Abstract
Colibacillosis caused by pathogenic Escherichia coli (E. coli) is one of the most serious infectious diseases, causing an extensive burden on animal husbandry and the human healthcare system. Vaccination is one of the ideal ways to prevent E. coli infection. In this work, recombinant outer membrane protein A (rOmpA), outer membrane protein C (rOmpC) and BamA (rBamA) from E. coli O78 (CVCC CAU0768) were expressed in a prokaryotic expression system with the concentration of 1–2 mg/mL after purification. Considerable immune responses could be triggered in mice that were immunized with these recombinant proteins, high antibody titers, high total IgG level and various antibody isotypes were detected in antisera after booster immunizations. Moreover, mice immunized with several recombinant proteins in combination showed a higher survival rate with the challenge of homologous strain E. coli O78 and a more significant cross-protection effect against heterologous strain E. coli O157:H7 (CICC 21530) in vivo than those of immunized alone. The antisera from immunized mice showed high affinity to multiple strains of Escherichia, Shigella and Salmonella in vitro, indicating that recombinant outer membrane proteins from E. coli O78 had the potential to be developed into universal antigenic substances against not only E. coli but also a variety of Gram-negative bacteria. rOmpA was considered as the most immunogenic protein in this work and the combination of different proteins could further enhance the immune response of immunized mice, which provided the reference for the construction of novel antigens with higher efficiency.
1. Introduction
There are two main pathogenic E. coli clinical pathotypes, including intestinal pathogenic E. coli (IPEC) and extraintestinal pathogenic E. coli (ExPEC), in which IPEC is the primary cause of diarrhea- and intestinal-related diseases, while ExPEC is the major cause of extraintestinal infections including urinary tract infections, neonatal sepsis, meningitis, etc. [1,2,3,4]. IPEC strains such as enterotoxigenic E. coli (ETEC), enteroaggregative E. coli (EAEC), enterohemorrhagic E. coli (EHEC), etc., primarily give rise to intestinal-related diseases and almost do not cause extraintestinal infection in the healthy hosts except in the developing world [2]. EHEC can lead to diarrhea, hemorrhagic colitis and hemolytic uremic syndrome, of which serotype O157:H7 plays a major role [5,6]. ExPEC strains such as avian pathogenic E. coli (APEC), septicemic pathogenic E. coli (SPEC) and uropathogenic E. coli (UPEC), etc., have a unique ability to invade various organs or tissues outside the gastrointestinal tract of hosts, resulting in various life-threatening diseases [3]. Reports showed that about 80% of APEC infections are caused by serotypes O78, O1 and O2 [7], leading to great economic losses to the poultry industry worldwide [8,9,10]. Antibiotics have made a great contribution to the treatment of E. coli infection, but the consequences of which cannot be ignored [11]. The rates of antibiotic resistance in E. coli are very high in China, India and the Middle East [2]. In addition, serotyping of E. coli is complex, with 173 kinds of O antigen, 80 kinds of K antigen, and 56 kinds of H antigen, which can be found in many possible combinations in nature, eventually forming 50,000–100,000 or more serotypes [12]. Although there are more and more studies and reports about vaccines for E. coli based on serotype, there will be a tremendous amount of work to do.
The academic, technical and industrial advantages of subunit vaccines are: moving the gateway of epidemic prevention ahead, mastering the risk of infection under control, delaying the mutation of pathogens, extending the validity span of vaccines, preventing the epidemics effectively at optimal ratio of cost performance in terms of time, results, scale and other input in whole. Therefore, subunit vaccines have become important candidate vaccine systems for rapid response to the spread of emergent diseases and pandemics. Outer membrane proteins (OMPs) broadly exist in Gram-negative bacteria and belong to the transmembrane β-barrel family. OMPs are not only involved in the processes of protein secretion, adhesion, virulence and biofilm formation of bacteria but also potential drug and vaccine targets as surface-exposed molecules [13]. As a consequence, the OMPs subunit vaccine has become an important research target for the treatment of Gram-negative bacteria. It has been reported that rOmpA, rOmpC and rBamA from E. coli CVCC 1515 exhibited a high immunogenicity, a strong humoral immune response could be detected in serum from mice that were immunized with these OMPs and Freund’s adjuvant [14,15,16]. Pore et al. found that rOmpA from Shigella flexneri 2a could evoke protective immunity in mice, which involved the participation of both the humoral and cellular immune responses [17]. Li et al. indicated that rOmpF could not only induce chicken to generate a high level of antibody but also induce chicken to produce a strong cell-mediated immune response, showing a high protective effect against Salmonella enteritidis infection [18]. Zhang et al. indicated that rOmpA from Proteus mirabilis as an antigen could enhance the immune response in chickens [19]. Zhang et al. found that OMPs of Klebsiella pneumoniae as immunogen could stimulate mice to produce significant antigen-specific IgG such as IgG2a and IgG1, which could be developed as serotype-independent and multivalent vaccine against infection caused by K. pneumoniae [20]. Duan et al. found that purified rOmpA from Edwardsiella anguillarum as an immunogen could significantly improve the specific and non-specific immune response in Japanese eels, thus promoting the resistance of eel to E. anguillarum infection. They indicated that it may be a feasible strategy for preparing practical aquatic vaccines [21]. Diao et al. indicated that the immunogenic surface recombinant protein rOmpC from Aeromonas salmonicida subsp. masoucida (ASM) could elicit a strong humoral immune response in rainbow trout and then provide a significant protective effect against ASM infection [22]. The above studies suggest that OMPs from different Gram-negative bacteria are ideal antigens in mice, chickens or fish, but not all OMPs are effective; more work is needed to screen out proteins with higher immunogenicity. In addition, most of the current studies only immunized a single OMP from Gram-negative bacteria in animals to characterize its anti-infection effect on the homologous strain. The lack of studies on both the anti-infection effect on the heterologous strain and the horizontal comparison among different proteins has made it difficult to find the most characteristic antigen.
The E. coli O78 (CVCC CAU0768), which belongs to extraintestinal APEC with various virulence factors, was isolated from the faeces of diarrhea avian. The OMPs of this strain have not been revealed. Therefore, to search for more characteristic antigenic proteins, the OMPs including OmpA, OmpC and BamA from clinical E. coli O78 were cloned and expressed in this work. The efficacy of these recombinant proteins as subunit vaccines was evaluated in mice by immunized alone or in combination, the protective effect against both homologous strain and heterologous strain in vivo and in vitro was validated, and the most characteristic antigenic protein was determined by horizontal comparisons with different proteins.
2. Materials and Methods
2.1. Bacterial Strains and Plasmids
E. coli O78 (CVCC CAU0768) was donated by Professor Aike Li from the Academy of National Food and Strategic Reserves Administration. Shigella sonnei CVCC 3926 was donated by Professor Bo Shi from the Feed Research Institute of Chinese Academy of Agricultural Sciences. Salmonella enteritidis CVCC 3377 were bought from the China Veterinary Culture Collection Center (CVCC) (Beijing, China). Sh. flexneri CMCC (B) 51571 was purchased from the National Center for Medical Culture Collection (CMCC) (Beijing, China). Pseudomonas aeruginosa CICC 21630, P. aeruginosa CICC 10419 and E. coli O157 (CICC 21530) were purchased from the China Center of Industrial Culture Collection (CICC) (Beijing, China). E. coli competent strains of BL21 (DE3) and DH5α were purchased from TransGen Biotech Co., Ltd. (Beijing, China). The vector of pET-28a (+) was purchased from Novagen (Madison, WI, USA) and pMDTM 19-T Simple was purchased from TaKaRa Biotechnology (Dalian, China).
2.2. Characterization of E. coli O78
Classical virulence factors of APEC were amplified by PCR to characterize E. coli O78 (CVCC CAU0768). Eight virulence genes including irp2, iss, fimC, iroN, mat, iebB, iucD and fyuA were detected in E. coli O78. The primers, irp2-F: 5′-CTGATGAACTCACTCGCTATCC-3′, irp2-R: 5′-AGCATCTCCTGGCTCTGCTC-3′, iss-F: 5′-ATCACATAGGATTCTGCCG-3′, iss-R: 5′-CAGCGGAGTATAGATGCCA-3′, fimC-F: 5′-GCCGATGGTGTAAAGGATGG-3′, fimC-R: 5′-GGGTAAGTGCGCCATAATCA-3′, iroN-F: 5′-CCTCCGACGATGATAATGACG-3′, iroN-R: 5′-GATACCATTATGCGTAATGCC-3′, mat-F: 5′-CGACCTGGTCAGCAACAGCC-3′, mat-R: 5′-TCCACGCCCACATTCAGTGT-3′, iebB-F: 5′-GTTCTCACTCAGCCAGAACG-3′, iebB-R: 5′-CATCCAGCACTTCCAGATAAC-3′, iucD-F: 5′-GAAGCATATGACACAATCCTG-3′, iucD-R: 5′-CAGAGTGAAGTCATCACGCAC-3′, fyuA-F: 5′-ATGTGAAACTGCGTCTGGCG-3′, fyuA-R: 5′-CGCAGTAGGCACGATGTTGT-3′, were obtained to amplify the genes [23,24,25,26].
2.3. Homological Analysis of the OmpA, OmpC and BamA
Gene sequences of OmpA, OmpC and BamA from E. coli O78 (CVCC CAU0768) were obtained by PCR amplification and amino acid sequences were translated. The amino acid sequences were blasted by online database UniProt [27] (https://www.uniprot.org/blast/, accessed on 26 November 2021) with the below parameters: Matrix BLOSUM-62, E-Threshold 10, no filtering, and the number of feedback alignments was 50. Based on similarities, the blasted data of Escherichia, Shigella, Salmonella and Klebsiella from the top 50 sequences were aligned, the phylogeny trees were constructed by MEGA (version 11, Kim Dotcom, Auckland, New Zealand) with the method of Neighbor-Joining and the number of bootstrap replications was 1000 [28,29].
2.4. Cloning of the ompA, ompC and bamA Gene
Genomic DNA of E. coli O78 (CVCC CAU0768) was extracted by Bacteria DNA Kit (Tiangen Biotech Co., Ltd., Beijing, China). The primers with the respective restriction sites, including rOmpA F-EcoRI: 5′-GAATTCGCTCCGAAAGATAACACCTGGTACAC-3′, rOmpA R-NotI: 5′-GCGGCCGCAGCTTGCGGTTGAGTTACTACGTC-3′, rOmpC F-BamHI, 5′-CGGGATCCGCTGAAGTTTACAACAAAGACG-3′, rOmpC R-XhoI, 5′-CCCTCGAGTTAGAACTGGTAAACCAGACCC-3′, rBamA F-EcoRI: 5′-GAATTCAATTGGTTAGGTACAGGTTATGC-3′, rBamA R-NotI: 5′-GCGGCCGCCCAGGTTTTGCCGATGTTGAACT-3′, were obtained and used for amplifying the genes by PCR according to the previous method with some modifications [14,15,16]. The target fragments were cloned into the pMDTM 19-T Simple vector and transformed into E. coli DH5α. The positive recombinant plasmids were isolated by the Plasmid Extraction Kit (Tiangen Biotech Co., Ltd., Beijing, China) and digested with the corresponding enzymes. After digestion, target DNA fragments were ligated into the expression vector pET-28a (+) to construct the plasmids pET28a-ompA/ompC/bamA. The recombinant plasmids with purpose fragments were further transformed into the expression strain E. coli BL21 (DE3); positive strains were screened by PCR amplification and sequencing of DNA.
2.5. Expression and Purification of the rOmpA, rOmpC and rBamA Protein
The recombinant proteins with 6× His-tag were expressed in BL21 (DE3) induced by Isopropyl β-D-1-Thiogalactopyranoside (IPTG) [30]. In simple terms, the positive target strains were cultured with 250 rpm at 37 °C in LB medium, when the optical density at 600 nm (OD600nm) ranged from 0.40 to 0.60, the IPTG was added into the medium with the final concentration of 1 mM. Then, the strains were continually cultured under the same condition for 6–8 h. The finished induced cells were obtained by centrifuging with 5000× g at 4 °C for 30 min, the supernatant was removed, and the precipitate was resuspended in 50 mM Tris–HCl lysis buffer, (pH 7.9, involving 5 mg of lysozyme/gram of cell paste) with a volume of 8 mL/g wet weight. To disrupt the cells adequately, the solution was sonicated on ice for 20 min by a probe ultrasonicator (SCIENTZ, Ningbo Science Biotechnol Co., Ltd., Ningbo, China) with a frequency of 10 s off and 5 s on. Then, the solution was centrifuged with 14,500× g at 4 °C for 20 min, the supernatant was removed, and the insoluble inclusion bodies (IBs) were obtained. Next, the precipitated IBs were dissolved by 10 mM Tris-HCl denaturing buffer (pH 7.5, involving 8 M urea). The denatured solution was centrifuged with 14,500× g at 4 °C for 20 min, the supernatant was collected, an equal volume of 20 mM Tris–HCl refolding buffer (pH 7.9, involving 5% (v/v) lauryl dimethyl amine oxide (LDAO) and 1 M NaCl) was added dropwise with rapid stirring to refold the recombinant proteins [31].
The refolded recombinant proteins were purified according to the previous methods with some modifications [14]. Firstly, the His Bind column of Ni2+-NTA affinity chromatography (AKTAxpress, Fairfield, CT, USA) was pre-equilibrated with 20 mM Tris–HCl equilibration buffer (pH 7.9, involving 0.1% (v/v) LDAO, 500 mM NaCl and 0.04 M imidazole). Next, the refolded recombinant protein solution was loaded into the column and then washed with 20 mM Tris–HCl elution buffer (pH 7.4, involving 0.1% (v/v) LDAO, 500 mM NaCl and 0.5 M imidazole), the recombinant proteins were eluted and collected. Finally, the collected recombinant proteins were desalted by dialyzing with 20 mM Tris–HCl buffer (pH 7.4, involving 0.1% (v/v) LDAO and 0.15 M NaCl). The concentration and purity of recombinant proteins were analyzed by the BCA Protein Assay Kit (Tiangen Biotech Co., Ltd., Beijing, China) and Gel-Pro Analyzer™ version 6.3 (Bio-Rad, CA, USA), respectively.
2.6. Mouse Immunization
The immunization programs were conducted by the previous method with some modifications [14,28]. The SPF BALB/c mice were purchased from Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China). A total of 135 female SPF BALB/c mice aged from 6–8 weeks were divided into 9 groups and immunized with the purified rOmpA-adjuvant, rOmpC-adjuvant, rBamA-adjuvant, rOmpA + rOmpC-adjuvant, rOmpA + rBamA-adjuvant, rBamA + rOmpC-adjuvant, rOmpA + rBamA + rOmpC-adjuvant, PBS-adjuvant, respectively. One group of mice was not immunized and raised under the same conditions as a negative control. The purified proteins were diluted with sterile PBS to a final concentration of 0.5 mg/mL and then mixed with complete Freund’s adjuvant in a volume ratio of 1:1 for the primary immunization and incomplete Freund’s adjuvant for the booster immunizations. A volume of 100 μL/mouse antigen mixture was vaccinated by hypodermic injection for the primary immunization on day 0 and intraperitoneal injection for the booster immunizations on day 21 and 35. All mice were monitored daily and housed reasonably in the cages with ventilation (Suhang Technology Equipment Co., Ltd., Suzhou, China) which were changed weekly. Blood was collected on day 5, day 26 and day 40 through the tail vein of mice. Sera were divided into small portions and stored at −80 °C until used.
2.7. Antibody Titer and Antibody Isotypes Detection by iELISA
Antibody titer and antibody isotypes were detected by protein iELISA. Firstly, the 96-well ELISA plates were coated with 100 μL of each protein (rOmpA, rOmpC, rBamA and an equimolar mix rOmpA + rOmpC, rOmpA + rBamA, rOmpC + rBamA, rOmpA + rOmpC + rBamA) at a final concentration of 2 µg/mL in coating buffer (pH 9.6, 35 mM sodium bicarbonate, 15 mM sodium carbonate) and incubated at 4 °C overnight. Then, the plates were washed by PBST (10 mM PBS, involving 0.05% Tween 20) for 4 times. Next, 5% BSA which dissolved in PBST with a volume of 200 μL was used to block the plates at 37 °C for 4 h. After washing, a volume of 100 μL serially diluted (from 1:1000) mouse serum was added and the plates were incubated at 37 °C for 1.5 h. After washing, a volume of 100 μL Horseradish Peroxidase (HRP) conjugated goat anti-mouse total IgG, IgG1, IgG2a, IgG2b, IgG3, IgA and IgM which diluted at 1:5000 was added to plates and incubated at 37 °C for 30 min. After washing, 3,3′,5,5′-tetramethylbenzidine (TMB) with 100 μL/well was added and the plates were incubated in the dark at 37 °C for 25 min. Finally, the reaction was terminated by adding 100 μL H2SO4 with a concentration of 2 mol/L. The OD value of each well was measured by a spectrophotometer (TECAN, Grödig, Austria) at 450 nm with the reference wavelength of 630 nm. Endpoint titers were calculated as the reciprocal of the last serum dilution that gave a value 2.1 times higher than non-immunized negative control with a minimum value of 0.05 [32,33].
2.8. Cross-Protection Property Detected by iELISA
The cross-reaction properties of antisera against different Gram-negative bacteria including E. coli O78 (CVCC CAU0768), E. coli O157 (CICC 21530), Sh. sonnei CVCC 3926, Sh. flexneri strain CMCC (B) 51571, S. pullorum CVCC 1789, S. enteritidis CVCC 3377, P. aeruginosa CICC 21630 and P. aeruginosa CICC 10419 were measured by the whole-cell iELISA. A volume of 150 μL/well stationary liquid (0.1 M NaHCO3, involving 5% glutaraldehyde) was added to pretreat the plate at 37 °C for 2 h. After washing, a volume of 100 μL solution with 107 CFU bacteria was added to each well and aired at 37 °C [28]. The next steps were the same as for protein iELISA above.
2.9. Double-Immunodiffusion Assay
For analyzing the level of total IgG, a double-immunodiffusion assay was applied as described previously with some modifications [34]. Briefly, agarose powder was dissolved with sterile 0.9% NaCl solution in a glass beaker to reach a final concentration of 1%. The mixture solution was heated at 100 °C about 5 min to obtain a homogeneous solution, which was moved to a 56 °C water bath for heat preservation. A volume of 10 mL melted solution at 56 °C was placed in a small beaker, and the goat anti-mouse IgG antibody was added with the concentration determined by the pre-test (final concentration 250 μg/mL). After thorough mixing, the solution with antibody was gently poured to plastic plates which were placed on a horizontal surface and kept for 30 min. When the solution solidified into a flat gel, a series of holes with an aperture of 3–4 mm and a hole distance of 10 mm were punched with a puncher. A volume of 10 μL standard mouse IgG with different concentrations (0.75, 0.5, 0.25, 0.125 and 0.0625 mg/mL) were accurately added into the hole with micropipette. Likewise, the serum to be tested was diluted into 1:2 with 0.9% NaCl solution and added to the hole. Then, the plates were covered and kept in a humidified chamber that contained wet cotton and placed on a horizontal surface at 37 °C for 24 h. The diameter of the precipitation ring was measured and recorded. With 5 gradient concentrations of standard mouse IgG as independent variables and the diameter of the precipitation ring as dependent variables, a regression equation was established through linear regression and correlation to calculate the IgG content in immune serum. All the experiments were performed in triplicate.
2.10. Opsonophagocytosis Assay
The serum on day 40 was used for the Opsonophagocytosis assay as described previously with some modifications [14,35]. Briefly, a volume of 10 μL antisera was incubated with 90 μL of mid-log phase E. coli O78 (CVCC CAU0768) cells with the concentration of 4 × 104 CFU/mL at 37 °C for 30 min. Then, a volume of 100 μL of phagocyte suspension which was obtained from peritoneal fluid of mice with the concentration of 4 × 106 cells/mL was added into the mixed solution with antisera and bacteria, and then incubated at 37 °C for 1 h. After that, sterile water was added to the mixture to lyse the phagocytes, then the mixture solution was diluted in serial, and available bacteria was evaluated by plate count. The formula (1 − (No. of CFU recovered in the presence of phagocytes/No. of CFU recovered in the absence of phagocytes) × 100) was used to calculate the bacterial killing rate [14].
2.11. Challenge Assay
The in vivo immune protection induced by recombinant protein in mice was characterized by the challenge with homologous and heterologous strains. An absolute lethal dose (LD100, 3 × 107 CFU/mice) of ExPEC E. coli O78 (CVCC CAU0768) in mice was determined in advance. On day 49, 10 mice of each group were intraperitoneally injected with 2 × LD100 mid-log phase E. coli to evaluate the effectiveness of recombinant proteins against a homologous strain. Mortality was recorded for 7 days. The remaining 5 mice of each group were intragastrically administrated with 1010 CFU mid-log phase IPEC E. coli O157 (CICC 21530) after fasting for 12 h to identify the protective effect of recombinant proteins on the heterologous strain. Fecal samples were collected every day during the 7-day observation period, and the fecal shedding of bacteria was determined by plate colony count [28].
3. Result
3.1. Characterization of E. coli O78
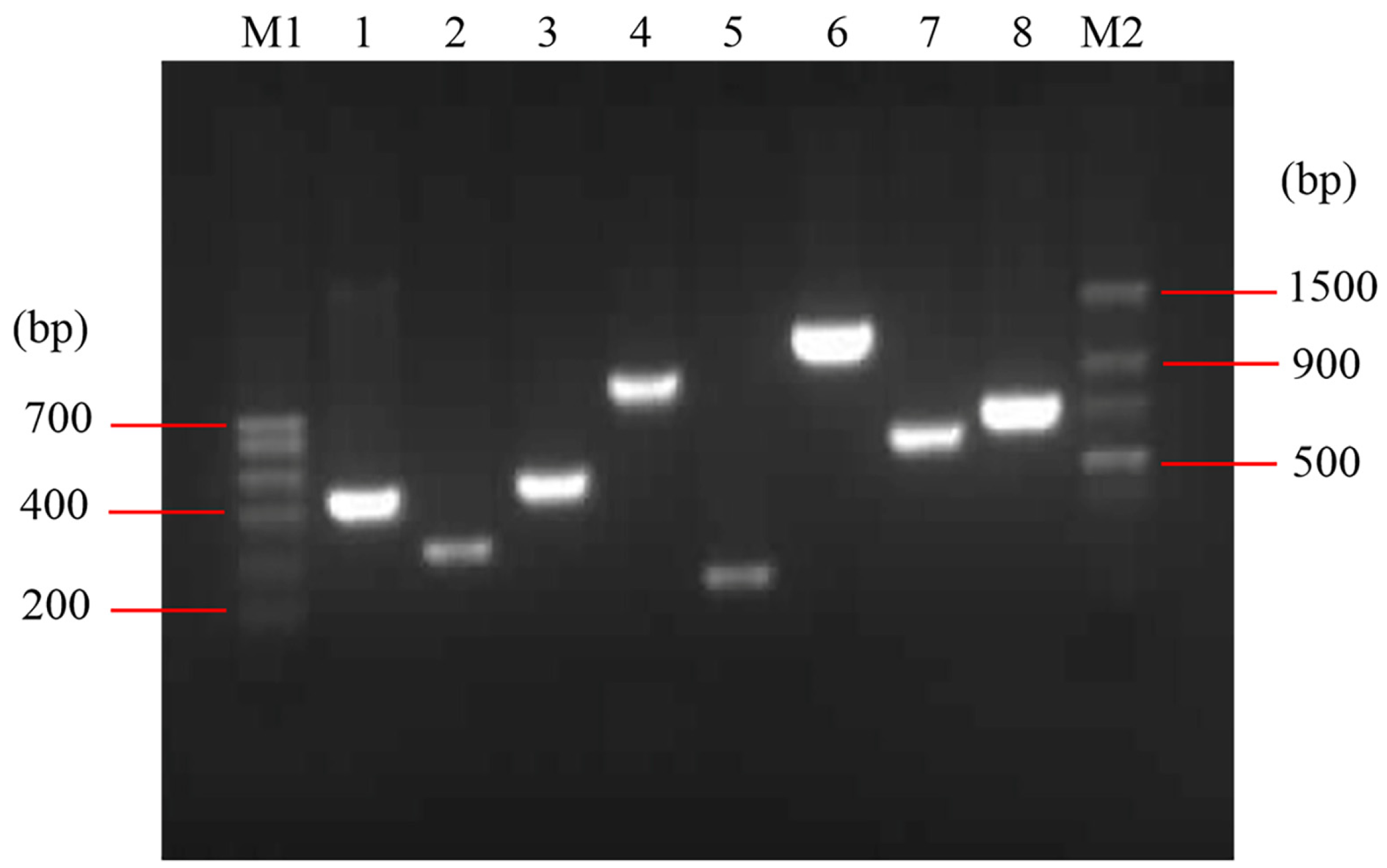
The detected virulence factors of E. coli O78 (CVCC CAU0768) were related to adhesion (mat, fimC), iron acquisition system (iroN, fyuA, irp2 and iucD), invasion (ibeB) and protectins/serum resistance (iss) (Figure 1). The mat is temperature-regulated fimbriae and associated with meningitis, fimC is responsible for adherence and colonization of epithelial cells. iroN is a catecholate siderophore (salmochelin) receptor, fyuA is a ferric yersiniabactin receptor, irp2 is responsible for iron repressible protein yersiniabactin synthesis, and iucD is related to the mediation of aerobactin synthesis. The iss promotes bacterium protection, which mainly relates to the action of the complement system, and ibeB is involved in APEC invasion and pathogenicity [23,24,25,26]. The result indicated that E. coli O78 (CVCC CAU0768) used in this work is a virulent strain with multiple virulence factors, which is a typical representative in APEC.
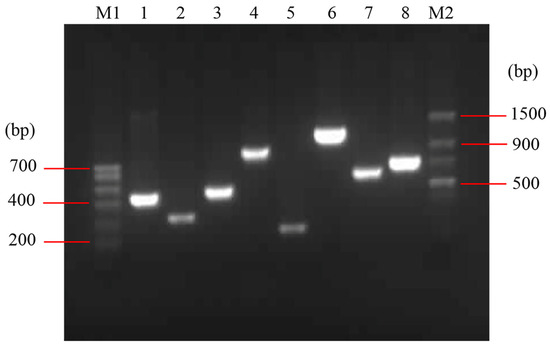
Figure 1.
Identification of virulence factors in E. coli O78 (CVCC CAU0768). M1: Trans DNA Marker I; M2: Trans DNA Marker II; lane 1–8: irp2, iss, fimC, iroN, mat, iebB, iucD and fyuA, respectively.
3.2. Homological Analysis of the OmpA, OmpC and BamA
Blasting was conducted by the UniProt database, the top 50 sequences with the highest similarity to OmpA, OmpC and BamA of E. coli O78 (CVCC CAU0768) were mostly from Escherichia, Shigella, Salmonella and Klebsiella. The OmpA shares 88.4–98.8% identity with other Escherichia, and shares 94.5–96.1%, 91.5–92.1% and 83.0–86.7% identity with Shigella, Salmonella and Klebsiella, respectively. The OmpC shares 91–97.7% identity with other Escherichia, and shares 92.4–94.3%, 79.2–80% and 79.2–81.7% identity with Shigella, Salmonella and Klebsiella, respectively. The BamA shares 86.5–100% identity with other Escherichia, and shares 100%, 92.3–93.7% and 85.5–87.4% identity with Shigella, Salmonella and Klebsiella, respectively. The results of blast and phylogenetic analysis showed that OmpA, OmpC and BamA from E. coli O78 (CVCC CAU0768) were highly homologous to those in Escherichia, Shigella, Salmonella and Klebsiella (Figure S1), indicating that these OMPs have potential to be developed into universal antigens against a variety of Gram-negative bacteria.
3.3. Cloning, Expression, and Purification of the Recombinant Proteins
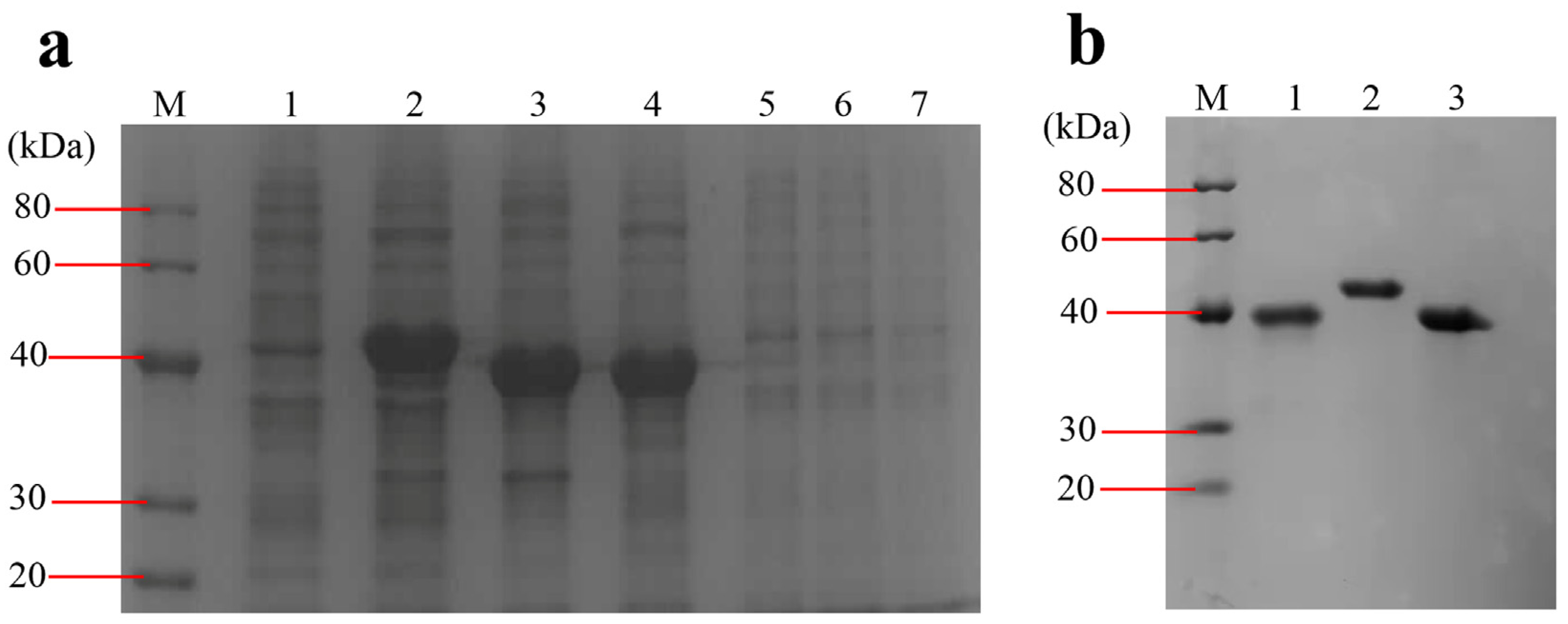
The DNA fragments of ompA (GenBank Accession No. OM350111), bamA (GenBank Accession No. OM365872) and ompC (GenBank Accession No. OM365873) with the size of 975 bp, 1089 bp and 1026 bp, respectively, were amplified from E. coli O78 (CVCC CAU0768) DNA genome. The digestion fragments were cloned into the pET28a expression vector and the recombinant proteins were expressed in E. coli BL21 (DE3) induced by IPTG. The recombinant proteins are present in the IBs and fused with an N-terminal 6× His tag. The molecular weight of rOmpA, rBamA and rOmpC was approximately 40 kDa, which accorded with their theoretical values (OmpA 35,158.34 Da, OmpC 37,935.18 Da, BamA 40,731.58 Da). After purification, the concentrations of rOmpA, rBamA and rOmpC were 1.122 mg/mL, 1.167 mg/mL and 2.046 mg/mL, respectively (Figure 2). After purification and refolding, the purities of rOmpA, rBamA and rOmpC were 95%, 98% and 97%, respectively.
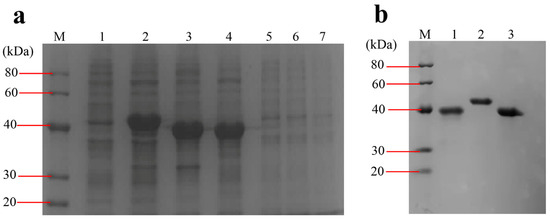
Figure 2.
Expression and purification of the OMPs. (a) Expression of the OMPs. Lane M, protein marker (20–40 kDa); lane 1, total proteins of BL21 harboring pET28a vector induced by IPTG; lane 2–4, total proteins of BL21 harboring pET28a-bamA, pET28a-ompA, pET28a-ompC induced by IPTG, respectively; lanes 5–7, total proteins of BL21 harboring pET28a-bamA, pET28a-ompA, pET28a-ompC without being induced by IPTG, respectively. (b) Purified OMPs. Lane 1–3, eluent of the purified rOmpA, rBamA, rOmpC, respectively.
3.4. Immunogenic Property of Recombinant Proteins in Mice
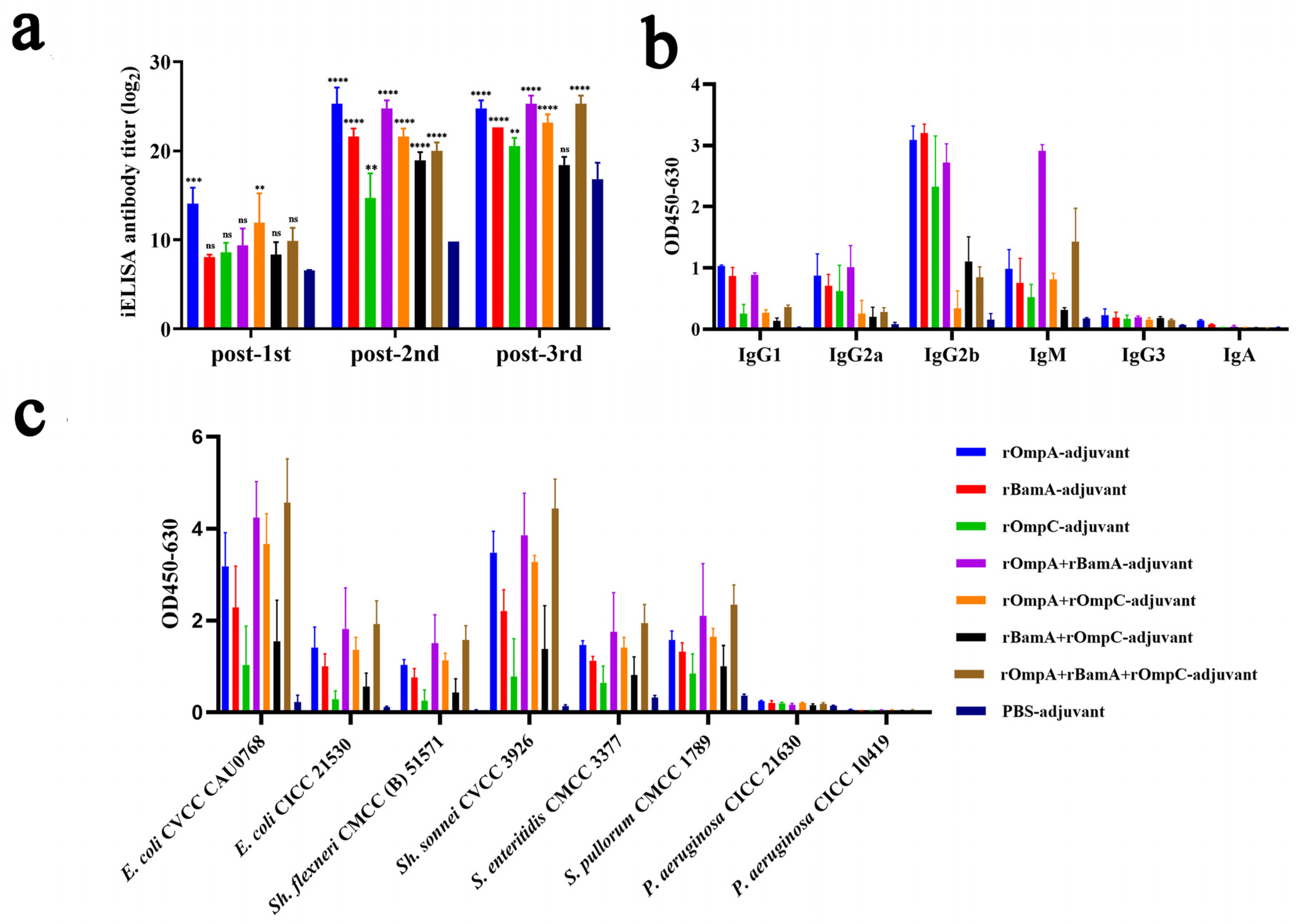
To evaluate the immunogenicity properties of these recombinant proteins, antisera were collected from mice on days 5, 26 and 40 and the specific reactivity with purified recombinant proteins was tested by iELISA. Results revealed that mice immunized with recombinant proteins alone or in combination both exhibited significant immune response; a strong and significant immune response of antisera to the recombinant proteins was detected after the second immunization (Figure 3a). For immunization alone groups, the highest antibody titer reached 1:32,805,000 from the rOmpA-adjuvant group after the third immunization, which was significantly higher than the rOmpC-adjuvant group (1:1,701,000) and rBamA-adjuvant group (1:6,561,000). For the immunization in combination groups, the highest antibody titer reached 1:45,927,000 from both the rOmpA + rBamA-adjuvant group and rOmpA + rBamA + rOmpC-adjuvant group after the third immunization. All recombinant protein immunization groups showed significant differences compared with the PBS-adjuvant group after the second immunization (Figure 3a).
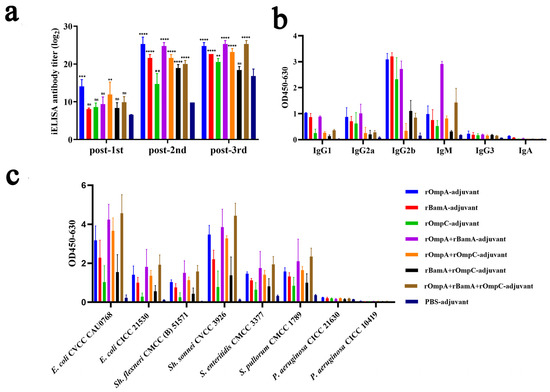
Figure 3.
Immune responses in antisera from mice immunized with recombinant proteins. (a) Antibody titers of the antisera against recombinant proteins. “**, *** and ****” indicates a significant difference between the immunized group and the PBS-adjuvant group (** p ≤ 0.01, *** p ≤ 0.001, **** p ≤ 0.0001). “ns” indicates that there is no significant difference between the recombinant protein immunized group and the PBS-adjuvant group. (b) Antibody isotypes in antisera from immunized mice. The serum dilution was 1:1000. (c) Cross-reaction properties of the antisera against different bacteria. The serum dilution was 1:27,000. Data are expressed as the means ± SD (n = 3).
Antibody isotypes in the serum of immunized mice were analyzed by protein iELISA, and the result showed that IgG1, IgG2a, IgG2b, and IgM were detected (Figure 3b); among them, IgG2b exhibited the highest OD value (0.17–3.35), and then in order, IgM (0.27–1.31), IgG2a (0.09–1.28) and IgG1 (0.08–1.05), and IgG3 and IgA were detected from the recombinant protein immunized groups with low OD values (<0.35). For the PBS-adjuvant group, the OD values of above all antibody isotypes were also very low (<0.26). Moreover, the cross-reaction properties of antisera against different Gram-negative bacteria including Escherichia, Shigella, Salmonella and Pseudomonas were conducted by whole-cell iELISA assay, among which Pseudomonas strains were detected as negative controls because of the low homology to E. coli O78 (CVCC CAU0768). The results showed that the antisera from recombinant protein immunized groups with high OD values had a high affinity to E. coli O78 (CVCC CAU0768) (0.47–5.43), E. coli O157 (CICC 21530) (0.16–2.85), Sh. sonnei CVCC 3926 (0.26–5.10), Sh. flexneri CMCC (B) 51571 (0.10–2.21), S. pullorum CVCC 1789 (0.52–2.80), and S. enteritidis CVCC 3377 (0.39–2.72) (Figure 3c), while the affinity of antisera to P. aeruginosa was low (<0.26). Furthermore, the antisera from the PBS-adjuvant group with low OD values have low affinity to all above strains (<0.39).
In general, rOmpA immunized alone or in combination with other proteins could stimulate mice to produce a higher antibody titer, and the antisera from these groups had the highest affinity capacity to a variety of Gram-negative bacteria, indicating that rOmpA was the most immunogenic outer membrane protein in this work, and the methods of combination immunization could improve the immune response in mice.
3.5. Double-Immunodiffusion Assay
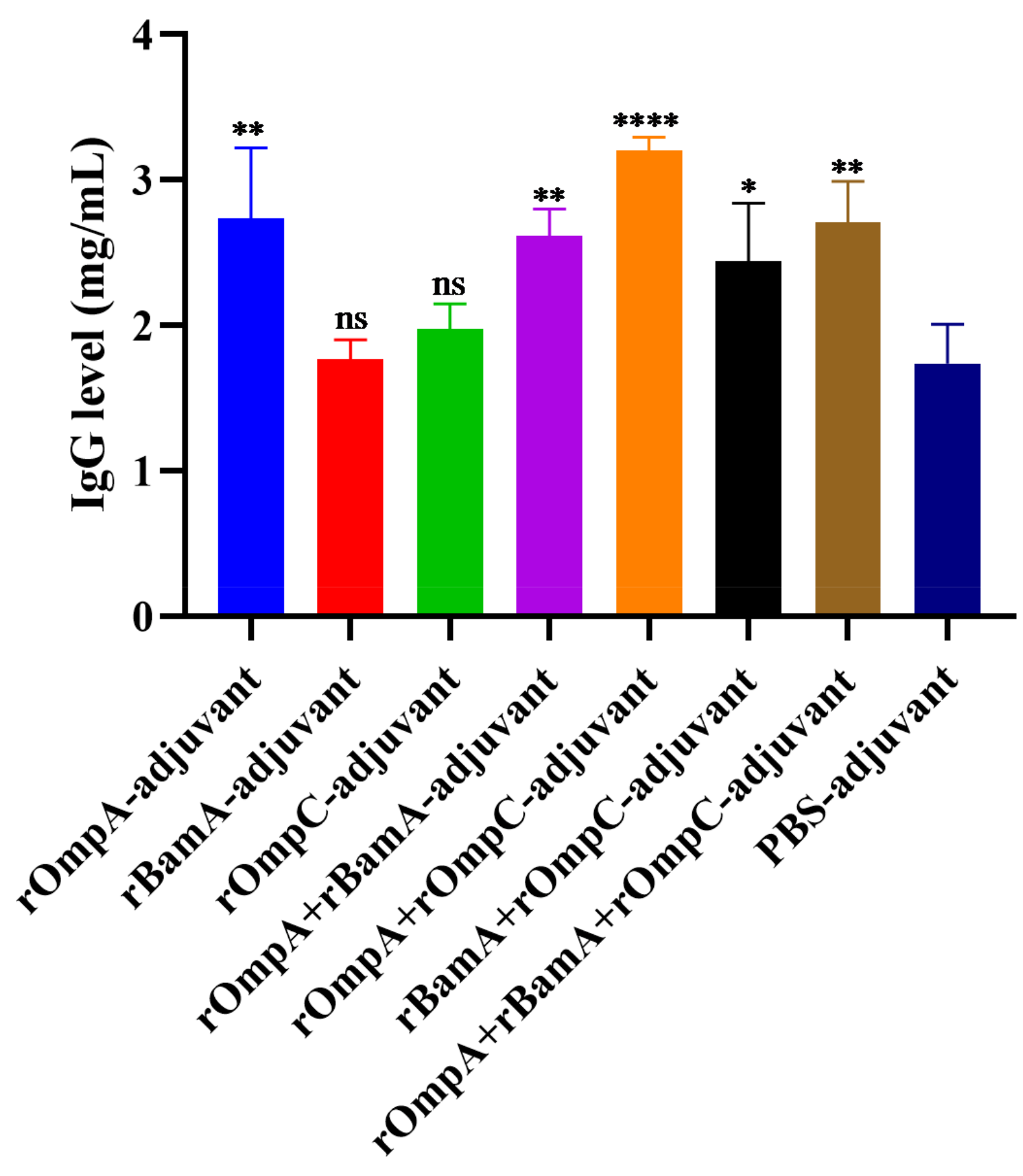
Quantitative analysis of total IgG levels in antisera after the third immunization was conducted by double-immunodiffusion assay. The level of total IgG in the rOmpA + rOmpC-adjuvant group (3.21 ± 0.09 mg/mL) was the highest among all immunized groups, which was extremely significantly different from the PBS-adjuvant group (1.74 ± 0.27 mg/mL). For immunization alone groups, only the rOmpA-adjuvant group (2.74 ± 0.49 mg/mL) had a significant difference compared with the PBS-adjuvant group (Figure 4). However, there was no significant difference between the PBS-adjuvant group and the rBamA-adjuvant group as well as the rOmpC-adjuvant group. The above results further indicated that rOmpA was the most potential antigenic protein compared with the other two proteins, and the methods of combination immunization could improve the immune response in mice.
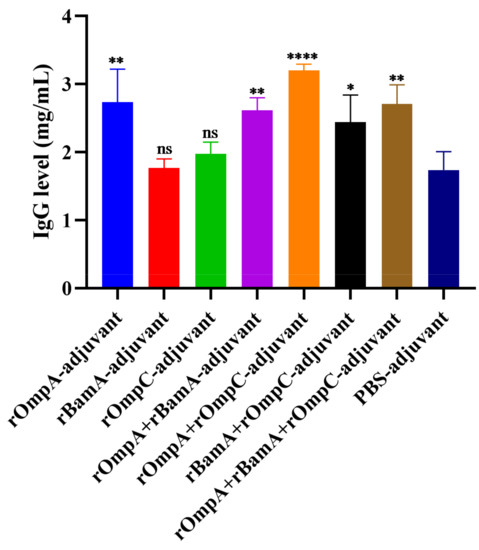
Figure 4.
The level of total IgG of the antisera. Data are expressed as the means ± SD (n = 3). “*, ** and ****” indicates a significant difference between the recombinant protein immunized group and the PBS-adjuvant group (* p ≤ 0.05, ** p ≤ 0.01, **** p ≤ 0.0001). “ns” indicates that there is no significant difference between the recombinant protein immunized group and the PBS-adjuvant group.
3.6. Opsonophagocytosis Assay
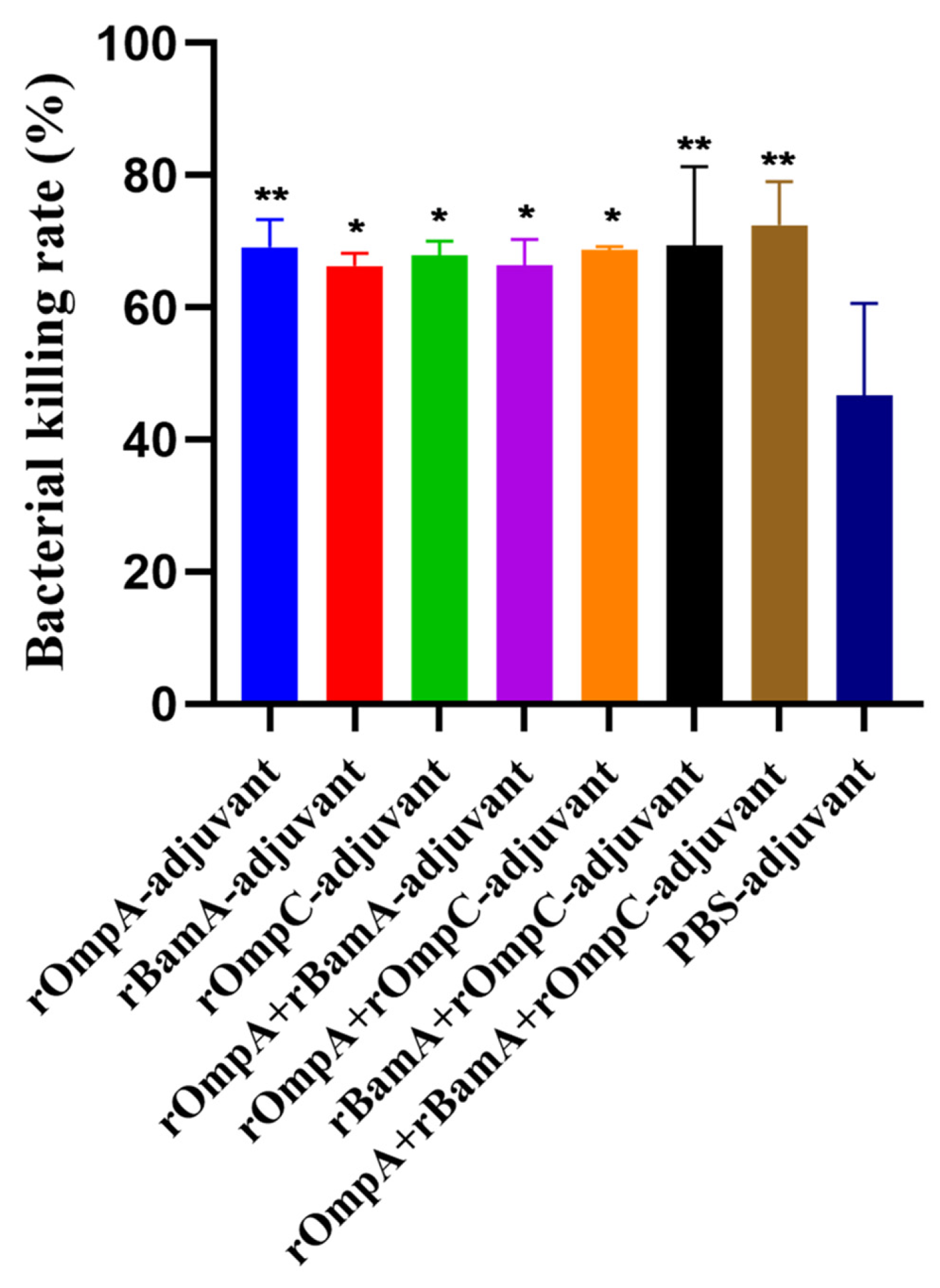
E. coli O78 (CVCC CAU0768) cells were incubated with phagocytes and antisera which were obtained from mice immunized with recombinant proteins or PBS. The bacterial killing rates of rOmpA-adjuvant, rBamA-adjuvant, rOmpC-adjuvant, rOmpA + rBamA-adjuvant, rOmpA + rOmpC-adjuvant, rBamA + rOmpC-adjuvant, rOmpA + rBamA + rOmpC-adjuvant and PBS-adjuvant immunization groups were 69.09 ± 4.19%, 66.28 ± 1.93%, 67.91 ± 2.15%, 66.34 ± 3.96%, 68.73 ± 0.49%, 69.35 ± 11.92%, 72.48 ± 6.56% and 46.73 ± 13.88%, respectively. The bacterial killing rates of recombinant protein immunization groups were all significantly higher than that of the PBS-adjuvant immunization group, the highest bacterial killing rate was from the rOmpA + rBamA + rOmpC-adjuvant group, and the bacterial killing rate of the rOmpA-adjuvant group was higher than those of rBamA-adjuvant group and rOmpC-adjuvant group (Figure 5). The above results showed that the antiserum of mice from the recombinant proteins immunization groups had a more significant opsonophagocytosis effect than that from the PBS-adjuvant group.
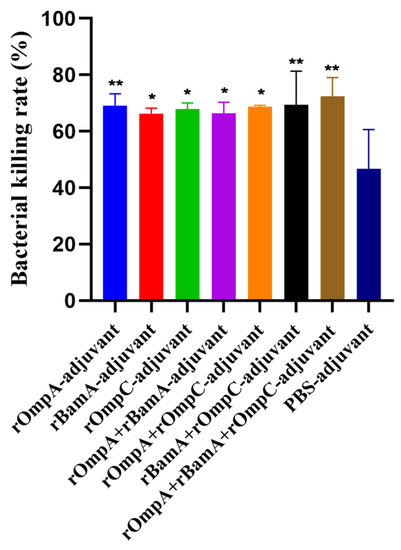
Figure 5.
Opsonophagocytosis effect of the antisera in vitro. Colony counts of different groups were determined by plate count. The E. coli O78 (CVCC CAU0768) was incubated with antisera of the recombinant proteins immunization group or the PBS immunization group. Data are expressed as means ± SD, n = 3. “* and **” indicates a significant difference between the immunized group and the PBS-adjuvant group (* p ≤ 0.05, ** p ≤ 0.01).
3.7. Protection Efficacy of Recombinant Proteins against E. coli In Vivo
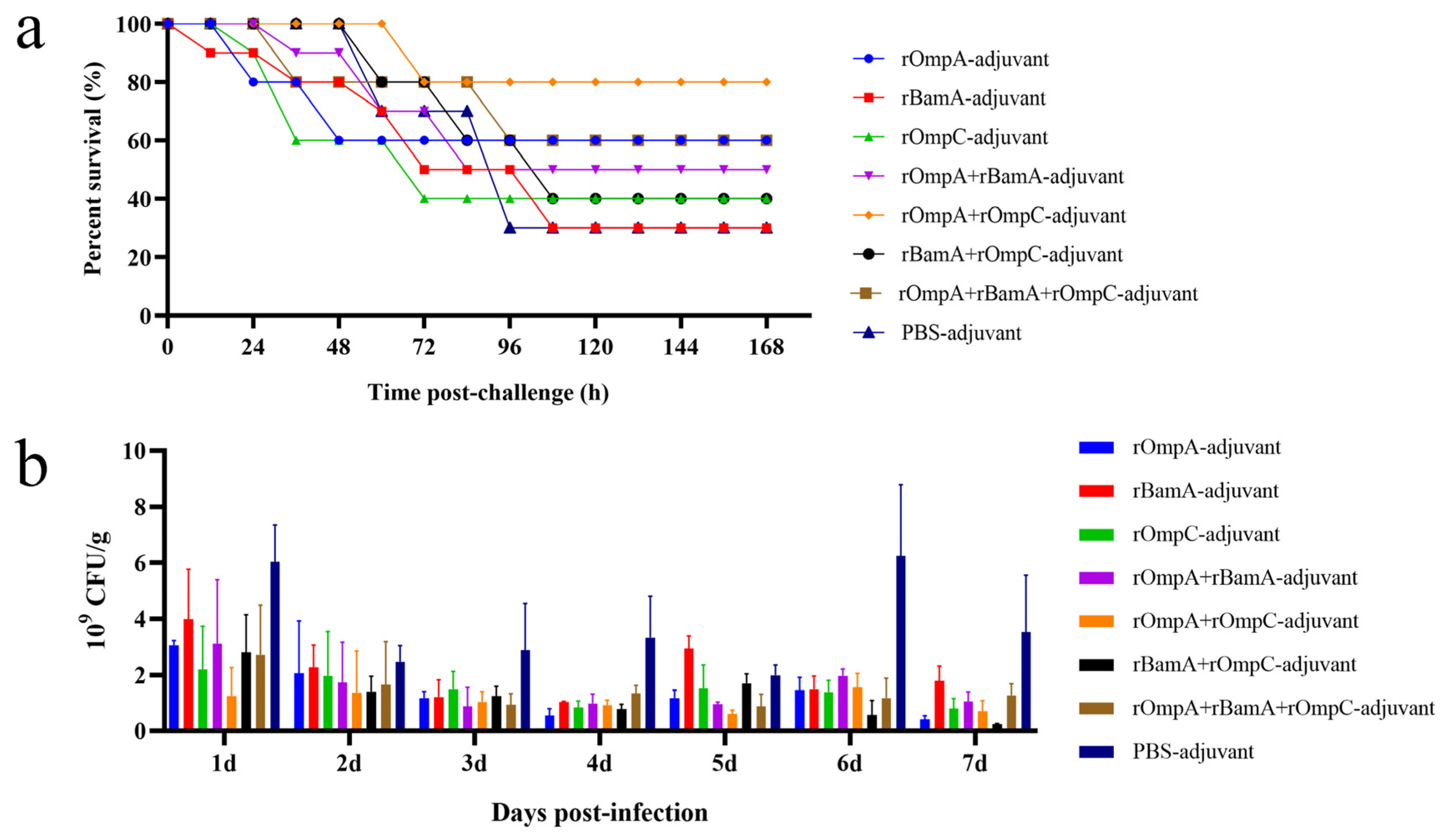
On day 49, mice were intraperitoneally injected with 2 × LD100 mid-log phase E. coli O78 (CVCC CAU0768) to evaluate the immune response induced by recombinant protein against homologous strain infection in vivo. The survival rate: rOmpA-adjuvant group 60%, rOmpC-adjuvant group 40%, rBamA-adjuvant group 30%, rOmpA + rOmpC-adjuvant group 80%, rOmpA + rBamA-adjuvant group 50%, rBamA + rOmpC-adjuvant group 40%, rOmpA + rBamA + rOmpC-adjuvant group 60%, PBS-adjuvant group 30% (Figure 6a). For immunization alone groups, the rOmpA-adjuvant group exhibited the highest survival rate of 60%; for the immunization in combination groups, the rOmpA + rOmpC-adjuvant group showed the highest survival rate of 80%. These results further indicated that rOmpA was the most potential antigenic protein compared with the other two proteins, and the methods of combination immunization could improve the immune response in mice, which was consistent with the result of antisera characterized in vitro.
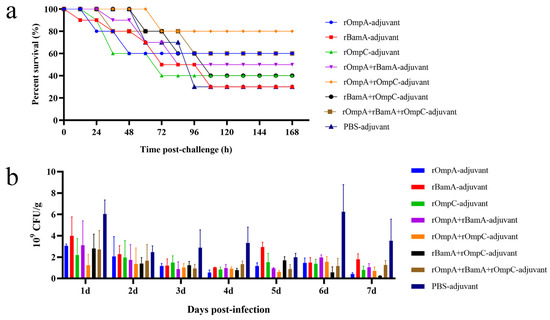
Figure 6.
Protection efficacy of the recombinant proteins against E. coli in vivo. (a) Immunized mice were intraperitoneally injected with mid-log phase E. coli O78 (CVCC CAU0768); survival rate was observed for 168 h after the challenge. (b) Immunized mice were challenged with mid-log phase E. coli O157 (CICC 21530) by gastric tube, and fecal shedding was observed for seven days after the challenge, data are expressed as the means ± SD, n = 3.
The antisera of immunized mice displayed cross-reaction to different Gram-negative strains in vitro (Figure 3c), including the IPEC E. coli O157 (CICC 21530). Therefore, immunized mice were intragastrically administrated with E. coli O157 to further identify the immune response induced by recombinant proteins against heterologous strain infection in vivo. The result of colony counting of fecal samples showed that the recombinant proteins immunization groups had a lower count and a gradual downward trend of E. coli in 7 days, while the colony count of the PBS + adjuvant group showed an unstable trend (Figure 6b). The colony counting of the rOmpA + rBamA-adjuvant group, rOmpA + rOmpC-adjuvant group and rOmpA + rBamA + rOmpC-adjuvant group had a significant difference compared with that of the PBS + adjuvant group from the third day after infection (Table S1), indicating that mice immunized with several recombinant proteins in combination could significantly reduce the colonization of heterologous E. coli in the intestine and maintain intestinal flora stability.
4. Discussion
There have been many studies on outer membrane proteins as subunit vaccines, some of which showed desirable immune effects while some of which were not satisfactory [14,15,16,28,36]. Previous studies showed that OmpA from Gram-negative bacteria was one of the most immunodominant antigens with various properties required for vaccine candidates, such as being involved in bacterial attachment, bacterial conjugation and as receptors for certain bacteriophages [17,37,38]. Therefore, in this work, the recombinant outer membrane proteins including OmpA, BamA and OmpC from highly toxic strain of E. coli O78 (CVCC CAU0768) were cloned, analyzed and expressed. The immune effects of these three proteins alone and in combination were evaluated in the mouse model to further identify proteins with more potential.
According to the result of blast on UniProt, the outer membrane proteins were highly conserved in different Gram-negative bacterial strains in the current work. Compared with our previous study [14,15,16], it was found that OmpA, BamA and OmpC from E. coli O78 (CVCC CAU0768) not only showed high homology to those in Escherichia, Salmonella and Shigella but also those in Klebsiella, among which BamA was the most conservative, and its identity with BamA of 26 strains including Escherichia coli and Shigella reached 100%. Secondly, OmpA had the highest identity with which of Escherichia coli strain K12 and O157:H7, reaching 98.8%. Finally, OmpC had the highest identity with which of Escherichia coli O157:H7, which was 97.7% (Figure S1). Previous studies showed that the high conserved property of OMPs made it potential for developing universal vaccines against multiple types of bacteria. Gao et al. showed that OMP VP1243 from Vibrio parahaemolyticus possessed immunogenicity and was highly conserved among the major Vibrio specie; cross-reactive immune response against several Vibrio species including V. parahaemolyticus, V. vulnificus and V. alginolyticus were induced in vaccination mice [39]. Peng et al. reported that OMPs from Vibrio parahaemolyticus were similar to those in P. fluorescens and A. hydrophila, which could induce protective immunity against not only Vibrio parahaemolyticus but also Pseudomonas fluorescens and A. hydrophila in vivo [40]. Wang et al. showed that OMPs from E. coli were highly conserved and had cross-reaction to different Gram-negative bacteria in vitro; they indicated that these OMPs had the potential to be developed into vaccines against Escherichia, Shigella and Salmonella [14,15,16]. Based on previous studies and the high conserved property of OmpA, BamA and OmpC from E. coli O78 in the current work, the cross-reaction of these three OMPs was characterized. The whole-cell iELISA assay showed that the antisera from mice immunized with these OMPs exhibited high affinity to multiple strains including Escherichia, Shigella and Salmonella in vitro (Figure 3c), which was consistent with the homological analysis and previous reports [14,15,16], indicating that the OMPs in this work also had the potential to be developed into vaccines against multiple Gram-negative bacteria. However, the cross-protection of OMPs against Gram-negative bacteria in vivo needs further study.
In this work, rOmpA, rBamA and rOmpC from E. coli O78 (CVCC CAU0768) were separately mixed with Freund’s adjuvant in a volume ratio of 1:1, and the antibody titer of antisera was 1:32,805,000, 1:6,561,000 and 1:1,701,000, respectively; the antibody titer of the PBS-adjuvant group was 1:171,000 (Figure 3a). Previous studies showed that rOmpA, rBamA and rOmpC from E. coli CVCC 1515 were separately mixed with Freund’s adjuvant in a volume ratio of 3:1, the antibody titer of antisera from immunized mice was 1:642,000, 1:736,000 and 1:240,000, respectively, and the antibody titer of the PBS-adjuvant group was lower than 1:100 [14,15,16]. The above results indicated that at the same injection volume, the proportion of Freund’s adjuvant had a considerable influence on the level of the immune response; the antibody titer in antisera may increase with the proportion of Freund’s adjuvant. In addition, the nonspecific immune response of the PBS-adjuvant group was also found in the assays of total IgG level detection (Figure 4). Studies have shown that Freund’s adjuvant was one of the most effective adjuvants for inducing high antibody titer and cell-mediated immunity (Th1) [41,42]. The previous report indicated that the function of complete Freund’s adjuvant (CFA) may be sufficient to prevent group B Coxsackieviruses infection, and the disease resistance ability of CFA may be attributed to the adjuvanticity of M. tuberculosis [43]. Therefore, the nonspecific immune response in the PBS-adjuvant group in this work may also be related to the presence of the cell wall component of M. tuberculosis in the CFA. Studies showed that the type of adjuvant, the mixture ratio of antigen and adjuvant, injection dose and administration mode all have a non-negligible influence on the generation of the immune response [44,45], which need to be further explored.
Previous reports showed that rOmpA from several Gram-negative bacteria including Escherichia, Shigella, Proteus and Edwardsiella was an ideal antigen and could trigger significant humoral and/or cell-mediated immunity in inoculated animals [15,17,19,21]; especially, Pore et al. indicated rOmpA showed a competency to coordinate not only adaptive but also innate responses immune in mice [17]. In the current work, compared with rBamA and rOmpC, rOmpA was the most efficient antigen with more significant humoral immunity in inoculated mice, indicating rOmpA from E. coli O78 was also an ideal antigen that could trigger significant protective humoral immunity in inoculated animal, while the cell-mediated immunity and innate immune responses needed a further study. Moreover, in vitro and in vivo assays showed that mice immunized with rOmpA in combination with the other two proteins were detected with the highest antibody titer (Figure 3a), the highest affinity to multiple Gram-negative bacteria (Figure 3c), the highest total IgG level (Figure 4), the most significant opsonization on macrophages (Figure 5), the highest survival rate of homologous strain (E. coli O78) challenge (Figure 6a) and the most effective anti-infection effect of heterologous strain (E. coli O157) challenge (Figure 6b), indicating that the methods of combination immunization could increase the abundance of antigens, thus effectively promoting the humoral immunity with more production of functional antibodies against not only homologous strain infection but also heterologous strain infection in mice. E. coli O157 (CICC 21530) belongs to EHEC of IPEC, while E. coli O78 (CVCC CAU0768) belongs to APEC of ExPEC [46,47,48]; there was a high identity between their outer membrane proteins according to the homology analysis (Figure S1), so the proteins from E. coli O78 (CVCC CAU0768) also had a cross-protective reaction against the E. coli O157 infection in vivo. Meanwhile, although most in vitro experiments showed that rOmpC and rBamA also stimulated a considerable immune response in mice; the protection effect of bacterial challenge in vivo was less than satisfactory compared with the PBS-adjuvant group. It has been shown that IgG1 antibody was elicited by Th2 lymphocytes, while antibodies of IgG2a, IgG2b and IgG3 were elicited by Th1 lymphocytes; IgG and IgM mainly exist in blood and play important role in antibacterial, antiviral, neutralizing toxins, etc. [49,50,51]. In this work, various antibody isotypes including IgG1, IgG2a, IgG2b, and IgM were detected (Figure 3b) in antisera, which may be a vital reason for the immunized mice to resist the challenge of both homologous and heterologous strain.
In conclusion, the high conserved property of OmpA, BamA and OmpC from E. coli O78 (CVCC CAU0768) indicated that they could be developed into universal antigens. In vitro assays showed that antisera from immunized mice had a high affinity to multiple Gram-negative bacteria; in vivo challenge showed that mice immunized with these recombinant proteins could resist not only homologous strains but also heterologous strains. rOmpA was characterized as the most efficient antigen in this work, which provided a basis for the selection of the most effective antigen in the future. Furthermore, it was found that a stronger immune response in mice could be triggered when immunized with several proteins in combination, which provided the reference for the construction of novel antigens with higher immunogenicity. However, there are still some limitations in this work; the cross-protection of OMPs against Gram-negative bacteria in vivo and more immune-related indexes are needed to be detected to further explore the immune mechanism of OMPs.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms10050982/s1, Figure S1: Phylogenetic trees of OmpA (a), BamA (b) and OmpC (c) in Escherichia, Shigella, Salmonella and Klebsiella constructed by MEGA; Table S1: Significance analysis for fecal shedding between the recombinant protein immunized group and the PBS-adjuvant group.
Author Contributions
Conceptualization, G.P., R.M., N.Y., D.T. and J.W.; methodology, G.P., R.M., N.Y., D.T. and J.W.; software, G.P. and R.M.; validation, G.P., R.M. and N.Y.; formal analysis, G.P., R.M. and N.Y.; investigation, G.P., R.M. and N.Y.; resources, Y.H.; data curation, G.P., R.M. and N.Y.; writing—original draft preparation, G.P.; writing—review and editing, R.M. and N.Y.; visualization, R.M. and N.Y.; supervision, J.W.; project administration, J.W.; funding acquisition, J.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (grants No. 31872393), the Direction of Antibiotic Peptides and Alternatives to Antibiotics, the Agricultural Science and Technology Innovation Program (CAAS-ASTIP-2013-FRI-02) and Key Project of Alternatives to Antibiotic for Animal Usages of ASTIP (CAAS-ZDRW202111) in CAAS and the Key Laboratory of Biological Products and Chemical Drugs for Animals, Ministry of Agriculture, China Animal Husbandry Industry Co., Ltd. (No. 2020110001002012).
Institutional Review Board Statement
The mouse experiment was performed according to the Animal Care and Use Committee of the Feed Research Institute of Chinese Academy of Agricultural Sciences (CAAS) and approved by the Laboratory Animal Ethical Committee and its Inspection of the Feed Research Institute of CAAS (AEC-CAAS-20090609).
Informed Consent Statement
Not applicable.
Data Availability Statement
All datasets generated for this study are included in the article/Supplementary Material.
Acknowledgments
We would like to thank Aike Li from the Academy of National Food and Strategic Reserves Administration and Bo Shi from the Feed Research Institute of Chinese Academy of Agricultural Sciences for the donation of bacterial strains.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Johnson, J.R.; Russo, T.A. Extraintestinal pathogenic Escherichia coli: “The other bad E coli”. J. Lab. Clin. Med. 2002, 139, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Vila, J.; Sáez-López, E.; Johnson, J.R.; Römling, U.; Dobrindt, U.; Cantón, R.; Giske, C.G.; Naas, T.; Carattoli, A.; Martínez-Medina, M.; et al. Escherichia coli: An old friend with new tidings. FEMS Microbiol. Rev. 2016, 40, 437–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, L.W. Distinguishing pathovars from nonpathovars: Escherichia coli. Microbiol. Spectr. 2020, 8, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Tapader, R.; Basu, S.; Pal, A. Secreted proteases: A new insight in the pathogenesis of extraintestinal pathogenic Escherichia coli. Int. J. Med. Microbiol. IJMM 2019, 309, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Joseph, A.; Cointe, A.; Mariani Kurkdjian, P.; Rafat, C.; Hertig, A. Shiga toxin-associated hemolytic uremic syndrome: A narrative review. Toxins 2020, 12, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwidder, M.; Heinisch, L.; Schmidt, H. Genetics, toxicity, and distribution of enterohemorrhagic Escherichia coli hemolysin. Toxins 2019, 11, 502. [Google Scholar] [CrossRef] [Green Version]
- Biran, D.; Ron, E.Z. Extraintestinal pathogenic Escherichia coli. Curr. Top. Microbiol. Immunol. 2018, 416, 149–161. [Google Scholar] [CrossRef]
- Vandekerchove, D.; De Herdt, P.; Laevens, H.; Pasmans, F. Colibacillosis in caged layer hens: Characteristics of the disease and the aetiological agent. Avian Pathol. 2004, 33, 117–125. [Google Scholar] [CrossRef]
- Guabiraba, R.; Schouler, C. Avian colibacillosis: Still many black holes. FEMS Microbiol. Lett. 2015, 362, fnv118. [Google Scholar] [CrossRef]
- Dziva, F.; Stevens, M.P. Colibacillosis in poultry: Unravelling the molecular basis of virulence of avian pathogenic Escherichia coli in their natural hosts. Avian Pathol. 2008, 37, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Guiral, E.; Bosch, J.; Vila, J.; Soto, S.M. Antimicrobial resistance of Escherichia coli strains causing neonatal sepsis between 1998 and 2008. Chemotherapy 2012, 58, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Orskov, F.; Orskov, I. Escherichia coli serotyping and disease in man and animals. Can. J. Microbiol. 1992, 38, 699–704. [Google Scholar] [CrossRef] [PubMed]
- Meuskens, I.; Michalik, M.; Chauhan, N.; Linke, D.; Leo, J.C. A new strain collection for improved expression of outer membrane proteins. Front. Cell. Infect. Microbiol. 2017, 7, 464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Guan, Q.; Wang, X.; Teng, D.; Mao, R.; Yao, J.; Wang, J. Paving the way to construct a new vaccine against Escherichia coli from its recombinant outer membrane protein C via a murine model. Process Biochem. 2015, 50, 1194–1201. [Google Scholar] [CrossRef]
- Guan, Q.; Wang, X.; Wang, X.; Teng, D.; Mao, R.; Zhang, Y.; Wang, J. Recombinant outer membrane protein A induces a protective immune response against Escherichia coli infection in mice. Appl. Microbiol. Biotechnol. 2015, 99, 5451–5460. [Google Scholar] [CrossRef]
- Guan, Q.; Wang, X.; Wang, X.; Teng, D.; Wang, J. In silico analysis and recombinant expression of BamA protein as a universal vaccine against Escherichia coli in mice. Appl. Microbiol. Biotechnol. 2016, 100, 5089–5098. [Google Scholar] [CrossRef]
- Pore, D.; Chakrabarti, M.K. Outer membrane protein A (OmpA) from Shigella flexneri 2a: A promising subunit vaccine candidate. Vaccine 2013, 31, 3644–3650. [Google Scholar] [CrossRef]
- Li, Q.; Ren, J.; Xian, H.; Yin, C.; Yuan, Y.; Li, Y.; Ji, R.; Chu, C.; Qiao, Z.; Jiao, X. rOmpF and OMVs as efficient subunit vaccines against Salmonella enterica serovar Enteritidis infections in poultry farms. Vaccine 2020, 38, 7094–7099. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, S.; Zhao, X.; Yang, Y.; Li, B.; Zhu, F.; Zhu, R. Immune enhancement of Taishan Robinia pseudoacacia polysaccharide on recombinant Proteus mirabilis OmpA in chickens. Int. Immunopharmacol. 2014, 22, 236–241. [Google Scholar] [CrossRef]
- Zhang, B.Z.; Hu, D.; Dou, Y.; Xiong, L.; Wang, X.; Hu, J.; Xing, S.Z.; Li, W.; Cai, J.P.; Jin, M.; et al. Identification and evaluation of recombinant outer membrane proteins as vaccine candidates against Klebsiella pneumoniae. Front. Immunol. 2021, 12, 730116. [Google Scholar] [CrossRef]
- Duan, L.; Feng, J.; Peng, L.; Guo, S.; He, L.; Xiao, Y. Evaluation of an outer membrane protein as a vaccine against Edwardsiella anguillarum in Japanese eels (Anguilla japonica). Aquaculture 2019, 498, 143–150. [Google Scholar]
- Diao, J.; Li, L.; Fan, Y.; Wang, S.; Gai, C.; Wang, Y.; Yu, X.; Wang, X.; Xu, L.; Liu, H.; et al. Recombinant outer membrane protein C of Aeromonas salmonicida subsp. masoucida, a potential vaccine candidate for rainbow trout (Oncorhynchus mykiss). Microb. Pathog. 2020, 145, 104211. [Google Scholar] [CrossRef] [PubMed]
- Ewers, C.; Janssen, T.; Kiessling, S.; Philipp, H.C.; Wieler, L.H. Rapid detection of virulence-associated genes in avian pathogenic Escherichia coli by multiplex polymerase chain reaction. Avian Dis. 2005, 49, 269–273. [Google Scholar] [CrossRef]
- Maciel, J.F.; Matter, L.B.; Trindade, M.M.; Camillo, G.; Lovato, M.; de Ávila Botton, S.; Castagna de Vargas, A. Virulence factors and antimicrobial susceptibility profile of extraintestinal Escherichia coli isolated from an avian colisepticemia outbreak. Microb. Pathog. 2017, 103, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Nayeem, M.M.H.; Sobur, M.A.; Ievy, S.; Islam, M.A.; Rahman, S.; Kafi, M.A.; Ashour, H.M.; Rahman, M.T. Virulence determinants and multidrug resistance of Escherichia coli isolated from migratory birds. Antibiotics 2021, 10, 190. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Shi, Z.; Xia, Y.; Li, H.; Kou, Y.; Bao, Y.; Dai, J.; Lu, C. IbeB is involved in the invasion and pathogenicity of avian pathogenic Escherichia coli. Vet. Microbiol. 2012, 159, 411–419. [Google Scholar] [CrossRef] [PubMed]
- UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [CrossRef]
- Wang, X.; Teng, D.; Guan, Q.; Mao, R.; Hao, Y.; Wang, X.; Yao, J.; Wang, J. Escherichia coli outer membrane protein F (OmpF): An immunogenic protein induces cross-reactive antibodies against Escherichia coli and Shigella. AMB Express 2017, 7, 155. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Ramos, C.R.; Abreu, P.A.; Nascimento, A.L.; Ho, P.L. A high-copy T7 Escherichia coli expression vector for the production of recombinant proteins with a minimal N-terminal His-tagged fusion peptide. Braz. J. Med. Biol. Res. Rev. Bras. De Pesqui. Med. E Biol. 2004, 37, 1103–1109. [Google Scholar] [CrossRef] [Green Version]
- Saleem, M.; Moore, J.; Derrick, J.P. Expression, purification, and crystallization of neisserial outer membrane proteins. Methods Mol. Biol. 2012, 799, 91–106. [Google Scholar] [CrossRef]
- Chang, P.; Huang, L.; Huang, M.; Tian, S.; Yang, Z. Improvement and optimization of a T-cell-dependent antibody response (TDAR) method for BALB/c mice using keyhole limpet hemocyanin (KLH) as specific antigen. J. Immunotoxicol. 2019, 16, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Gong, R.; Guo, A.; Chen, H. Protective effect of ligand-binding domain of fibronectin-binding protein on mastitis induced by Staphylococcus aureus in mice. Vaccine 2010, 28, 4038–4044. [Google Scholar] [CrossRef] [PubMed]
- Hornbeck, P. Double-immunodiffusion assay for detecting specific antibodies. Curr. Protoc. Immunol. 2001. [Google Scholar] [CrossRef]
- Swamydas, M.; Luo, Y.; Dorf, M.E.; Lionakis, M.S. Isolation of mouse neutrophils. Curr. Protoc. Immunol. 2015, 110, 3.20.21–3.20.15. [Google Scholar] [CrossRef] [Green Version]
- Grund, M.E.; Soo, J.C.; Cote, C.K.; Berisio, R.; Lukomski, S. Thinking outside the bug: Targeting outer membrane proteins for Burkholderia vaccines. Cells 2021, 10, 495. [Google Scholar] [CrossRef]
- Schweizer, M.; Henning, U. Action of a major outer cell envelope membrane protein in conjugation of Escherichia coli K-12. J. Bacteriol. 1977, 129, 1651–1652. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, D.W.; Ricker, N.; Barbieri, N.L.; Allen, H.K.; Nolan, L.K.; Logue, C.M. Outer membrane protein A (OmpA) of extraintestinal pathogenic Escherichia coli. BMC Res. Notes 2020, 13, 51. [Google Scholar] [CrossRef]
- Gao, Z.; Ye, C.; Zhou, L.; Zhang, Y.; Ge, Y.; Chen, W.; Pan, J. Evaluation of the β-barrel outer membrane protein VP1243 as a candidate antigen for a cross-protective vaccine against Vibrio infections. Microb. Pathog. 2020, 147, 104419. [Google Scholar] [CrossRef]
- Peng, B.; Ye, J.Z.; Han, Y.; Zeng, L.; Zhang, J.Y.; Li, H. Identification of polyvalent protective immunogens from outer membrane proteins in Vibrio parahaemolyticus to protect fish against bacterial infection. Fish Shellfish. Immunol. 2016, 54, 204–210. [Google Scholar] [CrossRef]
- Aguilar, J.C.; Rodríguez, E.G. Vaccine adjuvants revisited. Vaccine 2007, 25, 3752–3762. [Google Scholar] [CrossRef] [PubMed]
- Fontes, J.A.; Barin, J.G.; Talor, M.V.; Stickel, N.; Schaub, J.; Rose, N.R.; Čiháková, D. Complete Freund’s adjuvant induces experimental autoimmune myocarditis by enhancing IL-6 production during initiation of the immune response. Immun. Inflamm. Dis. 2017, 5, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Gangaplara, A.; Massilamany, C.; Lasrado, N.; Steffen, D.; Reddy, J. Evidence for anti-viral effects of complete Freund’s adjuvant in the mouse model of Enterovirus infection. Vaccines 2020, 8, 364. [Google Scholar] [CrossRef] [PubMed]
- Hajizade, A.; Salmanian, A.H.; Amani, J.; Ebrahimi, F.; Arpanaei, A. EspA-loaded mesoporous silica nanoparticles can efficiently protect animal model against enterohaemorrhagic E. coli O157:H7. Artif. Cells Nanomed. Biotechnol. 2018, 46, S1067–S1075. [Google Scholar] [CrossRef] [Green Version]
- Samo, M.; Choudhary, N.R.; Riebe, K.J.; Shterev, I.; Staats, H.F.; Sempowski, G.D.; Leduc, I. Immunization with the Haemophilus ducreyi trimeric autotransporter adhesin DsrA with alum, CpG or imiquimod generates a persistent humoral immune response that recognizes the bacterial surface. Vaccine 2016, 34, 1193–1200. [Google Scholar] [CrossRef]
- Ronco, T.; Stegger, M.; Olsen, R.H.; Sekse, C.; Nordstoga, A.B.; Pohjanvirta, T.; Lilje, B.; Lyhs, U.; Andersen, P.S.; Pedersen, K. Spread of avian pathogenic Escherichia coli ST117 O78:H4 in Nordic broiler production. BMC Genom. 2017, 18, 13. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Maurer, J.J.; Hubert, S.; De Villena, J.F.; McDermott, P.F.; Meng, J.; Ayers, S.; English, L.; White, D.G. Antimicrobial susceptibility and molecular characterization of avian pathogenic Escherichia coli isolates. Vet. Microbiol. 2005, 107, 215–224. [Google Scholar] [CrossRef]
- Moulin-Schouleur, M.; Répérant, M.; Laurent, S.; Brée, A.; Mignon-Grasteau, S.; Germon, P.; Rasschaert, D.; Schouler, C. Extraintestinal pathogenic Escherichia coli strains of avian and human origin: Link between phylogenetic relationships and common virulence patterns. J. Clin. Microbiol. 2007, 45, 3366–3376. [Google Scholar] [CrossRef] [Green Version]
- Pyrski, M.; Rugowska, A.; Wierzbiński, K.R.; Kasprzyk, A.; Bogusiewicz, M.; Bociąg, P.; Samardakiewicz, S.; Czyż, M.; Kurpisz, M.; Pniewski, T. HBcAg produced in transgenic tobacco triggers Th1 and Th2 response when intramuscularly delivered. Vaccine 2017, 35, 5714–5721. [Google Scholar] [CrossRef]
- Hussain, R.; Dawood, G.; Abrar, N.; Toossi, Z.; Minai, A.; Dojki, M.; Ellner, J.J. Selective increases in antibody isotypes and immunoglobulin G subclass responses to secreted antigens in tuberculosis patients and healthy household contacts of the patients. Clin. Diagn. Lab. Immunol. 1995, 2, 726–732. [Google Scholar] [CrossRef] [Green Version]
- Keyt, B.A.; Baliga, R.; Sinclair, A.M.; Carroll, S.F.; Peterson, M.S. Structure, function, and therapeutic use of IgM antibodies. Antibodies 2020, 9, 53. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).