
Salty Twins: Salt-Tolerance of Terrestrial Cyanocohniella Strains (Cyanobacteria) and Description of C. rudolphia sp. nov. Point towards a Marine Origin of the Genus and Terrestrial Long Distance Dispersal Patterns


Abstract
:1. Introduction
2. Material and Methods
2.1. Origin of Strain and Isolation
2.2. Molecular Characterization
2.3. Morphological Characterization
2.4. Holotype Preparation
2.5. Salt Assays
2.6. Physiology
3. Results
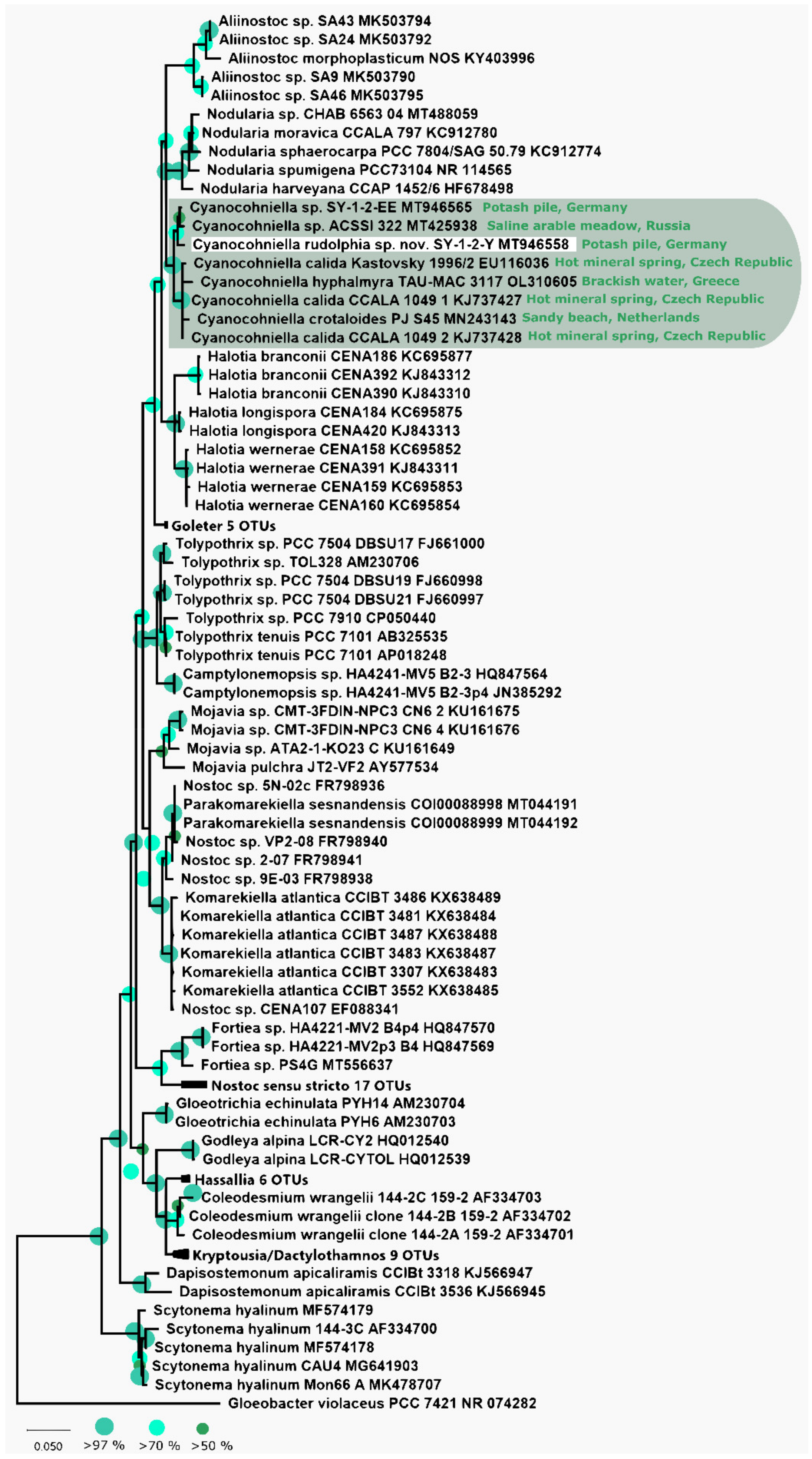
3.1. Phylogeny Based on the 16S rRNA Gene
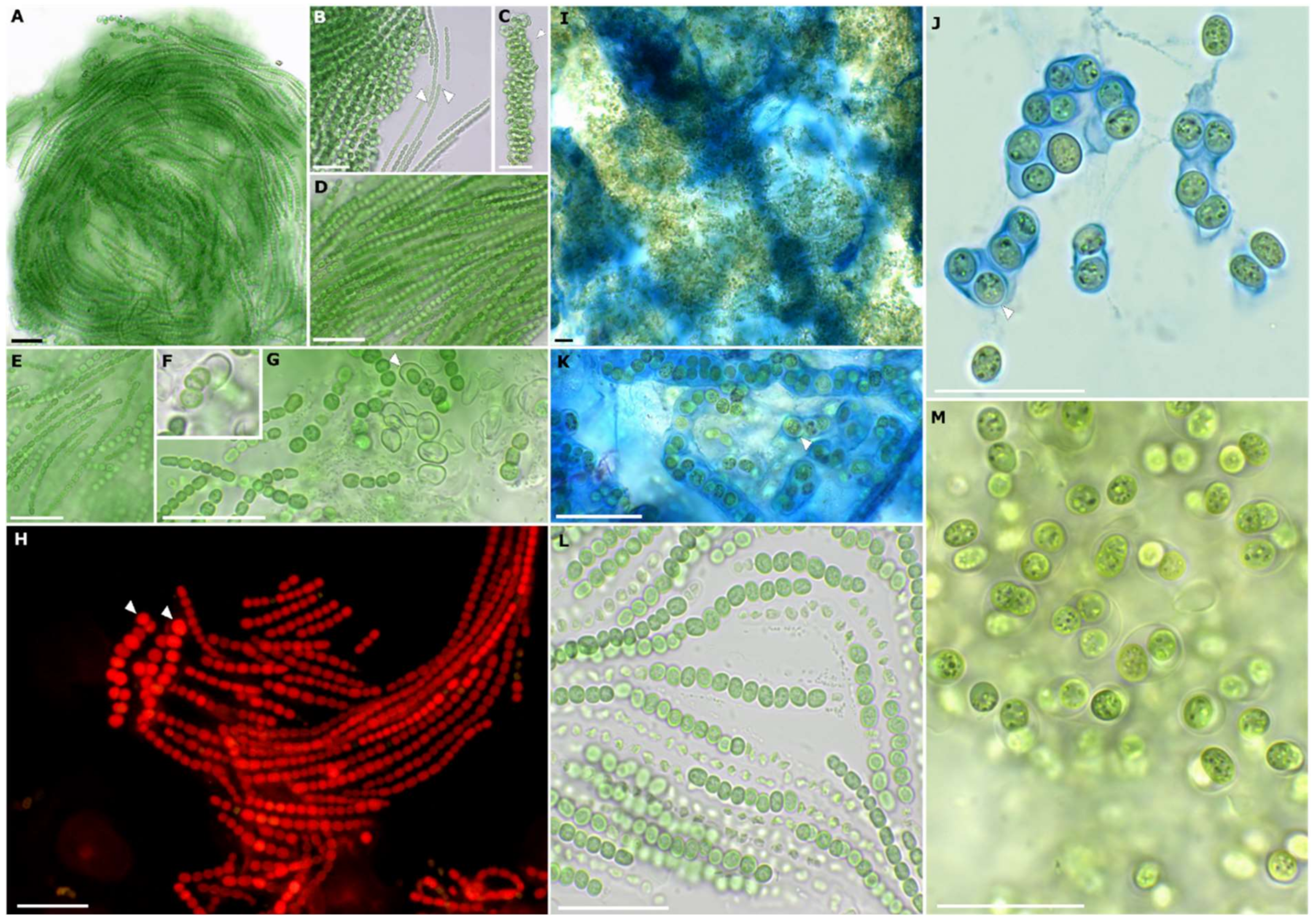
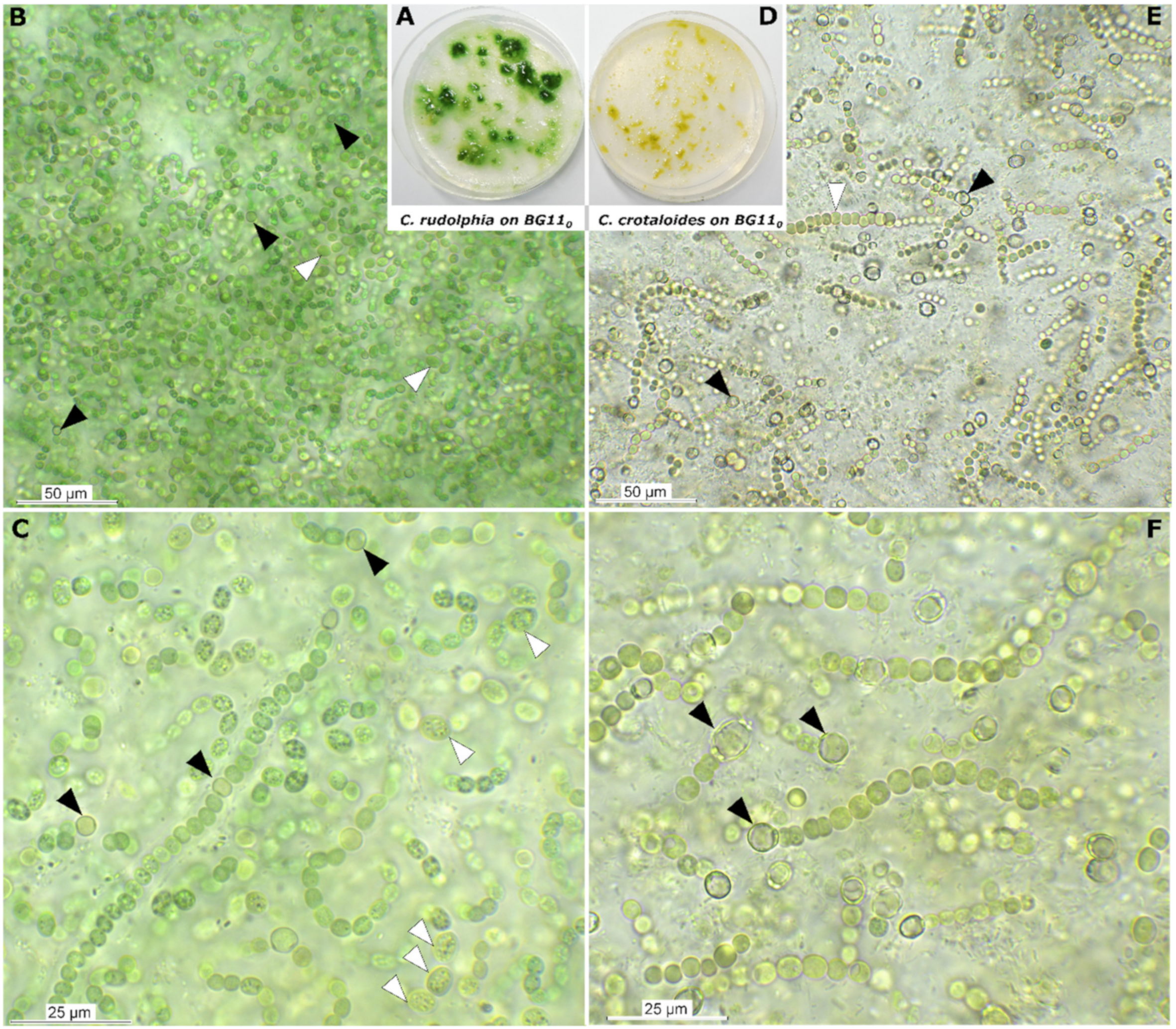
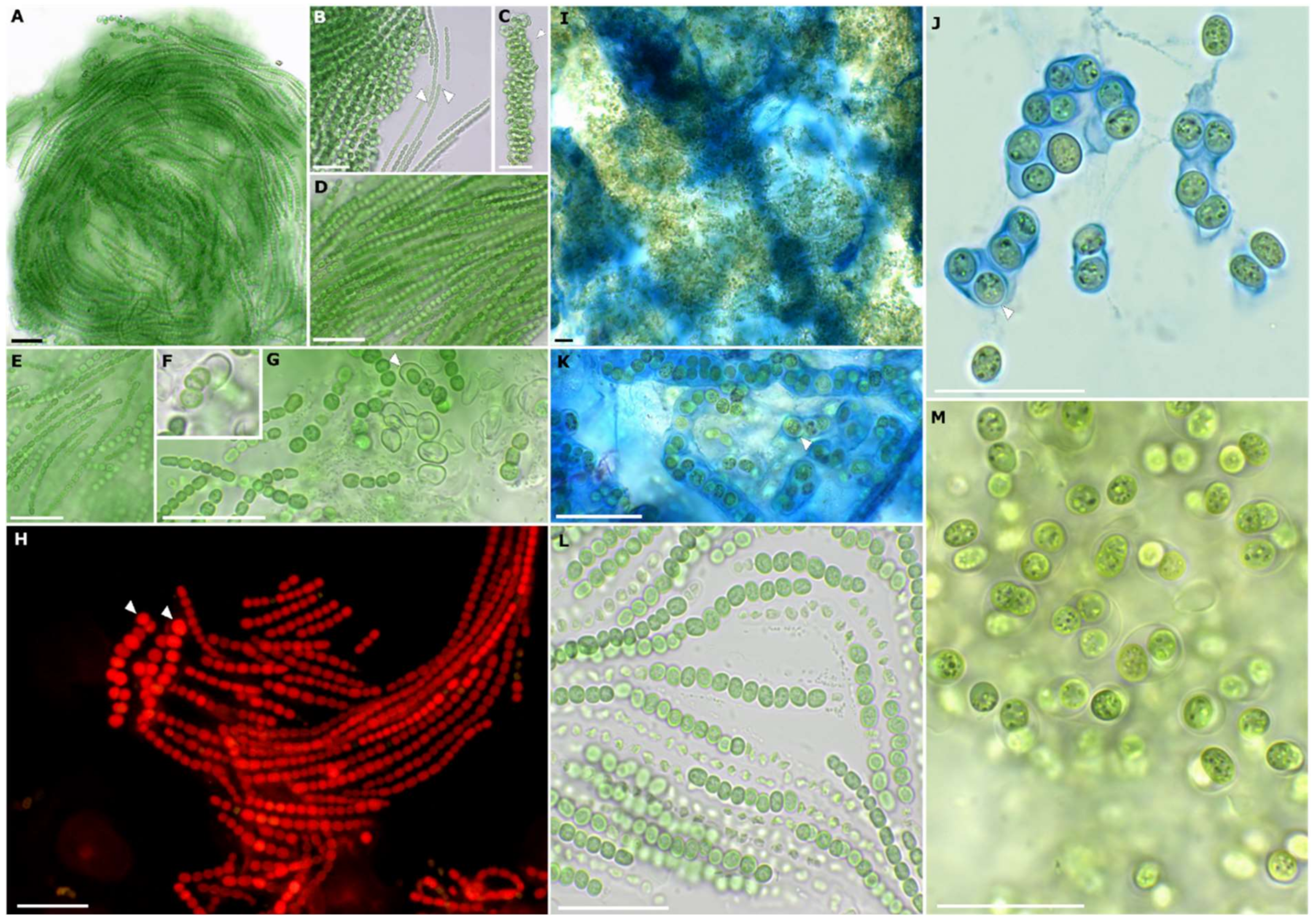
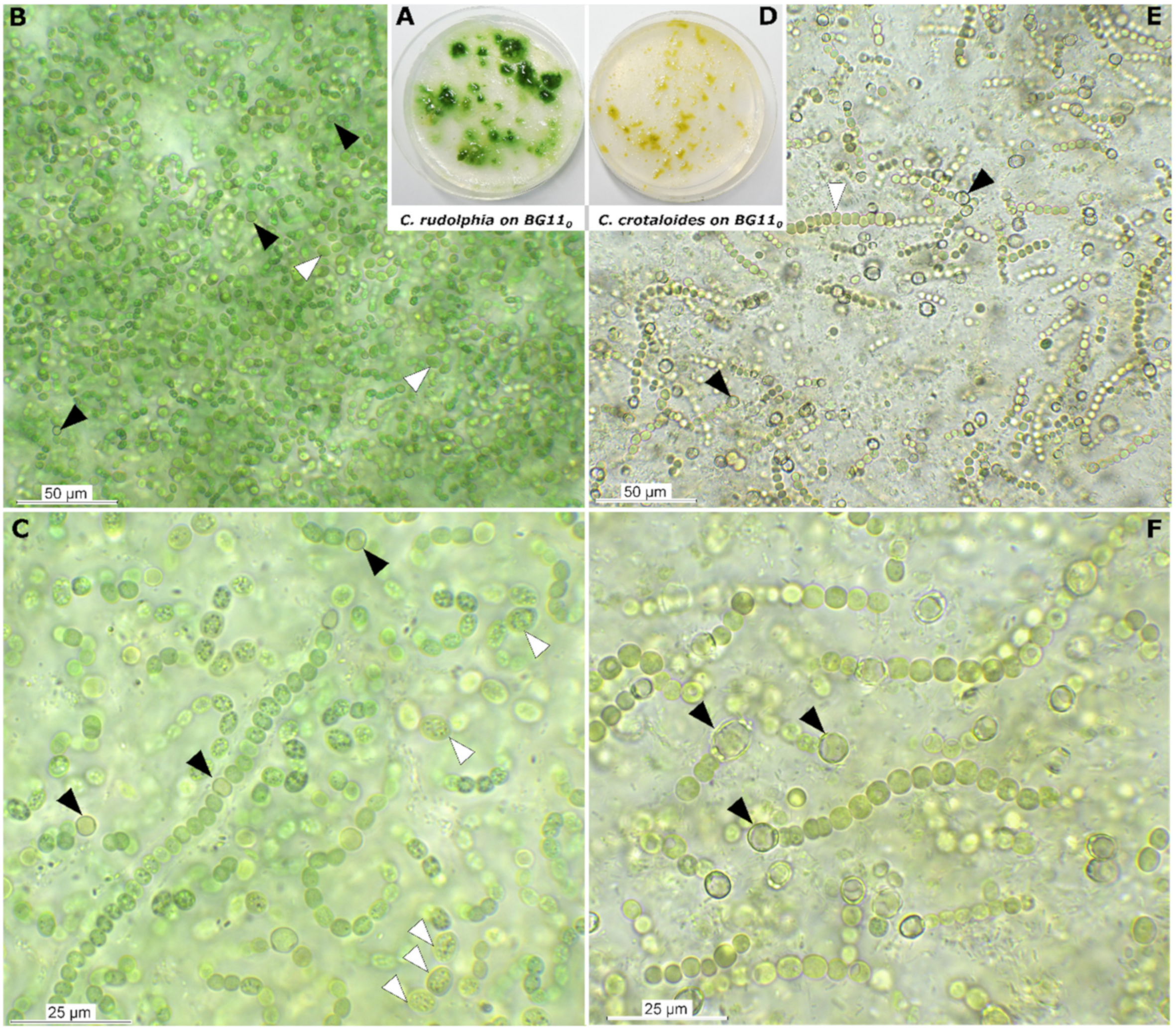
3.2. Strain Morphology and Nitrogen Fixation
3.3. Taxonomy
3.3.1. Cyanocohniella rudolphia sp. nov. P. Jung et V. Sommer
Description
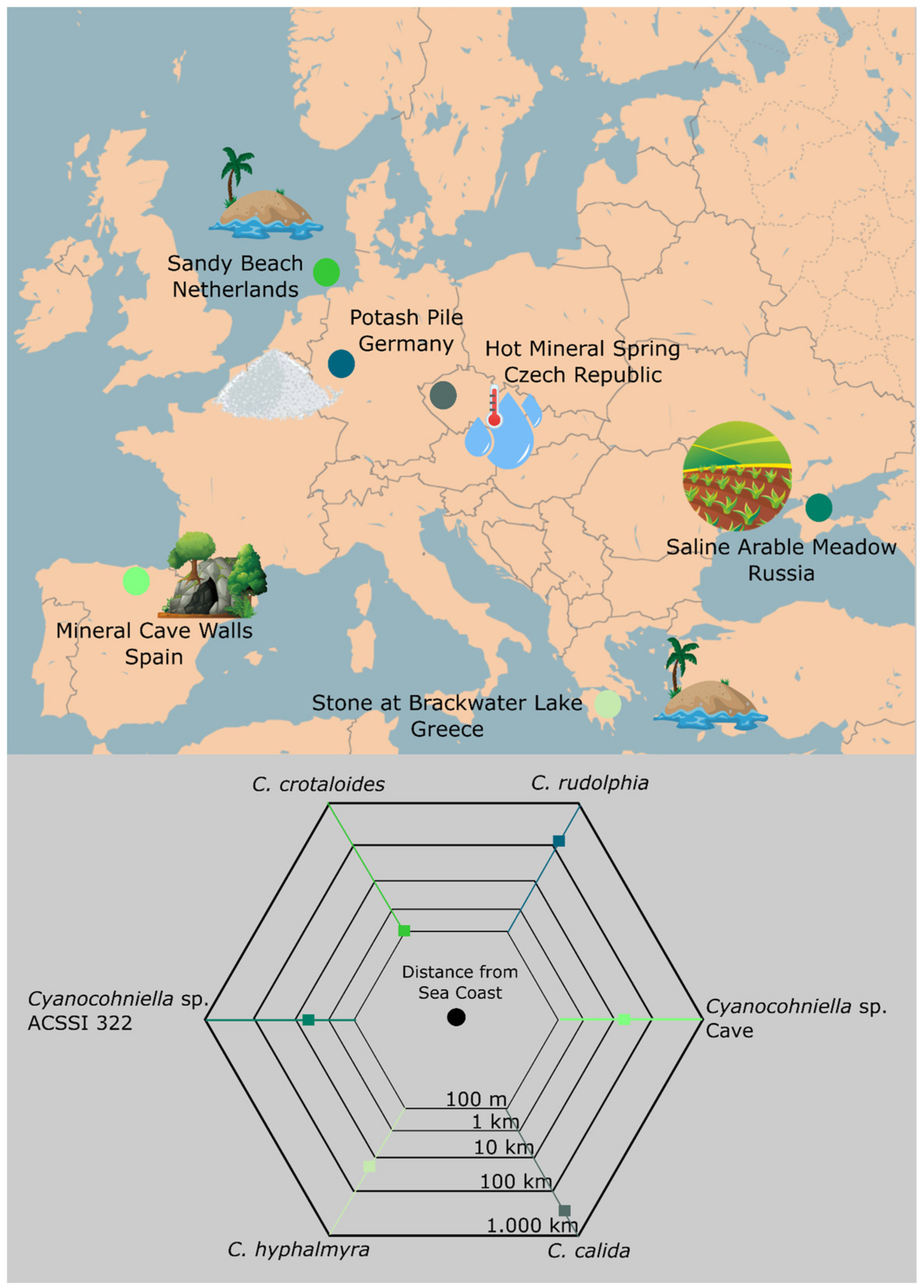
Habitat
Etymology
Type Locality
Holotype
Reference Strain
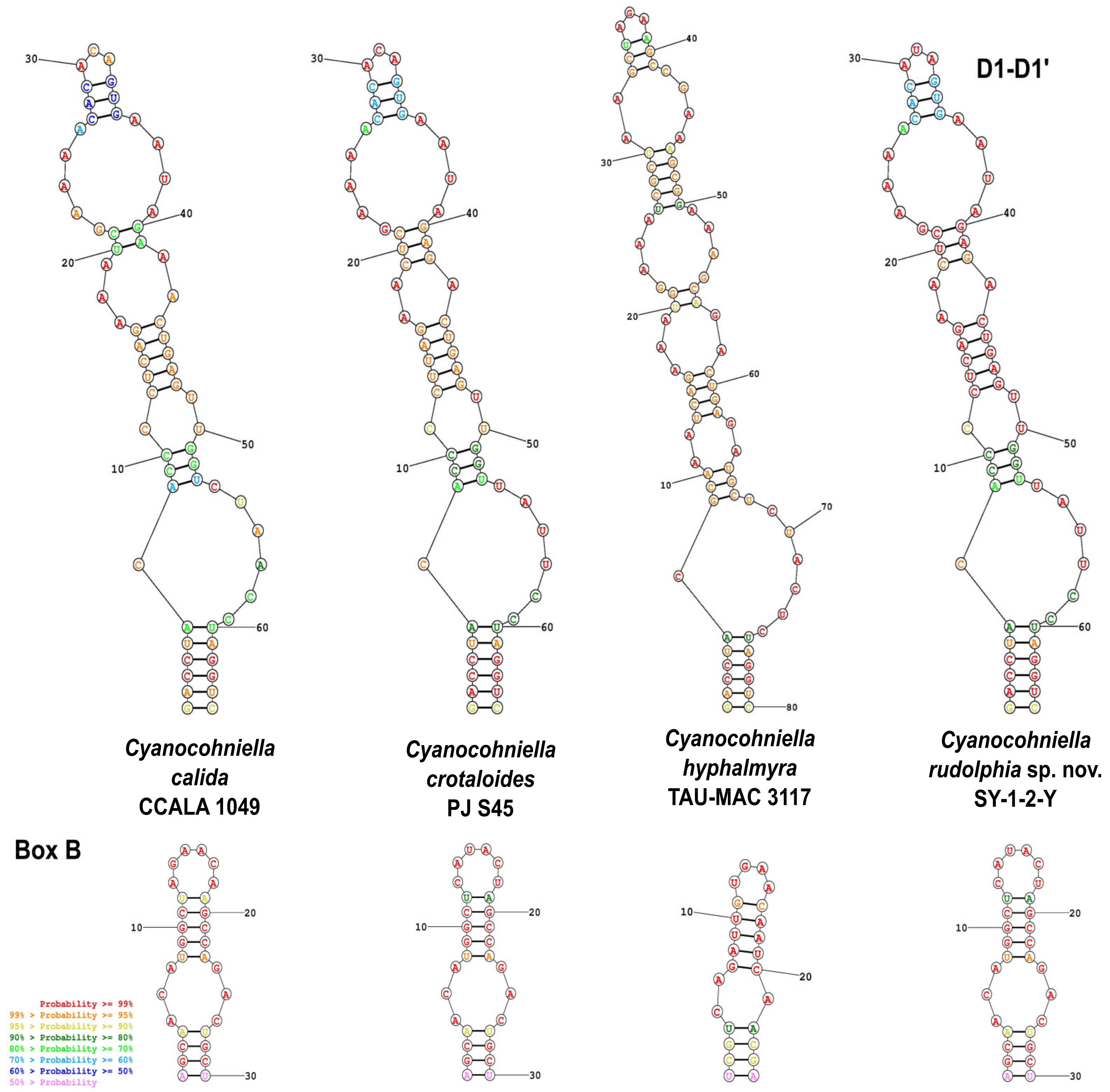
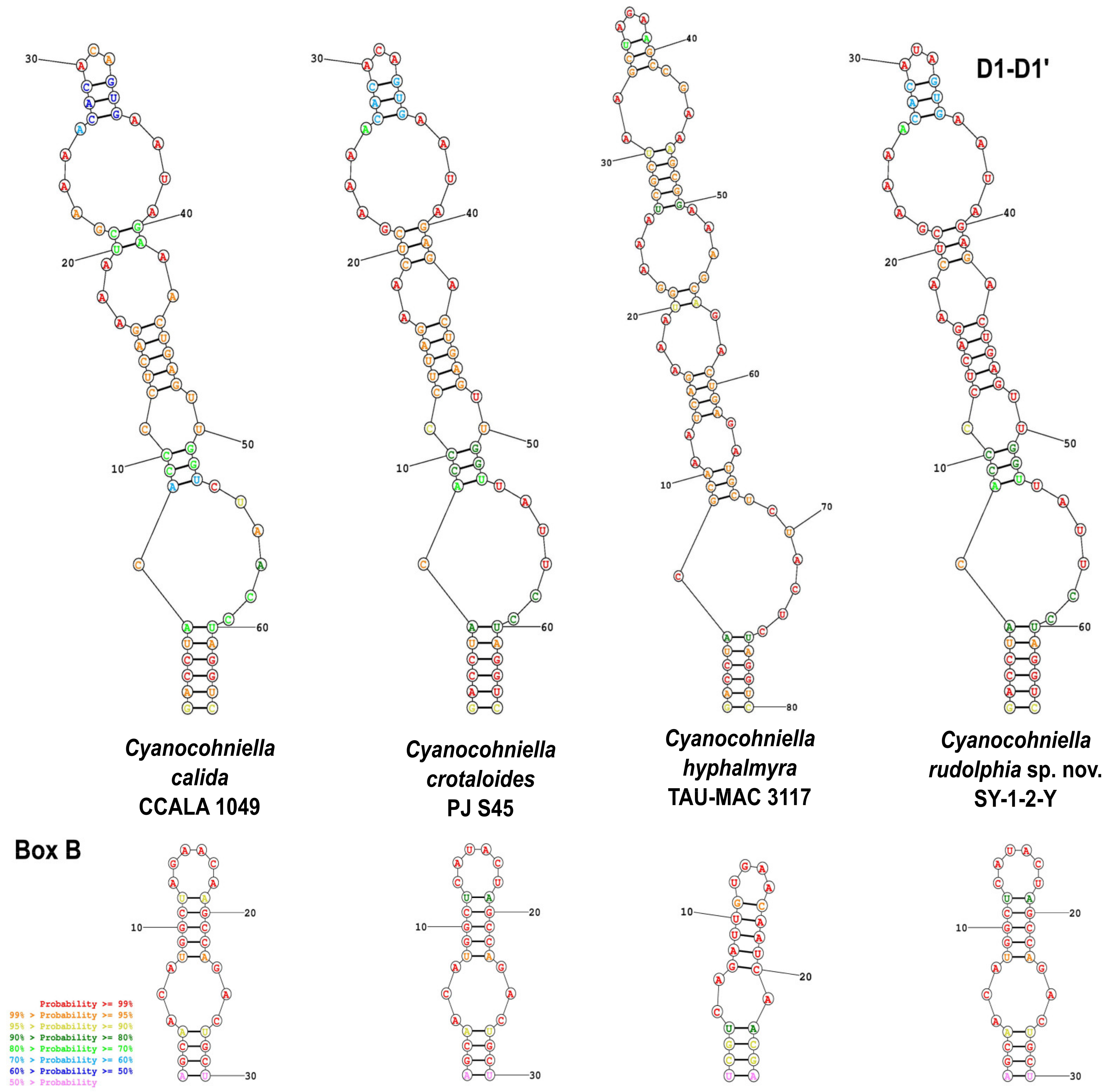
Phylogenetic Relation and Secondary Structure of the 16S-23S ITS Gene
Differentiation against Other Species
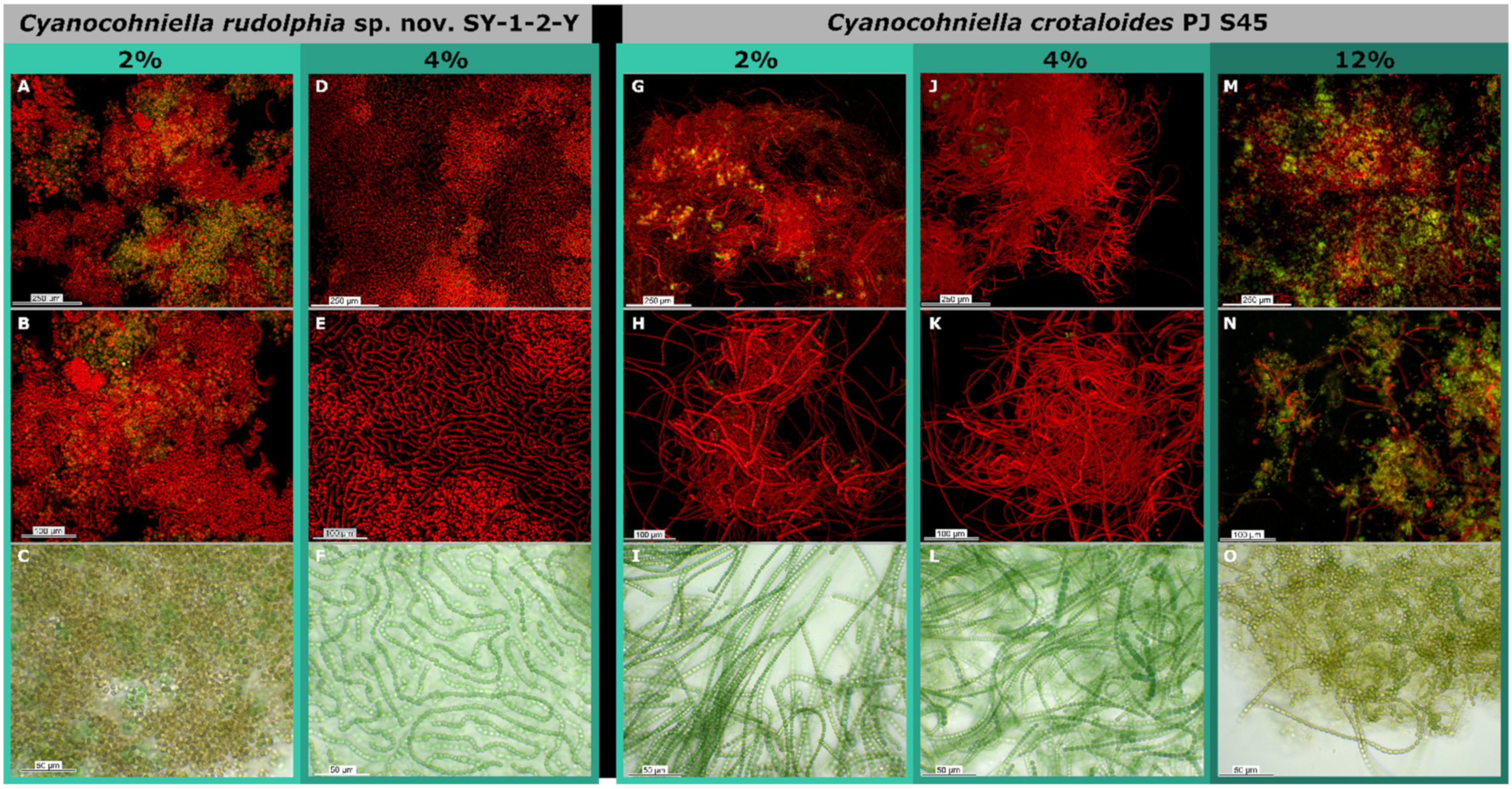
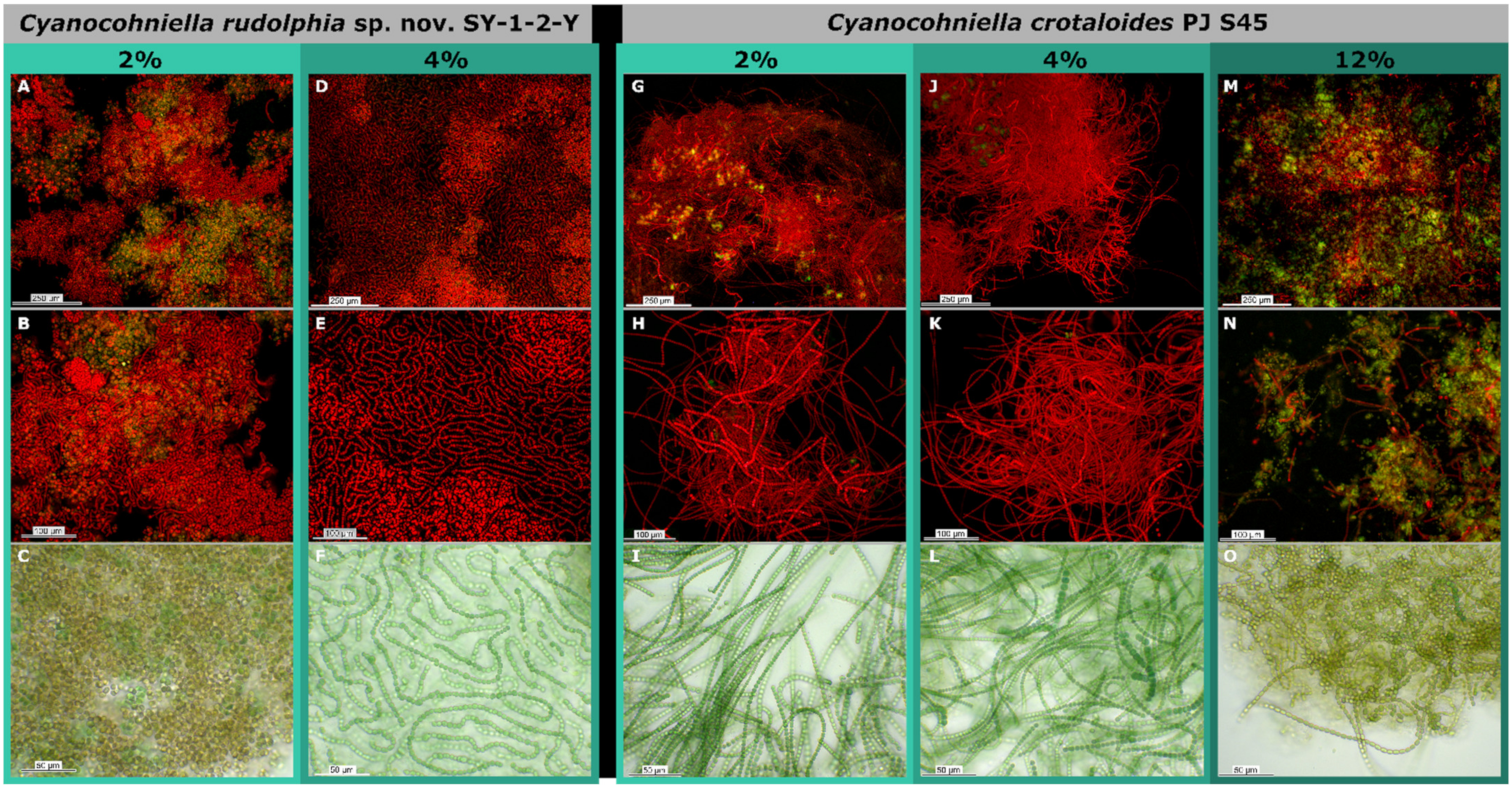
3.4. Salt Assays
4. Discussion
4.1. Implications for Taxonomy
4.2. Salt Tolerance
4.3. Ecological Importance and Biotechnology
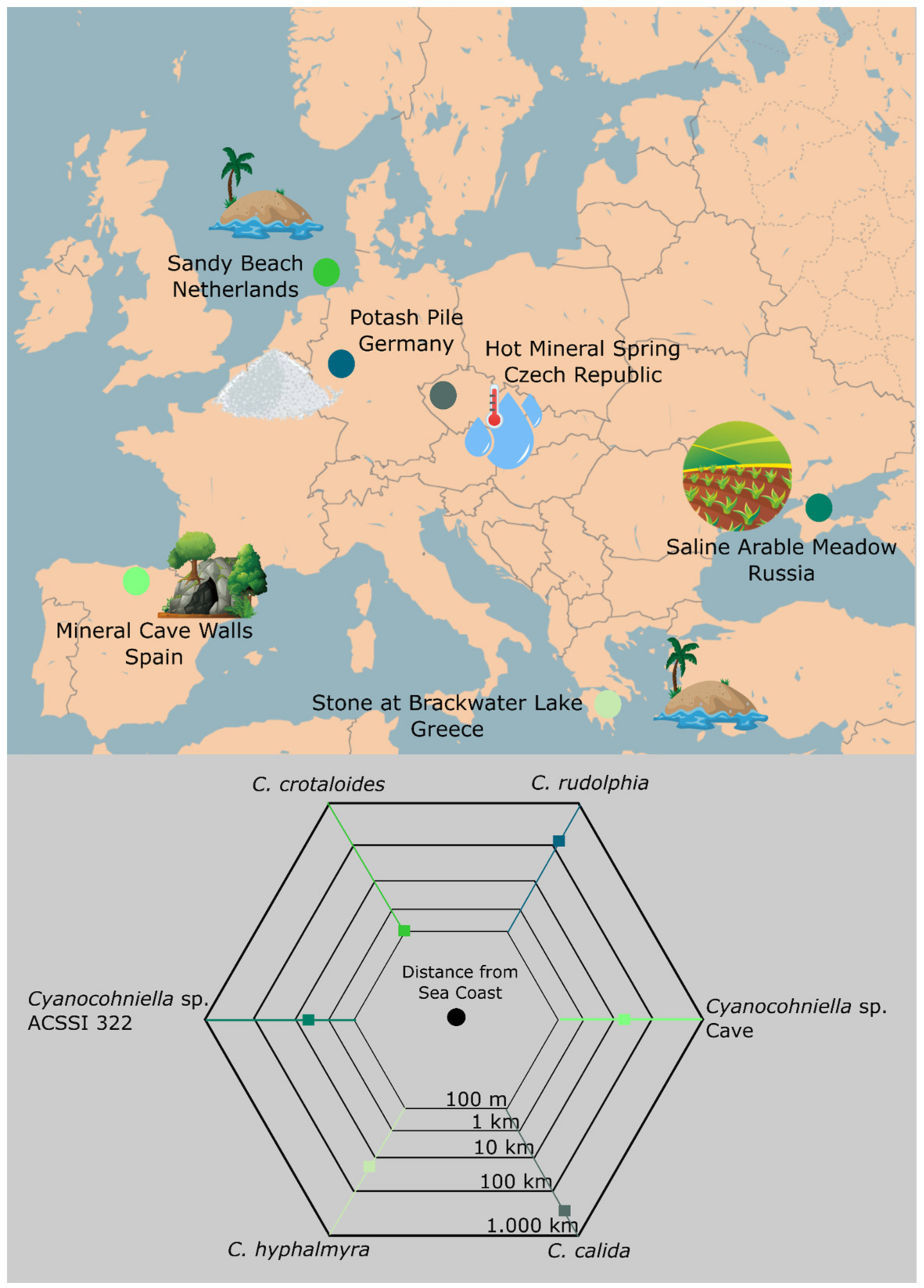
4.4. Implications for Biogeography
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bahl, J.; Lau, M.C.Y.; Smith, G.J.D.; Vijaykrishna, D.; Cary, S.C.; Lacap, D.C.; Lee, C.K.; Papke, R.T.; Warren-Rhodes, K.A.; Wong, F.K.Y.; et al. Ancient origins determine global biogeography of hot and cold desert cyanobacteria. Nat. Commun. 2011, 2, 163. [Google Scholar] [CrossRef] [PubMed]
- Moreira, C.; Fathalli, A.; Vasconcelos, V.; Antunes, A. Phylogeny and biogeography of the invasive cyanobacterium Cylindrospermopsis raciborskii. Arch. Microbiol. 2015, 197, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Del Giudice, D.; Scavia, D.; Binding, C.E.; Bridgeman, T.B.; Chaffin, J.D.; Evans, M.A.; Guinness, J.; Johengen, T.H.; Obenour, D.R. A space-time geostatistical model for probabilistic estimation of harmful algal bloom biomass and areal extent. Sci. Total Environ. 2019, 695, 133776. [Google Scholar] [CrossRef] [PubMed]
- Belnap, J. The world at your feet: Desert biological soil crusts. Front. Ecol. Environ. 2003, 1, 181–189. [Google Scholar] [CrossRef]
- Pushkareva, E.; Pessi, I.S.; Namsaraev, Z.; Mano, M.-J.; Elster, J.; Wilmotte, A. Cyanobacteria inhabiting biological soil crusts of a polar desert: Sør Rondane Mountains, Antarctica. Syst. Appl. Microbiol. 2018, 41, 363–373. [Google Scholar] [CrossRef]
- Behrendt, L.; Trampe, E.L.; Nord, N.B.; Nguyen, J.; Kühl, M.; Lonco, D.; Nyarko, A.; Dhinojwala, A.; Hershey, O.S.; Barton, H. Life in the dark: Far-red absorbing cyanobacteria extend photic zones deep into terrestrial caves. Environ. Microbiol. 2020, 22, 952–963. [Google Scholar] [CrossRef]
- Jung, P.; Brust, K.; Schultz, M.; Büdel, B.; Donner, A.; Lakatos, M. Opening the Gap: Rare Lichens with Rare Cyanobionts–Unexpected Cyanobiont Diversity in Cyanobacterial Lichens of the Order Lichinales. Front. Microbiol. 2021, 12, 728378. [Google Scholar] [CrossRef]
- Billi, D.; Verseux, C.; Fagliarone, C.; Napoli, A.; Baqué, M.; de Vera, J.P. A Desert Cyanobacterium under Simulated Mars-like Conditions in Low Earth Orbit: Implications for the Habitability of Mars. Astrobiology 2019, 19, 158–169. [Google Scholar] [CrossRef] [Green Version]
- Rosic, N. Molecular Mechanisms of Stress Tolerance in Cyanobacteria. In Ecophysiology and Biochemistry of Cyanobacteria; Springer: Singapore, 2021; pp. 131–153. [Google Scholar]
- Singh, H. Desiccation and radiation stress tolerance in cyanobacteria. J. Basic Microbiol. 2018, 58, 813–826. [Google Scholar] [CrossRef]
- Lakatos, M.; Strieth, D. Terrestrial Microalgae: Novel Concepts for Biotechnology and Applications. In Progress in Botany; Springer: Cham, Switzerland, 2017; Volume 79, pp. 269–312. [Google Scholar]
- Chen, M.Y.; Teng, W.K.; Zhao, L.; Hu, C.X.; Zhou, Y.K.; Han, B.P.; Song, L.R.; Shu, W.S. Comparative genomics reveals insights into cyanobacterial evolution and habitat adaptation. ISME J. 2021, 15, 211–227. [Google Scholar] [CrossRef]
- Oren, A. Cyanobacteria in hypersaline environments: Biodiversity and physiological properties. Biodivers. Conserv. 2015, 24, 781–798. [Google Scholar] [CrossRef]
- Hagemann, M. Molecular biology of cyanobacterial salt acclimation. FEMS Microbiol. Rev. 2011, 35, 87–123. [Google Scholar] [CrossRef] [PubMed]
- Hagemann, M.; Pade, N. Heterosides--compatible solutes occurring in prokaryotic and eukaryotic phototrophs. Plant Biol. 2015, 17, 927–934. [Google Scholar] [CrossRef] [PubMed]
- Kirsch, F.; Klähn, S.; Hagemann, M. Salt-Regulated Accumulation of the Compatible Solutes Sucrose and Glucosylglycerol in Cyanobacteria and Its Biotechnological Potential. Front. Microbiol. 2019, 10, 2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarenga, L.V.; de Lucius, S.; Vaz, M.G.M.V.; Araújo, W.L.; Hagemann, M. The novel strain Desmonostoc salinum CCM-UFV059 shows higher salt and desiccation resistance compared to the model strain Nostoc sp. PCC7120. J. Phycol. 2020, 56, 496–506. [Google Scholar] [CrossRef]
- Blumwald, E.; Tel-Or, E. Osmoregulation and cell composition in salt-adaptation of Nostoc muscorum. Arch. Microbiol. 1982, 132, 168–172. [Google Scholar] [CrossRef]
- Kirsch, F.; Pade, N.; Klähn, S.; Hess, W.R.; Hagemann, M. The glucosylglycerol-degrading enzyme GghA is involved in acclimation to fluctuating salinities by the cyanobacterium Synechocystis sp. strain PCC 6803. Microbiology 2017, 163, 1319–1328. [Google Scholar] [CrossRef]
- Cumbers, J.; Rothschild, L.J. Salt tolerance and polyphyly in the cyanobacterium Chroococcidiopsis (Pleurocapsales). J. Phycol. 2014, 50, 472–482. [Google Scholar] [CrossRef]
- Pade, N.; Michalik, D.; Ruth, W.; Belkin, N.; Hess, W.R.; Berman-Frank, I.; Hagemann, M. Trimethylated homoserine functions as the major compatible solute in the globally significant oceanic cyanobacterium Trichodesmium. Proc. Natl. Acad. Sci. USA 2016, 113, 13191–13196. [Google Scholar] [CrossRef] [Green Version]
- Sommer, V.; Mikhailyuk, T.; Glaser, K.; Karsten, U. Uncovering Unique Green Algae and Cyanobacteria Isolated from Biocrusts in Highly Saline Potash Tailing Pile Habitats, Using an Integrative Approach. Microorganisms 2020, 8, 1667. [Google Scholar] [CrossRef]
- Pushkareva, E.; Sommer, V.; Barrantes, I.; Karsten, U. Diversity of Microorganisms in Biocrusts Surrounding Highly Saline Potash Tailing Piles in Germany. Microorganisms 2021, 9, 714. [Google Scholar] [CrossRef] [PubMed]
- Sommer, V.; Karsten, U.; Glaser, K. Halophilic Algal Communities in Biological Soil Crusts Isolated from Potash Tailings Pile Areas. Front. Ecol. Evol. 2020, 8, 46. [Google Scholar] [CrossRef] [Green Version]
- Kaštovský, J.; Gomez, E.B.; Hladil, J.; Johansen, J.R. Cyanocohniella calida gen. et sp. nov. (Cyanobacteria: Aphanizomenonaceae) a new cyanobacterium from the thermal springs from Karlovy Vary, Czech Republic. Phytotaxa 2014, 181, 279. [Google Scholar] [CrossRef]
- Jung, P.; Mikhailyuk, T.; Emrich, D.; Baumann, K.; Dultz, S.; Büdel, B. Shifting Boundaries: Ecological and Geographical Range extension Based on Three New Species in the Cyanobacterial Genera Cyanocohniella, Oculatella, and, Aliterella. J. Phycol. 2020, 56, 1216–1231. [Google Scholar] [CrossRef] [PubMed]
- Panou, M.; Gkelis, S. Unravelling unknown cyanobacteria diversity linked with HCN production. Mol. Phylogenet. Evol. 2022, 166, 107322. [Google Scholar] [CrossRef]
- Turland, N.; Wiersema, J.; Barrie, F.; Greuter, W.; Hawksworth, D.; Herendeen, P.; Knapp, S.; Kusber, W.H.; Li, D.Z.; Marhold, K.; et al. International Code of Nomenclature for algae, fungi, and plants. IMA Fungus 2018, 159. [Google Scholar] [CrossRef]
- Marin, B.; Nowack, E.C.M.; Melkonian, M. A plastid in the making: Evidence for a second primary endosymbiosis. Protist 2005, 156, 425–432. [Google Scholar] [CrossRef]
- Wilmotte, A.; van der Auwera, G.; de Wachter, R. Structure of the 16 S ribosomal RNA of the thermophilic cyanobacterium chlorogloeopsis HTF (‘mastigocladus laminosus HTF’) strain PCC7518, and phylogenetic analysis. FEBS Lett. 1993, 317, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Bellaousov, S.; Reuter, J.S.; Seetin, M.G.; Mathews, D.H. RNAstructure: Web servers for RNA secondary structure prediction and analysis. Nucleic Acids Res. 2013, 41, W471–W474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genty, B.; Harbinson, J.; Briantais, J.M.; Baker, N.R. The relationship between non-photochemical quenching of chlorophyll fluorescence and the rate of photosystem 2 photochemistry in leaves. Photosynth. Res. 1990, 25, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Komarek, J.; Kaštovský, J.; Mares, J.; Johansen, J. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Caumette, P.; Matheron, R.; Raymond, N.; Relexans, J.C. Microbial mats in the hypersaline ponds of Mediterranean salterns (Salins-de-Giraud, France). FEMS Microbiol. Ecol. 1994, 13, 273–286. [Google Scholar] [CrossRef]
- Schneider, D.; Arp, G.; Reimer, A.; Reitner, J.; Daniel, R. Phylogenetic analysis of a microbialite-forming microbial mat from a hypersaline lake of the Kiritimati atoll, Central Pacific. PLoS ONE 2013, 8, e66662. [Google Scholar] [CrossRef] [Green Version]
- Shalygin, S.; Pietrasiak, N.; Gomez, F.; Mlewski, C.; Gerard, E.; Johansen, J.R. Rivularia halophila sp. nov. (Nostocales, Cyanobacteria): The first species of Rivularia described with the modern polyphasic approach. Eur. J. Phycol. 2018, 53, 537–548. [Google Scholar] [CrossRef]
- Brock, T.D. Halophilic-blue-green algae. Arch. Microbiol. 1976, 107, 109–111. [Google Scholar] [CrossRef]
- Karsten, U. Growth and organic osmolytes of geographically different isolates of Microcoleus chtonoplastes (Cyanobacteria) from benthic microbial mats: Response to salinity change. J. Phycol. 1996, 32, 501–506. [Google Scholar] [CrossRef]
- Wierzchos, J.; Ascaso, C.; McKay, C.P. Endolithic cyanobacteria in halite rocks from the hyperarid core of the Atacama Desert. Astrobiology 2006, 6, 415–422. [Google Scholar] [CrossRef]
- Robinson, C.K.; Wierzchos, J.; Black, C.; Crits-Christoph, A.; Ma, B.; Ravel, J.; Ascaso, C.; Artieda, O.; Valea, S.; Roldán, M.; et al. Microbial diversity and the presence of algae in halite endolithic communities are correlated to atmospheric moisture in the hyper-arid zone of the Atacama Desert. Environ. Microbiol. 2015, 17, 299–315. [Google Scholar] [CrossRef]
- Singh, S.P.; Montgomery, B.L. Salinity impacts photosynthetic pigmentation and cellular morphology changes by distinct mechanisms in Fremyella diplosiphon. Biochem. Biophys. Res. Commun. 2013, 433, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Bhadauriya, P.; Gupta, R.; Singh, S.; Bisen, P.S. Physiological and biochemical alterations in a diazotrophic cyanobacterium Anabaena cylindrica under NaCl stress. Curr. Microbiol. 2007, 55, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Verma, E.; Singh, S.; Niveshika; Mishra, A.K. Salinity-induced oxidative stress-mediated change in fatty acids composition of cyanobacterium Synechococcus sp. PCC7942. Int. J. Environ. Sci. Technol. 2019, 16, 875–886. [Google Scholar] [CrossRef]
- Meury, J. Glycine betaine reverses the effects of osmotic stress on DNA replication and cellular division in Escherichia coli. Arch. Microbiol. 1988, 149, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Piuri, M.; Sanchez-Rivas, C.; Ruzal, S.M. Cell wall modifications during osmotic stress in Lactobacillus casei. J. Appl. Microbiol. 2005, 98, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Yoshida, T.; Arima, H.; Hatanaka, Y.; Takani, Y.; Tamaru, Y. Accumulation of trehalose in response to desiccation and salt stress in the terrestrial cyanobacterium Nostoc commune. Phycol. Res. 2009, 57, 66–73. [Google Scholar] [CrossRef]
- Chen, L.; Li, D.; Liu, Y. Salt tolerance of Microcoleus vaginatus Gom., a cyanobacterium isolated from desert algal crust, was enhanced by exogenous carbohydrates. J. Arid. Environ. 2003, 55, 645–656. [Google Scholar] [CrossRef]
- Yoshimura, H.; Kotake, T.; Aohara, T.; Tsumuraya, Y.; Ikeuchi, M.; Ohmori, M. The role of extracellular polysaccharides produced by the terrestrial cyanobacterium Nostoc sp. strain HK-01 in NaCl tolerance. J. Appl. Phycol. 2012, 24, 237–243. [Google Scholar] [CrossRef]
- Mugnai, G.; Rossi, F.; Felde, V.J.M.N.L.; Colesie, C.; Büdel, B.; Peth, S.; Kaplan, A.; de Philippis, R. Development of the polysaccharidic matrix in biocrusts induced by a cyanobacterium inoculated in sand microcosms. Biol. Fertil. Soils 2018, 54, 27–40. [Google Scholar] [CrossRef]
- Chamizo, S.; Adessi, A.; Torzillo, G.; Philippis, R. Exopolysaccharide Features Influence Growth Success in Biocrust-forming Cyanobacteria, Moving from Liquid Culture to Sand Microcosms. Front. Microbiol. 2020, 11, 568224. [Google Scholar] [CrossRef]
- Chisti, Y. Constraints to commercialization of algal fuels. J. Biotechnol. 2013, 167, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Luley-Goedl, C.; Sawangwan, T.; Mueller, M.; Schwarz, A.; Nidetzky, B. Biocatalytic Process for Production of α-Glucosylglycerol Using Sucrose Phosphorylase. Food Technol. Biotechnol. 2010, 48, 276–283. [Google Scholar]
- Tan, X.; Du, W.; Lu, X. Photosynthetic and extracellular production of glucosylglycerol by genetically engineered and gel-encapsulated cyanobacteria. Appl. Microbiol. Biotechnol. 2015, 99, 2147–2154. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhao, Q.; Huang, H. Current states and challenges of salt-affected soil remediation by cyanobacteria. Sci. Total Environ. 2019, 669, 258–272. [Google Scholar] [CrossRef]
- Giraldo-Silva, A.; Nelson, C.; Penfold, C.; Barger, N.N.; Garcia-Pichel, F. Effect of preconditioning to the soil environment on the performance of 20 cyanobacterial strains used as inoculum for biocrust restoration. Restor. Ecol. 2020, 28, S187–S193. [Google Scholar] [CrossRef]
- Sommer, V.; Palm, A.; Schink, A.; Leinweber, P.; Gose, N.; Karsten, U.; Glaser, K. Artificial biocrust establishment on materials of potash tailings piles along a salinity gradient. J. Appl. Phycol. 2022, 34, 405–421. [Google Scholar] [CrossRef]
- Sharma, N.K.; Rai, A.K.; Singh, S.; Brown, R.M. Airborne Algae: Their Present Status and Relevance 1. J. Phycol. 2007, 43, 615–627. [Google Scholar] [CrossRef]
- Finlay, B.J. Global dispersal of free-living microbial eukaryote species. Science 2002, 296, 1061–1063. [Google Scholar] [CrossRef] [Green Version]
- Baas Becking, L.G.M. Geobiologie of Inleiding tot de Milieukunde; WP Van Stockum and Zoon: The Hague, The Netherlands, 1934. [Google Scholar]
- Fenchel, T.O.; Finlay, B.J. The Ubiquity of Small Species: Patterns of Local and Global Diversity. BioScience 2004, 54, 777. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NaCl Concentration (%) | Mean ΦPSII C. crotaloides | SD | Mean ΦPSII C. rudolphia | SD |
|---|---|---|---|---|
| Control | 0.206 | 0.033 | 0.173 | 0.007 |
| 0.5 | 0.258 | 0.007 | 0.167 | 0.022 |
| 0.75 | 0.263 | 0.013 | 0.233 | 0.022 |
| 1 | 0.229 | 0.036 | 0.171 | 0.012 |
| 2 | 0.276 | 0.004 | 0.193 | 0.090 |
| 4 | 0.292 | 0.008 | 0.473 | 0.174 |
| 12 | 0.014 | 0.020 | 0.023 | 0.004 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, P.; Sommer, V.; Karsten, U.; Lakatos, M. Salty Twins: Salt-Tolerance of Terrestrial Cyanocohniella Strains (Cyanobacteria) and Description of C. rudolphia sp. nov. Point towards a Marine Origin of the Genus and Terrestrial Long Distance Dispersal Patterns. Microorganisms 2022, 10, 968. https://doi.org/10.3390/microorganisms10050968
Jung P, Sommer V, Karsten U, Lakatos M. Salty Twins: Salt-Tolerance of Terrestrial Cyanocohniella Strains (Cyanobacteria) and Description of C. rudolphia sp. nov. Point towards a Marine Origin of the Genus and Terrestrial Long Distance Dispersal Patterns. Microorganisms. 2022; 10(5):968. https://doi.org/10.3390/microorganisms10050968
Chicago/Turabian StyleJung, Patrick, Veronika Sommer, Ulf Karsten, and Michael Lakatos. 2022. "Salty Twins: Salt-Tolerance of Terrestrial Cyanocohniella Strains (Cyanobacteria) and Description of C. rudolphia sp. nov. Point towards a Marine Origin of the Genus and Terrestrial Long Distance Dispersal Patterns" Microorganisms 10, no. 5: 968. https://doi.org/10.3390/microorganisms10050968
APA StyleJung, P., Sommer, V., Karsten, U., & Lakatos, M. (2022). Salty Twins: Salt-Tolerance of Terrestrial Cyanocohniella Strains (Cyanobacteria) and Description of C. rudolphia sp. nov. Point towards a Marine Origin of the Genus and Terrestrial Long Distance Dispersal Patterns. Microorganisms, 10(5), 968. https://doi.org/10.3390/microorganisms10050968