Decreased Ferulates. Am. J. Plant Sci. 2018, 09, 446–466. [Google Scholar] [CrossRef]
- Hatfield, R.D.; Marita, J.M. Maize Development: Cell Wall Changes in Leaves and Sheaths. Am. J. Plant Sci. 2017, 08, 1248–1263. [Google Scholar] [CrossRef]
- Ou, S.; Kwok, K.C. Ferulic acid: Pharmaceutical functions, preparation and applications in foods. J. Sci. Food Agric. 2004, 84, 1261–1269. [Google Scholar] [CrossRef]
- Russo, M.; Marquez, A.; Herrera, H.; Abeijon-Mukdsi, C.; Saavedra, L.; Hebert, E.; Gauffin-Cano, P.; Medina, R. Oral administration of Lactobacillus fermentum CRL1446 improves biomarkers of metabolic syndrome in mice fed a high-fat diet supplemented with wheat bran. Food Funct. 2020, 11, 3879–3894. [Google Scholar] [CrossRef]
- González-Ríos, H.; Dávila-Ramírez, J.L.; Peña-Ramos, E.A.; Valenzuela-Melendres, M.; Zamorano-García, L.; Islava-Lagarda, T.Y.; Valenzuela-Grijalva, N.V. Dietary supplementation of ferulic acid to steers under commercial feedlot feeding conditions improves meat quality and shelf life. Anim. Feed Sci. Technol. 2016, 222, 111–121. [Google Scholar] [CrossRef]
- Li, F.; Zhang, B.; Zhang, Y.; Zhang, X.; Usman, S.; Ding, Z.; Hao, L.; Guo, X. Probiotic effect of feruloyl esterase-producing Lactobacillus plantarum inoculated alfalfa silage on digestion, antioxidant, and immunity status of lactating dairy goats. Anim. Nutr. 2022. [Google Scholar] [CrossRef]
- Samad, K.A.; Zainol, N.; Yussof, H.W.; Khushairi, Z.A.; Mohd Sharif, N.S.A.; Mohd Syukri, N.S. Isolation, identification and characterization of soil bacteria for the production of ferulic acid through co-culture fermentation using banana stem waste. SN Appl. Sci. 2020, 2, 339. [Google Scholar] [CrossRef]
- Ding, Z.T.; Xu, D.M.; Bai, J.; Li, F.H.; Adesogan, A.T.; Zhang, P.; Yuan, X.J.; Guo, X.S. Characterization and identification of ferulic acid esterase-producing Lactobacillus species isolated from Elymus nutans silage and their application in ensiled alfalfa. J. Appl. Microbiol. 2019, 127, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; He, H.; Zhang, S.; Guo, T.; Kong, J. Characterization of Feruloyl Esterases Produced by the Four Lactobacillus Species: L. amylovorus, L. acidophilus, L. farciminis and L. fermentum, Isolated from Ensiled Corn Stover. Front. Microbiol. 2017, 8, 941. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Yao, D.; Li, D.; Lin, Y.; Bureenok, S.; Ni, K.; Yang, F. Effects of Lactic Acid Bacteria Isolated From Rumen Fluid and Feces of Dairy Cows on Fermentation Quality, Microbial Community, and in vitro Digestibility of Alfalfa Silage. Front. Microbiol. 2019, 10, 2998. [Google Scholar] [CrossRef] [PubMed]
- Donaghy, J.; Kelly, P.; McKay, A. Detection of ferulic acid esterase production by Bacillus spp. and lactobacilli. Appl. Microbiol. Biotechnol. 1998, 50, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Mukdsi, M.C.A.; Haro, C.; González, S.N.; Medina, R.B. Functional goat milk cheese with feruloyl esterase activity. J. Funct. Foods 2013, 5, 801–809. [Google Scholar] [CrossRef]
- Azcárate-Peril, M.A.; Raya, R.R. Methods for Plasmid and Genomic DNA Isolation from Lactobacilli. In Food Microbiology Protocols; Spencer, J.F.T., de Ragout Spencer, A.L., Eds.; Humana Press: Totowa, NJ, USA, 2001; pp. 135–139. [Google Scholar]
- Kullen, M.; Sanozky-Dawes, R.; Crowell, D.; Klaenhammer, T. Use of the DNA sequence of variable regions of the 16S rRNA gene for rapid and accurate identification of bacteria in the Lactobacillus acidophilus complex. J. Appl. Microbiol. 2000, 89, 511–516. [Google Scholar] [CrossRef]
- Rose, R.; Golosova, O.; Sukhomlinov, D.; Tiunov, A.; Prosperi, M. Flexible design of multiple metagenomics classification pipelines with UGENE. Bioinformatics 2018, 35, 1963–1965. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Aiyar, A. The Use of CLUSTAL W and CLUSTAL X for Multiple Sequence Alignment. In Bioinformatics Methods and Protocols; Misener, S., Krawetz, S.A., Eds.; Humana Press: Totowa, NJ, USA, 1999; pp. 221–241. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Efron, B.; Halloran, E.; Holmes, S. Bootstrap confidence levels for phylogenetic trees. Proc. Natl. Acad. Sci. USA 1996, 93, 7085–7090. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Distance Methods for Inferring Phylogenies: A Justification. Evol. Int. J. Org. Evol. 1984, 38, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento-Rubiano, L.A.; Berger, B.; Moine, D.; Zuniga, M.; Perez-Martinez, G.; Yebra, M.J. Characterization of a novel Lactobacillus species closely related to Lactobacillus johnsonii using a combination of molecular and comparative genomics methods. BMC Genom. 2010, 11, 504. [Google Scholar] [CrossRef]
- Maldonado, N.C.; Ficoseco, C.A.; Mansilla, F.I.; Melián, C.; Hébert, E.M.; Vignolo, G.M.; Nader-Macías, M.E.F. Identification, characterization and selection of autochthonous lactic acid bacteria as probiotic for feedlot cattle. Livest. Sci. 2018, 212, 99–110. [Google Scholar] [CrossRef]
- Zárate, G.; Chaia, A.P.; González, S.; Oliver, G. Viability and beta-galactosidase activity of dairy propionibacteria subjected to digestion by artificial gastric and intestinal fluids. J. Food Prot. 2000, 63, 1214–1221. [Google Scholar] [CrossRef] [PubMed]
- Oliszewski, R. Leche de Cabra: Su Calidad y Evaluación de Starters Lácticos Caprinos Regionales en la Manufactura de Productos Fermentados. Ph.D. Thesis, Universidad Nacional de Tucumán, Tucumán, Argentina, 2006. [Google Scholar]
- Babot, J.D.; Argañaraz-Martínez, E.; Saavedra, L.; Apella, M.C.; Chaia, A.P. Selection of indigenous lactic acid bacteria to reinforce the intestinal microbiota of newly hatched chicken–relevance of in vitro and ex vivo methods for strains characterization. Res. Vet. Sci. 2014, 97, 8–17. [Google Scholar] [CrossRef]
- Oladipo, I.; Adebiyi, A.; Ayandele, A. Toxin production in food as influenced by pH, thermal treatment and chemical preservatives. Afr. J. Biotechnol. 2008, 7, 1731–1739. [Google Scholar] [CrossRef]
- Yue, Q.; Yang, H.J.; Li, D.H.; Wang, J.Q. A comparison of HPLC and spectrophotometrical methods to determine the activity of ferulic acid esterase in commercial enzyme products and rumen contents of steers. Anim. Feed Sci. Technol. 2009, 153, 169–177. [Google Scholar] [CrossRef]
- Saarisalo, E.; Skytta, E.; Haikara, A.; Jalava, T.; Jaakkola, S. Screening and selection of lactic acid bacteria strains suitable for ensiling grass. J. Appl. Microbiol. 2007, 102, 327–336. [Google Scholar] [CrossRef]
- Nelson, N. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem 1944, 153, 375–380. [Google Scholar] [CrossRef]
- Cabrera, C.G.; Rodriguez, G.; Palacio, M.A.; Vera, T.A.; Dib, E.P.C. Red de Innovación para el Desarrollo Rural del Gran Chaco Americano en el Contexto del Cambio Producto# 2-Protocolos de la Cadena de Valor Apícola y Ganadera; Fontagro: Washington, DC, USA, 2022. [Google Scholar]
- Isas, A.S.; Mariotti Celis, M.S.; Pérez Correa, J.R.; Fuentes, E.; Rodríguez, L.; Palomo, I.; Mozzi, F.; Van Nieuwenhove, C. Functional fermented cherimoya (Annona cherimola Mill.) juice using autochthonous lactic acid bacteria. Food Res. Int. 2020, 138, 109729. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Xin, H.; Paengkoum, P.; Paengkoum, S.; Ban, C.; Sorasak, T. Effects of anthocyanin-rich purple corn (Zea mays L.) stover silage on nutrient utilization, rumen fermentation, plasma antioxidant capacity, and mammary gland gene expression in dairy goats1. J. Anim. Sci. 2018, 97, 1384–1397. [Google Scholar] [CrossRef]
- Zhuang, Y.; Chai, J.; Cui, K.; Bi, Y.; Diao, Q.; Huang, W.; Usdrowski, H.; Zhang, N. Longitudinal Investigation of the Gut Microbiota in Goat Kids from Birth to Postweaning. Microorganisms 2020, 8, 1111. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Versión 2011; Grupo Infostat, FCA, Universidad Nacional de Córdoba: Córdoba, Argentina, 2011. [Google Scholar]
- Russo, M.I. Empleo de Bacterias Lácticas con Actividad Cinamil Esterasa como Estrategia Nutricional para la Prevención del Síndrome Metabólico. Ph.D. Thesis, Universidad Nacional de Tucumán, Tucumán, Argentina, 2019. [Google Scholar]
- Draksler, D.; Locascio, M.; González, S.; Oliver, G. The development of faecal flora in young Creole goats. Small Rumin. Res. 2002, 46, 67–70. [Google Scholar] [CrossRef]
- Apas, A.L.; Dupraz, J.; Ross, R.; Gonzalez, S.N.; Arena, M.E. Probiotic administration effect on fecal mutagenicity and microflora in the goat’s gut. J. Biosci. Bioeng. 2010, 110, 537–540. [Google Scholar] [CrossRef]
- Wang, L.T.; Kuo, H.P.; Wu, Y.C.; Tai, C.J.; Lee, F.L. Lactobacillus taiwanensis sp. nov., isolated from silage. Int. J. Syst. Evol. Microbiol. 2009, 59, 2064–2068. [Google Scholar] [CrossRef] [PubMed]
- Nader-Macias, M.E.; Otero, M.C.; Espeche, M.C.; Maldonado, N.C. Advances in the design of probiotic products for the prevention of major diseases in dairy cattle. J. Ind. Microbiol. Biotechnol. 2008, 35, 1387–1395. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P.; Baah, J.; Beauchemin, K.A. Conservation, fiber digestibility, and nutritive value of corn harvested at 2 cutting heights and ensiled with fibrolytic enzymes, either alone or with a ferulic acid esterase-producing inoculant. J. Dairy Sci. 2015, 98, 1214–1224. [Google Scholar] [CrossRef] [PubMed]
- Addah, W.; Baah, J.; Okine, E.; McAllister, T. A third-generation esterase inoculant alters fermentation pattern and improves aerobic stability of barley silage and the efficiency of body weight gain of growing feedlot cattle1. J. Anim. Sci. 2012, 90, 1541–1552. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; He, H.; Zhang, S.; Kong, J. Effects of inoculants Lactobacillus brevis and Lactobacillus parafarraginis on the fermentation characteristics and microbial communities of corn stover silage. Sci. Rep. 2017, 7, 13614. [Google Scholar] [CrossRef] [PubMed]
- Su, R.; Ni, K.; Wang, T.; Yang, X.; Zhang, J.; Liu, Y.; Shi, W.; Yan, L.; Jie, C.; Zhong, J. Effects of ferulic acid esterase-producing Lactobacillus fermentum and cellulase additives on the fermentation quality and microbial community of alfalfa silage. PeerJ 2019, 7, e7712. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Bajagai, Y.S.; Klieve, A.V.; Dart, P.J.; Bryden, W.L. Probiotics in Animal Nutrition: Production, Impact and Regulation; FAO: Rome, Italy, 2016. [Google Scholar]
- Elghandour, M.M.Y.; Salem, A.Z.M.; Castañeda, J.S.M.; Camacho, L.M.; Kholif, A.E.; Chagoyán, J.C.V. Direct-fed microbes: A tool for improving the utilization of low quality roughages in ruminants. J. Integr. Agric. 2015, 14, 526–533. [Google Scholar] [CrossRef]
- Draksler, D.; Gonzales, S.; Oliver, G. Preliminary assays for the development of a probiotic for goats. Reprod. Nutr. Dev. 2004, 44, 397–405. [Google Scholar] [CrossRef]
- Apas, A.L.; Arena, M.E.; Elias, A.G.; González, F.; Gonzalez, S.N. Beneficial Effects of Fermented Sugarcane Residue with Goat Probiotic On Gut Health. Int. J. Agric. Innov. Res. 2015. [Google Scholar]
- EFSA. Opinion of the Scientific Committee on a request from EFSA on the introduction of a Qualified Presumption of Safety (QPS) approach for assessment of selected microorganisms referred to EFSA. EFSA J. 2007, 5, 587. [Google Scholar] [CrossRef]
- Weinberg, Z.G.; Muck, R.E.; Weimer, P.J.; Chen, Y.; Gamburg, M. Lactic acid bacteria used in inoculants for silage as probiotics for ruminants. Appl. Biochem. Biotechnol. 2004, 118, 1–9. [Google Scholar] [CrossRef]
- Han, H.; Ogata, Y.; Yamamoto, Y.; Nagao, S.; Nishino, N. Identification of lactic acid bacteria in the rumen and feces of dairy cows fed total mixed ration silage to assess the survival of silage bacteria in the gut. J. Dairy Sci. 2014, 97, 5754–5762. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, Z.; Muck, R.; Weimer, P. The survival of silage inoculant lactic acid bacteria in rumen fluid. J. Appl. Microbiol. 2003, 94, 1066–1071. [Google Scholar] [CrossRef] [PubMed]
- Naissinger da Silva, M.; Tagliapietra, B.L.; Flores, V.d.A.; Pereira dos Santos Richards, N.S. In vitro test to evaluate survival in the gastrointestinal tract of commercial probiotics. Curr. Res. Food Sci. 2021, 4, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Abeijón Mukdsi, M.C.; Medina, R.B.; Alvarez, M.d.F.; González, S.N. Ester synthesis by lactic acid bacteria isolated from goat’s and ewe’s milk and cheeses. Food Chem. 2009, 117, 241–247. [Google Scholar] [CrossRef]
- Couteau, D.; McCartney, A.L.; Gibson, G.R.; Williamson, G.; Faulds, C.B. Isolation and characterization of human colonic bacteria able to hydrolyse chlorogenic acid. J. Appl. Microbiol. 2001, 90, 873–881. [Google Scholar] [CrossRef]
- Ramos-de-la-Peña, A.M.; Contreras-Esquivel, J.C. Methods and substrates for feruloyl esterase activity detection, a review. J. Mol. Catal. B Enzym. 2016, 130, 74–87. [Google Scholar] [CrossRef]
- Fritsch, C.; Jansch, A.; Ehrmann, M.A.; Toelstede, S.; Vogel, R.F. Characterization of Cinnamoyl Esterases from Different Lactobacilli and Bifidobacteria. Curr. Microbiol. 2017, 74, 247–256. [Google Scholar] [CrossRef]
- Kim, J.-H.; Baik, S.-H. Probiotic properties of Lactobacillus strains with high cinnamoyl esterase activity isolated from jeot-gal, a high-salt fermented seafood. Ann. Microbiol. 2019, 69, 407–417. [Google Scholar] [CrossRef]
- Costa, D.M.; Carvalho, B.F.; Bernardes, T.F.; Schwan, R.F.; Ávila, C.L.D.S. New epiphytic strains of lactic acid bacteria improve the conservation of corn silage harvested at late maturity. Anim. Feed Sci. Technol. 2021, 274, 114852. [Google Scholar] [CrossRef]
- Avila, C.L.; Carvalho, B.F.; Pinto, J.C.; Duarte, W.F.; Schwan, R.F. The use of Lactobacillus species as starter cultures for enhancing the quality of sugar cane silage. J. Dairy Sci. 2014, 97, 940–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, P.; Borgo, M.F.; Binetti, A.; Puntillo, M.; Bergamini, C.; Paez, R.; Mazzoni, R.; Reinheimer, J.; Vinderola, G. Isolation, Characterization and Performance of Autochthonous Spray Dried Lactic Acid Bacteria in Maize Micro and Bucket-Silos. Front. Microbiol. 2018, 9, 2861. [Google Scholar] [CrossRef] [PubMed]
- Borreani, G.; Tabacco, E.; Schmidt, R.J.; Holmes, B.J.; Muck, R.E. Silage review: Factors affecting dry matter and quality losses in silages. J. Dairy Sci. 2018, 101, 3952–3979. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, O.C.; Arriola, K.G.; Daniel, J.L.; Adesogan, A.T. Effects of 8 chemical and bacterial additives on the quality of corn silage. J. Dairy Sci. 2013, 96, 5836–5843. [Google Scholar] [CrossRef]
- Comino, L.; Tabacco, E.; Righi, F.; Revello-Chion, A.; Quarantelli, A.; Borreani, G. Effects of an inoculant containing a Lactobacillus buchneri that produces ferulate-esterase on fermentation products, aerobic stability, and fibre digestibility of maize silage harvested at different stages of maturity. Anim. Feed Sci. Technol. 2014, 198, 94–106. [Google Scholar] [CrossRef]
- De Boever, J.; Dupon, E.; Vliegher, A.D.; Campeneere, S.D.; Latré, L. Can lactobacilli producing ferulate esterase improve the nutritive value of grass and maize silage? In Proceedings of the Grassland and Forages In High Output Dairy Farming Systems, 18th Symposium of the European Grassland Federation, Wageningen, The Netherlands, 15–17 June 2015; pp. 190–193. [Google Scholar]
- Li, F.; Ke, W.; Ding, Z.; Bai, J.; Zhang, Y.; Xu, D.; Li, Z.; Guo, X. Pretreatment of Pennisetum sinese silages with ferulic acid esterase-producing lactic acid bacteria and cellulase at two dry matter contents: Fermentation characteristics, carbohydrates composition and enzymatic saccharification. Bioresour. Technol. 2020, 295, 122261. [Google Scholar] [CrossRef]
- Xu, D.; Ding, Z.; Bai, J.; Ke, W.; Zhang, Y.; Li, F.; Guo, X. Evaluation of the effect of feruloyl esterase-producing Lactobacillus plantarum and cellulase pretreatments on lignocellulosic degradation and cellulose conversion of co-ensiled corn stalk and potato pulp. Bioresour. Technol. 2020, 310, 123476. [Google Scholar] [CrossRef]
- Xie, Y.; Guo, J.; Li, W.; Wu, Z.; Yu, Z. Effects of Ferulic Acid Esterase-Producing Lactic Acid Bacteria and Storage Temperature on the Fermentation Quality, In Vitro Digestibility and Phenolic Acid Extraction Yields of Sorghum (Sorghum bicolor L.) Silage. Microorganisms 2021, 9, 114. [Google Scholar] [CrossRef]
- Machado, E.; Matumoto Pintro, P.T.; Itavo, L.C.V.; Agustinho, B.C.; Daniel, J.L.P.; Santos, N.W.; Bragatto, J.M.; Ribeiro, M.G.; Zeoula, L.M. Reduction in lignin content and increase in the antioxidant capacity of corn and sugarcane silages treated with an enzymatic complex produced by white rot fungus. PLoS ONE 2020, 15, e0229141. [Google Scholar] [CrossRef]
- Taboada, A.; Novo-Uzal, E.; Flores, G.; Loureda, M.; Ros Barcelo, A.; Masa, A.; Pomar, F. Digestibility of silages in relation to their hydroxycinnamic acid content and lignin composition. J. Sci. Food Agric. 2010, 90, 1155–1162. [Google Scholar] [CrossRef]
- Puntillo, M.; Gaggiotti, M.; Oteiza, J.M.; Binetti, A.; Massera, A.; Vinderola, G. Potential of Lactic Acid Bacteria Isolated From Different Forages as Silage Inoculants for Improving Fermentation Quality and Aerobic Stability. Front. Microbiol. 2020, 11, 586716. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.; Wang, Y.; Li, D.; Cai, Y.; Pang, H. Characterization, identification and application of lactic Acid bacteria isolated from forage paddy rice silage. PLoS ONE 2015, 10, e0121967. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Fragment Size (bp) | Reference |
|---|---|---|---|
| pheS F | 5′-AAACTAGGGTGGTACCGCGA-3′ | 398 | This study |
| pheS R | 5′-GCTTTGAACCTAAATGTCCTTCAC-3′ | This study | |
| gyrB F | 5′-TCACATTTCTGCTGGAACGAT-3′ | 458 | This study |
| gyrB R | 5′-CTACAGAAGCACCAACACCGT-3′ | This study | |
| pyrG F | 5′-TTATGTTACYGAYGATGGTAC-3′ | 908 | Sarmiento Rubiano et al. [30] |
| pyrG R | 5′-ACCACGWGTACCAAAACCAC-3′ | [30] | |
| tuf F | 5′-ATGGCAGAAAAAGAACATTACG-3′ | 1176 | [30] |
| tuf R | 5′-AGTAACYTGACCRGCACCAAC-3′ | [30] |
| Temperature (°C) | Strain | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| ETC150 | ETC187 | ETC175 | CRL1446 | |||
| 37 | 218 Ba | 591 c | 211 Ba | 330 Bb | 9.5 | <0.0001 |
| 18 | 168 Aa,b | 513 c | 106 Aa | 216 Ab | ||
| SEM | 13.5 | |||||
| p-value | <0.0001 | |||||
| Item 1 | Time (h) | ETC150 | ETC175 | ETC187 | CRL1446 | SEM | p-Value |
|---|---|---|---|---|---|---|---|
| ∆pH | 12 | 0.2 a | 0.2 a | 0.6 b | 0.5 b | 0.02 | <0.001 |
| 24 | 0.8 a | 0.6 a | 1.6 b | 1.4 b | 0.06 | <0.001 | |
| 48 | 2.12 a | 2.29 b | 2.71 d | 2.46 c | 0.03 | <0.0001 | |
| RS | 48 | 21 b | 23.9 b | 7.3 a | 19.6 b | 1.12 | <0.01 |
| ∆Log CFU mL−1 | 48 | 4.72 b | 3.62 a | 4.63 b | 4.79 b | 0.05 | <0.01 |
| Item | UN | ETC150 | ETC175 | ETC187 | CRL1446 | SEM | p-Value | |
|---|---|---|---|---|---|---|---|---|
| pH | 3.83 a,b | 3.86 b | 3.77 a | 3.85 b | 3.82 a,b | 0.01 | 0.02 | * |
| Gas loss (%) | 9.8 b | 4 a | 3.5 a | 3 a | 2.3 a | 0.06 | <0.01 | ** |
| Microbial populations, Log UFC g FM−1 | ||||||||
| Lactic acid bacteria | 5.7 | 6.2 | 5.9 | 5.3 | 5.8 | 0.35 | 0.598 | ns |
| Total mesophilic bacteria | 5.0 | 5.3 | 5.4 | 5.4 | 6.6 | 0.2 | 0.053 | t |
| Yeasts | 4.8 | 3.3 | 3.3 | 2.7 | 3.6 | 0.35 | 0.062 | t |
| Fermentation products, g kg DM−1 | ||||||||
| Lactate | 39.1 | 36.1 | 40.9 | 38.1 | 30.3 | 1.38 | 0.07 | t |
| Acetate | 19.8 | 14.4 | 15.6 | 17.3 | 15.8 | 1.29 | 0.83 | ns |
| Total volatile acids | 59 | 51 | 57 | 55 | 46 | 4.8 | 0.45 | ns |
| Ethanol | 1.0 a,b | 0.2 a | 0.2 a | 0.1 a | 3.75 b | 0.5 | <0.05 | ns |
| Compositional and nutritional analysis, g kg DM−1 | ||||||||
| DM loss | 94 b | 20.5 a | 85.5 b | 71 b | 26.5 a | 7.32 | <0.01 | ** |
| Crude protein | 45 | 47 | 47 | 46 | 52 | 0.97 | 0.22 | ns |
| aNDF | 568 | 528 | 548 | 552 | 530 | 12 | 0.43 | ns |
| NDFD | 327 | 294 | 317 | 302 | 289 | 10 | 0.17 | ns |
| ADF | 269 c | 226 a | 255 b,c | 235 a,b | 231 a | 5.54 | <0.01 | ** |
| DMD | 607 a | 689 a,b | 631 a,b | 672 a,b | 703 b | 18 | <0.05 | * |
| Antioxidant determinations | ||||||||
| Ferulic acid (g kg DM−1) | 135 a | 144 a,b | 211 b | 139 a,b | 95 a | 15 | <0.01 | ** |
| DPPH scavenging activity (%) | 85 | 86 | 88 | 88 | 84 | 1.6 | 0.82 | ns |
| TPC (mg Eq AG 100 g−1) | 51 b,c | 52 c | 48.6 a,b | 46.4 a | 47.8 a | 0.5 | <0.01 | ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrada, E.; Mechoud, M.A.; Abeijón-Mukdsi, M.C.; Chagra Dib, E.P.; Cerviño, S.; Perez Chaia, A.; Medina, R.B. Ferulic Acid Esterase Producing Lactobacillus johnsonii from Goat Feces as Corn Silage Inoculants. Microorganisms 2022, 10, 1732. https://doi.org/10.3390/microorganisms10091732
Andrada E, Mechoud MA, Abeijón-Mukdsi MC, Chagra Dib EP, Cerviño S, Perez Chaia A, Medina RB. Ferulic Acid Esterase Producing Lactobacillus johnsonii from Goat Feces as Corn Silage Inoculants. Microorganisms. 2022; 10(9):1732. https://doi.org/10.3390/microorganisms10091732
Chicago/Turabian StyleAndrada, Estefania, Mónica Adriana Mechoud, María Claudia Abeijón-Mukdsi, Elsa Patricia Chagra Dib, Santiago Cerviño, Adriana Perez Chaia, and Roxana Beatriz Medina. 2022. "Ferulic Acid Esterase Producing Lactobacillus johnsonii from Goat Feces as Corn Silage Inoculants" Microorganisms 10, no. 9: 1732. https://doi.org/10.3390/microorganisms10091732
APA StyleAndrada, E., Mechoud, M. A., Abeijón-Mukdsi, M. C., Chagra Dib, E. P., Cerviño, S., Perez Chaia, A., & Medina, R. B. (2022). Ferulic Acid Esterase Producing Lactobacillus johnsonii from Goat Feces as Corn Silage Inoculants. Microorganisms, 10(9), 1732. https://doi.org/10.3390/microorganisms10091732