
Functionalization of Multi-Walled Carbon Nanotubes Changes Their Antibiofilm and Probiofilm Effects on Environmental Bacteria

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Medium
2.2. Multi-Walled Carbon Nanotubes
2.3. Determination of Biofilm Biomass
2.4. Biofilm Eradication in the Presence of pMWCNTs and Functionalized MWCNTs
2.5. Assessment of Respiratory Activity of Biofilms
2.6. Determination of Metabolic Activity of Biofilm Cells
2.7. Statistical Analysis
3. Results
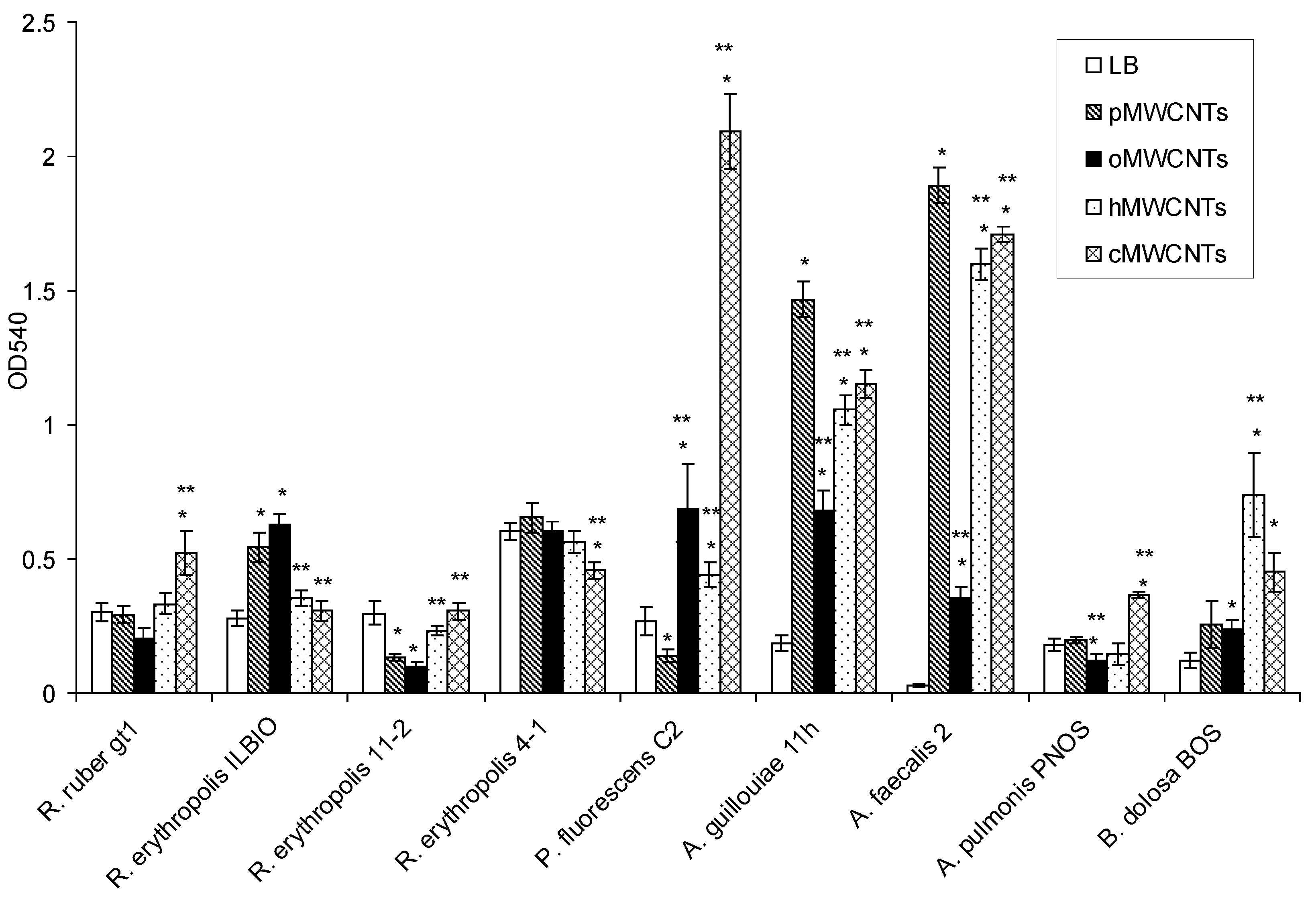
3.1. Effects of pMWCNTs and Functionalized MWCNTs on Bacterial Biofilm Formation
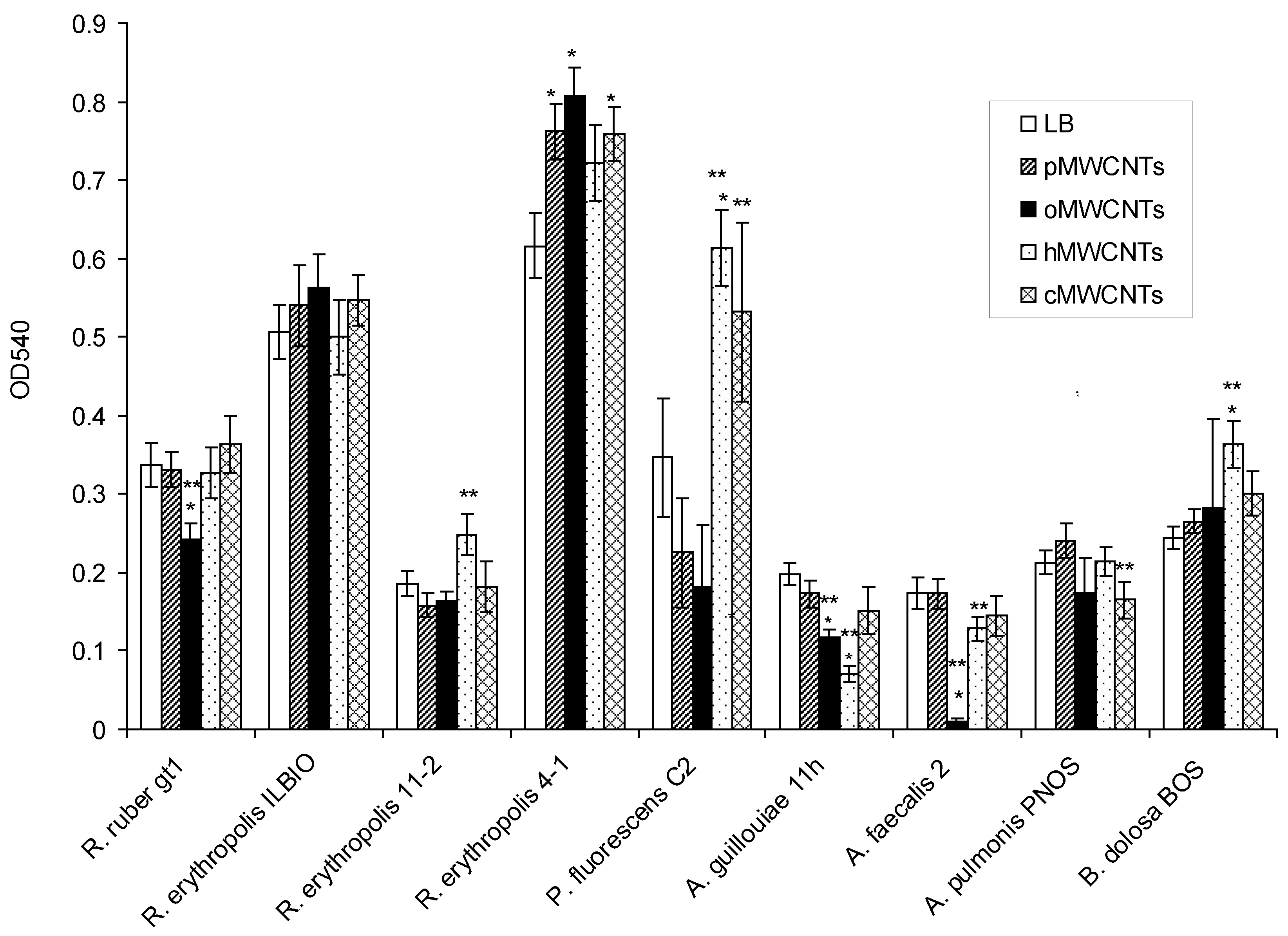
3.2. Effects of pMWCNTs and Functionalized MWCNTs on Biofilm Eradication
3.3. Effect of pMWCNTs and Functionalized MWCNTs on the Respiratory Activity of Biofilms
3.4. The Effects of pMWCNTs and Functionalized MWCNTs on the Metabolic Activity of Biofilms, Estimated by the Reduction of Resazurin to Rezarufan
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Aqel, A.; El-Nour, K.M.M.A.; Ammar, R.A.A.; Al-Warthan, A. Carbon nanotubes, science and technology part (I) structure, synthesis and characterisation. Arab. J. Chem. 2012, 5, 1–23. [Google Scholar] [CrossRef]
- Dong, Z.-J.; Sun, B.; Zhu, H.; Yuan, G.-M.; Li, B.-L.; Guo, J.-G.; Li, X.-K.; Cong, Y.; Zhang, J. A review of aligned carbon nanotube arrays and carbon/carbon composites: Fabrication, thermal conduction properties and applications in thermal management. New Carbon Mater. 2021, 36, 873–892. [Google Scholar] [CrossRef]
- Kang, S.; Pinault, M.; Pfefferle, L.D.; Elimelech, M. Single-walled carbon nanotubes exhibit strong antimicrobial activity. Langmuir 2007, 23, 8670–8673. [Google Scholar] [CrossRef]
- Kang, S.; Herzberg, M.; Rodrigues, D.F.; Elimelech, M. Antibacterial effects of carbon nanotubes: Size does matter! Langmuir 2008, 24, 6409–6413. [Google Scholar] [CrossRef]
- Ghasemi, M.; Daud, W.R.W.; Hassan, S.H.A.; Ohc, S.-E.; Ismail, M.; Rahimnejad, M.; Jahim, J.M. Nano-structured carbon as electrode material in microbial fuel cells: A comprehensive review. J. Alloys Compd. 2013, 580, 245–255. [Google Scholar] [CrossRef]
- Azizi-Lalabadi, M.; Hashemi, H.; Feng, J.; Jafari, S.M. Carbon nanomaterials against pathogens; the antimicrobial activity of carbon nanotubes, graphene/graphene oxide, fullerenes, and their nanocomposites. Adv. Colloid Interface Sci. 2020, 284, 102250. [Google Scholar] [CrossRef]
- Jackson, P.; Jacobsen, N.R.; Baun, A.; Birkedal, R.; Kühnel, D.; Jensen, K.A.; Vogel, U.; Wallin, H. Bioaccumulation and ecotoxicity of carbon nanotubes. Chem. Cent. J. 2013, 7, 154. [Google Scholar] [CrossRef]
- Seo, Y.; Hwang, J.; Lee, E.; Kim, Y.J.; Lee, K.; Park, C.; Choi, Y.; Jeonae, H.; Choi, J. Engineering copper nanoparticles synthesized on the surface of carbon nanotubes for anti-microbial and anti-biofilm applications. Nanoscale 2018, 10, 15529–15544. [Google Scholar] [CrossRef]
- Zardini, H.Z.; Amiri, A.; Shanbedi, M.; Maghrebi, M.; Baniadam, M. Enhanced antibacterial activity of amino acids-functionalized multi walled carbon nanotubes by a simple method. Colloids Surf. B Biointerfaces 2012, 92, 196–202. [Google Scholar] [CrossRef]
- Chi, M.-F.; Wu, W.-L.; Du, Y.; Chin, C.-J.M.; Lin, C.-C. Inactivation of Escherichia coli planktonic cells by multi-walled carbon nanotubes in suspensions: Effect of surface functionalization coupled with medium nutrition level. J. Hazard. Mater. 2016, 318, 507–514. [Google Scholar] [CrossRef]
- Gao, F.; Ma, B.; Zhao, Z.S.; Guo, L.; Jin, C.; Ga, M. Performance evaluation, enzymatic activity and microbial community of sequencing batch reactor under hydroxyl-functionalized multi-walled carbon nanotubes (MWCNTs-OH) stress. Environ. Technol. Innov. 2021, 21, 101213. [Google Scholar] [CrossRef]
- Abbasian, F.; Lockington, R.; Palanisami, T.; Megharaj, M.; Naidu, R. Multiwall carbon nanotubes increase the microbial community in crude oil contaminated fresh water sediments. Sci. Total Environ. 2016, 539, 370–380. [Google Scholar] [CrossRef]
- Song, B.; Gong, J.; Tang, W.; Zeng, G.; Chen, M.; Xu, P.; Shen, M.; Ye, S.; Feng, H.; Zhou, C.; et al. Influence of multi-walled carbon nanotubes on the microbial biomass, enzyme activity, and bacterial community structure in 2,4-dichlorophenol-contaminated sediment. Sci. Total Environ. 2020, 713, 136645. [Google Scholar] [CrossRef]
- Garrett, T.R.; Bhakoo, M.; Zhang, Z. Bacterial adhesion and biofilms on surfaces. Prog. Nat. Sci. 2008, 18, 1049–1056. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Rodrigues, D.F.; Elimelech, M. Toxic effects of single-walled carbon nanotubes in the development of E. coli biofilm. Environ. Sci. Technol. 2010, 44, 4583–4589. [Google Scholar] [CrossRef]
- Goyal, D.; Zhang, X.J.; Rooney-Varga, J.N. Impacts of single-walled carbon nanotubes on microbial community structure in activated sludge. Lett. Appl. Microbiol. 2010, 51, 428–435. [Google Scholar] [CrossRef]
- Mahto, K.U.; Das, S. Bacterial biofilm and extracellular polymeric substances in the moving bed biofilm reactor for wastewater treatment: A review. Bioresour. Technol. 2022, 345, 126476. [Google Scholar] [CrossRef]
- Li, L.; He, Z.; Liang, T.; Sheng, T.; Zhang, F.; Wu, D.; Ma, F. Colonization of biofilm in wastewater treatment: A review. Environ. Pollut. 2022, 293, 118514. [Google Scholar] [CrossRef]
- Zhang, K.; Li, J.; Zheng, Z.; Zhang, J.; Sun, M.; Huang, S. Analyzing the sludge characteristics and microbial communities of biofilm and activated sludge in the partial nitrification/anammox process. J. Water Process Eng. 2022, 46, 102618. [Google Scholar] [CrossRef]
- Mukherjee, M.; Zaiden, N.; Teng, A.; Hu, Y.; Cao, B. Shewanella biofilm development and engineering for environmental and bioenergy applications. Curr. Opin. Chem. Biol. 2020, 59, 84–92. [Google Scholar] [CrossRef]
- Albarracin-Arias, J.A.; Yu, C.-P.; Maeda, T.; Quintero, W.V.; Sanchez-Torres, V. Microbial community dynamics and electricity generation in MFCs inoculated with POME sludges and pure electrogenic culture. Int. J. Hydrog. Energy 2021, 46, 36903–36916. [Google Scholar] [CrossRef]
- Cai, T.; Zhang, Y.; Wang, N.; Zhang, Z.; Lu, X.; Zhen, G. Electrochemically active microorganisms sense charge transfer resistance for regulating biofilm electroactivity, spatio-temporal distribution, and catabolic pathway. Chem. Eng. J. 2022, 442, 136248. [Google Scholar] [CrossRef]
- Rosche, B.; Li, X.Z.; Hauer, B.; Schmid, A.; Buehler, K. Microbial biofilms: A concept for industrial catalysis? Trends Biotechnol. 2009, 27, 636–643. [Google Scholar] [CrossRef]
- Halan, B.; Buehler, K.; Schmid, A. Biofilms as living catalysts in continuous chemical syntheses. Trends Biotechnol. 2012, 30, 453–465. [Google Scholar] [CrossRef]
- Berlanga, M.; Guerrero, R. Living together in biofilms: The microbial cell factory and its biotechnological implications. Microb. Cell Factories 2016, 15, 165. [Google Scholar] [CrossRef]
- Sonawane, J.M.; Rai, A.K.; Sharma, M.; Tripathi, M.; Prasad, R. Microbial biofilms: Recent advances and progress in environmental bioremediation. Sci. Total Environ. 2022, 824, 153843. [Google Scholar] [CrossRef]
- Shukla, S.K.; Mangwani, N.; Rao, T.S.; Das, S. Biofilm-mediated bioremediation of polycyclic aromatic hydrocarbons. In Microbial Biodegradation and Bioremediation, 1st ed.; Das, S., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2014; pp. 203–232. [Google Scholar] [CrossRef]
- Catania, V.; Lopresti, F.; Cappello, S.; Scaffaro, R.; Quatrini, P. Innovative, ecofriendly biosorbent-biodegrading biofilms for bioremediation of oil- contaminated water. New Biotechnol. 2020, 58, 25–31. [Google Scholar] [CrossRef]
- Bouteh, E.; Ahmadi, N.; Abbasi, M.; Torabian, A.; van Loosdrecht, M.C.M.; Ducoste, J. Biodegradation of organophosphorus pesticides in moving bed biofilm reactors: Analysis of microbial community and biodegradation pathways. J. Hazard. Mater. 2021, 408, 124950. [Google Scholar] [CrossRef]
- Lee, S.W.; Phillips, K.S.; Gu, H.; Kazemzadeh-Narbat, M.; Ren, D. How microbes read the map: Effects of implant topography on bacterial adhesion and biofilm formation. Biomaterials 2021, 268, 120595. [Google Scholar] [CrossRef]
- Alizadeh, A.; Razmjou, A.; Ghaedi, M.; Jannesar, R. Nanoporous solid-state membranes modified with multi-wall carbon nanotubes with anti-biofouling property. Int. J. Nanomed. 2019, 14, 1669–1685. [Google Scholar] [CrossRef]
- Zhou, J.; Qi, X. Multi-walled carbon nanotubes⁄epilson-polylysine nanocomposite with enhanced antibacterial activity. Lett. Appl. Microbiol. 2010, 52, 76–83. [Google Scholar] [CrossRef]
- Benigno, E.; Lorente, M.A.; Olmosm, D.; González-Gaitano, G.; González-Benito, J. Nanocomposites based on low density polyethylene filled with carbon nanotubes prepared by high energy ball milling and their potential antibacterial activity. Polym. Int. 2019, 68, 1155–1163. [Google Scholar] [CrossRef]
- Ikuma, K.; Decho, A.W.; Lau, B.L.T. When nanoparticles meet biofilms—Interactions guiding the environmental fate and accumulation of nanoparticles. Front. Microbiol. 2015, 6, 291. [Google Scholar] [CrossRef]
- Malek, I.; Schaber, C.F.; Heinlein, T.; Schneider, J.J.; Gorb, S.N.; Schmitz, R.A. Vertically aligned multi walled carbon nanotubes prevent biofilm formation of medically relevant bacteria. J. Mater. Chem. B 2016, 4, 5228–5235. [Google Scholar] [CrossRef]
- Sun, Y.; Lang, Y.; Sun, T.; Liu, Q.; Pan, Y.; Qi, Z.; Ling, N.; Feng, Y.; Yu, M.; Ji, Y.; et al. Antifouling potential of multi-walled carbon nanotubes-modified chlorinated rubber-based composites on the colonization dynamics of pioneer biofilm-forming eukaryotic microbes. Int. Biodeterior. Biodegrad. 2020, 149, 104921. [Google Scholar] [CrossRef]
- Alvarez, A.; Saez, J.M.; Costa, J.S.D.; Colin, V.L.; Fuentes, M.S.; Cuozzo, S.A.; Benimeli, C.S.; Polti, M.A.; Amoroso, M.J. Actinobacteria: Current research and perspectives for bioremediation of pesticides and heavy metals. Chemosphere 2017, 166, 41–62. [Google Scholar] [CrossRef]
- Aparicio, J.D.; Raimondo, E.E.; Gil, R.A.; Benimeli, C.S.; Polti, M.A. Actinobacteria consortium as an efficient biotechnological tool for mixed polluted soil reclamation: Experimental factorial design for bioremediation process optimization. J. Hazard. Mater. 2018, 342, 408–417. [Google Scholar] [CrossRef]
- Maksimova, Y.G.; Sergeeva, A.A.; Ovechkina, G.V.; Maksimov, A.Y. Pyridine degradation by suspensions and biofilms of Achromobacter pulmonis PNOS and Burkholderia dolosa BOS strains isolated from activated sludge of sewage treatment plants. Biotechnology 2020, 36, 86–98. [Google Scholar] [CrossRef]
- Demakov, V.A.; Vasil’ev, D.M.; Maksimova, Y.G.; Pavlova, Y.A.; Ovechkina, G.V.; Maksimov, A.Y. Activated sludge bacteria transforming cyanopyridines and amides of pyridinecarboxylic acids. Microbiology 2015, 84, 433–441. [Google Scholar] [CrossRef]
- Maksimova, Y.G.; Maksimov, A.Y.; Demakov, V.A.; Kozlov, K.V.; Ovechkina, G.V.; Olontsev, V.F. Hydrolysis of acrylonitrile by nitrile-converting bacterial cells immobilized on fibrous carbon adsorbents. Appl. Biochem. Microbiol. 2011, 47, 681–687. [Google Scholar] [CrossRef]
- Maksimov, A.Y.; Kuznetsova, M.V.; Ovechkina, G.V.; Kozlov, S.V.; Maksimova, Y.G.; Demakov, V.A. Effects of nitriles and amides on the growth and nitrile hydratase activity of the Rhodococcus sp. strain gt1. Appl. Biochem. Microbiol. 2003, 39, 55–59. [Google Scholar]
- Pérez, L.M.; Alvarez, B.L.; Codony, F.; Fittipaldi, M.; Adrados, B.; Peñuela, G.; Morató, J. A new microtitre plate screening method for evaluating the viability of aerobic respiring bacteria in high surface biofilms. Lett. Appl. Microbiol. 2010, 51, 331–337. [Google Scholar] [CrossRef]
- Plakunov, V.K.; Mart’yanov, S.V.; Teteneva, N.A.; Zhurina, M.V. Controlling of microbial biofilms formation: Anti- and probiofilm agents. Microbiology 2017, 86, 423–438. [Google Scholar] [CrossRef]
- McNicholla, B.P.; McGrathb, J.W.; Quinn, J.P. Development and application of a resazurin-based biomass activity test for activated sludge plant management. Water Res. 2007, 41, 127–133. [Google Scholar] [CrossRef]
- Amiri, A.; Zare-Zardini, H.; Shanbedi, M.; Kazi, S.N.; Taheri-Kafrani, A.; Chew, B.T.; Zarrabi, A. Microbial toxicity of different functional groups-treated carbon nanotubes. In Surface Chemistry of Nanobiomaterials, 1st ed.; Grumezescu, A., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 33–70. [Google Scholar] [CrossRef]
- Arias, L.R.; Yang, L. Inactivation of bacterial pathogens by carbon nanotubes in suspensions. Langmuir 2009, 25, 3003–3012. [Google Scholar] [CrossRef]
- Gholami, S.; Llacuna, J.L.; Vatanpour, V.; Dehqan, A.; Paziresh, S.; Cortina, J.L. Impact of a new functionalization of multiwalled carbon nanotubes on antifouling and permeability of PVDF nanocomposite membranes for dye wastewater treatment. Chemosphere 2022, 294, 133699. [Google Scholar] [CrossRef]
- Alonso, B.; Cruces, R.; Pérez, A.; Sánchez-Carrillo, C.; Guembe, M. Comparison of the XTT and resazurin assays for quantification of the metabolic activity of Staphylococcus aureus biofilm. J. Microbiol. Methods 2017, 139, 135–137. [Google Scholar] [CrossRef]
- Zhu, C.; Mahmood, Z.; Zhang, W.; Akram, M.W.; Ainur, D.; Ma, H. In situ investigation of acute exposure of graphene oxide on activated sludge: Biofilm characteristics, microbial activity and cytotoxicity. Ecotoxicol. Environ. Saf. 2020, 199, 110639. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Strains | pMWCNTs | oMWCNTs | hMWCNTs | cMWCNTs | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| a | b | c | a | b | c | a | b | c | a | b | c | |
| R. ruber gt1 | 1.70 * | 1.50 * | 0.09 | 0.78 * | 1.51 * | 0.83 | 2.19 * | 1.30 | 0.21 | 1.83 * | 1.97 * | 0.12 |
| R. erythropolis IL BIO | 1.50 * | 1.47 | 0.91 | 2.30 * | 1.08 | 0.66 * | 1.57 * | 1.09 | 0.91 | 2.48 * | 1.55 * | 0.13 * |
| R. erythropolis 11-2 | 1.57 * | 1.21 | 0 | 1.34 * | 1.78 * | 0.70 | 1.43 | 1.01 | 0 | 1.14 | 1.30 | 0 |
| R. erythropolis 4-1 | 2.59 * | 0.67 | 0.22 * | 2.78 * | 1.09 | 0.40 | 1.66 * | 0.63 | 0.39 | 1.99 * | 1.09 | 0 |
| Ac. guillouiae 11h | 2.05 * | 0.60 * | 0 | 1.30 * | 0.89 | 0.61 * | 2.27 * | 0.47 * | 0 | 1.98 * | 0.79 | 0 |
| Al. faecalis 2 | 2.08 | 0.45 | 0 | 1.20 | 0.96 | 0.29 | 1.20 | 0.55 | 0 | 1.13 | 1.35 | 0 |
| P. fluorescens C2 | 1.19 * | 1.27 | 0.27 * | 0.94 | 1.25 * | 0.37 * | 1.25 * | 0.86 | 0 | 1.15 * | 2.05 * | 0 |
| A. pulmonis PNOS | 1.24 * | 0.58 * | 0 | 1.29 * | 1.49 | 0.83 | 0.87 * | 0.62 * | 0.13 * | 1.12 * | 1.20 | 0 |
| B. dolosa BOS | 1.04 | 1.04 | 0.38 * | 1.42 * | 1.85 * | 0.81 | 0.97 | 3.13 * | 0.88 | 1.03 | 2.15 * | 0 |
| Strains | pMWCNTs | oMWCNTs | hMWCNTs | cMWCNTs |
|---|---|---|---|---|
| R. ruber gt1 | 37.3 * | 75.6 | 58.3 | 29.9 * |
| R. erythropolis IL BIO | 30.3 * | 84.4 | 29.0 * | 58.2 * |
| R. erythropolis 11-2 | 8.9 * | 77.2 | 25.8 * | 60.1 |
| R. erythropolis 4-1 | 45.6 * | 116.5 | 26.8 * | 49.8 * |
| Ac. guillouiae 11 h | 87.3 | 142.1 | 42.5 | 56.6 * |
| Al. faecalis 2 | 90.8 | 123.5 | 61.1 | 72.5 |
| P. fluorescens C2 | 15.7 * | 127.7 | 19.9 * | 19.7 * |
| A. pulmonis PNOS | 23.8 * | 64.6 * | 33.8 * | 33.8 * |
| B. dolosa BOS | 4.8 * | 49.3 * | 44.7 * | 14.5 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maksimova, Y.; Bykova, Y.; Maksimov, A. Functionalization of Multi-Walled Carbon Nanotubes Changes Their Antibiofilm and Probiofilm Effects on Environmental Bacteria. Microorganisms 2022, 10, 1627. https://doi.org/10.3390/microorganisms10081627
Maksimova Y, Bykova Y, Maksimov A. Functionalization of Multi-Walled Carbon Nanotubes Changes Their Antibiofilm and Probiofilm Effects on Environmental Bacteria. Microorganisms. 2022; 10(8):1627. https://doi.org/10.3390/microorganisms10081627
Chicago/Turabian StyleMaksimova, Yuliya, Yana Bykova, and Aleksandr Maksimov. 2022. "Functionalization of Multi-Walled Carbon Nanotubes Changes Their Antibiofilm and Probiofilm Effects on Environmental Bacteria" Microorganisms 10, no. 8: 1627. https://doi.org/10.3390/microorganisms10081627
APA StyleMaksimova, Y., Bykova, Y., & Maksimov, A. (2022). Functionalization of Multi-Walled Carbon Nanotubes Changes Their Antibiofilm and Probiofilm Effects on Environmental Bacteria. Microorganisms, 10(8), 1627. https://doi.org/10.3390/microorganisms10081627