
Modeling the Impact of Management Changes on the Infection Dynamics of Extended-Spectrum Beta-Lactamase-Producing Escherichia coli in the Broiler Production


Abstract
1. Introduction
2. Materials and Methods
2.1. Mathematical Model
- Intake: feed, water and bedding material (including bacteria, Ning) are ingested.
- Growth: ESBL-producing E. coli grow in the chick’s intestinal tract (Nint).
- Excretion: Intestinal ESBLs were excreted with the feces (Nfec).
- All chickens share the same pen. The pen area is calculated for each scenario based on the chosen values of the parameters “total number of chickens”, “target weight” and “stocking density”.
- The excreted bacteria are immediately mixed evenly in the total amount of available litter. Each chick ingests the same amount of litter and, thus, bacteria.
- All the bacteria considered are resistant. If there is even a single resistant bacterium in the gastrointestinal tract of a chick, it is considered to be ESBL-positive.
- Once a viable colony-forming unit enters the intestinal tract, the growth rate, once assigned, does not change for these bacteria in this chick.
- Infection occurs only via positive chicks, artificial oral inoculation or ingestion of contaminated bedding material; we did not model a transmission via bird-to-bird-contact or air.
- The amount of excreted feces per chick per day is calculated from the difference between ingested feed and water (minus the amount of exhaled water) and daily weight gain. The loss of feed (energy) required for metabolism was not calculated and subtracted.
2.2. Modeling the Processes
2.2.1. Uptake of ESBL-Producing E. coli
2.2.2. Growth of ESBL-Producing E. coli in the Intestine
2.2.3. Excretion of ESBL-Producing E. coli into the Litter
2.3. Adaptation to Experimental Data
2.4. Modeling Different Ways of Infection
- The day-old chicks are positive for ESBL-producing E. coli.
- The pen is contaminated, and the chickens peck and ingest contaminated litter. Contamination may come from internal (e.g., previous positive flocks) or external sources (e.g., environment and rodents); the origin is not considered in our model.
2.5. Modeling Changes in Management Practice
- Amount of litter at the beginning of the fattening period: 3000, 6000 and 9000 g/m2, instead of 1000 g/m2;
- Breed Rowan x Ranger with a feeding duration of 47 days, daily feed intake and daily body weight gain according to the corresponding Aviagen manual [45] and a target weight of 1.911 kg on day 47, instead of Ross 308 with a feeding duration of 36 days, a target weight of 2.332 kg and daily intake and daily body weight gain according to the corresponding Aviagen manual [44];
- Stocking density: 20 or 25 kg/m2, instead of 39 kg/m2;
- Different combinations of the previously mentioned measures;
- Products with impact on the microbiota, such as prebiotics, probiotics, synbiotics, feed additives or competitive exclusion, were expected to reduce the prevalence of ESBL-producing E. coli [23,25]. We modeled the effect of these products with a lower maximum growth rate of ESBL-producing E. coli in the intestine—104 instead of 105.
2.6. Model Runs
3. Results
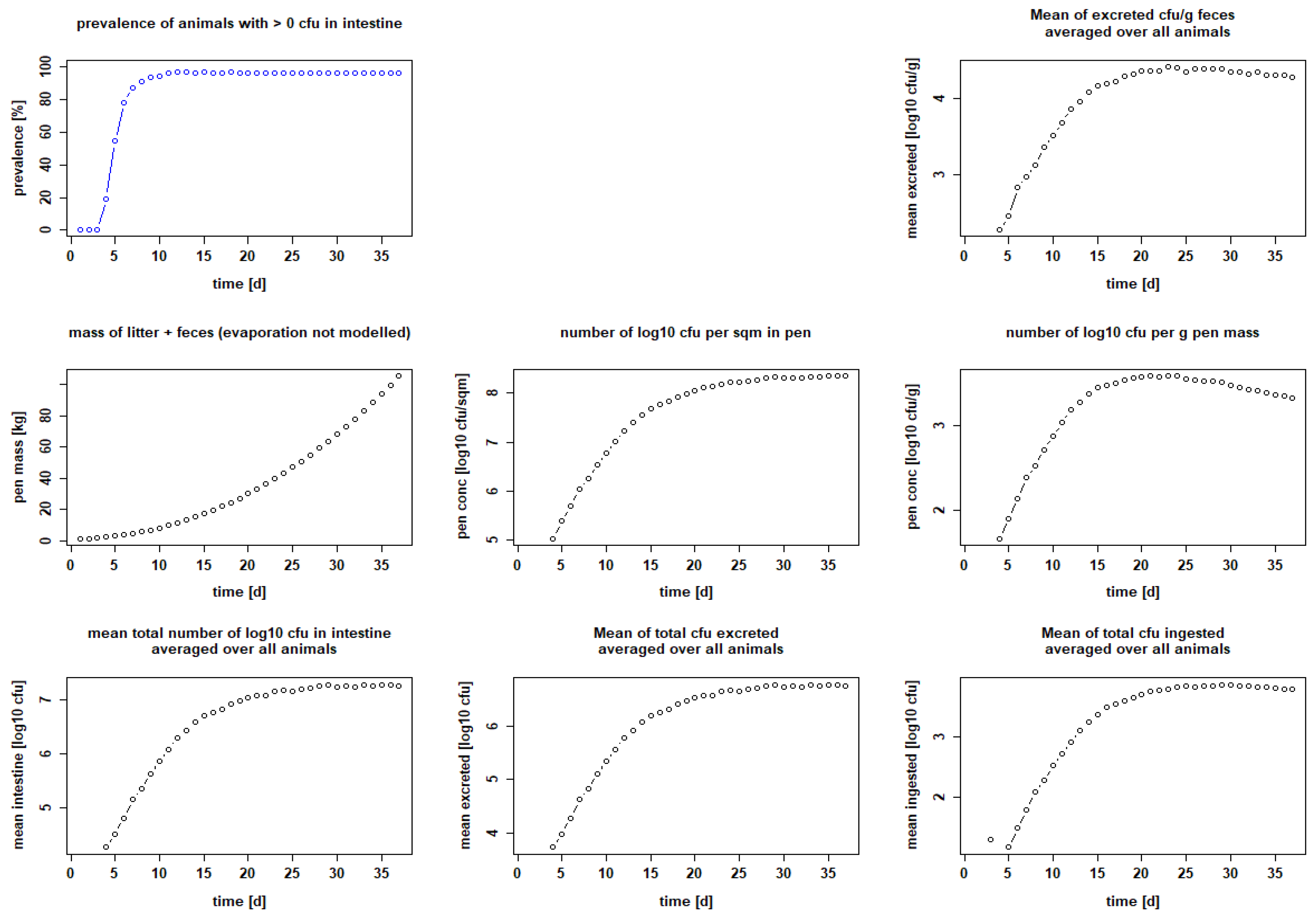
3.1. Adaptation to Experimental Data
3.2. Modeling Different Ways of Infection
3.3. Calculated Effects of Management Measures at Farm Level
3.3.1. Increasing the Amount of Litter per Square Meter
3.3.2. A Slow-Growing Breed
3.3.3. Lower Stocking Densities
3.3.4. Combination of Measures
3.3.5. Products with Impact on the Microbiota
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wielders, C.C.H.; van Hoek, A.; Hengeveld, P.D.; Veenman, C.; Dierikx, C.M.; Zomer, T.P.; Smit, L.A.M.; van der Hoek, W.; Heederik, D.J.; de Greeff, S.C.; et al. Extended-Spectrum Beta-Lactamase- and PAmpC-Producing Enterobacteriaceae among the General Population in a Livestock-Dense Area. Clin. Microbiol. Infect. 2017, 23, 120.e1–120.e8. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, M.; Irrgang, A.; Roschanski, N.; Brenner Michael, G.; Hamprecht, A.; Rieber, H.; Käsbohrer, A.; Schwarz, S.; Rösler, U.; Kreienbrock, L.; et al. Whole Genome Analyses of CMY-2-Producing Escherichia Coli Isolates from Humans, Animals and Food in Germany. BMC Genom. 2018, 19, 601. [Google Scholar] [CrossRef]
- Dierikx, C.M.; van Duijkeren, E.; Schoormans, A.H.; van Essen-Zandbergen, A.; Veldman, K.; Kant, A.; Huijsdens, X.W.; van der Zwaluw, K.; Wagenaar, J.A.; Mevius, D.J. Occurrence and Characteristics of Extended-Spectrum-Beta-Lactamase- and AmpC-Producing Clinical Isolates Derived from Companion Animals and Horses. J. Antimicrob. Chemother. 2012, 67, 1368–1374. [Google Scholar] [CrossRef] [PubMed]
- Dohmen, W.; Schmitt, H.; Bonten, M.; Heederik, D. Air Exposure as a Possible Route for ESBL in Pig Farmers. Environ. Res. 2017, 155, 359–364. [Google Scholar] [CrossRef]
- Hartmann, A.; Locatelli, A.; Amoureux, L.; Depret, G.; Jolivet, C.; Gueneau, E.; Neuwirth, C. Occurrence of CTX-M Producing Escherichia Coli in Soils, Cattle, and Farm Environment in France (Burgundy Region). Front. Microbiol. 2012, 3, 83. [Google Scholar] [CrossRef]
- Van Hoek, A.; Dierikx, C.; Bosch, T.; Schouls, L.; van Duijkeren, E.; Visser, M. Transmission of ESBL-Producing Escherichia Coli between Broilers and Humans on Broiler Farms. J. Antimicrob. Chemother. 2020, 75, 543–549. [Google Scholar] [CrossRef]
- Fischer, J.; Hille, K.; Ruddat, I.; Mellmann, A.; Kock, R.; Kreienbrock, L. Simultaneous Occurrence of MRSA and ESBL-Producing Enterobacteriaceae on Pig Farms and in Nasal and Stool Samples from Farmers. Vet. Microbiol. 2017, 200, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Huijbers, P.M.C.; van Hoek, A.H.A.M.; Graat, E.A.M.; Haenen, A.P.J.; Florijn, A.; Hengeveld, P.D.; van Duijkeren, E. Methicillin-Resistant Staphylococcus Aureus and Extended-Spectrum and AmpC β-Lactamase-Producing Escherichia Coli in Broilers and in People Living and/or Working on Organic Broiler Farms. Vet. Microbiol. 2015, 176, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Huijbers, P.M.; Blaak, H.; de Jong, M.C.; Graat, E.A.; Vandenbroucke-Grauls, C.M.; de Roda Husman, A.M. Role of the Environment in the Transmission of Antimicrobial Resistance to Humans: A Review. Environ. Sci. Technol. 2015, 49, 11993–12004. [Google Scholar] [CrossRef]
- Daehre, K.; Projahn, M.; Semmler, T.; Roesler, U.; Friese, A. Extended-Spectrum Beta-Lactamase-/AmpC Beta-Lactamase-Producing Enterobacteriaceae in Broiler Farms: Transmission Dynamics at Farm Level. Microb. Drug Resist. 2018, 24, 511–518. [Google Scholar] [CrossRef]
- Massella, E.; Reid, C.J.; Cummins, M.L.; Anantanawat, K.; Zingali, T.; Serraino, A.; Piva, S.; Giacometti, F.; Djordjevic, S.P. Snapshot Study of Whole Genome Sequences of Escherichia Coli from Healthy Companion Animals, Livestock, Wildlife, Humans and Food in Italy. Antibiotics 2020, 9, 782. [Google Scholar] [CrossRef]
- Leverstein-van Hall, M.A.; Dierikx, C.M.; Stuart, J.C.; Voets, G.M.; van den Munckhof, M.P.; van Essen-Zandbergen, A.; Platteel, T.; Fluit, A.C.; van de Sande-Bruinsma, N.; Scharinga, J.; et al. Dutch Patients, Retail Chicken Meat and Poultry Share the Same ESBL Genes, Plasmids and Strains. Clin. Microbiol. Infect. 2011, 17, 873–880. [Google Scholar] [CrossRef]
- World Health Organization. Global Action Plan on Antimicrobial Resistance. Available online: https://www.who.int/publications/i/item/9789241509763 (accessed on 14 April 2022).
- European Food Safety Authority (EFSA) The European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2019–2020. EFSA J. 2022, 20, e7209. (accessed on 14 April 2022). [CrossRef]
- Dahms, C.; Hubner, N.O.; Kossow, A.; Mellmann, A.; Dittmann, K.; Kramer, A. Occurrence of ESBL-Producing Escherichia Coli in Livestock and Farm Workers in Mecklenburg-Western Pomerania, Germany. PLoS ONE 2015, 10, e0143326. [Google Scholar] [CrossRef]
- Kaesbohrer, A.; Bakran-Lebl, K.; Irrgang, A.; Fischer, J.; Kämpf, P.; Schiffmann, A.; Werckenthin, C.; Busch, M.; Kreienbrock, L.; Hille, K. Diversity in Prevalence and Characteristics of ESBL/PAmpC Producing E. Coli in Food in Germany. Vet. Microbiol. 2019, 233, 52–60. [Google Scholar] [CrossRef]
- Clemente, L.; Leão, C.; Moura, L.; Albuquerque, T.; Amaro, A. Prevalence and Characterization of ESBL/AmpC Producing Escherichia Coli from Fresh Meat in Portugal. Antibiotics 2021, 10, 1333. [Google Scholar] [CrossRef]
- Motola, G.; Hafez, H.M.; Bruggemann-Schwarze, S. Efficacy of Six Disinfection Methods against Extended-Spectrum Beta-Lactamase (ESBL) Producing E. Coli on Eggshells in Vitro. PLoS ONE 2020, 15, e0238860. [Google Scholar] [CrossRef]
- Tebrun, W.; Motola, G.; Hafez, M.H.; Bachmeier, J.; Schmidt, V.; Renfert, K.; Reichelt, C.; Bruggemann-Schwarze, S.; Pees, M. Preliminary Study: Health and Performance Assessment in Broiler Chicks Following Application of Six Different Hatching Egg Disinfection Protocols. PLoS ONE 2020, 15, e0232825. [Google Scholar] [CrossRef]
- Projahn, M.; Daehre, K.; Roesler, U.; Friese, A. Extended-Spectrum-Beta-Lactamase- and Plasmid-Encoded Cephamycinase-Producing Enterobacteria in the Broiler Hatchery as a Potential Mode of Pseudo-Vertical Transmission. Appl. Environ. Microbiol. 2017, 83, e02364-16. [Google Scholar] [CrossRef]
- Robe, C.; Daehre, K.; Merle, R.; Friese, A.; Guenther, S.; Roesler, U. Impact of Different Management Measures on the Colonization of Broiler Chickens with ESBL- and PAmpC-Producing Escherichia Coli in an Experimental Seeder-Bird Model. PLoS ONE 2021, 16, e0245224. [Google Scholar] [CrossRef]
- Saliu, E.-M.; Ren, H.; Goodarzi Boroojeni, F.; Zentek, J.; Vahjen, W. The Impact of Direct-Fed Microbials and Phytogenic Feed Additives on Prevalence and Transfer of Extended-Spectrum Beta-Lactamase Genes in Broiler Chicken. Microorganisms 2020, 8, 322. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Vahjen, W.; Dadi, T.; Saliu, E.M.; Boroojeni, F.G.; Zentek, J. Synergistic Effects of Probiotics and Phytobiotics on the Intestinal Microbiota in Young Broiler Chicken. Microorganisms 2019, 7, 684. [Google Scholar] [CrossRef] [PubMed]
- Methner, U.; Friese, A.; Rosler, U. Competitive Exclusion: A Tool to Combat Extended-Spectrum Beta-Lactamase-Producing Escherichia Coli Strains in Chickens. Res. Vet. Sci. 2019, 123, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Methner, U.; Rosler, U. Efficacy of a Competitive Exclusion Culture against Extended-Spectrum Beta-Lactamase-Producing Escherichia Coli Strains in Broilers Using a Seeder Bird Model. BMC Vet. Res. 2020, 16, 143. [Google Scholar] [CrossRef]
- Projahn, M.; Sachsenroeder, J.; Correia-Carreira, G.; Becker, E.; Martin, A.; Thomas, C.; Hobe, C.; Reich, F.; Robe, C.; Roesler, U.; et al. Impact of On-Farm Interventions against CTX-Resistant Escherichia Coli on the Contamination of Carcasses before and during an Experimental Slaughter. Antibiotics 2021, 10, 228. [Google Scholar] [CrossRef]
- Thomas, C.; Schonknecht, A.; Puning, C.; Alter, T.; Martin, A.; Bandick, N. Effect of Peracetic Acid Solutions and Lactic Acid on Microorganisms in On-Line Reprocessing Systems for Chicken Slaughter Plants. J. Food Prot. 2020, 83, 615–620. [Google Scholar] [CrossRef]
- Reich, F.; Atanassova, V.; Klein, G. Extended-Spectrum Beta-Lactamase- and AmpC-Producing Enterobacteria in Healthy Broiler Chickens, Germany. Emerg. Infect. Dis. 2013, 19, 1253–1259. [Google Scholar] [CrossRef]
- Von Tippelskirch, P.; Golz, G.; Projahn, M.; Daehre, K.; Friese, A.; Roesler, U.; Alter, T.; Orquera, S. Prevalence and Quantitative Analysis of ESBL and AmpC Beta-Lactamase Producing Enterobacteriaceae in Broiler Chicken during Slaughter in Germany. Int. J. Food Microbiol. 2018, 281, 82–89. [Google Scholar] [CrossRef]
- Thomas, C.; Idler, C.; Ammon, C.; Herrmann, C.; Amon, T. Inactivation of ESBL-/AmpC-Producing Escherichia Coli during Mesophilic and Thermophilic Anaerobic Digestion of Chicken Manure. Waste Manag. 2019, 84, 74–82. [Google Scholar] [CrossRef]
- Siller, P.; Daehre, K.; Thiel, N.; Nubel, U.; Roesler, U. Impact of Short-Term Storage on the Quantity of Extended-Spectrum Beta-Lactamase-Producing Escherichia Coli in Broiler Litter under Practical Conditions. Poult. Sci. 2020, 99, 2125–2135. [Google Scholar] [CrossRef]
- Becker, E.; Projahn, M.; Burow, E.; Käsbohrer, A. Are There Effective Intervention Measures in Broiler Production against the ESBL/AmpC Producer Escherichia Coli? Pathogens 2021, 10, 608. [Google Scholar] [CrossRef]
- Rawson, T.; Dawkins, M.S.; Bonsall, M.B. A Mathematical Model of Campylobacter Dynamics Within a Broiler Flock. Front. Microbiol. 2019, 10, 1940. [Google Scholar] [CrossRef]
- Bastard, J.; Haenni, M.; Gay, E.; Glaser, P.; Madec, J.-Y.; Temime, L.; Opatowski, L. Drivers of ESBL-Producing Escherichia Coli Dynamics in Calf Fattening Farms: A Modelling Study. One Health 2021, 12, 100238. [Google Scholar] [CrossRef]
- Huijbers, P.M.C.; Graat, E.A.M.; van Hoek, A.; Veenman, C.; de Jong, M.C.M.; van Duijkeren, E. Transmission Dynamics of Extended-Spectrum Beta-Lactamase and AmpC Beta-Lactamase-Producing Escherichia Coli in a Broiler Flock without Antibiotic Use. Prev. Vet. Med. 2016, 131, 12–19. [Google Scholar] [CrossRef]
- Plaza Rodriguez, C.; Correia Carreira, G.; Kasbohrer, A. A Probabilistic Transmission Model for the Spread of Extended-Spectrum-Beta-Lactamase and AmpC-Beta-Lactamase-Producing Escherichia Coli in the Broiler Production Chain. Risk Anal. 2018, 38, 2659–2682. [Google Scholar] [CrossRef]
- Correia-Carreira, G.; Langkabel, N.; Projahn, M.; Becker, E.; Käsbohrer, A. Modelling of Interventions for Reducing External Enterobacteriaceae Contamination of Broiler Carcasses during Processing. Risk Anal. 2022; submitted. [Google Scholar]
- Birkegard, A.C.; Halasa, T.; Toft, N.; Folkesson, A.; Graesboll, K. Send More Data: A Systematic Review of Mathematical Models of Antimicrobial Resistance. Antimicrob. Resist. Infect. Control 2018, 7, 117. [Google Scholar] [CrossRef]
- Hill, A.; Simons, R.; Ramnial, V.; Tennant, J.; Denman, S.; Cheney, T.; Snary, E.; Swart, A.; Evers, E.; Nauta, M.; et al. Quantitative Microbiological Risk Assessment on Salmonella in Slaughter and Breeder Pigs: Final Report. EFSA Support. Publ. 2010, 7, 46E. [Google Scholar] [CrossRef]
- Græsbøll, K.; Nielsen, S.S.; Toft, N.; Christiansen, L.E. How Fitness Reduced, Antimicrobial Resistant Bacteria Survive and Spread: A Multiple Pig—Multiple Bacterial Strain Model. PLoS ONE 2014, 9, e100458. [Google Scholar] [CrossRef]
- R Development Core Team. A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2008. Available online: https://www.R-project.org (accessed on 14 April 2022).
- Malone, G.W.; Chaloupka, G.W.; Saylor, W.W. Influence of Litter Type and Size on Broiler Performance. Poult. Sci. 1983, 62, 1741–1746. [Google Scholar] [CrossRef]
- Dame-Korevaar, A.; Fischer, E.A.J.; van der Goot, J.; Velkers, F.; Ceccarelli, D.; Mevius, D.; Stegeman, A. Early Life Supply of Competitive Exclusion Products Reduces Colonization of Extended Spectrum Beta-Lactamase-Producing Escherichia Coli in Broilers. Poult. Sci. 2020, 99, 4052–4064. [Google Scholar] [CrossRef]
- Aviagen. ROSS 308/ROSS 308 FF BROILER: Performance Objectives. Available online: http://en.aviagen.com/assets/Tech_Center/Ross_Broiler/Ross308-308FF-BroilerPO2019-EN.pdf (accessed on 14 April 2022).
- Aviagen. Managing the Rowan Ranger. Available online: http://en.aviagen.com/assets/Tech_Center/Rowan_Range/RowanRangerManagement2016EN.pdf (accessed on 14 April 2022).
- Watkins, S.; Tabler, G.T. Avian Advice, Summer 2009. pp. 8–12. Available online: https://scholarworks.uark.edu/cgi/viewcontent.cgi?article=1029&context=avian-advice (accessed on 14 April 2022).
- Kirkpatrick, K.F. Aviagen Brief—Water Quality; Aviagen: Huntsville, AL, USA, 2008; Available online: https://eu.aviagen.com/assets/Tech_Center/Broiler_Breeder_Tech_Articles/English/AviagenBrief_WaterQuality_Feb08.pdf (accessed on 14 April 2022).
- Muschkowitz, C. Impedimetrische Bestimmung von Generationszeiten Bei Stämmen Lebensmittelhygienisch Relevanter Enterobacteriaceaespezies Und -Genera. Ph.D. Thesis, Freie Universität, Berlin, Germany, 1997. [Google Scholar] [CrossRef]
- Robe, C.; Blasse, A.; Merle, R.; Friese, A.; Roesler, U.; Guenther, S. Low Dose Colonization of Broiler Chickens With ESBL-/AmpC-Producing Escherichia Coli in a Seeder-Bird Model Independent of Antimicrobial Selection Pressure. Front. Microbiol. 2019, 10, 2124. [Google Scholar] [CrossRef]
- Dame-Korevaar, A.; Fischer, E.A.J.; van der Goot, J.; Velkers, F.; van den Broek, J.; Veldman, K.; Ceccarelli, D.; Mevius, D.; Stegeman, A. Effect of Challenge Dose of Plasmid-Mediated Extended-Spectrum Beta-Lactamase and AmpC Beta-Lactamase Producing Escherichia Coli on Time-until-Colonization and Level of Excretion in Young Broilers. Vet. Microbiol. 2019, 239, 108446. [Google Scholar] [CrossRef]
- Rougiere, N.; Carre, B. Comparison of Gastrointestinal Transit Times between Chickens from D+ and D- Genetic Lines Selected for Divergent Digestion Efficiency. Animal 2010, 4, 1861–1872. [Google Scholar] [CrossRef]
- Svihus, B.; Itani, K. Intestinal Passage and Its Relation to Digestive Processes. J. Appl. Poult. Res. 2019, 28, 546–555. [Google Scholar] [CrossRef]
- Ravindran, V. Feed Enzymes: The Science, Practice, and Metabolic Realities. J. Appl. Poult. Res. 2013, 22, 628–636. [Google Scholar] [CrossRef]
- Blaak, H.; van Hoek, A.H.; Hamidjaja, R.A.; van der Plaats, R.Q.; Kerkhof-de Heer, L.; de Roda Husman, A.M.; Schets, F.M. Distribution, Numbers, and Diversity of ESBL-Producing E. Coli in the Poultry Farm Environment. PLoS ONE 2015, 10, e0135402. [Google Scholar] [CrossRef]
- Ebenhöh, W. Zur Bedeutung Der Mathematischen Modellierung. Available online: https://wiki.hhu.de/download/attachments/157483130/BedeutungDerMathematischenModellierung_WolfgangEbenhoeh.pdf?version=1&modificationDate=1584892377000&api=v2 (accessed on 14 April 2022).
- Opatowski, L.; Guillemot, D.; Boelle, P.Y.; Temime, L. Contribution of Mathematical Modeling to the Fight against Bacterial Antibiotic Resistance. Curr. Opin. Infect. Dis. 2011, 24, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Gänzle, M. Prädiktive Mikrobiologie. Available online: https://roempp.thieme.de/lexicon/RD-16-03896 (accessed on 14 April 2022).
- Jurburg, S.D.; Brouwer, M.S.M.; Ceccarelli, D.; van der Goot, J.; Jansman, A.J.M.; Bossers, A. Patterns of Community Assembly in the Developing Chicken Microbiome Reveal Rapid Primary Succession. Microbiologyopen 2019, 8, e00821. [Google Scholar] [CrossRef] [PubMed]
- Schokker, D.; Veninga, G.; Vastenhouw, S.A.; Bossers, A.; de Bree, F.M.; Kaal-Lansbergen, L.M.; Rebel, J.M.; Smits, M.A. Early Life Microbial Colonization of the Gut and Intestinal Development Differ between Genetically Divergent Broiler Lines. BMC Genom. 2015, 16, 418. [Google Scholar] [CrossRef] [PubMed]
- Laube, H.; Friese, A.; von Salviati, C.; Guerra, B.; Kasbohrer, A.; Kreienbrock, L.; Roesler, U. Longitudinal Monitoring of Extended-Spectrum-Beta-Lactamase/AmpC-Producing Escherichia Coli at German Broiler Chicken Fattening Farms. Appl. Environ. Microbiol. 2013, 79, 4815–4820. [Google Scholar] [CrossRef]
- Dorado-García, A.; Smid, J.H.; van Pelt, W.; Bonten, M.J.M.; Fluit, A.C.; van den Bunt, G.; Wagenaar, J.A.; Hordijk, J.; Dierikx, C.M.; Veldman, K.T.; et al. Molecular Relatedness of ESBL/AmpC-Producing Escherichia Coli from Humans, Animals, Food and the Environment: A Pooled Analysis. J. Antimicrob. Chemother. 2018, 73, 339–347. [Google Scholar] [CrossRef]
- Huijbers, P.M.; Graat, E.A.; Haenen, A.P.; van Santen, M.G.; van Essen-Zandbergen, A.; Mevius, D.J.; van Duijkeren, E.; van Hoek, A.H. Extended-Spectrum and AmpC Beta-Lactamase-Producing Escherichia Coli in Broilers and People Living and/or Working on Broiler Farms: Prevalence, Risk Factors and Molecular Characteristics. J. Antimicrob. Chemother. 2014, 69, 2669–2675. [Google Scholar] [CrossRef]
- Guenther, S.; Bethe, A.; Fruth, A.; Semmler, T.; Ulrich, R.G.; Wieler, L.H.; Ewers, C. Frequent Combination of Antimicrobial Multiresistance and Extraintestinal Pathogenicity in Escherichia Coli Isolates from Urban Rats (Rattus Norvegicus) in Berlin, Germany. PLoS ONE 2012, 7, e50331. [Google Scholar] [CrossRef]
- Ewers, C.; Bethe, A.; Wieler, L.H.; Guenther, S.; Stamm, I.; Kopp, P.A.; Grobbel, M. Companion Animals: A Relevant Source of Extended-Spectrum Beta-Lactamase-Producing Fluoroquinolone-Resistant Citrobacter Freundii. Int. J. Antimicrob. Agents 2011, 37, 86–87. [Google Scholar] [CrossRef]
- Delpont, M.; Guinat, C.; Guérin, J.-L.; Le Leu, E.; Vaillancourt, J.-P.; Paul, M.C. Biosecurity Measures in French Poultry Farms Are Associated with Farm Type and Location. Prev. Vet. Med. 2021, 195, 105466. [Google Scholar] [CrossRef]
- Luyckx, K.Y.; Van Weyenberg, S.; Dewulf, J.; Herman, L.; Zoons, J.; Vervaet, E.; Heyndrickx, M.; De Reu, K. On-Farm Comparisons of Different Cleaning Protocols in Broiler Houses. Poult. Sci. 2015, 94, 1986–1993. [Google Scholar] [CrossRef]
- Mueller-Doblies, D.; Carrique-Mas, J.J.; Sayers, A.R.; Davies, R.H. A Comparison of the Efficacy of Different Disinfection Methods in Eliminating Salmonella Contamination from Turkey Houses. J. Appl. Microbiol. 2010, 109, 471–479. [Google Scholar] [CrossRef]
- Bolan, N.S.; Szogi, A.A.; Chuasavathi, T.; Seshadri, B.; Rothrock, M.J.; Panneerselvam, P. Uses and Management of Poultry Litter. Worlds Poult. Sci. J. 2019, 66, 673–698. [Google Scholar] [CrossRef]
- Paul, S.S.; Chatterjee, R.N.; Raju, M.V.L.N.; Prakash, B.; Rama Rao, S.V.; Yadav, S.P.; Kannan, A. Gut Microbial Composition Differs Extensively among Indian Native Chicken Breeds Originated in Different Geographical Locations and a Commercial Broiler Line, but Breed-Specific, as Well as Across-Breed Core Microbiomes, Are Found. Microorganisms 2021, 9, 391. [Google Scholar] [CrossRef]
- Ramírez, G.A.; Richardson, E.; Clark, J.; Keshri, J.; Drechsler, Y.; Berrang, M.E.; Meinersmann, R.J.; Cox, N.A.; Oakley, B.B. Broiler Chickens and Early Life Programming: Microbiome Transplant-Induced Cecal Community Dynamics and Phenotypic Effects. PLoS ONE 2020, 15, e0242108. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, T.; Zhang, N.; Li, J.; Wang, Y.; Kulyar, M.F.; Han, Z.; Li, Y. Effect of Stocking Density and Age on Physiological Performance and Dynamic Gut Bacterial and Fungal Communities in Langya Hens. Microb. Cell Factories 2021, 20, 218. [Google Scholar] [CrossRef]
- Guardia, S.; Konsak, B.; Combes, S.; Levenez, F.; Cauquil, L.; Guillot, J.F.; Moreau-Vauzelle, C.; Lessire, M.; Juin, H.; Gabriel, I. Effects of Stocking Density on the Growth Performance and Digestive Microbiota of Broiler Chickens. Poult. Sci. 2011, 90, 1878–1889. [Google Scholar] [CrossRef] [PubMed]
- Torok, V.A.; Hughes, R.J.; Ophel-Keller, K.; Ali, M.; Macalpine, R. Influence of Different Litter Materials on Cecal Microbiota Colonization in Broiler Chickens. Poult. Sci. 2009, 88, 2474–2481. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Lopez, P.; Sarmiento-Franco, L.A.; Santos-Ricalde, R. Effect of Stocking Density on Performance, Infection by Eimeria Spp., Intestinal Lesions and Foot Pad Injuries in Broilers with Outdoor Access under Tropical Conditions. Br. Poult. Sci. 2021, 63, 108–114. [Google Scholar] [CrossRef] [PubMed]
- McKeith, A.; Loper, M.; Tarrant, K.J. Research Note: Stocking Density Effects on Production Qualities of Broilers Raised without the Use of Antibiotics. Poult. Sci. 2020, 99, 698–701. [Google Scholar] [CrossRef]
- Bokkers, E.A.; de Boer, I.J. Economic, Ecological, and Social Performance of Conventional and Organic Broiler Production in the Netherlands. Br. Poult. Sci. 2009, 50, 546–557. [Google Scholar] [CrossRef]
- Simitzis, P.E.; Kalogeraki, E.; Goliomytis, M.; Charismiadou, M.A.; Triantaphyllopoulos, K.; Ayoutanti, A.; Niforou, K.; Hager-Theodorides, A.L.; Deligeorgis, S.G. Impact of Stocking Density on Broiler Growth Performance, Meat Characteristics, Behavioural Components and Indicators of Physiological and Oxidative Stress. Br. Poult. Sci. 2012, 53, 721–730. [Google Scholar] [CrossRef]
- Dame-Korevaar, A.; Kers, J.G.; van der Goot, J.; Velkers, F.C.; Ceccarelli, D.; Mevius, D.J.; Stegeman, A.; Fischer, E.A.J. Competitive Exclusion Prevents Colonization and Compartmentalization Reduces Transmission of ESBL-Producing Escherichia Coli in Broilers. Front. Microbiol. 2020, 11, 566619. [Google Scholar] [CrossRef]
- Ceccarelli, D.; van Essen-Zandbergen, A.; Smid, B.; Veldman, K.T.; Boender, G.J.; Fischer, E.A.J.; Mevius, D.J.; van der Goot, J.A. Competitive Exclusion Reduces Transmission and Excretion of Extended-Spectrum-Beta-Lactamase-Producing Escherichia Coli in Broilers. Appl. Environ. Microbiol. 2017, 83, e03439-16. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Definition | Value and Unit | Reference |
|---|---|---|---|
| Feeding duration | Total duration of the fattening period | 36 days (Ross 308) 47 days (Rowan x Ranger) | [21,43] |
| Feed intake | Daily feed intake for all days of the defined fattening period according to the information provided by the breeder | According to manual [g] | Aviagen manual Ross 308 [44], Table “As-Hatched Performance”, days 2–37; Aviagen manual Rowan Ranger [45], Table “As-Hatched”, days 2–48 |
| Water intake | Daily intake of water for all days of the defined fattening period according to the information provided by the breeder | According to manual [g] | [46]; 70% of the quantities, see manual Aviagen Brief [47] |
| king | Litter uptake and intake of ESBLs with contaminated litter | Factor 0.014 (= 1.38% of the amount of feed intake [g]) | Estimated [42] |
| Litter amount | Litter quantity per square meter at the beginning of the fattening period | 1000 g/m2 | Commercial standard in Europe |
| Stocking density | Maximum number of chickens per square meter (refers to the target weight) | 39 kg/m2 | (Tierschutz-Nutztierhaltungs-verordnung—TierSchNutztV), 2001, § 19, Absatz 7, (3) |
| kvia | Random factor for bacteria which do remain capable of reproduction when ingested | 0–0.5 | Assumed |
| kgro | Random growth rate for ESBL-producing E. coli in the chicken’s intestine | 100–105 | Maximum estimated [48] |
| Carrying capacity | Maximum number of CFU ESBL-producing E. coli in colon | 8 log10/g | [49,50] |
| Transition factor | The factor by which the total amount in the chick within 24 h, 1 day, has to be divided to obtain the content of the intestine (in grams) | 4.85 | Transition time estimated [51,52,53] |
| kexc | Factor of excreted bacteria | 0.3 | Assumed |
| ksur | Daily survival rate of ESBL in litter | 0.5 | Estimated [31,54] |
| Target weight | Weight of the chickens at the end of the fattening period (see “feeding duration” above) | 2.332 kg (Ross 308) 1.911 kg (Rowan x Ranger) | Aviagen manual Ross308-308FF-Broiler [44], Table “As-Hatched Performance”. Aviagen manual Rowan Ranger [45] (Appendix 3, Table 1), “As-hatched broiler performance”. |
| Seeder–Sentinel Ratio | No. of Chicks | Feeding Duration [Days] | Inoculation | Prevalence * [%] | ESBL-Producing E. coli in Feces * [log10 CFU/g] | ESBL-Producing E. coli in Litter * [log10 CFU/g] | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Amount | On Day | Maximum | On Day | Max. | On Day | Max. | On Day | |||
| 1:1 | 10 | 21 | 102 CFU | 1 | 99.2 ± 4.42 | 7 | 4.51 ± 4.42 | 21 | 3.71 ± 3.66 | 17 |
| 1:5 | 20 | 36 | 102 CFU | 3 | 97.15 ± 14.67 | 15 | 4.41 ± 4.26 | 23 | 3.6 ± 3.43 | 23 |
| Scenario | Feeding Duration [Days] | Prevalence * [%] | ESBL-Producing E. coli in Feces * [log10 CFU/g] | ESBL-Producing E. coli in Litter * [log10 CFU/g] | |||
|---|---|---|---|---|---|---|---|
| Maximum | On Day | Maximum | On Day | Maximum | On Day | ||
| Reference groups | |||||||
| Chicks positive | 36 | 99.38 ± 1.98 | 6 | 4.63 ± 4.08 | 20 | 3.9 ± 3.47 | 18 |
| Pen positive | 36 | 99.79 ± 0.61 | 7 | 4.51 ± 3.85 | 26 | 3.68 ± 3.36 | 20 |
| Management measure 3000 g litter/m2 | |||||||
| Chicks positive | 36 | 98.53 ± 6.24 | 11 | 4.19 ± 3.92 | 34 | 3.25 ± 3.04 | 30 |
| Pen positive | 36 | 32.46 ± 4.42 | 3 | 1.48 ± 1.14 | 3 | 1.47 ± Inf | 1 |
| Management measure 6000 g litter/m2 | |||||||
| Chicks positive | 36 | 57.23 ± 25.11 | 7 | 2.63 ± 2.37 | 2 | 1.13 ± 0.77 | 3 |
| Pen positive | 36 | 0 | 0 | 0 | 0 | 1.18 ± Inf | 1 |
| Management measure 9000 g litter/m2 | |||||||
| Chicks positive | 36 | 29.72 ± 16.97 | 5 | 2.64 ± 2.43 | 2 | 0.94 ± 0.59 | 3 |
| Pen positive | 36 | 0 | 0 | 0 | 0 | 1.01 ± Inf | 1 |
| Management measure slow growing breed | |||||||
| Chicks positive | 47 | 99.59 ± 1.82 | 6 | 4.23 ± 4.01 | 25 | 3.4 ± 3.33 | 21 |
| Pen positive | 47 | 99.19 ± 1 | 5 | 4.01 ± 3.7 | 30 | 3.11 ± 2.84 | 27 |
| Management measure stocking density 25 kg/m2 | |||||||
| Chicks positive | 36 | 99.16 ± 4.15 | 7 | 4.5 ± 4.04 | 22 | 3.7 ± 3.29 | 21 |
| Pen positive | 36 | 90.64 ± 28.71 | 16 | 4.1 ± 3.98 | 30 | 3.2 ± 3.04 | 32 |
| Management measure stocking density 20 kg/m2 | |||||||
| Chicks positive | 36 | 99.04 ± 5.68 | 7 | 4.44 ± 3.92 | 26 | 3.59 ± 3.15 | 23 |
| Pen positive | 36 | 49.28 ± 5.21 | 3 | 2.7 ± 3.43 | 35 | 1.71 ± 2.45 | 35 |
| Combination of measures litter 3000 g/m2 and stocking density 20 kg/m2 (Combination 1) | |||||||
| Chicks positive | 36 | 56.91 ± 28.45 | 8 | 2.61 ± 2.38 | 2 | 1.47 ± 1.93 | 37 |
| Pen positive | 36 | 0 | 0 | 0 | 0 | 1.19 ± Inf | 1 |
| Combination of measures breed RxR, litter 3000 g/m2 and stocking density 25 kg/m2 (Combination 2) | |||||||
| Chicks positive | 47 | 72.03 ± 28 | 6 | 3.3 ± 3.07 | 2 | 1.41 ± 1.14 | 4 |
| Pen positive | 47 | 32.12 ± 5.09 | 3 | 1.29 ± 0.95 | 3 | 1.37 ± Inf | 1 |
| Measure on microbiota: reduced growth of ESBL E. coli in the intestine | |||||||
| Chicks positive | 36 | 20.68 ± 6.97 | 3 | 1.71 ± 1.46 | 2 | 0.81 ± 0.56 | 2 |
| Pen positive | 36 | 77.04 ± 4.79 | 4 | 1.25 ± 0.63 | 3 | 1.9 ± Inf | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becker, E.; Correia-Carreira, G.; Projahn, M.; Käsbohrer, A. Modeling the Impact of Management Changes on the Infection Dynamics of Extended-Spectrum Beta-Lactamase-Producing Escherichia coli in the Broiler Production. Microorganisms 2022, 10, 981. https://doi.org/10.3390/microorganisms10050981
Becker E, Correia-Carreira G, Projahn M, Käsbohrer A. Modeling the Impact of Management Changes on the Infection Dynamics of Extended-Spectrum Beta-Lactamase-Producing Escherichia coli in the Broiler Production. Microorganisms. 2022; 10(5):981. https://doi.org/10.3390/microorganisms10050981
Chicago/Turabian StyleBecker, Evelyne, Guido Correia-Carreira, Michaela Projahn, and Annemarie Käsbohrer. 2022. "Modeling the Impact of Management Changes on the Infection Dynamics of Extended-Spectrum Beta-Lactamase-Producing Escherichia coli in the Broiler Production" Microorganisms 10, no. 5: 981. https://doi.org/10.3390/microorganisms10050981
APA StyleBecker, E., Correia-Carreira, G., Projahn, M., & Käsbohrer, A. (2022). Modeling the Impact of Management Changes on the Infection Dynamics of Extended-Spectrum Beta-Lactamase-Producing Escherichia coli in the Broiler Production. Microorganisms, 10(5), 981. https://doi.org/10.3390/microorganisms10050981