
Fusarium Graminearum Virus-1 Strain FgV1-SD4 Infection Eliminates Mycotoxin Deoxynivalenol Synthesis by Fusarium graminearum in FHB


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and Fungal Materials
2.2. Confirmation of FgV1
2.3. Comparison of Pathogenicity between Fg-4-1 and Fg-4-2
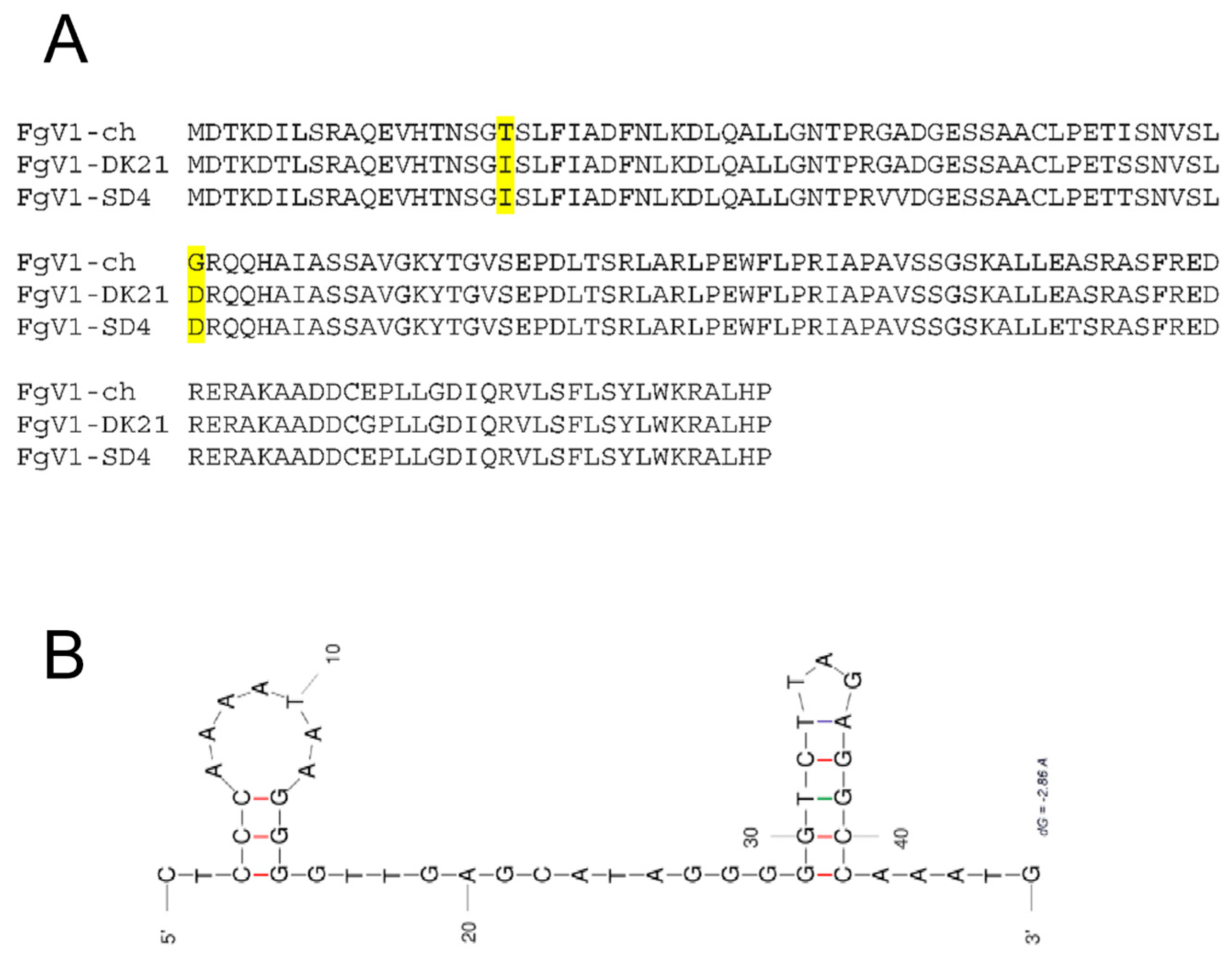
2.4. Analysis of Viral Genomic Sequences
3. Results
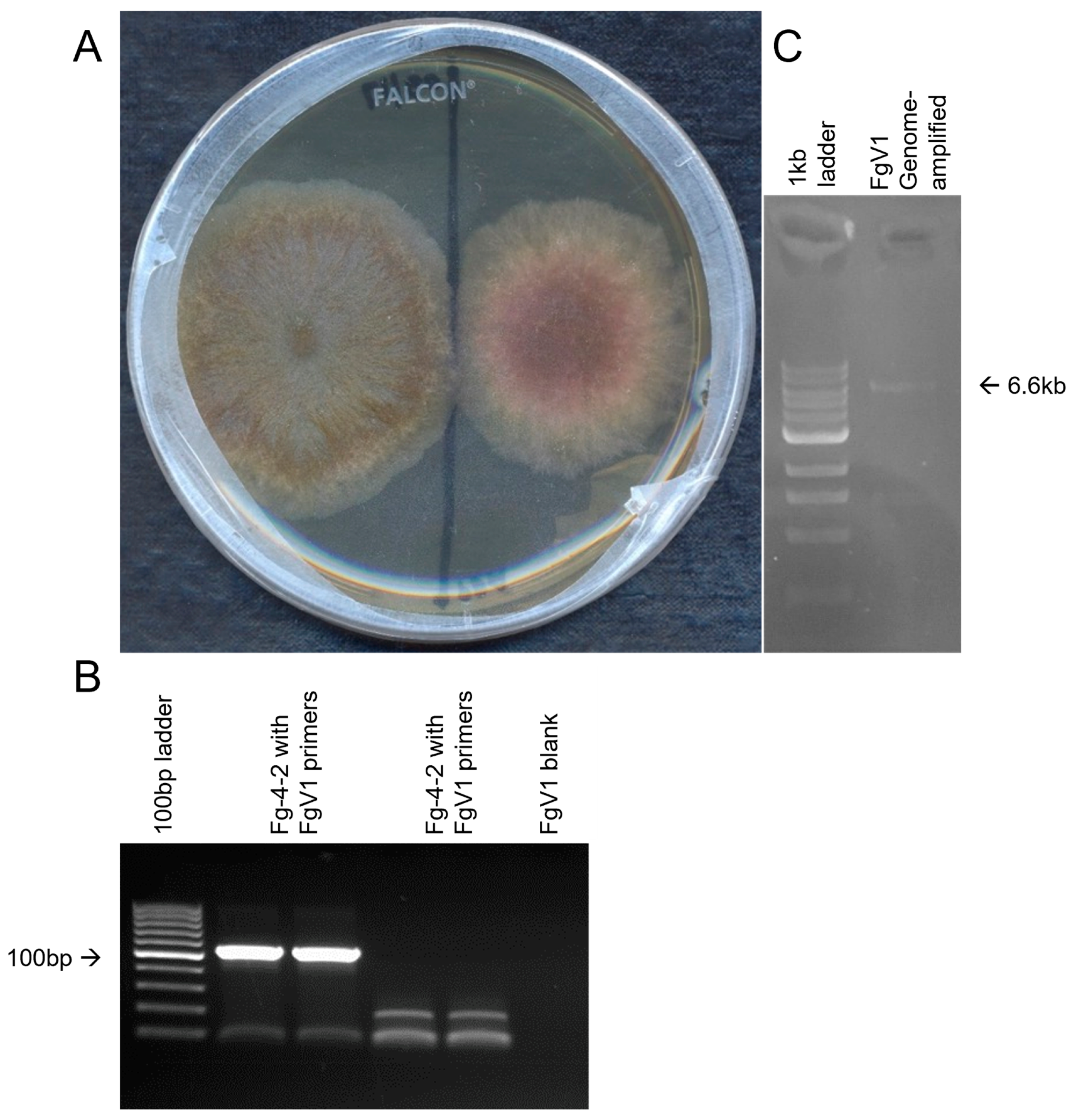
3.1. The Discovery of FgV1-SD4
3.2. Detection and Confirmation of FgV1-SD4
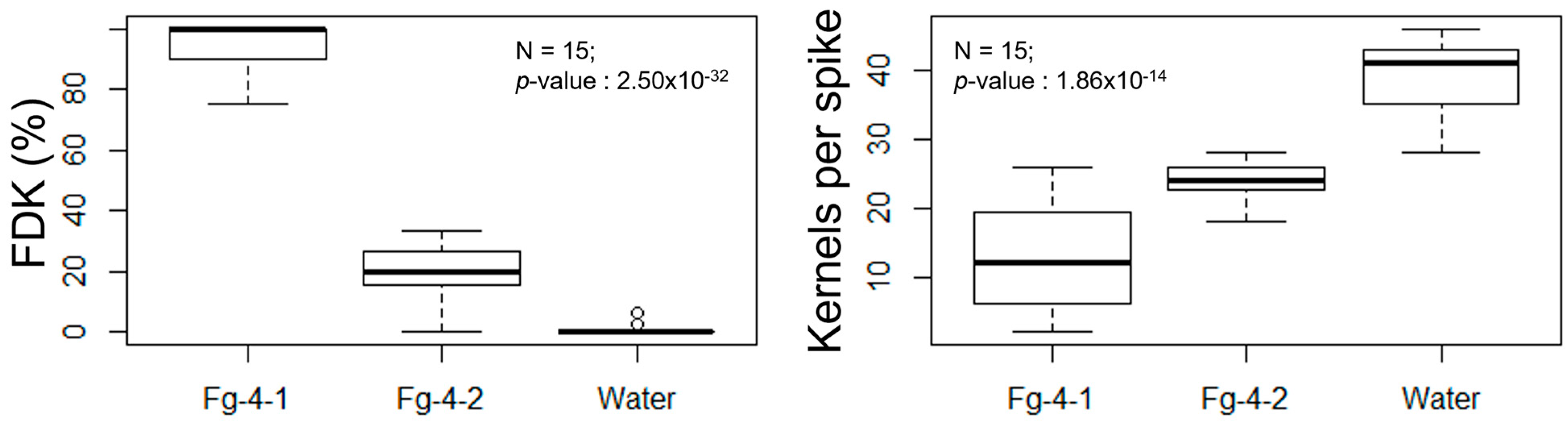
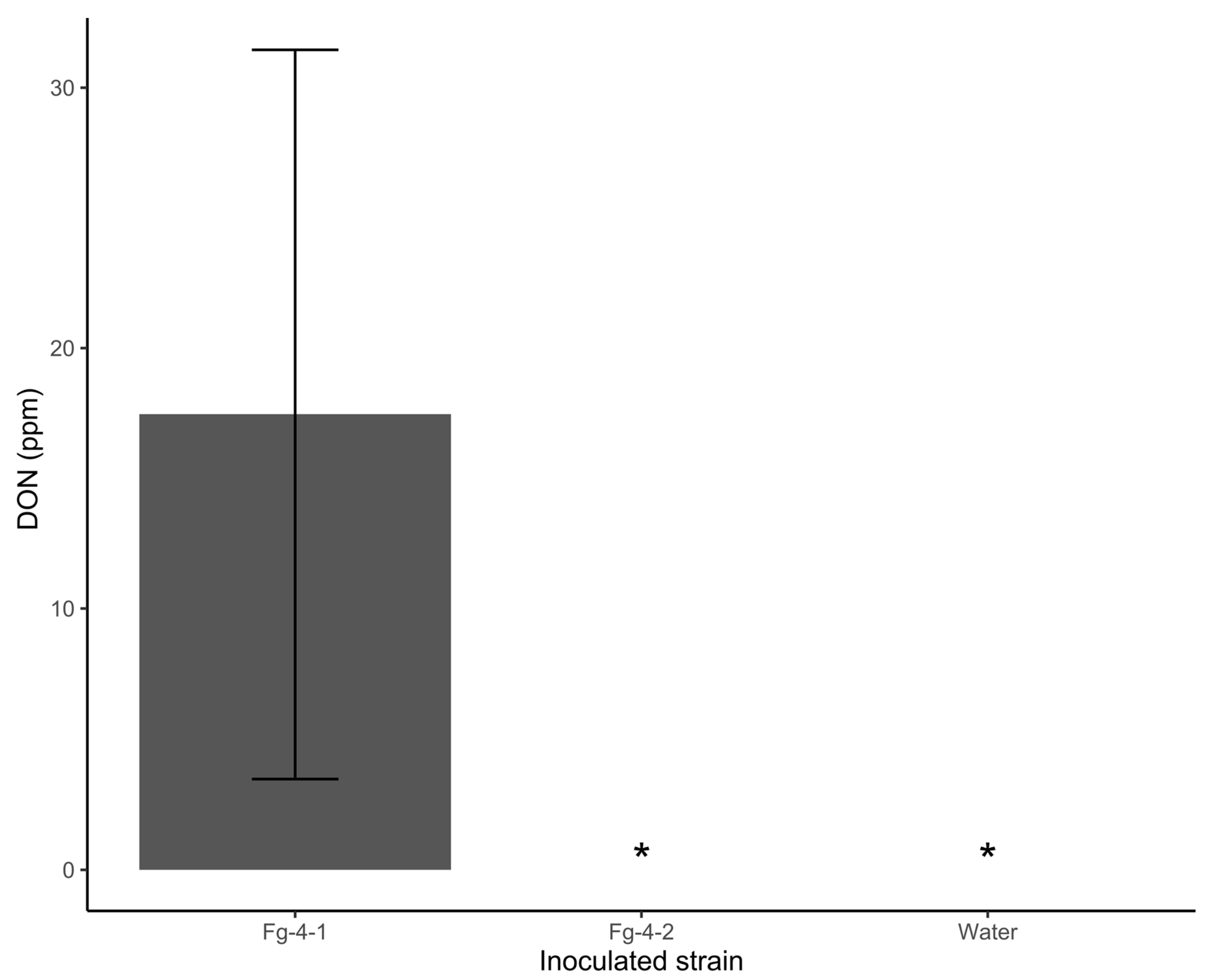
3.3. Pathogenicity in FHB
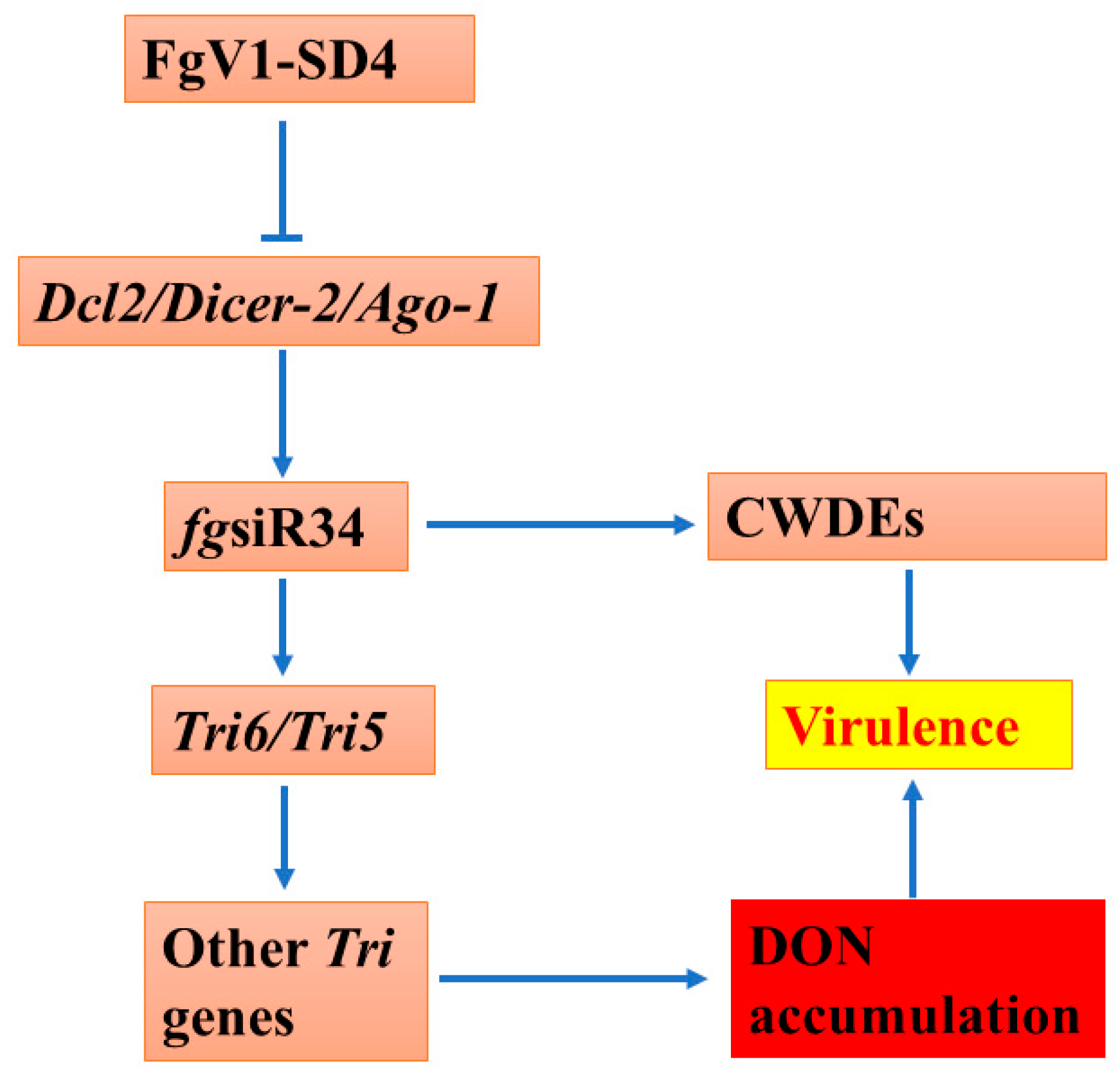
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cook, S.; Chung, B.Y.W.; Bass, D.; Moureau, G.; Tang, S.; McAlister, E.; Culverwell, C.L.; Glücksman, E.; Wang, H.; Brown, T.D.K. Novel virus discovery and genome reconstruction from field RNA samples reveals highly divergent viruses in dipteran hosts. PLoS ONE 2013, 8, e80720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzano, S.L.; Nelson, B.D.; Ajayi-Oyetunde, O.; Bradley, C.A.; Hughes, T.J.; Hartman, G.L.; Eastburn, D.M.; Domier, L.L. Identification of diverse mycoviruses through metatranscriptomics characterization of the viromes of five major fungal plant pathogens. J. Virol. 2016, 90, 6846–6863. [Google Scholar] [CrossRef] [Green Version]
- Rosario, K.; Breitbart, M. Exploring the viral world through metagenomics. Curr. Opin. Virol. 2011, 1, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Choi, G.H.; Dawe, A.L.; Churbanov, A.; Smith, M.L.; Milgroom, M.G.; Nuss, D.L. Molecular characterization of vegetative incompatibility genes that restrict hypovirus transmission in the chestnut blight fungus Cryphonectria parasitica. Genetics 2012, 190, 113–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Li, B.; Fu, Y.; Xie, J.; Cheng, J.; Ghabrial, S.A.; Li, G.; Yi, X.; Jiang, D. Extracellular transmission of a DNA mycovirus and its use as a natural fungicide. Proc. Natl. Acad. Sci. USA 2013, 110, 1452–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, C.; Feng, J.; Wang, Z.; Pedersen, C.; Wang, X.; Saleem, H.; Domier, L.; Marzano, S.Y.L. Identification of the viral determinant of hypovirulence and host range in Sclerotiniaceae of a genomovirus reconstructed from the plant metagenome. J. Virol. 2021, 95, e00264-21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Hisano, S.; Tani, A.; Kondo, H.; Kanematsu, S.; Suzuki, N. A capsidless ssRNA virus hosted by an unrelated dsRNA virus. Nat. Microbiol. 2016, 1, 15001. [Google Scholar] [CrossRef] [PubMed]
- Rochon, D.A.; Kakani, K.; Robbins, M.; Reade, R. Molecular aspects of plant virus transmission by olpidium and plasmodiophorid vectors. Annu. Rev. Phytopathol. 2004, 42, 211–241. [Google Scholar] [CrossRef]
- Yu, J.; Kwon, S.J.; Lee, K.M.; Son, M.; Kim, K.H. Complete nucleotide sequence of double-stranded RNA viruses from Fusarium graminearum strain DK3. Arch. Virol. 2009, 154, 1855. [Google Scholar] [CrossRef]
- Kwon, S.J.; Lim, W.S.; Park, S.H.; Park, M.R.; Kim, K.H. Molecular characterization of a dsRNA mycovirus, Fusarium graminearum virus-DK21, which is phylogenetically related to hypoviruses but has a genome organization and gene expression strategy resembling those of plant potex-like viruses. Mol. Cells 2007, 23, 304. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.M.; Jeon, J.J.; Yea, S.J.; Kim, Y.H.; Yun, S.H.; Lee, Y.W.; Kim, K.H. Double-stranded RNA mycovirus from Fusarium graminearum. Appl. Environ. Microbiol. 2002, 68, 2529–2534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stack, R.W.; Leonard, K.; Bushnell, W. History of Fusarium head blight with emphasis on North America. In Fusarium Head Blight Wheat Barley; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2003; pp. 1–34. [Google Scholar]
- Zhang, L.; Chen, X.; Bhattacharjee, P.; Shi, Y.; Guo, L.; Wang, S. Molecular Characterization of a Novel Strain of Fusarium graminearum Virus 1 Infecting Fusarium graminearum. Viruses 2020, 12, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cogoni, C.; Macino, G. Gene silencing in Neurospora crassa requires a protein homologous to RNA-dependent RNA polymerase. Nature 1999, 399, 166. [Google Scholar] [CrossRef]
- Zhang, X.; Segers, G.C.; Sun, Q.; Deng, F.; Nuss, D.L. Characterization of hypovirus-derived small RNAs generated in the chestnut blight fungus by an inducible DCL-2-dependent pathway. J. Virol. 2008, 82, 2613–2619. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Gao, Q.; Huang, M.; Liu, Y.; Liu, Z.; Liu, X.; Ma, Z. Characterization of RNA silencing components in the plant pathogenic fungus Fusarium graminearum. Sci. Rep. 2015, 5, 12500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galla, A. Role of Fusarium graminearum sRNAs in Fusarium Head Blight Pathogenesis on Wheat; South Dakota State University: Brookings, SD, USA, 2014. [Google Scholar]
- Lee, K.-M.; Cho, W.K.; Yu, J.; Son, M.; Choi, H.; Min, K.; Lee, Y.-W.; Kim, K.-H. A comparison of transcriptional patterns and mycological phenotypes following infection of Fusarium graminearum by four mycoviruses. PLoS ONE 2014, 9, e100989. [Google Scholar] [CrossRef]
- Andika, I.B.; Jamal, A.; Kondo, H.; Suzuki, N. SAGA complex mediates the transcriptional up-regulation of antiviral RNA silencing. Proc. Natl. Acad. Sci. USA 2017, 114, E3499–E3506. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Lee, K.M.; Cho, W.K.; Park, J.Y.; Kim, K.H. Differential contribution of RNA interference components in response to distinct Fusarium graminearum virus infections. J. Virol. 2018, 92, e01756-17. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.Y.; Naupane, A.; Feng, J.H.; Pedersen, C.; Marzano, S.L. Direct metatranscriptomics survey of the sunflower microbiome and virome. Viruses 2021, 13, 1867. [Google Scholar] [CrossRef]
- Li, G.; Yen, Y. Jasmonate and ethylene signaling pathway may mediate Fusarium head blight resistance in wheat. Crop Sci. 2008, 48, 1888–1896. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markham, N.R.; Zuker, M. DINAMelt web server for nucleic acid melting prediction. Nucleic Acids Res. 2005, 33 (Suppl. S2), W577–W581. [Google Scholar] [CrossRef] [PubMed]
- Andika, I.B.; Maruyama, K.; Sun, L.; Kondo, H.; Tamada, T.; Suzuki, N. Differential contributions of plant Dicer-like proteins to antiviral defenses against potato virus X in leaves and roots. Plant J. 2015, 8, 781–793. [Google Scholar] [CrossRef] [PubMed]
- Campo, S.; Gilbert, K.B.; Carrington, J.C. Small RNA-based antiviral defense in the phytopathogenic fungus Colletotrichum higginsianum. PLoS Pathog. 2016, 12, e1005640. [Google Scholar] [CrossRef] [Green Version]
- Segers, G.C.; Zhang, X.; Deng, F.; Sun, Q.; Nuss, D.L. Evidence that RNA silencing functions as an antiviral defense mechanism in fungi. Proc. Natl. Acad. Sci. USA 2007, 104, 12902–12906. [Google Scholar] [CrossRef] [Green Version]
- Segers, G.C.; Van Wezel, R.; Zhang, X.; Hong, Y.; Nuss, D.L. Hypovirus papain-like protease p29 suppresses RNA silencing in the natural fungal host and in a heterologous plant system. Eukaryot. Cell 2006, 5, 896–904. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Choi, G.H.; Nuss, D.L. A single Argonaute gene is required for induction of RNA silencing antiviral defense and promotes viral RNA recombination. Proc. Natl. Acad. Sci. USA 2009, 106, 17927–17932. [Google Scholar] [CrossRef] [Green Version]
- Dahal, S. Role of Silencing RNA fgsiR34 in Fusarium graminearum’s Pathogenicity to Wheat; South Dakota State University: Brookings, SD, USA, 2016. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence | Purpose |
|---|---|---|
| FgV1-F | GTTGCGTTGGAGGTTGACAC | RT-PCR |
| FgV1-R | CCAAAAACCACACGTCGTCC | RT-PCR, cDNA synthesis |
| FgV1-F2 | GGGGTATACTCTGATTATTTGAATTT | Whole viral genome RT-PCR amplification, Sanger sequencing |
| FgV1-R2 | CATTTGGCCTCTAGACCCCCTATGCT | Whole viral genome RT-PCR amplification, Reverse transcription, Sequencing |
| FgMV-F | ACCATATCCCTTTTGGGGCTG | RT-PCR |
| FgMV-R | GTGCTCTTCCGATCTCCGTG | RT-PCR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paudel, B.; Pedersen, C.; Yen, Y.; Marzano, S.-Y.L. Fusarium Graminearum Virus-1 Strain FgV1-SD4 Infection Eliminates Mycotoxin Deoxynivalenol Synthesis by Fusarium graminearum in FHB. Microorganisms 2022, 10, 1484. https://doi.org/10.3390/microorganisms10081484
Paudel B, Pedersen C, Yen Y, Marzano S-YL. Fusarium Graminearum Virus-1 Strain FgV1-SD4 Infection Eliminates Mycotoxin Deoxynivalenol Synthesis by Fusarium graminearum in FHB. Microorganisms. 2022; 10(8):1484. https://doi.org/10.3390/microorganisms10081484
Chicago/Turabian StylePaudel, Bimal, Connor Pedersen, Yang Yen, and Shin-Yi Lee Marzano. 2022. "Fusarium Graminearum Virus-1 Strain FgV1-SD4 Infection Eliminates Mycotoxin Deoxynivalenol Synthesis by Fusarium graminearum in FHB" Microorganisms 10, no. 8: 1484. https://doi.org/10.3390/microorganisms10081484
APA StylePaudel, B., Pedersen, C., Yen, Y., & Marzano, S.-Y. L. (2022). Fusarium Graminearum Virus-1 Strain FgV1-SD4 Infection Eliminates Mycotoxin Deoxynivalenol Synthesis by Fusarium graminearum in FHB. Microorganisms, 10(8), 1484. https://doi.org/10.3390/microorganisms10081484