
Sorption of Cellulases in Biofilm Enhances Cellulose Degradation by Bacillus subtilis


Abstract
1. Introduction
2. Materials and Methods
2.1. Culture Media
2.2. Bacterial Strains and Construction of Biofilm Knockout Mutants
2.3. Estimation of Submerged Biofilm Formation on Cellulose
2.4. Measurement of Cellulase in Biofilm and Supernatant
2.5. Pretreatment of Cellulose and Filter Paper for Efficient Degradation
2.6. Degradation of Regenerated Amorphous Cellulose by Biofilm and Enzyme Complexes
2.7. Visualization of Biofilm on Cellulose
2.8. Engineering Bacteria to Produce a Secretory Cellulase
2.9. Filter Paper Degradation by Engineered Bacteria
2.10. Statistical Analysis
3. Results
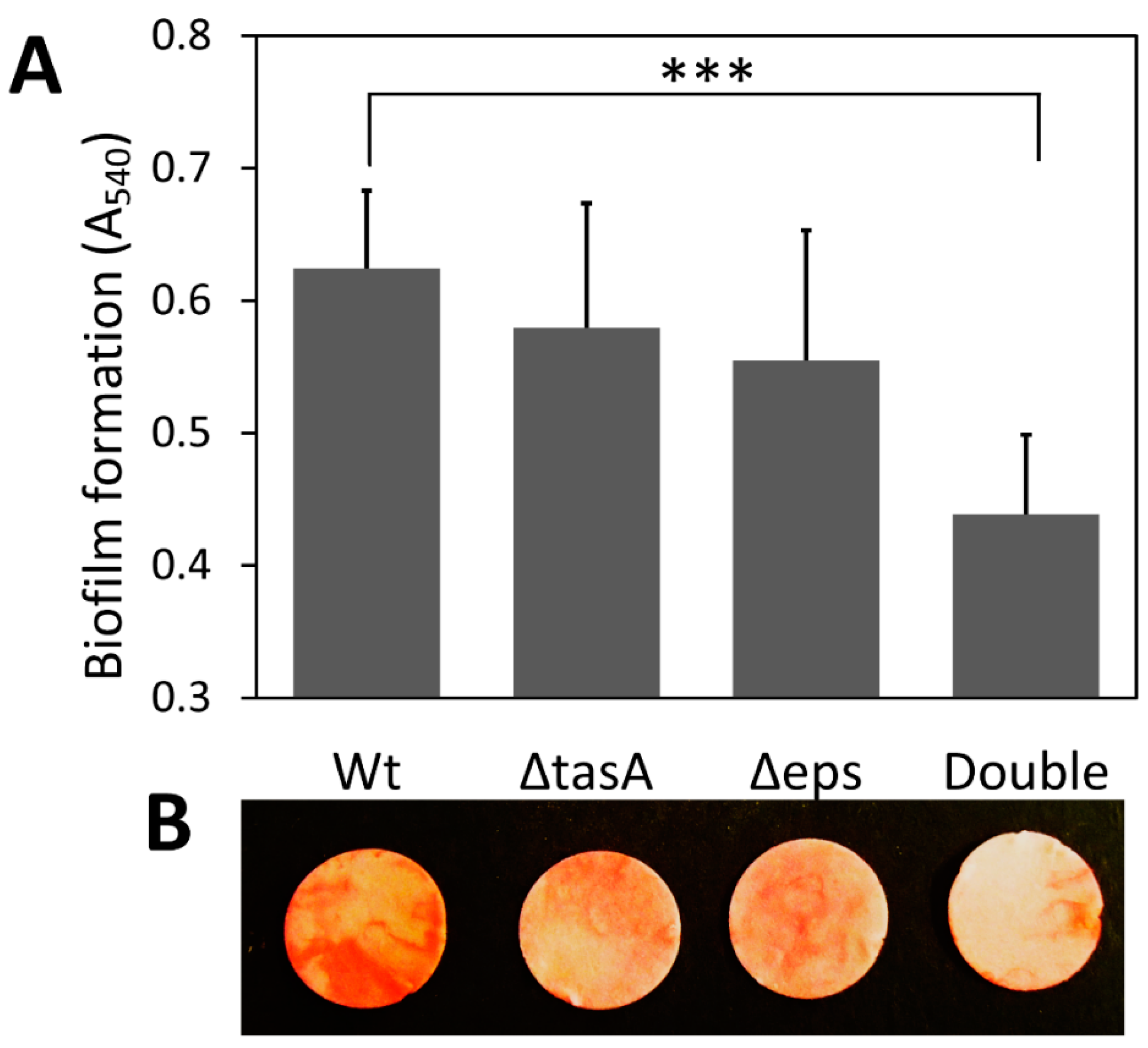
3.1. Differential Biofilm Formation on Cellulose Discs
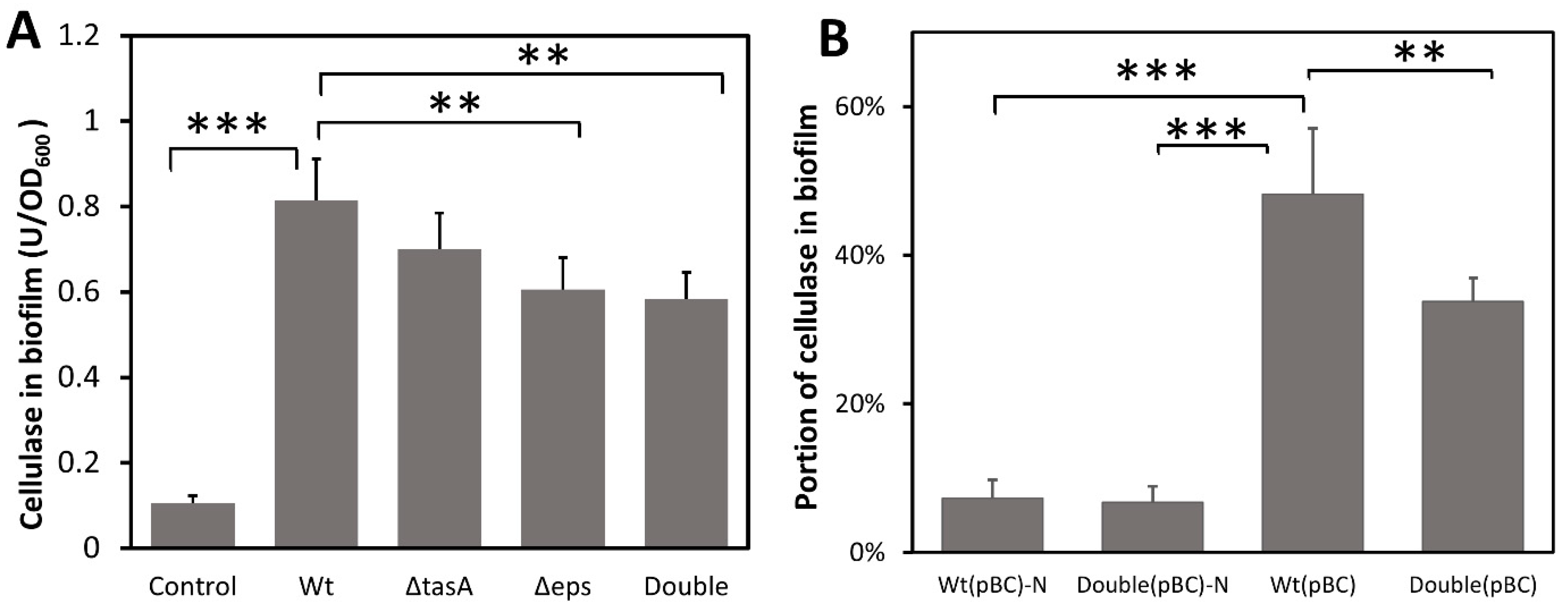
3.2. Biofilm Is Able to Capture and Retain Cellulase
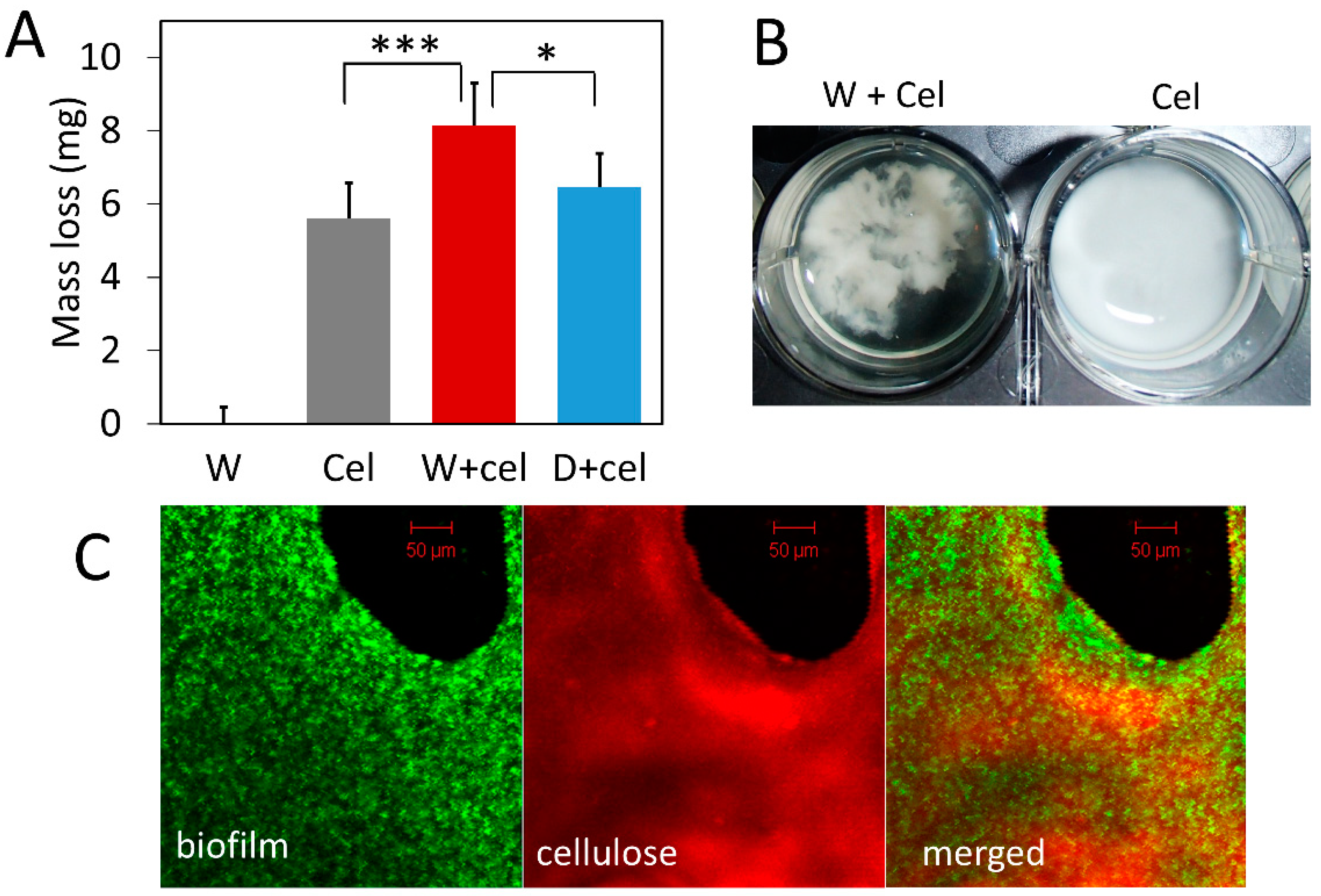
3.3. Biofilm Concentrates Exogenous Cellulase on Cellulose and Enhances Its Degradation
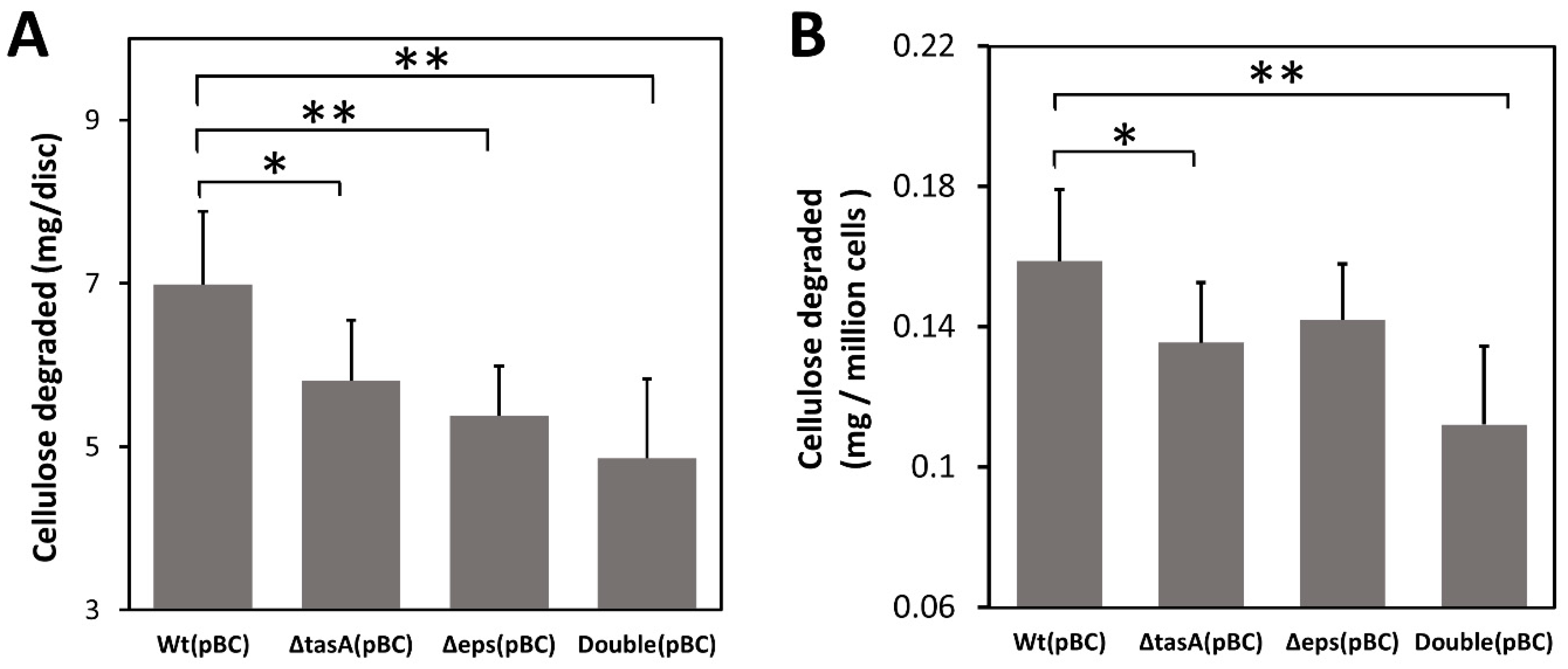
3.4. Biofilm Concentrates Endogenous Cellulase and Enhances Cellulose Degradation
4. Discussion
4.1. Potential Roles of Biofilm in Cellulose Degradation
4.2. Biofilm Resolves the Dilemma of Bacteria Producing Free Cellulases in the Environment
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flemming, H.-C.; Wingender, J. The Biofilm Matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Sutherland, I.W. The Biofilm Matrix—An Immobilized but Dynamic Microbial Environment. Trends Microbiol. 2001, 9, 222–227. [Google Scholar] [CrossRef]
- Davey, M.E.; Toole, G.A.O. Microbial Biofilms: From Ecology to Molecular Genetics. Microbiol. Mol. Biol. Rev. 2000, 64, 847–867. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An Emergent Form of Bacterial Life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Berlanga, M.; Guerrero, R. Living Together in Biofilms: The Microbial Cell Factory and Its Biotechnological Implications. Microb. Cell Fact. 2016, 15, 165. [Google Scholar] [CrossRef] [PubMed]
- Weimer, P.J.; Hatfield, R.D.; Buxton, D.R. Inhibition of Ruminal Cellulose Fermentation by Extracts of the Perennial Legume Cicer Milkvetch (Astragalus cicer). Appl. Envir. Microbiol. 1993, 59, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.W.; Lee, S.H.; Elkins, J.G.; Morrell-Falvey, J.L. Spatial and Temporal Dynamics of Cellulose Degradation and Biofilm Formation by Caldicellulosiruptor Obsidiansis and Clostridium Thermocellum. AMB Express 2011, 1, 30. [Google Scholar] [CrossRef] [PubMed]
- Young, J.M.; Leschine, S.B.; Reguera, G. Reversible Control of Biofilm Formation by Cellulomonas Spp. in Response to Nitrogen Availability. Environ. Microbiol. 2012, 14, 594–604. [Google Scholar] [CrossRef] [PubMed]
- Dumitrache, A.; Wolfaardt, G.; Allen, G.; Liss, S.N.; Lynd, L.R. Form and Function of Clostridium Thermocellum Biofilms. Appl. Environ. Microbiol. 2013, 79, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Dumitrache, A.; Wolfaardt, G.M.; Allen, D.G.; Liss, S.N.; Lynd, L.R. Tracking the Cellulolytic Activity of Clostridium Thermocellum Biofilms. Biotechnol. Biofuels 2013, 6, 175. [Google Scholar] [CrossRef] [PubMed]
- Lynd, L.R.; Weimer, P.J.; van Zyl, W.H.; Pretorius, I.S. Microbial Cellulose Utilization: Fundamentals and Biotechnology. Microbiol. Mol. Biol. Rev. 2002, 66, 506–577. [Google Scholar] [CrossRef] [PubMed]
- Bogino, P.C.; Oliva, M.D.L.M.; Sorroche, F.G.; Giordano, W. The Role of Bacterial Biofilms and Surface Components in Plant-Bacterial Associations. Int. J. Mol. Sci. 2013, 14, 15838–15859. [Google Scholar] [CrossRef] [PubMed]
- Brethauer, S.; Shahab, R.L.; Studer, M.H. Impacts of Biofilms on the Conversion of Cellulose. Appl. Microbiol. Biotechnol. 2020, 104, 5201–5212. [Google Scholar] [CrossRef]
- Wang, Z.W.; Chen, S. Potential of Biofilm-Based Biofuel Production. Appl Microbiol. Biotechnol 2009, 83, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Earl, A.M.; Losick, R.; Kolter, R. Ecology and Genomics of Bacillus Subtilis. Trends Microbiol. 2008, 16, 269–275. [Google Scholar] [CrossRef]
- Alcaraz, L.D.; Moreno-Hagelsieb, G.; Eguiarte, L.E.; Souza, V.; Herrera-Estrella, L.; Olmedo, G. Understanding the Evolutionary Relationships and Major Traits of Bacillus through Comparative Genomics. BMC Genom. 2010, 11, 332. [Google Scholar] [CrossRef]
- Hamon, M.A.; Lazazzera, B.A. The Sporulation Transcription Factor Spo0A Is Required for Biofilm Development in Bacillus Subtilis. Mol. Microbiol. 2001, 42, 1199–1209. [Google Scholar] [CrossRef]
- Bridier, A.; Le Coq, D.; Dubois-Brissonnet, F.; Thomas, V.; Aymerich, S.; Briandet, R. The Spatial Architecture of Bacillus Subtilis Biofilms Deciphered Using a Surface-Associated Model and in Situ Imaging. PLoS ONE 2011, 6, e16177. [Google Scholar] [CrossRef] [PubMed]
- Vlamakis, H.; Chai, Y.; Beauregard, P.; Losick, R.; Kolter, R. Sticking Together: Building a Biofilm the Bacillus Subtilis Way. Nat. Rev. Microbiol. 2013, 11, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Branda, S.S.; González-Pastor, J.E.; Ben-Yehuda, S.; Losick, R.; Kolter, R. Fruiting Body Formation by Bacillus Subtilis. Proc. Natl. Acad. Sci. USA 2001, 98, 11621–11626. [Google Scholar] [CrossRef]
- Branda, S.S.; Chu, F.; Kearns, D.B.; Losick, R.; Kolter, R. A Major Protein Component of the Bacillus Subtilis Biofilm Matrix. Mol. Microbiol. 2006, 59, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Kesel, S.; Von Bronk, B.; Falcón García, C.; Götz, A.; Lieleg, O.; Opitz, M. Matrix Composition Determines the Dimensions of: Bacillus Subtilis NCIB 3610 Biofilm Colonies Grown on LB Agar. RSC Adv. 2017, 7, 31886–31898. [Google Scholar] [CrossRef]
- Peng, N.; Cai, P.; Mortimer, M.; Wu, Y.; Gao, C.; Huang, Q. The Exopolysaccharide–EDNA Interaction Modulates 3D Architecture of Bacillus Subtilis Biofilm. BMC Microbiol. 2020, 20, 115. [Google Scholar] [CrossRef] [PubMed]
- Kearns, D.B.; Chu, F.; Branda, S.S.; Kolter, R.; Losick, R. A Master Regulator for Biofilm Formation by Bacillus Subtilis. Mol. Microbiol. 2005, 55, 739–749. [Google Scholar] [CrossRef]
- Branda, S.S.; Gonzalez-Pastor, J.E.; Dervyn, E.; Ehrlich, S.D.; Losick, R.; Kolter, R. Genes Involved in Formation of Structured Multicellular Communities by Bacillus Subtilis. J. Bacteriol. 2004, 186, 3970–3979. [Google Scholar] [CrossRef]
- Terra, R.; Stanley-Wall, N.R.; Cao, G.; Lazazzera, B.A. Identification of Bacillus Subtilis SipW as a Bifunctional Signal Peptidase That Controls Surface-Adhered Biofilm Formation. J. Bacteriol. 2012, 194, 2781–2790. [Google Scholar] [CrossRef]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia Coli K-12 In-frame, Single-gene Knockout Mutants: The Keio Collection. Mol. Syst. Biol. 2006, 2, 2006–2008. [Google Scholar] [CrossRef]
- Chai, Y.; Chu, F.; Kolter, R.; Losick, R. Bistability and Biofilm Formation in Bacillus Subtilis. Mol. Microbiol. 2008, 67, 254–263. [Google Scholar] [CrossRef]
- Wach, A. PCR-Synthesis of Marker Cassettes with Long Flanking Homology Regions for Gene Disruptions in S. Cerevisiae. Yeast 1996, 12, 259–265. [Google Scholar] [CrossRef]
- Yan, X.; Yu, H.-J.; Hong, Q.; Li, S.-P. Cre/Lox System and PCR-Based Genome Engineering in Bacillus Subtilis. Appl. Environ. Microbiol. 2008, 74, 5556–5562. [Google Scholar] [CrossRef]
- Anagnostopoulos, C.; Spizizen, J. Requirement for Transformation in Bacillus Subtilis. J. Bacteriol. 1961, 81, 741–746. [Google Scholar] [CrossRef] [PubMed]
- van den Burg, B.; Enequist, H.G.; van der Haar, M.E.; Eijsink, V.G.; Stulp, B.K.; Venema, G. A Highly Thermostable Neutral Protease from Bacillus Caldolyticus: Cloning and Expression of the Gene in Bacillus Subtilis and Characterization of the Gene Product. J. Bacteriol. 1991, 173, 4107–4115. [Google Scholar] [CrossRef] [PubMed]
- Overkamp, W.; Beilharz, K.; Detert Oude Weme, R.; Solopova, A.; Karsens, H.; Kovács, Á.T.; Kok, J.; Kuipers, O.P.; Veening, J.-W. Benchmarking Various Green Fluorescent Protein Variants in Bacillus Subtilis, Streptococcus Pneumoniae, and Lactococcus Lactis for Live Cell Imaging. Appl. Environ. Microbiol. 2013, 79, 6481–6490. [Google Scholar] [CrossRef] [PubMed]
- Hochstim, C.J.; Choi, J.Y.; Lowe, D.; Masood, R.; Rice, D.H. Biofilm Detection with Hematoxylin-Eosin Staining. Arch. Otolaryngol. Head. Neck Surg. 2010, 136, 453–456. [Google Scholar] [CrossRef]
- Tóth, L.; Csomor, P.; Sziklai, I.; Karosi, T. Biofilm Detection in Chronic Rhinosinusitis by Combined Application of Hematoxylin-Eosin and Gram Staining. Eur. Arch. Oto-Rhino-Laryngol. 2011, 268, 1455–1462. [Google Scholar] [CrossRef]
- Hulme, E.C.; Trevethick, M.A. Ligand Binding Assays at Equilibrium: Validation and Interpretation. Br. J. Pharmacol. 2010, 161, 1219–1237. [Google Scholar] [CrossRef]
- Jarmoskaite, I.; Alsadhan, I.; Vaidyanathan, P.P.; Herschlag, D. How to Measure and Evaluate Binding Affinities. Elife 2020, 9, e57264. [Google Scholar] [CrossRef]
- Xiao, Z.; Storms, R.; Tsang, A. Microplate-Based Carboxymethylcellulose Assay for Endoglucanase Activity. Anal. Biochem. 2005, 342, 176–178. [Google Scholar] [CrossRef]
- Zhang, Y.-H.P.; Cui, J.; Lynd, L.R.; Kuang, L.R. A Transition from Cellulose Swelling to Cellulose Dissolution by O-Phosphoric Acid: Evidence from Enzymatic Hydrolysis and Supramolecular Structure. Biomacromolecules 2006, 7, 644–648. [Google Scholar] [CrossRef]
- Zhang, X.Z.; Sathitsuksanoh, N.; Zhu, Z.; Percival Zhang, Y.H. One-Step Production of Lactate from Cellulose as the Sole Carbon Source without Any Other Organic Nutrient by Recombinant Cellulolytic Bacillus Subtilis. Metab. Eng. 2011, 13, 364–372. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Zhang, X.Z.; Zhang, Y.H.P. Simple, Fast and High-Efficiency Transformation System for Directed Evolution of Cellulase in Bacillus Subtilis. Microb. Biotechnol. 2011, 4, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-Z.; Cui, Z.-L.; Hong, Q.; Li, S.-P. High-Level Expression and Secretion of Methyl Parathion Hydrolase in Bacillus Subtilis WB800. Appl. Environ. Microbiol. 2005, 71, 4101–4103. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, C.; Govindarajan, S.; Minshull, J. Codon Bias and Heterologous Protein Expression. Trends Biotechnol. 2004, 22, 346–353. [Google Scholar] [CrossRef]
- Salis, H.M. The Ribosome Binding Site Calculator. Available online: https://salis.psu.edu/software/ (accessed on 24 September 2014).
- Salis, H.M.; Mirsky, E.A.; Voigt, C.A. Automated Design of Synthetic Ribosome Binding Sites to Control Protein Expression. Nat. Biotechnol. 2009, 27, 946–950. [Google Scholar] [CrossRef] [PubMed]
- Espah Borujeni, A.; Channarasappa, A.S.; Salis, H.M. Translation Rate Is Controlled by Coupled Trade-Offs between Site Accessibility, Selective RNA Unfolding and Sliding at Upstream Standby Sites. Nucleic Acids Res. 2014, 42, 2646–2659. [Google Scholar] [CrossRef]
- Tailliez, P.; Girard, H.; Millet, J.; Beguin, P. Enhanced Cellulose Fermentation by an Asporogenous and Ethanol-Tolerant Mutant of Clostridium Thermocellum. Appl. Environ. Microbiol. 1989, 55, 207–211. [Google Scholar] [CrossRef]
- Updegraff, D.M. Semimicro Determination of Cellulose Inbiological Materials. Anal. Biochem. 1969, 32, 420–424. [Google Scholar] [CrossRef]
- Dumitrache, A.; Klingeman, D.M.; Natzke, J.; Rodriguez, M.; Giannone, R.J.; Hettich, R.L.; Davison, B.H.; Brown, S.D. Specialized Activities and Expression Differences for Clostridium Thermocellum Biofilm and Planktonic Cells. Sci. Rep. 2017, 7, 43583. [Google Scholar] [CrossRef]
- Zhao, S.; Liu, Q.; Wang, J.X.; Liao, X.Z.; Guo, H.; Li, C.X.; Zhang, F.F.; Liao, L.S.; Luo, X.M.; Feng, J.X. Differential Transcriptomic Profiling of Filamentous Fungus during Solid-State and Submerged Fermentation and Identification of an Essential Regulatory Gene PoxMBF1 That Directly Regulated Cellulase and Xylanase Gene Expression. Biotechnol. Biofuels 2019, 12, 103. [Google Scholar] [CrossRef]
- Romani, A.M.; Fund, K.; Artigas, J.; Schwartz, T.; Sabater, S.; Obst, U. Relevance of Polymeric Matrix Enzymes during Biofilm Formation. Microb. Ecol. 2008, 56, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Frolund, B.; Griebe, T.; Nielsen, P.H. Enzymatic Activity in the Activated-Sludge Floc Matrix. Appl. Microbiol. Biotechnol. 1995, 43, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.H.; Lynd, L.R. Toward an Aggregated Understanding of Enzymatic Hydrolysis of Cellulose: Noncomplexed Cellulase Systems. Biotechnol. Bioeng. 2004, 88, 797–824. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-Z.; Sathitsuksanoh, N.; Lynd, L.R.; Percival Zhang, Y.H.; You, C.; Zhang, Y.-H.P. Enhanced Microbial Utilization of Recalcitrant Cellulose by an Ex Vivo Cellulosome-Microbe Complex. Appl. Environ. Microbiol. 2012, 78, 1437–1444. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, Y.-H.P.; Lynd, L.R. Enzyme-Microbe Synergy during Cellulose Hydrolysis by Clostridium Thermocellum. Proc. Natl. Acad. Sci. USA 2006, 103, 16165–16169. [Google Scholar] [CrossRef]
- Van Dyk, J.S.; Pletschke, B.I. A Review of Lignocellulose Bioconversion Using Enzymatic Hydrolysis and Synergistic Cooperation between Enzymes—Factors Affecting Enzymes, Conversion and Synergy. Biotechnol. Adv. 2012, 30, 1458–1480. [Google Scholar] [CrossRef]
- Marvasi, M.; Visscher, P.T.; Casillas Martinez, L. Exopolymeric Substances (EPS) from Bacillus Subtilis: Polymers and Genes Encoding Their Synthesis. FEMS Microbiol. Lett. 2010, 313, 207–212. [Google Scholar] [CrossRef]
- Shoham, Y.; Lamed, R.; Bayer, E.A. The Cellulosome Concept as an Efficient Microbial Strategy for the Degradation of Insoluble Polysaccharides. Trends Microbiol. 1999, 7, 275–281. [Google Scholar] [CrossRef]
- Deng, Y.-J.; Wang, S.Y. Synergistic Growth in Bacteria Depends on Substrate Complexity. J. Microbiol. 2016, 54, 23–30. [Google Scholar] [CrossRef]
- Jiang, Y.; Liu, Y.; Zhang, X.; Gao, H.; Mou, L.; Wu, M.; Zhang, W.; Xin, F.; Jiang, M. Biofilm Application in the Microbial Biochemicals Production Process. Biotechnol. Adv. 2021, 48, 107724. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain (Plasmid) | Relevant Genotype or Property | Source |
|---|---|---|
| Wt | Wild type 168, trpC2 | BGSC * |
| Δeps | Δeps::zeoR; eps operon deleted from Wt168 | This study |
| ΔtasA | ΔtasA::ermR; tasA deleted from Wt168 | BGSC |
| Double | Δeps::zeoR, ΔtasA::ermR; Δeps ΔtasA double knockout from Wt 168 | This study |
| E. coli DH5α (p7Z6) | ampR, zeoR, lox71-zeo-lox66 | [30] |
| B. subtilis 1012M15 (pGDV1) | High-copy-number plasmid, camR; carrying multiple cloning site mp18 | [32] |
| E. coli DH5α (pDR111-GFP(Sp)) | ampR, specR, carrying GFP; integration vector for B. subtilis | [33] |
| Wt(pBC) | Wt168 carrying cellulase over-expression plasmid pBC; camR | This study |
| Δeps(pBC) | Δeps carrying pBC | This study |
| ΔtasA(pBC) | ΔtasA carrying pBC | This study |
| Double(pBC) | Δeps ΔtasA carrying pBC | This study |
| Wt_GFP | Wt168 with GFP incorporated into its genome by plasmid pDR111-GFP(Sp) | This Study |
| Primer Name | Primer Sequence 5′-----3′ |
|---|---|
| eps-up-F | ACGCCATTGTCCGACAGG |
| eps-up-R | AATCTCTAGAGGATCCCCGGGTACCGAGCTC CTCATTCATGTATTCATAGCCTTCAGC |
| zeo-F | GAGCTCGGTACCCGGGGATCCT |
| zeo-R | GCTTGCATGCCTGCAGGTCGAC |
| eps-dw-F | GTAGAATCGTCGACCTGCAGGCATGCAAGCATCCAGCACGCCTCA AAGAAG |
| eps-dw-R | TATCTTGAATGGTATGAAGCGGAAT |
| epsCF-F | GACTGAGCAGGTTCCAATC |
| epsCF-R | CCAAGTTGAGCGAGTGTTTC |
| tasA-F | CATCAGGTACGCTTGATTTATCTG |
| tasA-R | GTTTTATCATCCTTGAATTGGATTTCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, Y.; Wang, S.Y. Sorption of Cellulases in Biofilm Enhances Cellulose Degradation by Bacillus subtilis. Microorganisms 2022, 10, 1505. https://doi.org/10.3390/microorganisms10081505
Deng Y, Wang SY. Sorption of Cellulases in Biofilm Enhances Cellulose Degradation by Bacillus subtilis. Microorganisms. 2022; 10(8):1505. https://doi.org/10.3390/microorganisms10081505
Chicago/Turabian StyleDeng, Yijie, and Shiao Y. Wang. 2022. "Sorption of Cellulases in Biofilm Enhances Cellulose Degradation by Bacillus subtilis" Microorganisms 10, no. 8: 1505. https://doi.org/10.3390/microorganisms10081505
APA StyleDeng, Y., & Wang, S. Y. (2022). Sorption of Cellulases in Biofilm Enhances Cellulose Degradation by Bacillus subtilis. Microorganisms, 10(8), 1505. https://doi.org/10.3390/microorganisms10081505