
Variations in Marine Bacterial and Archaeal Communities during an Ulva prolifera Green Tide in Coastal Qingdao Areas

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. DNA Extraction and Illumina High-Throughput Sequencing
2.3. Sequence Processing
2.4. Statistical Analysis
3. Results
3.1. Variations in Environmental Factors during the Green Tide
3.2. Variations in Bacterial and Archaeal Richness and Diversity during the Green Tide
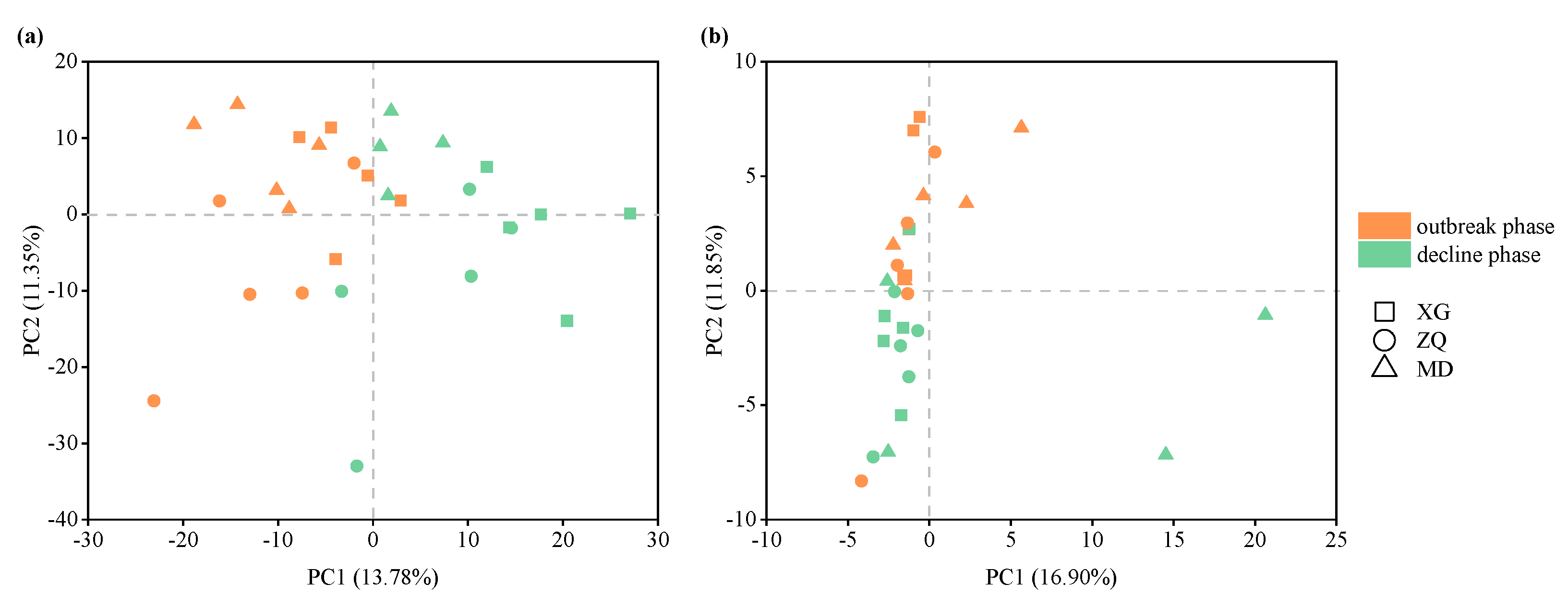
3.3. Variations in Bacterial and Archaeal Community Structure during the Green Tide
3.4. Correlation Analysis of Environmental Factors with Bacterial and Archaeal Communities
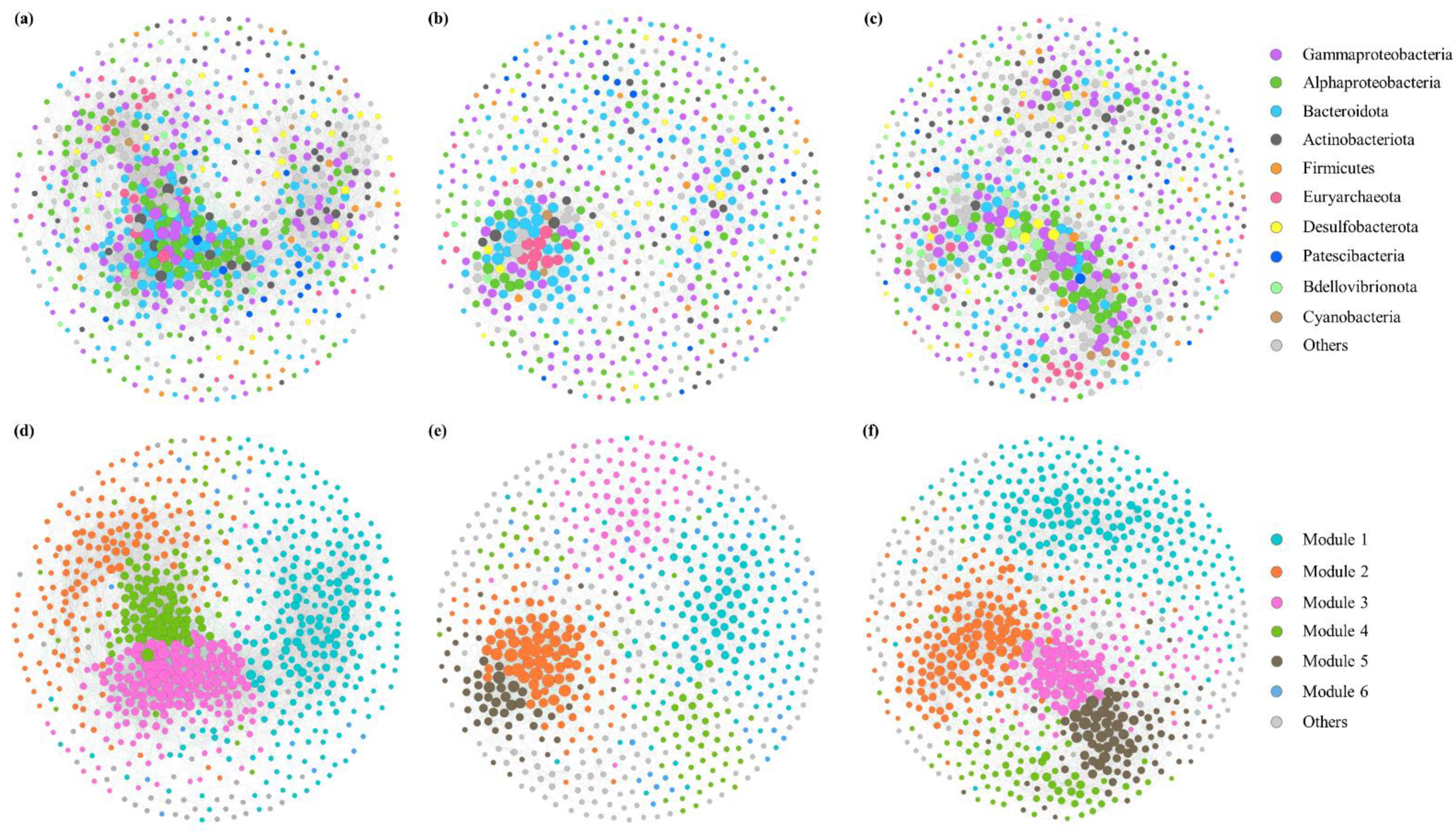
3.5. Microbial Co-Occurrence Network Analysis during the Green Tide
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, J.; Jiang, P.; Liu, Z.; Wei, W.; Lin, H.; Li, F.; Wang, J.; Qin, S. The Yellow Sea green tides were dominated by one species, Ulva (Enteromorpha) prolifera, from 2007 to 2011. Chin. Sci. Bull. 2013, 58, 2298–2302. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Wu, Y.; Fang, Z.; Cui, X.; Liang, J.; Song, X. Spatiotemporal patterns and morphological characteristics of Ulva prolifera distribution in the Yellow Sea, China in 2016–2018. Remote Sens. 2019, 11, 445. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; He, P.; Li, H.; Li, G.; Liu, J.; Jiao, F.; Zhang, J.; Huo, Y.; Shi, X.; Su, R.; et al. Ulva prolifera green-tide outbreaks and their environmental impact in the Yellow Sea, China. Natl. Sci. Rev. 2019, 6, 825–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Wang, Z. A review of the green tides in the Yellow Sea, China. Mar. Environ. Res. 2016, 119, 189–196. [Google Scholar] [CrossRef]
- Song, X.; Shi, Y.; Liu, A.; Xing, H.; Jiang, H.; Wang, W.; Zhang, L. The impact of green tide on the phytoplankton community in Yellow Sea. Appl. Mech. Mater. 2013, 260–261, 1130–1137. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, X. Release and microbial degradation of dissolved organic matter (DOM) from the macroalgae Ulva prolifera. Mar. Pollut. Bull. 2017, 125, 192–198. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, G.; Sun, X.; Zhang, F.; Zhang, X. Temporal variability in zooplankton community in the western Yellow Sea and its possible links to green tides. PeerJ 2019, 7, e6641. [Google Scholar] [CrossRef]
- Qu, T.; Zhao, X.; Hao, Y.; Zhong, Y.; Guan, C.; Hou, C.; Tang, X.; Wang, Y. Ecological effects of Ulva prolifera green tide on bacterial community structure in Qingdao offshore environment. Chemosphere 2020, 244, 125477. [Google Scholar] [CrossRef]
- O’Boyle, S.; McDermott, G.; Silke, J.; Cusack, C. Potential impact of an exceptional bloom of Karenia mikimotoi on dissolved oxygen levels in waters off western Ireland. Harmful Algae 2016, 53, 77–85. [Google Scholar] [CrossRef]
- Achá, D.; Guédron, S.; Amouroux, D.; Point, D.; Lazzaro, X.; Fernandez, P.E.; Sarret, G. Algal bloom exacerbates hydrogen sulfide and methylmercury contamination in the emblematic high-altitude Lake Titicaca. Geosciences 2018, 8, 438. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Yu, R.C.; Zhou, M.J. Effects of the decomposing green macroalgae Ulva (Enteromorpha) prolifera on the growth of four red-tide species. Harmful Algae 2012, 16, 12–19. [Google Scholar] [CrossRef]
- Ducklow, H.W.; Purdie, D.A.; Williams, P.J.L.; Davies, J.M. Bacterioplankton: A sink for carbon in a coastal marine plankton community. Science 1986, 232, 865–867. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, H.G.; Gocke, K.; Koppe, R.; Begler, C. Bacterial growth and primary production along a north-south transect of the Atlantic Ocean. Nature 2002, 416, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Seymour, J.R.; Stocker, M.R. Resource patch formation and exploitation throughout the marine microbial food web. Am. Nat. 2009, 173, E15–E29. [Google Scholar] [CrossRef] [Green Version]
- Paerl, H.W.; Dyble, J.; Moisander, P.H.; Noble, R.T.; Piehler, M.F.; Pinckney, J.L.; Steppe, T.F.; Twomey, L.; Valdes, L.M. Microbial indicators of aquatic ecosystem change: Current applications to eutrophication studies. FEMS Microbiol. Ecol. 2003, 46, 233–246. [Google Scholar] [CrossRef]
- Su, X.; Steinman, A.D.; Tang, X.; Xue, Q.; Zhao, Y.; Xie, L. Response of bacterial communities to cyanobacterial harmful algal blooms in Lake Taihu, China. Harmful Algae 2017, 68, 168–177. [Google Scholar] [CrossRef]
- Zhang, H.; Hou, F.; Xie, W.; Wang, K.; Zhou, X.; Zhang, D.; Zhu, X. Interaction and assembly processes of abundant and rare microbial communities during a diatom bloom process. Mar. Microb. Ecol. 2020, 22, 1707–1719. [Google Scholar] [CrossRef]
- Klindworth, A.; Mann, A.J.; Huang, S.; Wichels, A.; Quast, C.; Waldmann, J.; Teeling, H.; Glöckner, F.O. Diversity and activity of marine bacterioplankton during a diatom bloom in the North Sea assessed by total RNA and pyrotag sequencing. Mar. Genom. 2014, 18, 185–192. [Google Scholar] [CrossRef]
- Zhou, J.; Richlen, M.L.; Sehein, T.R.; Kulis, D.M.; Anderson, D.M.; Cai, Z. Microbial community structure and associations during a marine dinoflagellate bloom. Front. Microbiol. 2018, 9, 1201. [Google Scholar] [CrossRef]
- Geng, H.; Yu, R.; Zhang, Q.; Yan, T.; Kong, F.; Zhou, M. Tracing the settlement region of massive floating green algae in the Yellow Sea. J. Oceanol. Limnol. 2019, 37, 1555–1565. [Google Scholar] [CrossRef]
- Zhang, X.; Song, Y.; Liu, D.; Keesing, J.K.; Gong, J. Macroalgal blooms favor heterotrophic diazotrophic bacteria in nitrogen-rich and phosphorus-limited coastal surface waters in the Yellow Sea. Estuar. Coast. Shelf Sci. 2015, 163, 75–81. [Google Scholar] [CrossRef]
- Lin, G.; Sun, F.; Wang, C.; Zhang, L.; Zhang, X. Assessment of the effect of Enteromorpha prolifera on bacterial community structures in aquaculture environment. PLoS ONE 2017, 12, e0179792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Li, H.; Zhang, Z.; He, C.; Shi, Q.; Jiao, N.; Zhang, Y. DOC dynamics and bacterial community succession during long-term degradation of Ulva prolifera and their implications for the legacy effect of green tides on refractory DOC pool in seawater. Water Res. 2020, 185, 116268. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Wang, C.; Chen, H.; Zheng, Z. Metagenomic analysis of the effect of Enteromorpha prolifera on microbial community and function in aquaculture environment. Curr. Microbiol. 2020, 77, 816–825. [Google Scholar] [CrossRef]
- Liang, J.; Liu, J.; Zhan, Y.; Zhou, S.; Xue, C.X.; Sun, C.; Lin, Y.; Luo, C.; Wang, X.; Zhang, X.H. Succession of marine bacteria in response to Ulva prolifera-derived dissolved organic matter. Environ. Int. 2021, 155, 106687. [Google Scholar] [CrossRef]
- Needham, D.M.; Fuhrman, J.A. Pronounced daily succession of phytoplankton, archaea and bacteria following a spring bloom. Nat. Microbiol. 2016, 1, 16005. [Google Scholar] [CrossRef]
- Lupatini, M.; Suleiman, A.K.A.; Jacques, R.J.S.; Antoniolli, Z.I.; de Siqueira Ferreira, A.; Kuramae, E.E.; Roesch, L.F.W. Network topology reveals high connectance levels and few key microbial genera within soils. Front. Environ. Sci. 2014, 2, 10. [Google Scholar] [CrossRef] [Green Version]
- Fuhrman, J.A.; Cram, J.A.; Needham, D.M. Marine microbial community dynamics and their ecological interpretation. Nat. Rev. Microbiol. 2015, 13, 133–146. [Google Scholar] [CrossRef]
- Dai, T.; Zhang, Y.; Ning, D.; Su, Z.; Tang, Y.; Huang, B.; Mu, Q.; Wen, D. Dynamics of sediment microbial functional capacity and community interaction networks in an urbanized coastal estuary. Front. Microbiol. 2018, 9, 2731. [Google Scholar] [CrossRef]
- Chaffron, S.; Rehrauer, H.; Pernthaler, J.; von Mering, C. A global network of coexisting microbes from environmental and whole-genome sequence data. Genome Res. 2010, 20, 947–959. [Google Scholar] [CrossRef] [Green Version]
- Konopka, A.; Lindemann, S.; Fredrickson, J. Dynamics in microbial communities: Unraveling mechanisms to identify principles. ISME J. 2015, 9, 1488–1495. [Google Scholar] [CrossRef] [PubMed]
- Hou, F.; Zhang, H.; Xie, W.; Zhou, X.; Zhu, X.; Zhang, D. Co-occurrence patterns and assembly processes of microeukaryotic communities in an early-spring diatom bloom. Sci. Total Environ. 2020, 711, 134624. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Lao, Y.M.; Song, J.T.; Jin, H.; Zhu, J.M.; Cai, Z.H. Temporal heterogeneity of microbial communities and metabolic activities during a natural algal bloom. Water Res. 2020, 183, 116020. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, H.; Zhang, Y.; Si, D.; Chen, Q. Evolution of microbial community along with increasing solid concentration during high-solids anaerobic digestion of sewage sludge. Bioresour. Technol. 2016, 216, 87–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, N.; Tan, G.; Wang, H.; Gai, X. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An open source software for exploring and manipulating networks. In Proceedings of the International AAAI Conference on Weblogs and Social Media, San Jose, CA, USA, 17–20 May 2009. [Google Scholar]
- Zhang, Y.H.; Yao, P.; Sun, C.; Li, S.Z.; Shi, X.C.; Zhang, X.H.; Liu, J.W. Vertical diversity and association pattern of total, abundant and rare microbial communities in deep-sea sediments. Mol. Ecol. 2021, 30, 2800–2816. [Google Scholar] [CrossRef]
- Simon, H.; Lipsewers, Y.A.; Giebel, H.A.; Wiltshire, K.H.; Simon, M. Temperature effects on aggregation during a spring diatom bloom. Limnol. Oceanogr. 2014, 59, 2089–2100. [Google Scholar] [CrossRef]
- Skerratt, J.H.; Bowman, J.P.; Hallegraeff, G.; James, S.; Nichols, P.D. Algicidal bacteria associated with blooms of a toxic dinoflagellate in a temperate Australian estuary. Mar. Ecol. Prog. Ser. 2002, 244, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Brinkhoff, T.; Giebel, H.A.; Simon, M. Diversity, ecology, and genomics of the Roseobacter clade: A short overview. Arch. Microbiol. 2008, 189, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Lenk, S.; Moraru, C.; Hahnke, S.; Arnds, J.; Richter, M.; Kube, M.; Reinhardt, R.; Brinkhoff, T.; Harder, J.; Amann, R.; et al. Roseobacter clade bacteria are abundant in coastal sediments and encode a novel combination of sulfur oxidation genes. ISME J. 2012, 6, 2178–2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voget, S.; Wemheuer, B.; Brinkhoff, T.; Vollmers, J.; Dietrich, S.; Giebel, H.A.; Beardsley, C.; Sardemann, C.; Bakenhus, I.; Billerbeck, S.; et al. Adaptation of an abundant Roseobacter RCA organism to pelagic systems revealed by genomic and transcriptomic analyses. ISME J. 2015, 9, 371–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.X.; Zhang, H.; Chen, X.H.; Xie, Z.X.; Zhang, Y.; Zhang, S.F.; Lin, L.; Chen, F.; Wang, D.Z. Metaproteomics reveals major microbial players and their metabolic activities during the blooming period of a marine dinoflagellate Prorocentrum donghaiense. Environ. Microbiol. 2018, 20, 632–644. [Google Scholar] [CrossRef]
- Tada, Y.; Taniguchi, A.; Nagao, I.; Miki, T.; Uematsu, M.; Tsuda, A.; Hamasaki, K. Differing growth responses of major phylogenetic groups of marine bacteria to natural phytoplankton blooms in the western north Pacific Ocean. Appl. Environ. Microbiol. 2011, 77, 4055–4065. [Google Scholar] [CrossRef] [Green Version]
- Imai, I.; Ishida, Y.; Sakaguchi, K.; Hata, Y. Algicidal marine bacteria isolated from northern Hiroshima Bay, Japan. Fish. Sci. 1995, 61, 628–636. [Google Scholar] [CrossRef] [Green Version]
- Umetsu, S.; Kanda, M.; Imai, I.; Sakai, R.; Fujita, M.J. Questiomycins, algicidal compounds produced by the marine bacterium Alteromonas sp. D and their production cue. Molecules 2019, 24, 4522. [Google Scholar] [CrossRef] [Green Version]
- Mayali, X.; Azamn, F. Algicidal bacteria in the sea and their impact on algal blooms. J. Eukaryot. Microbiol. 2004, 51, 139–144. [Google Scholar] [CrossRef]
- Orsi, W.D.; Smith, J.M.; Wilcox, H.M.; Swalwell, J.E.; Carini, P.; Worden, A.Z.; Santoro, A.E. Ecophysiology of uncultivated marine euryarchaea is linked to particulate organic matter. ISME J. 2015, 9, 1747–1763. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.L.; Xie, W.; Martin-Cuadrado, A.B.; Rodriguez-Valera, F. Marine Group II archaea, potentially important players in the global ocean carbon cycle. Front. Microbiol. 2015, 6, 1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Zhang, C.L.; Yang, C.; Chen, S.; Cao, Z.; Zhang, Z.; Tian, J. Marine Group II dominates planktonic archaea in water column of the northeastern South China Sea. Front. Microbiol. 2017, 8, 1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, W.; Luo, H.; Murugapiran, S.K.; Dodsworth, J.A.; Chen, S.; Sun, Y.; Hedlund, B.P.; Wang, P.; Fang, H.; Deng, M.; et al. Localized high abundance of Marine Group II archaea in the subtropical Pearl River Estuary: Implications for their niche adaptation. Environ. Microbiol. 2018, 20, 734–754. [Google Scholar] [CrossRef]
- Christman, H.; Campbell, E.; Meeks, J. Global transcription profiles of the nitrogen stress response resulting in heterocyst or hormogonium development in Nostoc punctiforme. J. Bacteriol. 2011, 193, 6874–6886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morono, Y.; Terada, N.; Nishizawa, M.; Ito, M.; Hillion, F.; Takahata, N.; Sano, Y.; Inagaki, F. Carbon and nitrogen assimilation in deep subseafloor microbial cells. Proc. Natl. Acad. Sci. USA 2011, 108, 18295–18300. [Google Scholar] [CrossRef] [Green Version]
- Stahl, D.A.; de la Torre, J.R. Physiology and diversity of ammonia-oxidizing archaea. Annu. Rev. Microbiol. 2012, 66, 83–101. [Google Scholar] [CrossRef]
- Offre, P.; Spang, A.; Schleper, C. Archaea in biogeochemical cycles. Annu. Rev. Microbiol. 2013, 67, 437–457. [Google Scholar] [CrossRef] [Green Version]
- Bothe, H.; Jost, G.; Schloter, M.; Ward, B.B.; Witzel, K.P. Molecular analysis of ammonia oxidation and denitrification in natural environments. FEMS Microbiol. Rev. 2000, 24, 673–690. [Google Scholar] [CrossRef]
- Niu, Y.; Shen, H.; Chen, J.; Xie, P.; Yang, X.; Tao, M.; Ma, Z.; Qi, M. Phytoplankton community succession shaping bacterioplankton community composition in Lake Taihu, China. Water Res. 2011, 45, 4169–4182. [Google Scholar] [CrossRef]
- Xing, P.; Li, H.; Liu, Q.; Zhang, J. Composition of the archaeal community involved in methane production during the decomposition of Microcystis blooms in the laboratory. Can. J. Microbiol. 2012, 58, 1153–1158. [Google Scholar] [CrossRef]
- Bergen, B.; Endres, S.; Engel, A.; Zark, M.; Dittmar, T.; Sommer, U.; Jürgens, K. Acidification and warming affect prominent bacteria in two seasonal phytoplankton bloom mesocosms. Environ. Microbiol. 2016, 18, 4579–4595. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.; Wichels, A.; Teeling, H.; Chafee, M.; Scharfe, M.; Gerdts, G. Annual dynamics of North Sea bacterioplankton: Seasonal variability superimposes short-term variation. FEMS Microbiol. Ecol. 2015, 91, fiv099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele, J.A.; Countway, P.D.; Xia, L.; Vigil, P.D.; Beman, J.M.; Kim, D.Y.; Chow, C.T.; Sachdeva, R.; Jones, A.C.; Schwalbach, M.S.; et al. Marine bacterial, archaeal and protistan association networks reveal ecological linkages. ISME J. 2011, 5, 1414–1425. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, F.; Zhang, C.; Wang, M.; Kan, J. Estuarine gradients dictate spatiotemporal variations of microbiome networks in the Chesapeake Bay. Environ. Microbiome 2021, 16, 22. [Google Scholar] [CrossRef]
- Herren, C.M.; McMahon, K.D. Keystone taxa predict compositional change in microbial communities. Environ. Microbiol. 2018, 20, 2207–2217. [Google Scholar] [CrossRef] [Green Version]
- Mayali, X.; Franks, P.J.S.; Azam, F. Cultivation and ecosystem role of a marine Roseobacter clade-affiliated cluster bacterium. Appl. Environ. Microbiol. 2008, 74, 2595–2603. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Date | Sample Name | ||
|---|---|---|---|
| XG | ZQ | MD | |
| 13 June 2019 | XG0613 | ZQ0613 | MD0613 |
| 27 June 2019 | XG0627 | ZQ0627 | MD0627 |
| 11 July 2019 | XG0711 | ZQ0711 | MD0711 |
| 18 July 2019 | XG0718 | ZQ0718 | MD0718 |
| 22 July 2019 | XG0722 | ZQ0722 | MD0722 |
| 8 August 2019 | XG0806 | ZQ0806 | MD0806 |
| 16 August 2019 | XG0816 | ZQ0816 | / |
| 23 August 2019 | XG0823 | ZQ0823 | MD0823 |
| 29 August 2019 | XG0829 | ZQ0829 | MD0829 |
| 5 September 2019 | XG0905 | ZQ0905 | MD0905 |
| Temperature (°C) | Salinity | pH | DO (mg/L) | chla (μg/L) | NH4+ (μmol/L) | NO2− (μmol/L) | NO3− (μmol/L) | PO43− (μmol/L) | |
|---|---|---|---|---|---|---|---|---|---|
| XG0613 | 18.73 | 30.15 | 8.05 | 7.61 | 1.67 | 4.27 | 0.13 | 1.53 | 2.88 |
| XG0627 | 20.90 | 29.96 | 8.05 | 7.56 | 1.07 | 24.35 | 0.14 | 1.67 | 4.31 |
| XG0711 | 22.80 | 29.83 | 7.98 | 7.31 | 1.18 | 5.37 | 0.08 | 1.15 | 0.30 |
| XG0718 | 23.60 | 29.78 | 8.00 | 6.74 | / | 23.47 | 0.17 | 2.30 | 0.26 |
| XG0722 | 24.77 | 29.49 | 7.99 | 7.34 | 2.96 | 13.37 | 0.33 | 5.02 | 0.44 |
| XG0806 | 25.83 | 29.95 | 7.58 | 7.62 | 0.75 | 12.66 | 0.26 | 2.90 | 0.61 |
| XG0816 | 26.10 | 29.28 | 7.75 | 7.48 | 0.83 | 18.42 | 0.65 | 9.57 | 0.30 |
| XG0823 | 26.10 | 29.66 | 8.02 | 10.71 | 1.51 | 15.32 | 0.27 | 3.70 | 0.21 |
| XG0829 | 26.07 | 29.79 | 8.37 | 6.47 | / | 8.01 | 0.23 | 3.08 | 0.10 |
| XG0905 | 25.37 | 29.61 | 8.09 | 9.42 | 0.65 | 28.70 | 0.52 | 6.84 | 3.92 |
| ZQ0613 | 20.10 | 30.08 | 8.11 | 9.39 | 2.92 | 28.66 | 0.13 | 1.76 | 0.24 |
| ZQ0627 | 22.00 | 30.07 | 8.05 | 7.24 | 3.91 | 18.78 | 0.11 | 1.37 | 0.89 |
| ZQ0711 | 23.80 | 30.01 | 8.04 | 5.59 | 3.37 | 5.69 | 0.07 | 0.73 | / |
| ZQ0718 | 23.70 | 29.86 | 8.01 | 5.08 | / | 21.13 | 0.11 | 1.38 | / |
| ZQ0722 | 25.90 | 29.86 | 8.04 | 7.94 | 4.86 | 5.71 | 0.08 | 1.06 | / |
| ZQ0806 | 26.00 | 30.39 | 7.82 | 7.68 | 0.78 | 11.17 | 0.20 | 2.09 | 2.68 |
| ZQ0816 | 26.20 | 30.00 | 8.02 | 9.10 | 1.51 | 10.14 | 0.21 | 3.98 | 0.44 |
| ZQ0823 | 26.10 | 30.04 | 8.11 | 10.20 | 1.13 | 20.74 | 0.40 | 5.03 | / |
| ZQ0829 | 26.50 | 30.17 | 8.25 | 12.17 | / | 16.01 | 0.16 | 1.87 | 0.27 |
| ZQ0905 | 25.20 | 29.88 | 8.10 | 12.86 | 0.93 | 21.07 | 0.27 | 3.56 | 1.83 |
| MD0613 | 18.60 | 30.22 | 8.11 | 8.74 | / | 3.25 | 0.04 | 0.59 | 0.44 |
| MD0627 | 20.80 | 30.04 | 8.13 | 9.07 | / | 7.69 | 0.08 | 1.07 | 0.27 |
| MD0711 | 22.90 | 29.98 | 8.13 | 8.37 | / | 18.40 | 0.05 | 0.73 | 0.25 |
| MD0718 | 23.00 | 29.87 | 8.05 | 7.84 | / | 4.91 | 0.07 | 0.97 | 0.35 |
| MD0722 | 24.30 | 29.77 | 8.05 | 8.19 | / | 23.02 | 0.11 | 1.93 | 0.18 |
| MD0806 | 25.80 | 29.88 | 8.01 | 8.05 | / | 21.40 | 0.20 | 2.66 | 0.32 |
| MD0823 | 26.20 | 29.90 | 7.99 | 3.92 | / | 19.95 | 0.08 | 0.99 | 0.27 |
| MD0829 | 26.40 | 29.91 | 8.21 | 11.05 | / | 22.23 | 0.31 | 4.76 | 0.22 |
| MD0905 | 25.50 | 29.74 | 8.08 | 8.35 | / | 23.03 | 0.38 | 5.45 | 4.67 |
| Bacterial Community | Archaeal Community | |||||||
|---|---|---|---|---|---|---|---|---|
| OTUs | Chao1 | Shannon | Coverage | OTUs | Chao1 | Shannon | Coverage | |
| XG0613 | 643 | 750.36 | 4.44 | 0.9965 | 57 | 65.27 | 1.64 | 0.9995 |
| XG0627 | 588 | 705.50 | 3.80 | 0.9963 | 53 | 60.50 | 1.65 | 0.9996 |
| XG0711 | 659 | 824.28 | 3.43 | 0.9958 | 82 | 83.00 | 2.47 | 0.9999 |
| XG0718 | 768 | 871.21 | 4.61 | 0.9963 | 101 | 123.00 | 2.43 | 0.9996 |
| XG0722 | 735 | 851.32 | 4.55 | 0.9962 | 89 | 102.00 | 1.85 | 0.9995 |
| XG0806 | 671 | 825.01 | 4.28 | 0.9956 | 57 | 72.00 | 1.85 | 0.9994 |
| XG0816 | 750 | 856.65 | 4.62 | 0.9961 | 59 | 85.25 | 1.76 | 0.9994 |
| XG0823 | 756 | 854.70 | 4.31 | 0.9963 | 106 | 115.23 | 2.55 | 0.9994 |
| XG0829 | 787 | 890.15 | 4.87 | 0.9964 | 89 | 114.00 | 2.45 | 0.9991 |
| XG0905 | 674 | 765.88 | 4.69 | 0.9967 | 53 | 62.00 | 2.33 | 0.9996 |
| ZQ0613 | 642 | 738.04 | 3.84 | 0.6697 | 64 | 65.20 | 1.92 | 0.9999 |
| ZQ0627 | 719 | 784.32 | 4.63 | 0.9973 | 73 | 79.07 | 1.61 | 0.9995 |
| ZQ0711 | 625 | 748.04 | 4.04 | 0.9959 | 59 | 69.91 | 2.11 | 0.9994 |
| ZQ0718 | 813 | 932.72 | 4.76 | 0.9962 | 83 | 92.07 | 2.13 | 0.9994 |
| ZQ0722 | 811 | 903.24 | 4.79 | 0.9963 | 92 | 117.50 | 1.96 | 0.9993 |
| ZQ0806 | 803 | 935.37 | 4.68 | 0.9959 | 90 | 106.87 | 2.19 | 0.9991 |
| ZQ0816 | 712 | 789.11 | 4.55 | 0.9969 | 102 | 122.00 | 2.11 | 0.9994 |
| ZQ0823 | 650 | 797.66 | 3.99 | 0.9956 | 62 | 66.67 | 2.49 | 0.9997 |
| ZQ0829 | 837 | 949.26 | 4.85 | 0.9962 | 112 | 120.57 | 2.60 | 0.9994 |
| ZQ0905 | 762 | 915.25 | 4.41 | 0.9958 | 76 | 93.10 | 2.39 | 0.9993 |
| MD0613 | 632 | 753.15 | 3.89 | 0.9967 | 67 | 67.75 | 1.95 | 0.9999 |
| MD0627 | 687 | 781.61 | 4.45 | 0.9969 | 49 | 56.00 | 1.61 | 0.9997 |
| MD0711 | 646 | 769.10 | 3.34 | 0.9960 | 70 | 75.60 | 2.42 | 0.9997 |
| MD0718 | 806 | 916.57 | 4.52 | 0.9963 | 83 | 113.00 | 2.18 | 0.9991 |
| MD0722 | 750 | 830.53 | 4.23 | 0.9965 | 82 | 84.50 | 2.37 | 0.9998 |
| MD0806 | 792 | 945.38 | 4.37 | 0.9954 | 86 | 98.67 | 2.19 | 0.9993 |
| MD0823 | 395 | 539.73 | 3.43 | 0.9968 | 57 | 64.50 | 2.95 | 0.9998 |
| MD0829 | 438 | 631.80 | 2.31 | 0.9960 | 75 | 80.60 | 2.36 | 0.9997 |
| MD0905 | 617 | 812.63 | 3.83 | 0.9951 | 83 | 87.58 | 2.39 | 0.9996 |
| Phases | Bacterial Community | Archaeal Community | ||||||
|---|---|---|---|---|---|---|---|---|
| Chao1 | Shannon | Rhodobacterales | Flavobacteriales | Chao1 | Shannon | MGII | ||
| chla content | green tide | −0.212 | 0.018 | 0.326 | 0.638 ** | 0.209 | −0.250 | −0.165 |
| outbreak phase | 0.548 | 0.857 ** | −0.048 | 0.762 * | 0.619 | 0.000 | −0.476 | |
| decline phase | 0.048 | −0.476 | 0.690 | 0.333 | 0.667 | 0.476 | 0.143 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, G.; He, H.; Wang, H.; Liang, Y.; Guo, C.; Shao, H.; Jiang, Y.; Wang, M. Variations in Marine Bacterial and Archaeal Communities during an Ulva prolifera Green Tide in Coastal Qingdao Areas. Microorganisms 2022, 10, 1204. https://doi.org/10.3390/microorganisms10061204
Zhao G, He H, Wang H, Liang Y, Guo C, Shao H, Jiang Y, Wang M. Variations in Marine Bacterial and Archaeal Communities during an Ulva prolifera Green Tide in Coastal Qingdao Areas. Microorganisms. 2022; 10(6):1204. https://doi.org/10.3390/microorganisms10061204
Chicago/Turabian StyleZhao, Guihua, Hui He, Hualong Wang, Yantao Liang, Cui Guo, Hongbing Shao, Yong Jiang, and Min Wang. 2022. "Variations in Marine Bacterial and Archaeal Communities during an Ulva prolifera Green Tide in Coastal Qingdao Areas" Microorganisms 10, no. 6: 1204. https://doi.org/10.3390/microorganisms10061204
APA StyleZhao, G., He, H., Wang, H., Liang, Y., Guo, C., Shao, H., Jiang, Y., & Wang, M. (2022). Variations in Marine Bacterial and Archaeal Communities during an Ulva prolifera Green Tide in Coastal Qingdao Areas. Microorganisms, 10(6), 1204. https://doi.org/10.3390/microorganisms10061204