
Yellow Fever: Roles of Animal Models and Arthropod Vector Studies in Understanding Epidemic Emergence

 ,
,
Abstract
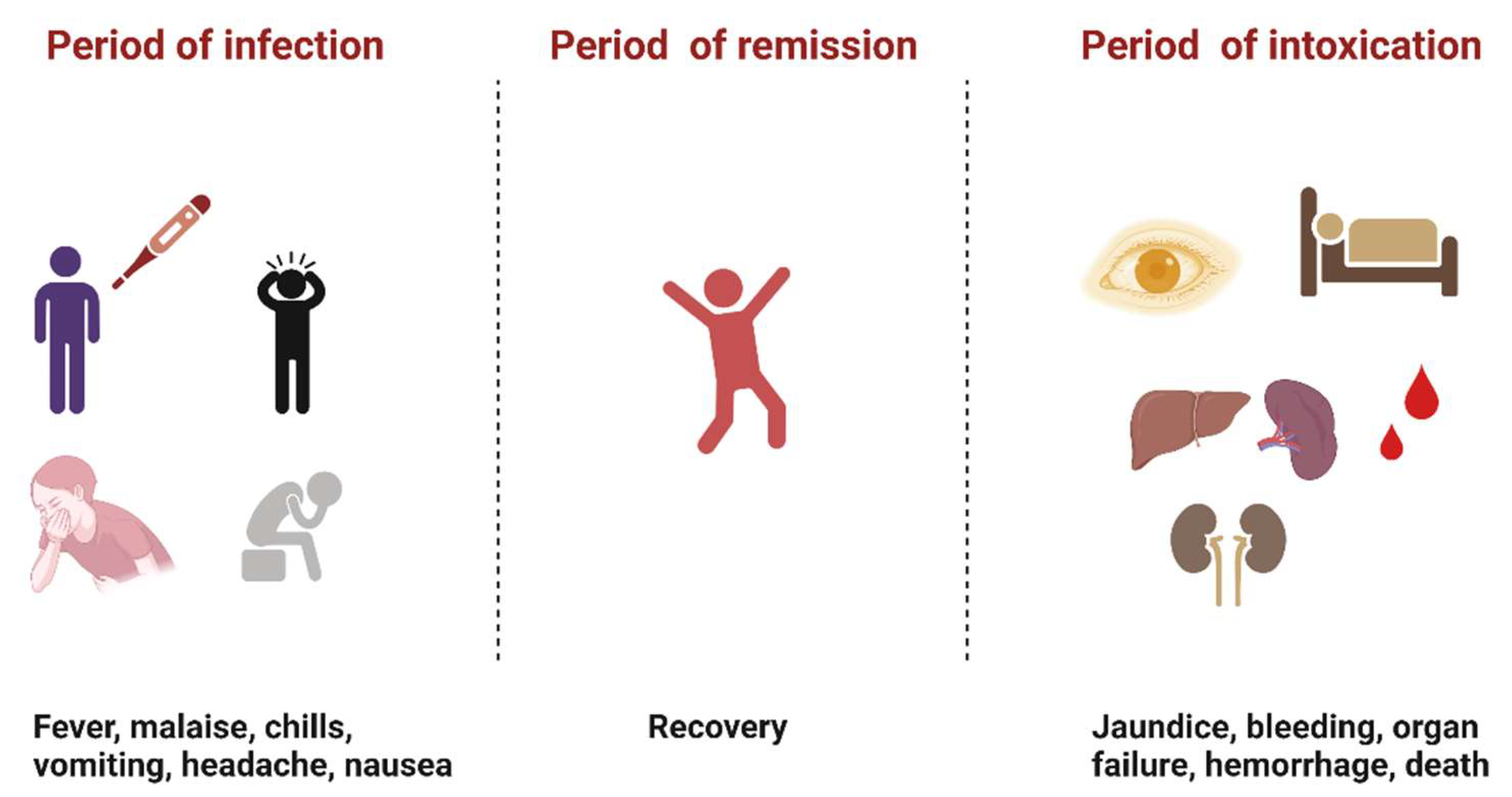
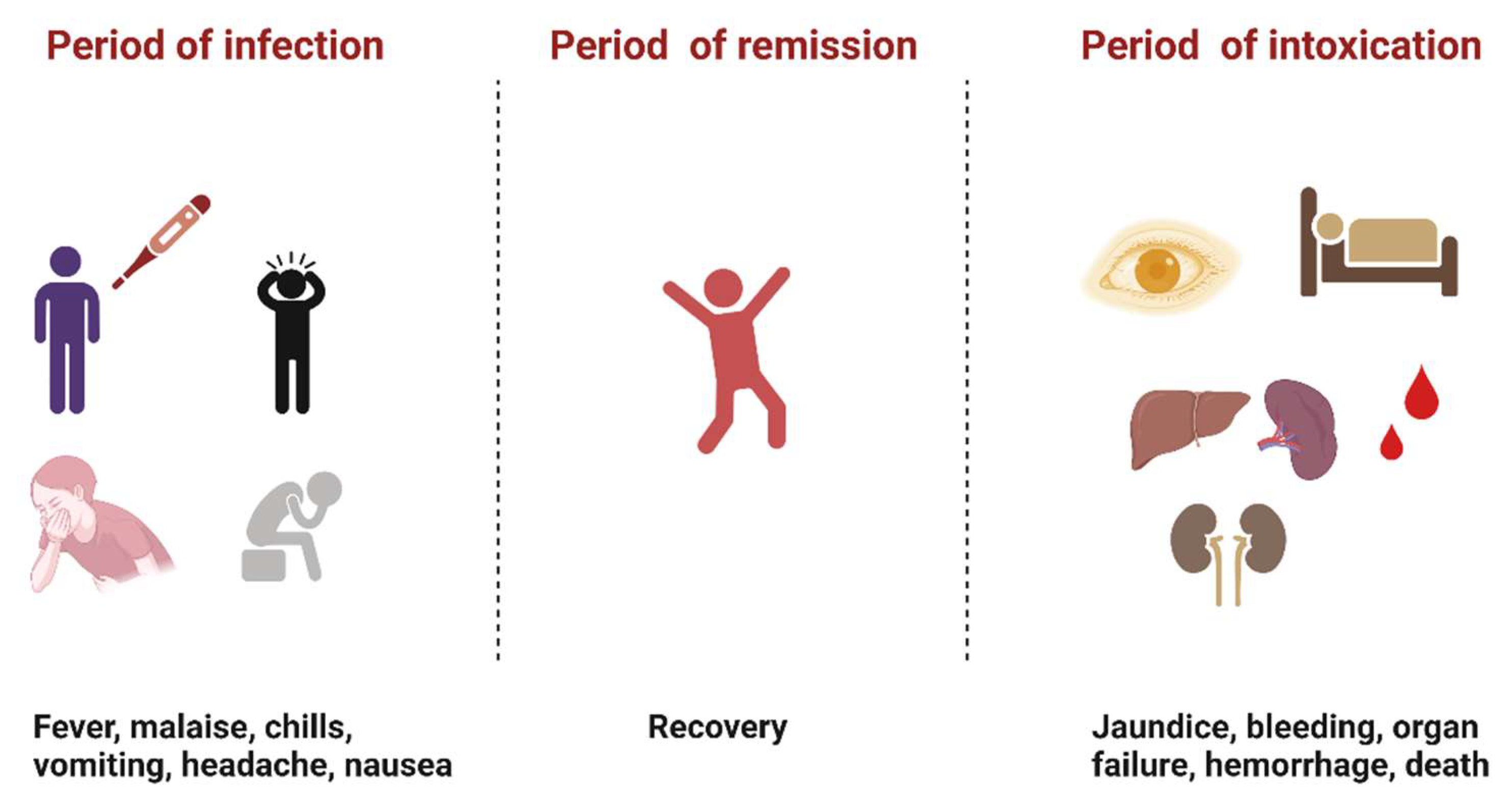
:1. Introduction
2. Brief History of YFV Research
3. Animal Models
3.1. Mice (Mus Musculus)

{kind=link}
{kind=link}
| Mouse | Age | Virus Strain | Route of Inoculation/Dose | Disease Outcome | Reference |
|---|---|---|---|---|---|
| Immunocompetent | |||||
| C57BL/6 | 3–4-week-old | 17D * | 1 IM; 104, 107 4 PFU | No signs of disease up to 30 days | [32] |
| 2 IP; 104 PFU | No signs of disease up to 30 days | [32] | |||
| WT129 | 3–4-week-old | 17D-204 # | 3 SQ (both footpad); 106 PFU | No signs of disease | [27] |
| Asibi; # Angola73 # | SQ (both footpad); 106 PFU | No signs of disease | [27] | ||
| Immunocompromised | |||||
| C57BL/6 (IFNAR−/−) | 3–4-week-old | 17D * | SQ; 104 PFU | No signs of disease | [32] |
| Footpad; 104 PFU | Transient swelling at site of inoc; fully recovered by 6 DPI | [32] | |||
| IM; 104 PFU | 32% of mice had neurotropic or viscerotropic disease | [32] | |||
| IM; 107 PFU | 42% of mice had neurotropic or viscerotropic | [32] | |||
| 6–7-week-old | 17D * | IM; 107 PFU | No signs of disease | [32] | |
| Independent of age | Angola71 # | SQ (both rear footpads); 2 × 104 PFU | 100% lethal by 6–9 DPI | [28] | |
| A129 | 3–4-week-old | 17D-204 # | SQ (both footpad); 106 PFU | No signs of disease | [27] |
| Asibi; # Angola 73 # | SQ (both footpad); 104 PFU | Clinical signs such as lethargy, hunched posture, swelling at inoculation site; weight loss; lethal by 7–8 DPI | [27] | ||
| AG129 | 3–4-week-old | 17D-204 # | SQ (both footpad); 106 PFU | Weight loss; lethal by 10–11 DPI | [27] |
| 6–7-week-old | 17D-204 † | IP; 104 PFU | Neurotropic; lethal; 5 AST (days): 17.2 ± 1.1 | [26] | |
| IP; 105 PFU | Neurotropic; lethal; AST (days): 15.2 ± 3.3 | [26] | |||
| IP; 106 PFU | Neurotropic; lethal; AST (days): 12.0 ± 3.6 | [26] | |||
| SQ; 104 PFU | Neurotropic; 70% mortality; AST (days): 20.8 ± 4.6 | [26] | |||
| SQ; 105 PFU | Neurotropic; lethal; AST (days): 16.8 ± 2.1 | [26] | |||
| 3–4-week-old | Asibi; # Angola73 # | SQ (both footpad); 104 PFU | Clinical signs such as lethargy, hunched posture, swelling at inoculation site; weight loss; lethal by 6–7 DPI | [27] | |
| G129 | 3–4-week-old | 17D-204 # | SQ (both footpad); 106 PFU | No signs of disease | [27] |
| Asibi; # Angola73 # | SQ (both footpad); 104 PFU | No signs of disease | [27] | ||
| STAT129 | 3–4-week-old | 17D-204 # | SQ (both footpad); 106 PFU | No signs of disease | [27] |
| Asibi # | SQ (both footpad); 104 PFU | Lethal by 7–8 DPI | [27] | ||
| STAT2−/− | Unknown | Asibi | Hock; 104 PFU | Viral burden and histopathological abormalities in spleen, liver; viremia on day 3 PI | [29] |
3.2. Hamsters (Mesocricetus Auratus)
3.3. Non-Human Primates
| NHP Species | YFV Strain | Dose/Route of Inoculation | Outcome | Reference |
|---|---|---|---|---|
| Old World NHPs | ||||
| Macaca mullata (Rhesus macaque) | DakH129 * | 25 1 TCID50; 2 SQ | 75% lethal (3/4) by 5–7 DPI; 39–79% hepatocellular damage in animals with lethal outcome | [47] |
| 102 TCID50; SQ | 75% lethal (3/4) by 5–7 DPI; 36–57% hepatocellular damage in animals with lethal outcome | [47] | ||
| 103 TCID50; SQ | 84% lethal (5/6) by 4–6 DPI; 33–77% hepatocellular damage in animals with lethal outcome | [47] | ||
| 104 TCID50; SQ | 50% lethal (1/2) by 4–5 DPI; 68% hepatocellular damage in the animal with lethal outcome | [47] | ||
| 5 × 104 TCID50; SQ | 75% lethal (3/4) by 4–5 DPI; 70–81% hepatocellular damage in animals with lethal outcome | [47] | ||
| Asibi | 800 adult mouse 3 ic LD50; 4 IP | 100% lethal (3/3) by 5–6 DPI | [53] | |
| French Viscerotropic | 8000 baby mouse ic LD50; 5 ID | 100% lethal (2/2) by 5–6 DPI | [53] | |
| JSS (South American) | 3 baby mouse ic LD50; SQ | 0% lethal (0/2) | [53] | |
| Macaca fascicularis (Cynomolgus macaque) | Asibi # | 900 6 PFU; SQ | 100% lethal (1/1) by Day 7 PI; Peak viremia 9 log10 PFU/mL; elevated levels of 7 AST and 8 ALT | [54] |
| Asibi # | 104 TCID50; SQ | 100% lethal (6/6) by 6 DPI; liver swelling, jaundice, pathological abnormalities in liver, spleen, lymph node, kidney; peak viremia 3 × 109 genome copies/mL; increased ALT, AST, AP, Bilirubin | [55] | |
| 17D # | 104 TCID50; SQ | No clinical signs of disease | [55] | |
| Galago crassicaudatus (Bush baby) | Unknown | Unknown | 50% lethal; viremia duration 4–8 days | [44] |
| Cercopithecus spp. (various species) | Unknown | Unknown | Intense viremia for 3–4 days followed by immunity | [44] |
| Colobus abyssinicus | East African strain | 2–6000 mouse LD50 | No apparent signs of disease; Viremia duration 6–9 days | [45] |
| Cercocebus spp. | Unknown | Unknown | Viremia duration 2 days (Only C. torquatus develops high enough viremia to infect mosquitoes) | [44] |
| Pan troglodytes (Chimpanzee) | African | 400 mouse LD50 | Viremia duration 3 days | [50] |
| Perodicticus potto (African potto) | Intense viremia, high enough to infect mosquitoes; viremia duration 4–8 days; no clinical signs of disease | [49] | ||
| New World NHPs | ||||
| Saimiri spp. (Squirrel monkey) | BeH655417 * (South American) | 1 × 106 PFU/mL; ID | Viscerotropic disease; pathological abnormalities and viral burden in liver, spleen, kidney, lymph node, heart, lung, brain, stomach | [56] |
| Callithrix albicollis (Marmoset) | Asibi | Unknown (blood transferred from other NHPs or via mosquito bites) | 33% lethal (2/6); febrile, pathological abnormalities in liver and kidney | [57] |
| S.R. | Unknown (blood transferred from other NHPs or via mosquito bites) | 80% lethal (4/5); febrile, pathological abnormalities in liver, kidney, spleen | [57] | |
| Leontocebus ursulus (Tamarin) | Asibi | Unknown (blood transferred from other NHPs or via mosquito bites) | 100% lethal (5/5); febrile, pathological abnormalities in liver and kidney | [57] |
| Cebus spp. (Capuchin) | Asibi; S.R. | Unknown (blood transferred from other NHPs or via mosquito bites) | Very low mortality, viremia may or may not be high enough to infect mosquitoes | [58] |
3.4. Other Animal Models
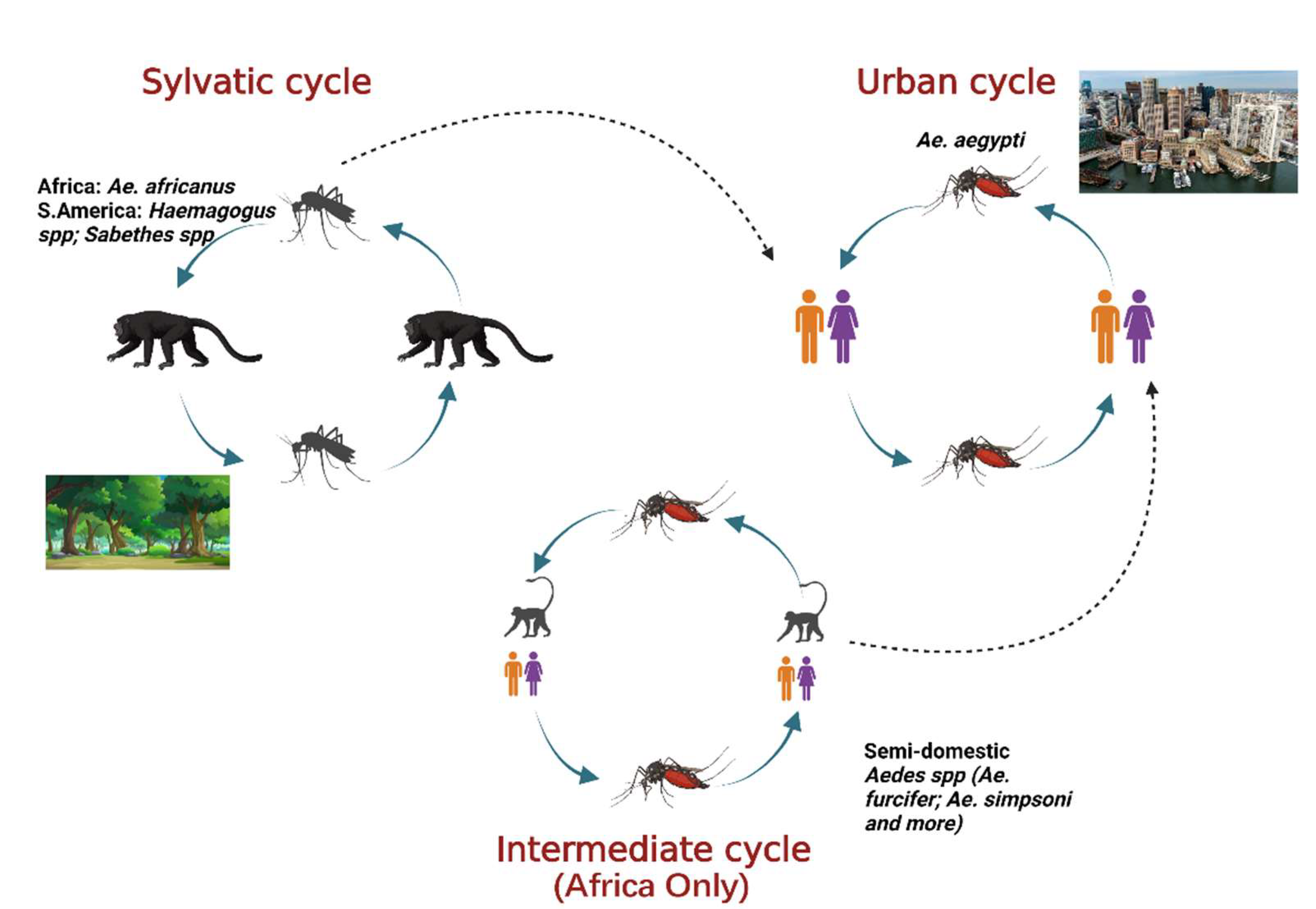
4. Arthropod Vectors
4.1. The Urban Cycle of Transmission
| Colony | Infectious Dose/Route | YFV Strain | Results | Reference |
|---|---|---|---|---|
| Santos, Brazil | 7–7.8 Log10 1 PFU/mL; 2 ABM | Brazilian (MG2001) | 4 IR: 35%, 6 TR: 28% at 11–14 3 DPF | [72] |
| 6.3 Log10 PFU/mL; ABM | Brazilian (MG2001) | TR: 23% at 21 DPF | [72] | |
| Respublic of Vanuatu | 7–7.8 Log10 PFU/mL; ABM | Brazilian (MG2001) | IR: 18%, TR: 12% at 11–14 DPF | [72] |
| 6.3 Log10 PFU/mL; ABM | Brazilian (MG2001) | TR: 17% at 21 DPF | [72] | |
| Goias, Brazil | 6 Log10 PFU/mL; ABM | Brazilian (4408-1E) | IR: 0, ~30, ~70, ~10 % at 3, 7, 14 and 21 DPF 5 DR: 0, ~25, ~70, 100% at 3, 7, 14 and 21 DPF TR: 0, 0, ~20, ~50% at 3, 7, 14 and 21 DPF | [66] |
| Brazilian (74018-1D) | IR: 0, ~30, ~80, ~65% at 3, 7, 14 and 21 DPF DR: 0, ~35, ~65, ~90% at 3, 7, 14 and 21 DPF TR: 0, 0, ~20, 0% at 3, 7, 14 and 21 DPF | [66] | ||
| Senegalese (S-79) | IR: 0, ~30, ~80, 0% at 3, 7, 14 and 21 DPF DR: 0, ~35, ~75,0% at 3, 7, 14 and 21 DPF TR: 0% at 3, 7, 14 and 21 DPF | [66] | ||
| Manaus, Brazil | 6 Log10 PFU/mL; ABM | Brazilian (4408-1E) | IR: ~55%, DR:~85%, TR: ~25% at 14–21 DPF | [66] |
| Brazilian (74018-1D) | IR: ~55%, DR: ~60%, TR: ~15% at 14–21 DPF | [66] | ||
| Senegalese (S-79) | IR: ~30%, DR: ~50%, TR: ~35% at 14–21 DPF | [66] | ||
| Rio, Brazil | 6 Log10 PFU/mL; ABM | Brazilian (4408-1E) | IR: ~85%, DR: ~60%, TR: ~60% at 14–21 DPF | [66] |
| Brazilian (74018-1D) | IR: ~45%, DR: ~60%, TR: ~35% at 14–21 DPF | [66] | ||
| Senegalese (S-79) | IR: ~50%, DR: ~65%, TR: ~40% at 14–21 DPF | [66] | ||
| Fatick, Senegal | 6.22 Log10 PFU/mL; ABM | Nigerian (BA-55) | 7 MIR: 100%, 8 DIR:59% at 14 DPF | [70] |
| 5.9 Log10 PFU/mL; ABM | Nigerian (DAK 1279) | MIR: 17%, DIR: 0% at 14 DPF | [70] | |
| Bignona, Senegal | 6.22 Log10 PFU/mL; ABM | Nigerian (BA-55) | MIR: 83%, DIR: 13% at 14 DPF | [70] |
| 7.79 Log10 PFU/mL; ABM | Nigerian (DAK 1279) | MIR: 33%, DIR:0% at 14 DPF | [70] | |
| Richard Toll, Senegal | 6.32 Log10 PFU/mL; ABM | Nigerian (BA-55) | MIR: 57%, DIR: 10% at 14 DPF | [70] |
| 7.79 Log10 PFU/mL; ABM | Nigerian (DAK 1279) | MIR: 57%, DIR: 17% at 14 DPF | [70] | |
| Goudiry, Senegal | 6.04 Log10 PFU/mL; ABM | Nigerian (BA-55) | MIR: 53%, DIR: 0% at 14 DPF | [70] |
| 5.9 Log10 PFU/mL; ABM | Nigerian (DAK 1279) | MIR: 10%, DIR: 0% at 14 DPF | [70] | |
| Kedougou, Senegal | 5.34 Log10 PFU/mL; ABM | Nigerian (BA-55) | MIR: 35%, DIR: 0% at 14 DPF | [70] |
| 5.9 Log10 PFU/mL; ABM | Nigerian (DAK 1279) | MIR: 10%, DIR: 0% at 14 DPF | [70] | |
| PK10, Senegal | 6.04 Log10 PFU/mL; ABM | Nigerian (BA-55) | MIR: 27%, DIR: 3% at 14 DPF | [70] |
| 5.9 Log10 PFU/mL; ABM | Nigerian (DAK 1279) | MIR: 22%, DIR: 0% at 14 DPF | [70] | |
| Mont Rolland, Senegal | 6.2 Log10 PFU/mL; ABM | Nigerian (BA-55) | MIR: 27%, DIR: 0% at 14 DPF | [70] |
| 5.9 Log10 PFU/mL; ABM | Nigerian (DAK 1279) | MIR: 20%, DIR: 3% at 14 DPF | [70] | |
| Rufisque, Senegal | 6.13 Log10 PFU/mL; ABM | Nigerian (BA-55) | MIR: 17%, DIR: 0% at 14 DPF | [70] |
| 5.9 Log10 PFU/mL; ABM | Nigerian (DAK 1279) | MIR: 11%, DIR: 0% | [70] | |
| Cairns, Australia | 7.2 Log10 TCID50/mL; ABM | Nigerian (BA-55) | IR: 80%, DIR: 72%, 9 TIR: 52% at 14 DPF | [71] |
| 6.7 Log10 TCID50/mL; ABM | Bolivian (Cinetrop 28) | IR: 24%, DIR: 24%, TIR: 24% at 15 DPF | [71] | |
| 8 Log10 TCID50/mL; ABM | African (Asibi) | IR: 92%, DIR: 80% At 14 DPF | [71] | |
| Townsville, Australia | 7.2 Log10 TCID50/mL; ABM | Nigerian (BA-55) | IR: 72%, DIR: 60%, TIR: 60% at 14 DPF | [71] |
| 6.7 Log10 TCID50/mL; ABM | Bolivian (Cinetrop 28) | IR: 36%, DIR: 32%, TIR: 28% at 14 DPF | [71] | |
| 8 Log10 TCID50/mL ABM | African (Asibi) | IR: 96%, DIR: 100% | [71] | |
| Cambodia, Asia | 7 Log10 PFU/mL; ABM | Senegalese (S-79) | IR: 40%, DR: 60%, TR: 0% at 14 DPF IR: 40%, DR: 90%, TR: 35% at 21 DPF | [73] |
| Vietnam, Asia | 7 Log10 PFU/mL; ABM | Senegalese (S-79) | IR: 60%, DR: 70%, TR: 0% at 14 DPF IR: 70%, DR: 55%, TR: 5% at 21 DPF | [73] |
| Trung, Asia | 7 Log10 PFU/mL; ABM | Senegalese (S-79) | IR: 95%, DR: 75%, TR: 0% at 14 DPF IR: 90%, DR: 90%, TR: 30% at 21 DPF | [73] |
| Laos, Asia | 7 Log10 PFU/mL; ABM | Senegalese (S-79) | IR: 75%, DR: 62%, TR: 0% at 14 DPF IR: 100%, DR: 90%, TR: 40% at 21 DPF | [73] |
| Thailand, Asia | 7 Log10 PFU/mL; ABM | Senegalese (S-79) | IR: 100%, DR: 85%, TR: 45% at 14 DPF IR: 100%, DR: 100%, TR: 58% at 21 DPF | [73] |
| Singapore, Asia | 7 Log10 PFU/mL; ABM | Senegalese (S-79) | IR: 80%, DR: 80%, TR: 10% at 14 DPF IR: 100%, DR: 50%, TR: 5% at 21 DPF | [73] |
| New Caledonia, Asia | 7 Log10 PFU/mL; ABM | Senegalese (S-79) | IR: 100%, DR: 100%, TR: 10% at 14 DPF IR: 100%, DR: 75%, TR: 25% at 21 DPF | [73] |
| Taiwan, Asia | 7 Log10 PFU/mL; ABM | Senegalese (S-79) | IR: 65%, DR: 55%, TR: 0% at 14 DPF IR: 85%, DR: 50%, TR: 50% at 21 DPF | [73] |
4.2. The Sylvatic Cycle of Transmission
5. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Simmonds, P.; Becher, P.; Bukh, J.; Gould, E.A.; Meyers, G.; Monath, T.; Muerhoff, S.; Pletnev, A.; Rico-Hesse, R.; Smith, D.B.; et al. ICTV Virus Taxonomy Profile: Flaviviridae. J. Gen. Virol. 2017, 98, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Rice, C.M.; Lenches, E.M.; Eddy, S.R.; Shin, S.J.; Sheets, R.L.; Strauss, J.H. Nucleotide sequence of yellow yever virus: Implications for flavivirus gene expression and evolution. Science 1985, 229, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Hanley, K.A.; Monath, T.P.; Weaver, S.C.; Rossi, S.L.; Richman, R.L.; Vasilakis, N. Fever versus Fever: The role of host and vector susceptibility and interspecific competition in shaping the current and future distributions of the sylvatic cycles of dengue virus and yellow fever virus. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2013, 19, 292–311. [Google Scholar] [CrossRef] [PubMed]
- Angola: Epidemic (Yellow Fever) Emergency Plan of Action Preliminary Final Report (MDRAO006)—Angola|ReliefWeb. Available online: https://reliefweb.int/report/angola/angola-epidemic-yellow-fever-emergency-plan-action-preliminary-final-report-mdrao006 (accessed on 30 June 2022).
- Germain, M.; Cornet, M.; Mouchet, J.; Monath, T.P.; Hervé, J.P.; Salaun, J.J.; Cordellier, R.; Saluzzo, J.F.; Camicas, J.L.; Hervy, J.P.; et al. Recent advances in research regarding sylvatic yellow fever in West and Central Africa. Bull. De L’institut Pasteur 1982, 80, 315–330. Available online: https://horizon.documentation.ird.fr/exl-doc/pleins_textes/pleins_textes_5/b_fdi_04-05/03774.pdf (accessed on 21 July 2022).
- Abreu, F.V.S.D.; Ribeiro, I.P.; Ferreira-de-Brito, A.; Santos, A.A.C.D.; Miranda, R.M.D.; Bonelly, I.D.S.; Neves, M.S.A.S.; Bersot, M.I.; Santos, T.P.D.; Gomes, M.Q.; et al. Haemagogus leucocelaenus and Haemagogus janthinomys are the primary vectors in the major yellow fever outbreak in Brazil, 2016–2018. Emerg. Microbes Infect. 2019, 8, 218–231. [Google Scholar] [CrossRef]
- Mutebi, J.-P.; Barrett, A.D.T. The epidemiology of yellow fever in Africa. Microbes Infect. 2002, 4, 1459–1468. [Google Scholar] [CrossRef]
- Yellow Fever. Available online: https://www.who.int/news-room/fact-sheets/detail/yellow-fever (accessed on 18 May 2022).
- Silva, N.I.O.; Sacchetto, L.; de Rezende, I.M.; Trindade, G.D.S.; LaBeaud, A.D. Recent sylvatic yellow fever virus transmission in Brazil: The news from an old disease. Virol. J. 2020, 17, 9. [Google Scholar] [CrossRef]
- Gershman, M.D. Addressing a yellow fever vaccine shortage—United States, 2016–2017. MMWR Morb. Mortal Wkly. Rep. 2017, 66, 457. [Google Scholar] [CrossRef]
- Monath, T.P.; Cetron, M.S.; McCarthy, K.; Nichols, R.; Archambault, W.T.; Weld, L.; Bedford, P. Yellow fever 17D vaccine safety and immunogenicity in the elderly. Hum. Vaccines 2005, 1, 207–214. [Google Scholar] [CrossRef]
- Monath, T.P.; Barrett, A.D.T. Pathogenesis and Pathophysiology of Yellow Fever. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2003; Volume 60, pp. 343–395. [Google Scholar] [CrossRef]
- Symptoms, Diagnosis, & Treatment 26 February 2020. Available online: https://www.cdc.gov/yellowfever/symptoms/index.html (accessed on 18 May 2022).
- Bryant, J.E.; Holmes, E.C.; Barrett, A.D.T. Out of Africa: A molecular perspective on the introduction of yellow fever virus into the Americas. PLoS Pathog. 2007, 3, e75. [Google Scholar] [CrossRef]
- To, K.K.; Yuen, K.-Y. In memory of Patrick Manson, founding father of tropical medicine and the discovery of vector-borne infections. Emerg. Microbes Infect. 2012, 1, 1–7. [Google Scholar] [CrossRef]
- Clements, A.N.; Harbach, R.E. History of the discovery of the mode of transmission of yellow fever virus. J. Vector Ecol. J. Soc. Vector Ecol. 2017, 42, 208–222. [Google Scholar] [CrossRef]
- Finlay, C. The mosquito hypothetically considered as an agent in the transmission of yellow fever poison. Yale J. Biol. Med. 1973, 9, 589–604. [Google Scholar]
- Reed, W.; Carroll, J.; Agramonte, A.; Lazear, J.W. The etiology of yellow fever—A preliminary note. Public Health Pap. Rep. 1900, 26, 37–53. [Google Scholar]
- Sanitation in Panama/by William Crawford Gorgas. Wellcome Collection. Available online: https://wellcomecollection.org/works/f97wdqvn/items (accessed on 18 May 2022).
- Hudson, N.P. Adrian stokes and yellow fever research: A tribute. Trans. R. Soc. Trop. Med. Hyg. 1966, 60, 170–174. [Google Scholar] [CrossRef]
- Norrby, E. Yellow fever and Max Theiler: The only Nobel Prize for a virus vaccine. J. Exp. Med. 2007, 204, 2779–2784. [Google Scholar] [CrossRef] [PubMed]
- Camargo, S. History of Aedes aegypti eradication in the Americas. Bull. World Health Organ. 1967, 36, 602–603. [Google Scholar] [PubMed]
- Aedes Aegypti Eradication Program; Communicable Disease Center: Atlanta, GA, USA, 1966.
- Soper, F.L. The elimination of urban yellow fever in the Americas through the eradication of Aedes aegypti. Am. J. Public Health Natl. Health 1963, 53, 7–16. [Google Scholar] [CrossRef]
- Status of Aedes aegypti eradication in Americas. Pan American Health Organization Meeting, August 1967. Available online: https://iris.paho.org/bitstream/handle/10665.2/5757/48939.pdf?sequence=1&isAllowed=y (accessed on 17 May 2022).
- Thibodeaux, B.A.; Garbino, N.C.; Liss, N.M.; Piper, J.; Blair, C.D.; Roehrig, J.T. A small animal peripheral challenge model of yellow fever using interferon-receptor deficient mice and the 17D-204 vaccine strain. Vaccine 2012, 30, 3180–3187. [Google Scholar] [CrossRef]
- Meier, K.C.; Gardner, C.L.; Khoretonenko, M.V.; Klimstra, W.B.; Ryman, K.D. A mouse model for studying viscerotropic disease caused by yellow fever virus infection. PLoS Pathog. 2009, 5, e1000614. [Google Scholar] [CrossRef]
- Watson, A.M.; Lam, L.K.M.; Klimstra, W.B.; Ryman, K.D. The 17D-204 vaccine strain-induced protection against virulent yellow fever virus is mediated by humoral immunity and CD4+ but not CD8+ T Cells. PLoS Pathog. 2016, 12, e1005786. [Google Scholar] [CrossRef]
- Miorin, L.; Laurent-Rolle, M.; Pisanelli, G.; Co, P.H.; Albrecht, R.A.; García-Sastre, A.; Morrison, J. Host-Specific NS5 Ubiquitination Determines Yellow Fever Virus Tropism. J. Virol. 2019, 93, e00151-19. [Google Scholar] [CrossRef]
- Strom, S.C.; Davila, J.; Grompe, M. Chimeric mice with humanized liver: Tools for the study of drug metabolism, excretion, and toxicity. Methods Mol. Biol. 2010, 640, 491–509. [Google Scholar] [CrossRef]
- Bailey, A.L.; Kang, L.I.; De Assis Barros, D. Consumptive coagulopathy of severe yellow fever occurs independently of hepatocellular tropism and massive hepatic injury. Proc. Natl. Acad. Soc. USA 2020, 117, 32648–32656. [Google Scholar] [CrossRef]
- Erickson, A.K.; Pfeiffer, J.K. Spectrum of disease outcomes in mice infected with YFV-17D. J. Gen. Virol. 2015, 96, 1328–1339. [Google Scholar] [CrossRef]
- Tesh, R.B.; Guzman, H.; da Rosa, A.P.; Vasconcelos, P.F.; Dias, L.B.; Bunnell, J.E.; Zhang, H.; Xiao, S.Y. Experimental yellow fever virus infection in the Golden Hamster (Mesocricetus auratus). I. Virologic, biochemical, and immunologic studies. J. Infect. Dis. 2001, 183, 1431–1436. [Google Scholar] [CrossRef]
- Xiao, S.; Zhang, H.; Guzman, H.; Tesh, R.B. Experimental yellow fever virus infection in the Golden Hamster (Mesocricetus auratus). II. Pathology. J. Infect. Dis. 2001, 183, 1437–1444. [Google Scholar] [CrossRef]
- McArthur, M.A.; Zhang, S.L.; Li, L.; Tesh, R.B.; Barrett, A.D.T. Molecular characterization of hamster-adapted yellow fever virus. Vector Borne Zoonotic Dis. 2020, 20, 222–227. [Google Scholar] [CrossRef]
- McArthur, M.A.; Suderman, M.T.; Mutebi, J.-P.; Xiao, S.-Y.; Barrett, A.D.T. Molecular characterization of a hamster viscerotropic strain of yellow fever virus. J. Virol. 2003, 77, 1462–1468. [Google Scholar] [CrossRef]
- Kumm, H.W.; Laemmert, H.W., Jr. A study of the concentration of yellow fever virus which will infect certain species of Aedes mosquitoes. Am. J. Trop. Med. 1950, 30, 1950. Available online: https://www.cabdirect.org/cabdirect/abstract/19531000394 (accessed on 14 July 2021). [CrossRef]
- Julander, J.G.; Furuta, Y.; Shafer, K.; Sidwell, R.W. Activity of T-1106 in a hamster model of yellow fever virus Infection. Antimicrob. Agents Chemother. 2007, 51, 1962–1966. [Google Scholar] [CrossRef]
- Julander, J.G.; Shafer, K.; Smee, D.F.; Morrey, J.D.; Furuta, Y. Activity of T-705 in a hamster model of yellow fever virus infection in comparison with that of a chemically related compound, T-1106. Antimicrob. Agents Chemother. 2009, 53, 202–209. [Google Scholar] [CrossRef]
- Sbrana, E.; Xiao, S.; Guzman, H.; Ye, M.; Tesh, R.B. Efficacy of post-exposure treatment of yellow fever with ribavirin in a hamster model of the disease. Am. J. Trop. Med. Hyg. 2004, 71, 306–312. [Google Scholar] [CrossRef]
- Monath, T.P.; Lee, C.K.; Julander, J.G.; Brown, A.; Beasley, D.W.; Watts, D.M.; Hayman, E.; Guertin, P.; Makowiecki, J.; Crowell, J.; et al. Inactivated yellow fever 17D vaccine: Development and nonclinical safety, immunogenicity and protective activity. Vaccine 2010, 28, 3827–3840. [Google Scholar] [CrossRef]
- Julander, J.G.; Trent, D.W.; Monath, T.P. Immune correlates of protection against yellow fever determined by passive immunization and challenge in the hamster model. Vaccine 2011, 29, 6008–6016. [Google Scholar] [CrossRef]
- Sacchetto, L.; Silva, N.I.O.; Rezende, I.M.D.; Arruda, M.S.; Costa, T.A.; de Mello, E.M.; Oliveira, G.F.G.; Alves, P.A.; de Mendonca, V.E.; Stumpp, R.G.A.V.; et al. Neighbor danger: Yellow fever virus epizootics in urban and urban-rural transition areas of Minas Gerais state, during 2017-2018 yellow fever outbreaks in Brazil. PLoS Negl. Trop. Dis. 2020, 14, e0008658. [Google Scholar] [CrossRef]
- Rodhain, F. The role of monkeys in the biology of dengue and yellow fever. Comp. Immunol. Microbiol. Infect. Dis. 1991, 1, 9–19. [Google Scholar] [CrossRef]
- Woodall, J.P.; Dykes, J.R.W.; Williams, M.C. The reaction of a species of colobus monkey to inoculation with yellow fever virus. Ann. Trop. Med. Parasitol. 1968, 62, 528–535. [Google Scholar] [CrossRef]
- Stokes, A.; Bauer, J.H.; Hudson, N.P.; Mortimer, R.B.P.P. The transmission of yellow fever to Macacus Rhesus. Rev. Med. Virol. 2001, 11, 141–148. [Google Scholar] [CrossRef]
- Engelmann, F.; Josset, L.; Girke, T.; Park, B.; Barron, A.; Dewane, J.; Hammarlund, E.; Lewis, A.; Axthelm, M.K.; Slifka, M.K.; et al. Pathophysiologic and transcriptomic analyses of viscerotropic yellow fever in a rhesus macaque model. PLoS Negl. Trop. Dis. 2014, 8, e3295. [Google Scholar] [CrossRef]
- Monath, T.P.; Brinker, K.R.; Cropp, C.B.; Chandler, F.W.; Kemp, G.E. Pathophysiologic correlations in a rhesus monkey model of yellow fever: With special observations on the acute necrosis of B cell areas of lymphoid tissues*. Am. J. Trop. Med. Hyg. 1981, 30, 431–443. [Google Scholar] [CrossRef]
- Risk assessment on yellow fever virus circulation in endemic countries. Available online: https://apps.who.int/iris/bitstream/handle/10665/112751/WHO_HSE_PED_CED_2014.2_eng.pdf?sequence=1 (accessed on 15 February 2022).
- Smithburn, K.C.; Haddow, A.J. The susceptibility of African wild animals to yellow fever: I. Monkeys. Am. J. Trop. Med. Hyg. 1949, s1–s29, 389–423. [Google Scholar] [CrossRef]
- Galindo, P. Monkeys and Yellow Fever. In Non-Human Primates and Medical Research; Academic Press, Inc.: New York, NY, USA; London, UK, 1973; Available online: http://www.gorgas.gob.pa/BiblioGorgas/pdf/monkey%20and%20yellow%20fever.%20pedro%20galindo.pdf (accessed on 5 June 2022).
- da Vasconcelos, P.F.C. Febre amarela. Rev. Soc. Bras. Med. Trop. 2003, 36, 275–293. [Google Scholar] [CrossRef]
- Theiler, M.; Anderson, C.R. The relative resistance of dengue-immune monkeys to yellow fever virus. Am. J. Trop. Med. Hyg. 1975, 24, 115–117. [Google Scholar] [CrossRef]
- Barrett, A.D.T.; Monath, T.P.; Cropp, C.B.; Adkins, J.A.; Ledger, T.N.; Gould, E.A.; Schlesinger, J.J.; Kinney, R.M.; Trent, D.W. Attenuation of wild-type yellow fever virus by passage in HeLa cells. J. Gen. Virol. 1990, 71, 2301–2306. [Google Scholar] [CrossRef]
- McGee, C. Evaluation of arthropod-Borne Virus Recombination Potential and Phenotype: In Vitro and In Vivo Analysis of Flavivirus and Alphavirus Model Systems; University of Texas Medical Branch: Galveston, TX, USA, 2010; Available online: https://utmb-ir.tdl.org/bitstream/handle/2152.3/117/CEMcGeeDissertationFinalFinal.pdf?sequence=1&isAllowed=y (accessed on 1 June 2022).
- Ferreira, M.S.; Júnior, P.S.B.; Cerqueira, V.D.; Rivero, G.R.C.; Júnior, C.A.O.; Castro, P.H.G.; Silva, G.A.D.; Silva, W.B.D.; Imbeloni, A.A.; Sousa, J.R.; et al. Experimental yellow fever virus infection in the squirrel monkey (Saimiri spp.) I: Gross anatomical and histopathological findings in organs at necropsy. Memórias Do Inst. Oswaldo Cruz 2020, 115, e190501. [Google Scholar] [CrossRef]
- Davis, N.C. The susceptibility of marmosets to yellow fever virus. J. Exp. Med. 1930, 52, 405–416. [Google Scholar] [CrossRef]
- Davis, N.C. Susceptibility of capuchin (Cebus) monkeys to yellow fever virus. Am. J. Epidemiol. 1930, 11, 321–334. [Google Scholar] [CrossRef]
- Whitman, L. The response to yellow fever virus in the nonsusceptible rabbit. J. Immunol. 1935, 29, 12. [Google Scholar]
- Hughes, T.P.; Perlowagora, A. The application of immunological tests of sera from captured wild animals to the study of yellow fever epidemiology. Am. J. Trop. Med. Hyg. 1950, s1-30, 835–853. [Google Scholar] [CrossRef]
- Powell, J.R.; Gloria-Soria, A.; Kotsakiozi, P. Recent History of Aedes aegypti: Vector Genomics and Epidemiology Records. Bioscience 2018, 68, 854–860. [Google Scholar] [CrossRef]
- Christophers, S.R. Aedes Aegypti: The Yellow Fever Mosquito. In Aedes Aegypti: The Yellow Fever Mosquito. Its Life History, Binomics and Structure; The Syndics of The Cambridge University Press: London, UK, 1960. [Google Scholar]
- Crawford, J.E.; Alves, J.M.; Palmer, W.J.; Day, J.P.; Sylla, M.; Ramasamy, R.; Surendran, S.N.; Black, W.C.; Pain, A.; Jiggins, F.M.; et al. Population genomics reveals that an anthropophilic population of Aedes aegypti mosquitoes in West Africa recently gave rise to American and Asian populations of this major disease vector. BMC Biol. 2017, 15, 16. [Google Scholar] [CrossRef]
- Tabachnick, W.J.; Powell, J.R. A world-wide survey of genetic variation in the yellow fever mosquito, Aedes aegypti*. Genet. Res. 1979, 34, 215–229. [Google Scholar] [CrossRef]
- Failloux, A.-B.; Vazeille, M.; Rodhain, F. Geographic Genetic Variation in Populations of the Dengue Virus Vector Aedes aegypti. J. Mol. Evol. 2002, 55, 653–663. [Google Scholar] [CrossRef]
- Couto-Lima, D.; Madec, Y.; Bersot, M.I.; Campos, S.S.; Motta, M.D.A.; Santos, F.B.D.; Vazeille, M.; Vasconcelos, P.F.D.C.; Lourenço-de-Oliveira, R.; Failloux, A.B. Potential risk of re-emergence of urban transmission of Yellow Fever virus in Brazil facilitated by competent Aedes populations. Sci. Rep. 2017, 7, 4848. [Google Scholar] [CrossRef]
- Mutebi, J.-P.; Gianella, A.; da Rosa, A.T.; Tesh, R.B.; Barrett, A.D.T.; Higgs, S. Yellow fever virus infectivity for Bolivian Aedes aegypti mosquitoes. Emerg. Infect. Dis. 2004, 10, 1657–1660. [Google Scholar] [CrossRef]
- Lourenço-de-Oliveira, R.; Vazeille, M.; de Filippis, A.M.B.; Failloux, A.-B. Oral susceptibility to yellow fever virus of Aedes aegypti from Brazil. Memórias Do Inst. Oswaldo Cruz 2002, 97, 437–439. [Google Scholar] [CrossRef]
- Souza-Neto, J.A.; Powell, J.R.; Bonizzoni, M. Aedes aegypti vector competence studies: A review. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2019, 67, 191–209. [Google Scholar] [CrossRef]
- Dickson, L.B.; Sanchez-Vargas, I.; Sylla, M.; Fleming, K.; Iv, W.C.B. Vector competence in West African Aedes aegypti Is flavivirus species and genotype dependent. PLoS Negl. Trop. Dis. 2014, 8, e3153. [Google Scholar] [CrossRef]
- Van Den Hurk, A.F.; McElroy, K.; Pyke, A.T.; McGee, C.E.; Hall-Mendelin, S.; Day, A.; Ryan, P.A.; Ritchie, S.A.; Vanlandingham, D.L.; Higgs, S. Vector competence of Australian mosquitoes for yellow fever virus. Am. J. Trop. Med. Hyg. 2011, 85, 446–451. [Google Scholar] [CrossRef]
- Johnson, B.W.; Chambers, T.V.; Crabtree, M.B.; Filippis, A.M.; Vilarinhos, P.T.; Resende, M.C.; Macoris, M.D.L.G.; Miller, B.R. Vector competence of Brazilian Aedes aegypti and Ae. albopictus for a Brazilian yellow fever virus isolate. Trans. R. Soc. Trop. Med. Hyg. 2002, 96, 611–613. [Google Scholar] [CrossRef]
- Lataillade, L.D.G.D.; Vazeille, M.; Obadia, T.; Madec, Y.; Mousson, L.; Kamgang, B.; Chen, C.H.; Failloux, A.B.; Yen, P.S. Risk of yellow fever virus transmission in the Asia-Pacific region. Nat. Commun. 2020, 11, 5801. [Google Scholar] [CrossRef] [PubMed]
- Diallo, D.; Diagne, C.T.; Hanley, K.A.; Sall, A.A.; Buenemann, M.; Ba, Y.; Dia, I.; Weaver, S.C.; Diallo, M. Larval ecology of mosquitoes in sylvatic arbovirus foci in southeastern Senegal. Parasites Vectors 2012, 5, 286. [Google Scholar] [CrossRef]
- Monath, T.P.; Vasconcelos, P.F.C. Yellow fever. J. Clin. Virol. 2015, 64, 160–173. [Google Scholar] [CrossRef] [PubMed]
- de Rodaniche, E.; Galindo, P. Isolation of yellow fever virus from Haemagogus mesodentatus, H. equinus and Sabethes chloropterus captured in Guatemala in 1956. Am. J. Trop. Med. Hyg. 1957, 6, 232–237. [Google Scholar] [CrossRef]
- Mondet, B.; Vasconcelos, P.F.C.; Travassos da Rosa, A.P.A.; Travassos da Rosa, E.S.; Rodrigues, S.G.; Travassos da Rosa, J.F.S.; Bicout, D.J. Isolation of yellow fever virus from nulliparous Haemagogusjanthinomys in Eastern Amazonia. Vector-Borne Zoonotic Dis. 2002, 2, 47–50. [Google Scholar] [CrossRef] [PubMed]
- ter Meulen, J.; Sakho, M.; Koulemou, K.; Magassouba, N.F.; Bah, A.; Preiser, W.; Daffis, S.; Klewitz, C.; Bae, H.G.; Niedrig, M.; et al. Activation of the cytokine network and unfavorable outcome in patients with yellow fever. J. Infect. Dis. 2004, 190, 1821–1827. [Google Scholar] [CrossRef]
- Quaresma, J.A.; Barros, V.L.; Pagliari, C.; Fernandes, E.R.; Guedes, F.; Takakura, C.F.; Andrade, H.F., Jr.; Vasconcelos, P.F.; Duarte, M.I. Revisiting the liver in human yellow fever: Virus-induced apoptosis in hepatocytes associated with TGF-β, TNF-α and NK cells activity. Virology 2006, 345, 22–30. [Google Scholar] [CrossRef]
- Quaresma, J.A.S.; Barros, V.L.R.S.; Fernandes, E.R.; Pagliari, C.; Takakura, C.; da Costa Vasconcelos, P.F.; de Andrade, H.F., Jr.; Duarte, M.I.S. Reconsideration of histopathology and ultrastructural aspects of the human liver in yellow fever. Acta Trop. 2005, 94, 116–127. [Google Scholar] [CrossRef]
- Tabachnick, W.J.; Wallis, G.P.; Aitken, T.H.; Miller, B.R.; Amato, G.D.; Lorenz, L.; Powell, J.R.; Beaty, B.J. Oral infection of Aedes aegypti with yellow fever virus: Geographic variation and genetic consideration. Am. J. Trop. Med. Hyg. 1985, 34, 1219–1224. [Google Scholar] [CrossRef]
- Lorenz, L.; Beaty, B.J.; Aitken, T.H.G.; Wallis, G.P.; Tabachnick, W.J. The effect of colonization upon Aedes aegypti susceptibility to oral infection with yellow fever virus*. Am. J. Trop. Med. Hyg. 1984, 33, 690–694. [Google Scholar] [CrossRef]
- Miot, E.F.; Aubry, F.; Dabo, S.; Mendenhall, I.H.; Marcombe, S.; Tan, C.H.; Ng, L.C.; Failloux, A.B.; Pompon, J.; Brey, P.T.; et al. A peridomestic Aedes malayensis population in Singapore can transmit yellow fever virus. PLoS Negl. Trop. Dis. 2019, 13, e0007783. [Google Scholar] [CrossRef] [PubMed]
- Kuno, G. The absence of yellow fever in Asia: History, hypotheses, vector dispersal, possibility of YF in Asia, and other enigmas. Viruses 2020, 12, 1349. [Google Scholar] [CrossRef] [PubMed]
- Agampodi, S.B.; Wickramage, K. Is there a risk of yellow fever virus transmission in South Asian countries with hyperendemic dengue? BioMed. Res. Int. 2013, 2013, e905043. [Google Scholar] [CrossRef] [PubMed]
- Home—ProMED. ProMED-Mail. Available online: https://promedmail.org/ (accessed on 8 June 2022).
| Animal model | Advantages | Disadvantages |
|---|---|---|
| Mice | Small, easy to handle, cost-effective; reagents available; can conduct studies involving large number of animals; can use wild-type strains of YFV | Need to use immunocompromised mice; generally, viremia is not high enough to conduct mosquito experiments; disease outcome not similar to humans |
| Hamster | Viscerotropic disease outcome typical of YFV; can use immune-intact animals; generates high enough viremia for mosquito experiments | Need to use hamster-adapted strains; reagents are limited |
| Non-human primates | Disease outcome similar to humans; intact immunity; best representative model for vaccine and therapeutic studies; generally required in the past prior to clinical trials | Expensive; require specialized facilities, training; cannot use large groups due to ethical restrictions |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shinde, D.P.; Plante, J.A.; Plante, K.S.; Weaver, S.C. Yellow Fever: Roles of Animal Models and Arthropod Vector Studies in Understanding Epidemic Emergence. Microorganisms 2022, 10, 1578. https://doi.org/10.3390/microorganisms10081578
Shinde DP, Plante JA, Plante KS, Weaver SC. Yellow Fever: Roles of Animal Models and Arthropod Vector Studies in Understanding Epidemic Emergence. Microorganisms. 2022; 10(8):1578. https://doi.org/10.3390/microorganisms10081578
Chicago/Turabian StyleShinde, Divya P., Jessica A. Plante, Kenneth S. Plante, and Scott C. Weaver. 2022. "Yellow Fever: Roles of Animal Models and Arthropod Vector Studies in Understanding Epidemic Emergence" Microorganisms 10, no. 8: 1578. https://doi.org/10.3390/microorganisms10081578
APA StyleShinde, D. P., Plante, J. A., Plante, K. S., & Weaver, S. C. (2022). Yellow Fever: Roles of Animal Models and Arthropod Vector Studies in Understanding Epidemic Emergence. Microorganisms, 10(8), 1578. https://doi.org/10.3390/microorganisms10081578