
The Genus Iodidimonas: From Its Discovery to Potential Applications


Abstract
1. Introduction
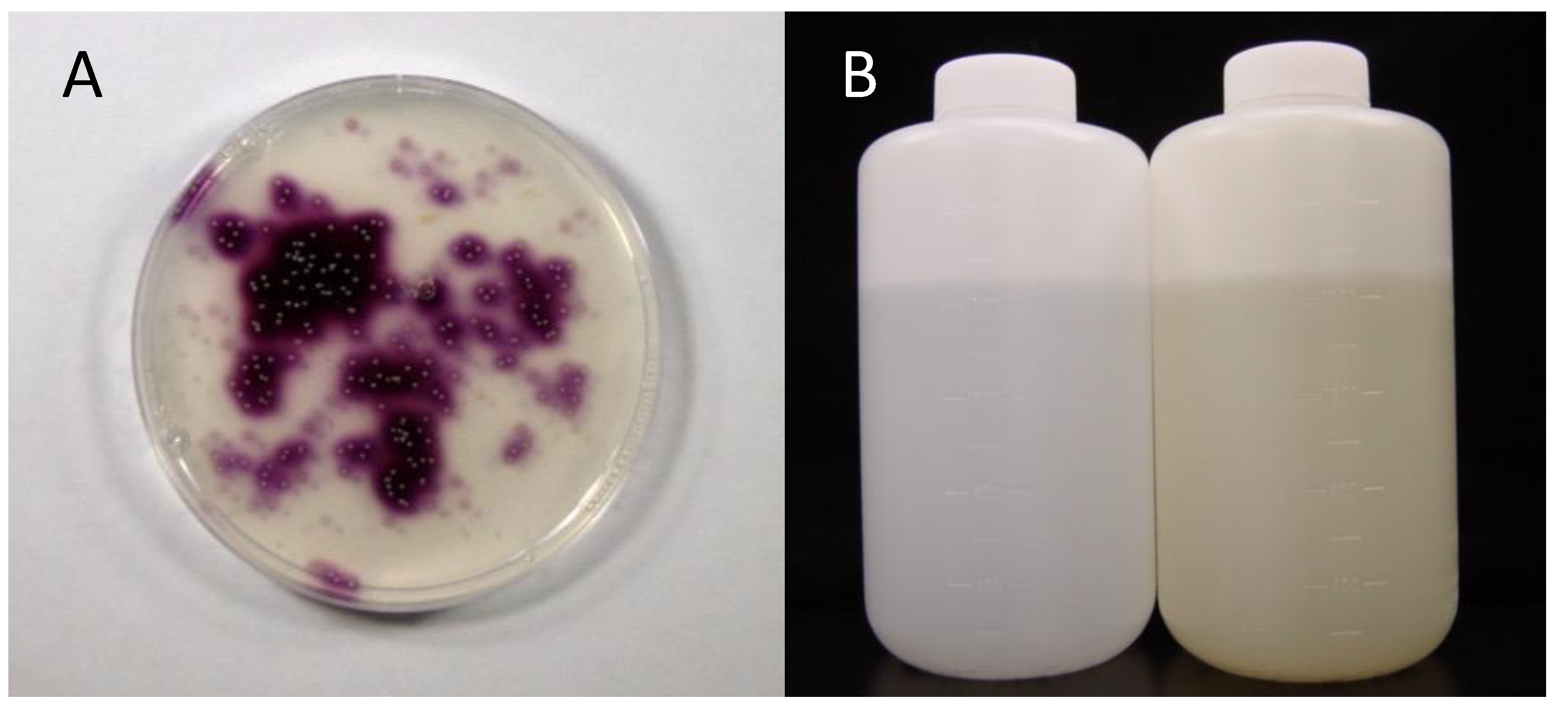
2. Discovery of Iodidimonas from Natural Gas Brine Waters
3. Enrichment of Iodidimonas in Seawater
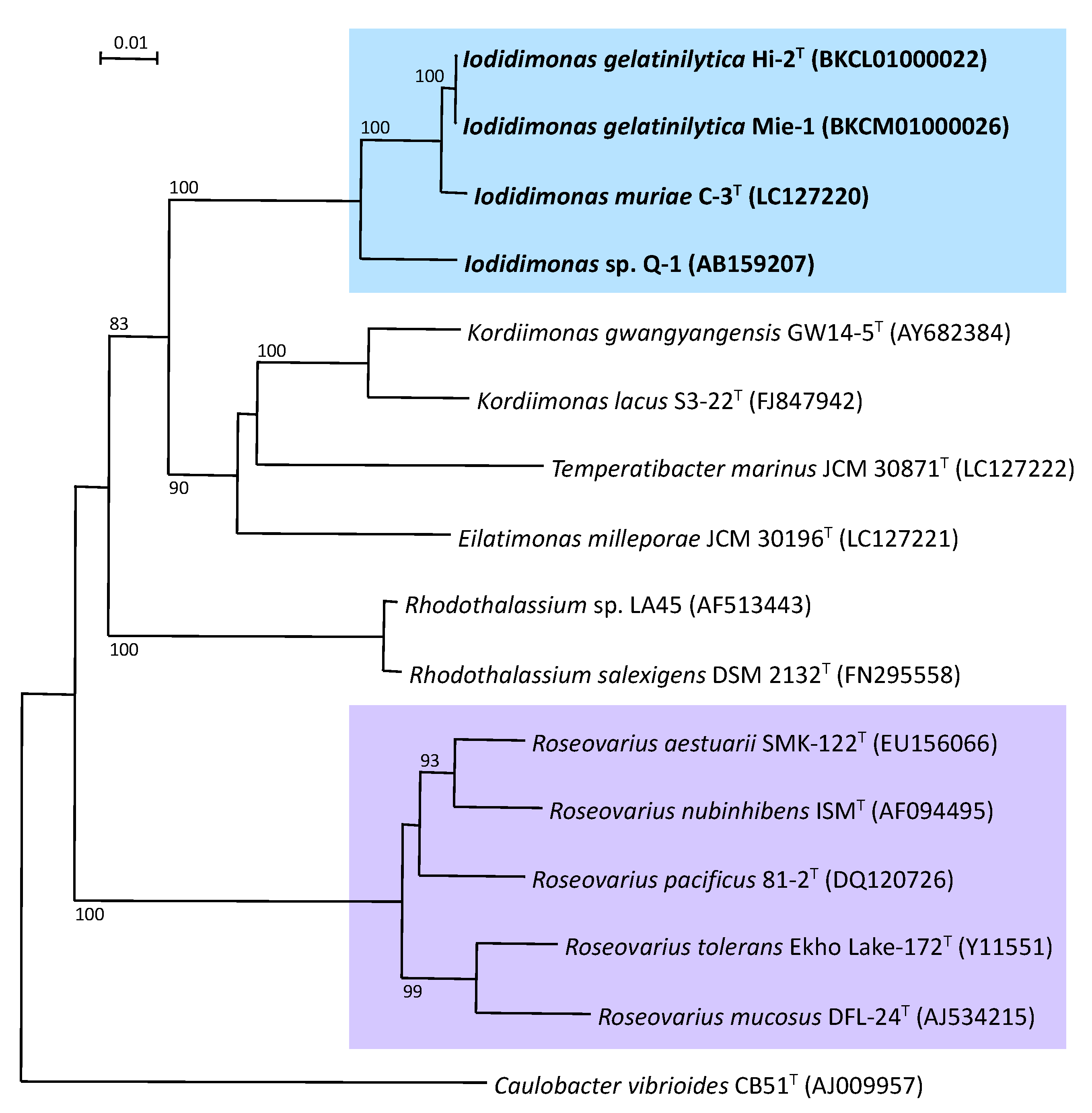
4. Molecular Phylogeny and Systematics of Iodidimonas
5. Biochemistry of Iodide Oxidation by Iodidimonas
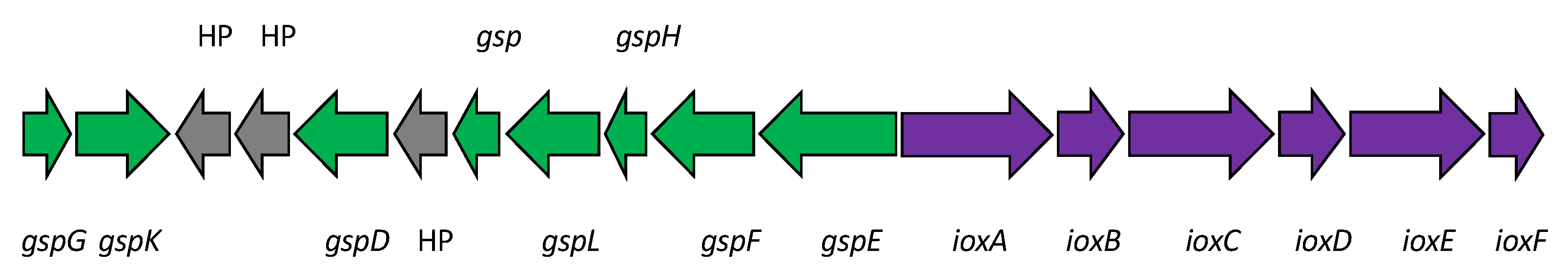
6. Genome Analysis of Iodidimonas
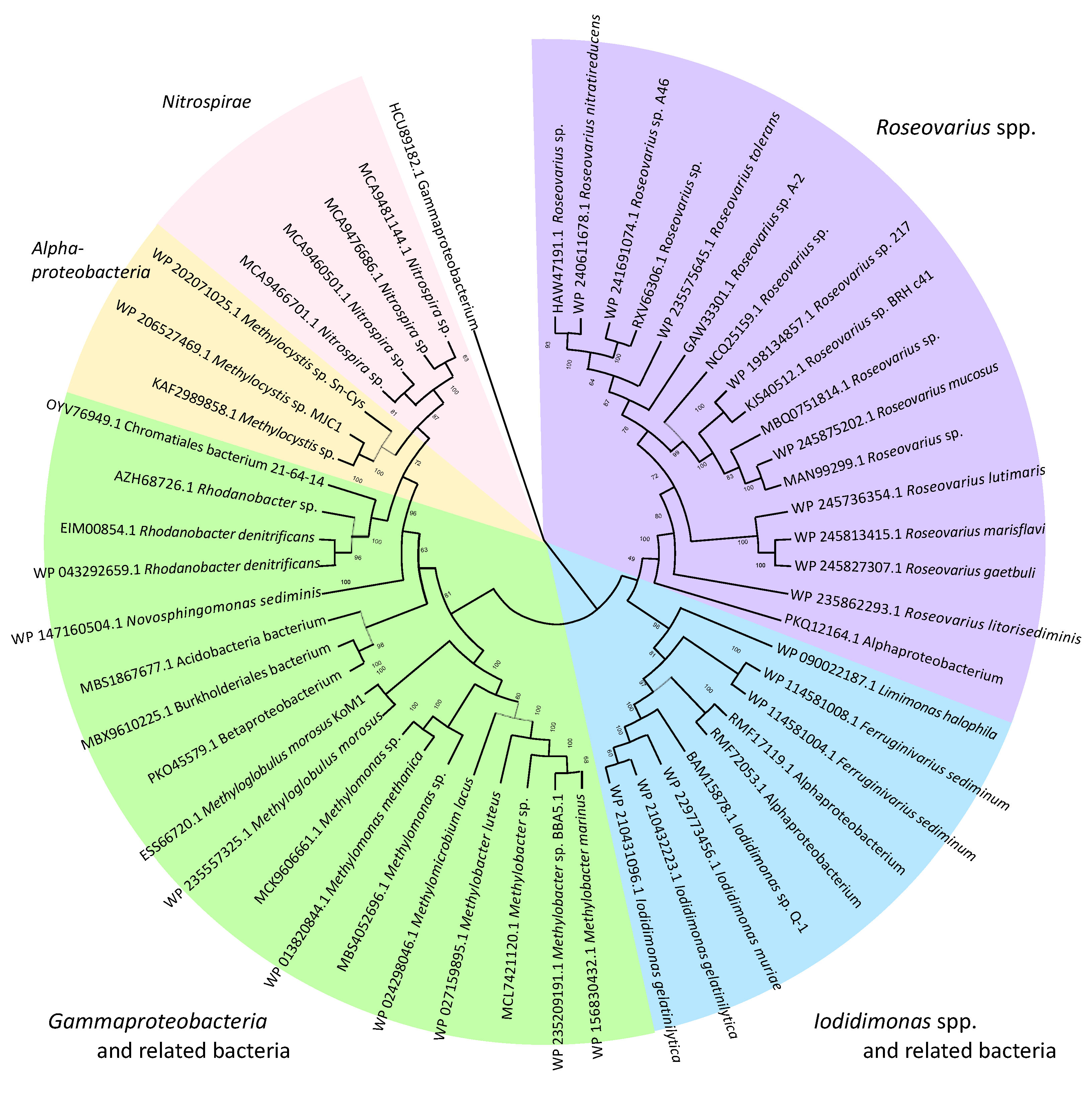
7. Habitat and Distribution of Iodidimonas in Nature
8. Potential Application of Iodidimonas
9. Future Aspects
Funding
Conflicts of Interest
References
- Hetzel, B.S. Iodine deficiency disorders (IDD) and their eradication. Lancet 1983, 2, 1126–1129. [Google Scholar] [CrossRef]
- Hetzel, B.S.; Mano, M.T. A review of experimental studies of iodine deficiency during fetal development. J. Nutr. 1989, 119, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Buraglio, N.; Aldahan, A.; Possnert, G.; Vintersved, I. 129I from the nuclear reprocessing facilities traced in precipitation and runoff in northern Europe. Environ. Sci. Technol. 2001, 35, 1579–1586. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.L.; Dahlgaard, H.; Nielsen, S.P. Iodine-129 time series in Danish, Norwegian and northwest Greenland coast and the Baltic Sea by seaweed. Estuar. Coast. Shelf Sci. 2000, 51, 571–584. [Google Scholar] [CrossRef]
- Moran, J.E.; Oktay, S.; Santschi, P.H.; Schink, D.R. Atmospheric dispersal of 129Iodine from nuclear fuel reprocessing facilities. Environ. Sci. Technol. 1999, 33, 2536–2542. [Google Scholar] [CrossRef]
- Raisbeck, G.M.; Yiou, F. 129I in the oceans: Origin and applications. Sci. Total Environ. 1999, 238, 31–41. [Google Scholar] [CrossRef]
- Cohen, B.L. The origin of I in soil and the 129I problem. Health Phys. 1985, 49, 279–285. [Google Scholar] [CrossRef]
- Fuge, R.; Johnson, C.C. Iodine and human health, the role of environmental geochemistry and diet, a review. Appl. Geochem. 2015, 63, 282–302. [Google Scholar] [CrossRef]
- Wong, G.T.F. The marine geochemistry of iodine. Rev. Aquat. Sci. 1991, 4, 45–73. [Google Scholar]
- Sillen, L.G. The physical chemistry of seawater. In Oceanography; Sears, M., Ed.; American Association for the Advancement of Science: Washington, DC, USA, 1961; pp. 549–581. [Google Scholar]
- Campos, M.L.A.M.; Farrenkopf, A.M.; Jickells, T.D.; Luther, G.W., III. A comparison of dissolved iodine cycling at the Bermuda Atlantic Time-series Station and Hawaii Ocean Time-series Station. Deep-Sea Res. Part II 1996, 43, 455–466. [Google Scholar] [CrossRef]
- Tian, R.C.; Marty, J.C.; Nicolas, E.; Chiavérini, J.; Ruiz-Pino, D.; Pizay, M.D. Iodine speciation: A potential indicator to evaluate new production versus regenerated production. Deep-Sea Res. Part I 1996, 43, 723–738. [Google Scholar] [CrossRef]
- Tian, R.C.; Nicolas, E. Iodine speciation in the northwest Mediterranean Sea: Method and vertical profile. Mar. Chem. 1995, 48, 151–156. [Google Scholar] [CrossRef]
- Councell, T.B.; Landa, E.R.; Lovley, D.R. Microbial reduction of iodate. Water Air Soil Pollut. 1997, 100, 99–106. [Google Scholar] [CrossRef]
- Farrenkopf, A.M.; Dollhopf, M.E.; Chadhain, S.N.; Luther III, G.W.; Nealson, K.H. Reduction of iodate in seawater during Arabian Sea shipboard incubations and in laboratory cultures of the marine bacterium Shewanella putrefaciens strain MR-4. Mar. Chem. 1997, 57, 347–354. [Google Scholar] [CrossRef]
- Yamazaki, C.; Kashiwa, S.; Horiuchi, A.; Kasahara, Y.; Yamamura, S.; Amachi, S. A novel dimethylsulfoxide reductase family of molybdoenzyme, Idr, is involved in iodate respiration by Pseudomonas sp. SCT. Environ. Microbiol. 2020, 22, 2196–2212. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.T.F.; Piumsomboon, A.U.; Dunstan, W.M. The transformation of iodate to iodide in marine phytoplankton cultures. Mar. Ecol. Prog. Ser. 2002, 237, 27–39. [Google Scholar] [CrossRef]
- Gozlan, R.S. Isolation of iodine-producing bacteria from aquaria. Antonie Van Leeuwenhoek 1968, 34, 226. [Google Scholar] [CrossRef]
- Gozlan, R.S.; Margalith, P. Iodide oxidation by a marine bacterium. J. Appl. Bacteriol. 1973, 36, 407–417. [Google Scholar] [CrossRef]
- Gozlan, R.S.; Margalith, P. Iodide oxidation by Pseudomonas iodooxidans. J. Appl. Bacteriol. 1974, 37, 493–499. [Google Scholar] [CrossRef]
- Amachi, S.; Muramatsu, Y.; Akiyama, Y.; Miyazaki, K.; Yoshiki, S.; Hanada, S.; Kamagata, Y.; Ban-nai, T.; Shinoyama, H.; Fujii, T. Isolation of iodide-oxidizing bacteria from iodide-rich natural gas brines and seawaters. Microb. Ecol. 2005, 49, 547–557. [Google Scholar] [CrossRef]
- Muramatsu, Y.; Fehn, U.; Yoshida, S. Recycling of iodine in fore-arc areas: Evidence from the iodine brines in Chiba, Japan. Earth Planet. Sci. Lett. 2001, 192, 583–593. [Google Scholar] [CrossRef]
- Iino, T.; Ohkuma, M.; Kamagata, Y.; Amachi, S. Iodidimonas muriae gen. nov., sp. nov., an aerobic iodide-oxidizing bacterium isolated from brine of a natural gas and iodine recovery facility, and proposals of Iodidimonadaceae fam. nov., Iodidimonadales ord. nov., Emcibacteraceae fam. nov. and Emcibacterales ord. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5016–5022. [Google Scholar] [PubMed]
- Arakawa, Y.; Akiyama, Y.; Furukawa, H.; Suda, W.; Amachi, S. Growth stimulation of iodide-oxidizing α-Proteobacteria in iodide-rich environments. Microb. Ecol. 2012, 63, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Wakai, S.; Ito, K.; Iino, T.; Tomoe, Y.; Mori, K.; Harayama, S. Corrosion of iron by iodide-oxidizing bacteria isolated from brine in an iodine production facility. Microb. Ecol. 2014, 68, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Sugai, Y.; Sasaki, K.; Wakizono, R.; Higuchi, Y.; Muraoka, N. Considerations on the possibility of microbial clogging of re-injection wells of the wastewater generated in a water-dissolved natural gas field. Int. Biodeterior. Biodegrad. 2013, 81, 35–43. [Google Scholar] [CrossRef]
- Iino, T.; Oshima, K.; Hattori, M.; Ohkuma, M.; Amachi, S. Iodidimonas gelatinilytica sp. nov., aerobic iodide-oxidizing bacteria isolated from brine water and surface seawater. Antonie Van Leeuwenhoek 2021, 114, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Yarza, P.; Yilmaz, P.; Pruesse, E.; Glöckner, F.O.; Ludwig, W.; Schleifer, K.-H.; Whitman, W.B.; Euzéby, J.; Amann, R.; Rosselló-Móra, R. Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nat. Rev. Microbiol. 2014, 12, 635–645. [Google Scholar] [CrossRef]
- Meier-Kothoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar]
- Suzuki, M.; Eda, Y.; Ohsawa, S.; Kanesaki, Y.; Yoshikawa, H.; Tanaka, K.; Muramatsu, Y.; Yoshikawa, J.; Sato, I.; Fujii, T.; et al. Iodide oxidation by a novel multicopper oxidase from the Alphaproteobacterium strain Q-1. Appl. Environ. Microbiol. 2012, 78, 3941–3949. [Google Scholar] [CrossRef]
- Ehara, A.; Suzuki, H.; Kanesaki, Y.; Yoshikawa, H.; Amachi, S. Draft genome sequence of strain Q-1, an iodide-oxidizing Alphaproteobacterium isolated from natural gas brine water. Genome Announc. 2014, 2, e00659-14. [Google Scholar] [CrossRef]
- Schulze, M.; Roedel, G. Accumulation of the cytochrome c oxidase subunits I and II in yeast requires a mitochondrial membrane-associated protein, encoded by the nuclear SCO1 gene. Mol. Gen. Genet. 1989, 216, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Shiroyama, K.; Kawasaki, Y.; Unno, Y.; Amachi, S. A putative multicopper oxidase, IoxA, is involved in iodide oxidation by Roseovarius sp. strain A-2. Biosci. Biotechnol. Biochem. 2015, 79, 1898–1905. [Google Scholar] [CrossRef] [PubMed]
- Solomon, E.I.; Sundaram, U.M.; Machonkin, T.E. Multicopper oxidases and oxygenases. Chem. Rev. 1996, 96, 2563–2605. [Google Scholar] [CrossRef] [PubMed]
- Mohan, A.M.; Hartsock, A.; Hammack, R.W.; Vidic, R.D. Microbial communities in flowback water impoundments from hydraulic fracturing for recovery of shale gas. FEMS Microbiol. Ecol. 2013, 86, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Almaraz, N.; Regnery, J.; Vanzin, G.F.; Riley, S.M.; Ahoor, D.C.; Cath, T.Y. Emergence and fate of volatile iodinated organic compounds during biological treatment of oil and gas produced water. Sci. Total Environ. 2020, 699, 134202. [Google Scholar] [CrossRef]
- Van Houghton, B.D.; Acharya, S.M.; Rosenblum, J.S.; Chakraborty, R.; Tringe, S.G.; Cath, T.Y. Membrane bioreactor pretreatment of high-salinity O&G produced water. ACS EST Water 2022, 2, 484–494. [Google Scholar]
- Wen, P.; Huang, Y.; Qiu, Z.; Li, Q. Microbial response during treatment of different types of landfill leachate in a semi-aerobic aged refuse biofilter. Chemosphere 2021, 262, 127822. [Google Scholar] [CrossRef]
- Toshakov, S.V.; Izotova, A.O.; Vinogradova, E.N.; Kachmazov, G.S.; Tuaeva, A.Y.; Abaev, V.T.; Evteeva, M.A.; Gunitseeva, N.M.; Korzhenkov, A.A.; Elcheninov, A.G.; et al. Culture-independent survey of thermophilic microbial communities of the North Caucasus. Biology 2021, 10, 1352. [Google Scholar] [CrossRef]
- Cooper, R.A. Iodine revisited. Int. Wound J. 2007, 4, 124–137. [Google Scholar] [CrossRef]
- Gottardi, W. Chapter 8: Iodine and iodine compounds. In Disinfection, Sterilization and Preservation, 3rd ed.; Block, S.S., Ed.; Lea & Febiger: Philadelphia, PA, USA, 1983; pp. 183–196. [Google Scholar]
- Hickey, J.; Panicucci, R.; Duan, Y.; Dinehart, K.; Murphy, J.; Kessler, J.; Gottardi, W. Control of the amount of free molecular iodine in iodine germicides. J. Pharm. Pharmacol. 1997, 49, 1195–1199. [Google Scholar] [CrossRef]
- Hansen, E.H.; Albertsen, L.; Schäfer, T.; Johansen, C.; Frisvad, J.C.; Molin, S.; Gram, L. Curvularia haloperoxidase: Antimicrobial activity and potential application as a surface disinfectant. Appl. Environ. Microbiol. 2003, 69, 4611–4617. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yuliana, T.; Ebihara, K.; Suzuki, M.; Shimonaka, C.; Amachi, S. A novel enzyme-based antimicrobial system comprising iodide and a multicopper oxidase isolated from Alphaproteobacterium strain Q-1. Appl. Microbiol. Biotechnol. 2015, 99, 10011–10018. [Google Scholar] [CrossRef] [PubMed]
- Morozova, O.V.; Shumakovich, G.P.; Shleev, S.V.; Yaropolov, Y.I. Laccase-mediator systems and their applications: A review. Appl. Biochem. Microbiol. 2007, 43, 523–535. [Google Scholar] [CrossRef]
- Riva, S. Laccases: Blue enzymes for green chemistry. Trends Biotechnol. 2006, 24, 219–226. [Google Scholar] [CrossRef]
- Taguchi, T.; Ebihara, K.; Yanagisaki, C.; Yoshikawa, J.; Horiguchi, H.; Amachi, S. Decolorization of recalcitrant dyes by a multicopper oxidase produced by Iodidimonas sp. Q-1 with iodide as a novel inorganic natural redox mediator. Sci. Rep. 2018, 8, 6717. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, K.; Yoshikawa, J.; Horiguchi, H.; Amachi, S. Decolorization of cationic dyes under alkaline conditions by Iodidimonas sp. Q-1 multicopper oxidase. J. Biosci. Bioeng. 2022, 133, 323–328. [Google Scholar] [CrossRef]
- Kaksonen, A.H.; Mudunuru, B.M.; Hackl, R. The role of microorganisms in gold processing and recovery—A review. Hydrometallurgy 2014, 142, 70–83. [Google Scholar] [CrossRef]
- Khaing, S.Y.; Sugai, Y.; Sasaki, K. Gold dissolution from ore with iodide-oxidising bacteria. Sci. Rep. 2019, 9, 4178. [Google Scholar] [CrossRef]
- Kudpeng, K.; Bohu, T.; Morris, C.; Thiravetyan, P.; Kaksonen, A.H. Bioleaching of gold from sulfidic gold ore concentrate and electronic waste by Roseovarius tolerans and Roseovarius mucosus. Microorganisms 2020, 8, 1783. [Google Scholar] [CrossRef]
- Esposti, M.D.; Geiger, O.; Sanchez-Flores, A.; Esposti, M.D. On the bacterial ancestry of mitochondria: New insights with triangulated approaches. bioRxiv 2022. [Google Scholar] [CrossRef]
- Muramatsu, Y.; Wedepohl, K.H. The distribution of iodine in the earth’s crust. Chem. Geol. 1998, 147, 201–216. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | 1 | 2 |
|---|---|---|
| Hydrolysis of aesculin | + | − |
| Hydrolysis of gelatin | − | + |
| API 20E | ||
| L-Arabinose | − | + |
| D-Maltose | + | w/− |
| Cellular fatty acid composition (%) | ||
| C16:1ω5c | 17.2 | 11.5–13.4 |
| C17:1ω6c | 8.4 | 4.0–4.2 |
| 11-methyl C18:1ω7c | 3.0 | 4.2–7.5 |
| C18:1 2-OH | 8.4 | 10.8–14.1 |
| Summed feature 8 a | 38.0 | 42.0–49.1 |
| Strain | Assembly | Size (Mb) | GC (%) | CDS | Contigs | |
|---|---|---|---|---|---|---|
| Iodidimonas muriae | C-3T | GCA_014647255.1 | 3.01 | 55.6 | 2718 | 26 |
| Iodidimonas gelatinilytica | Hi-2T | GCA_008579125.1 | 2.85 | 55.4 | 2709 | 30 |
| Iodidimonas gelatinilytica | Mie-1 | GCA_008579145.1 | 2.80 | 55.3 | 2644 | 47 |
| Iodidimonas sp. | Q-1 | GCA_000710935.1 | 3.09 | 56.1 | 2788 | 109 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amachi, S.; Iino, T. The Genus Iodidimonas: From Its Discovery to Potential Applications. Microorganisms 2022, 10, 1661. https://doi.org/10.3390/microorganisms10081661
Amachi S, Iino T. The Genus Iodidimonas: From Its Discovery to Potential Applications. Microorganisms. 2022; 10(8):1661. https://doi.org/10.3390/microorganisms10081661
Chicago/Turabian StyleAmachi, Seigo, and Takao Iino. 2022. "The Genus Iodidimonas: From Its Discovery to Potential Applications" Microorganisms 10, no. 8: 1661. https://doi.org/10.3390/microorganisms10081661
APA StyleAmachi, S., & Iino, T. (2022). The Genus Iodidimonas: From Its Discovery to Potential Applications. Microorganisms, 10(8), 1661. https://doi.org/10.3390/microorganisms10081661