
A Novel Isolate of Spherical Multicellular Magnetotactic Prokaryotes Has Two Magnetosome Gene Clusters and Synthesizes Both Magnetite and Greigite Crystals

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Enrichment of MMPs
2.2. Optical and Electron Microscopy
2.3. Genomic DNA Extraction, Whole Genome Amplification, and Phylogenetic Analysis of 16S rRNA Genes
2.4. Fluorescence In Situ Hybridization (FISH)
2.5. Sequencing, Assembly, and Genome Annotation, and Comparative Analysis of Magnetosome Genes and Proteins
3. Results
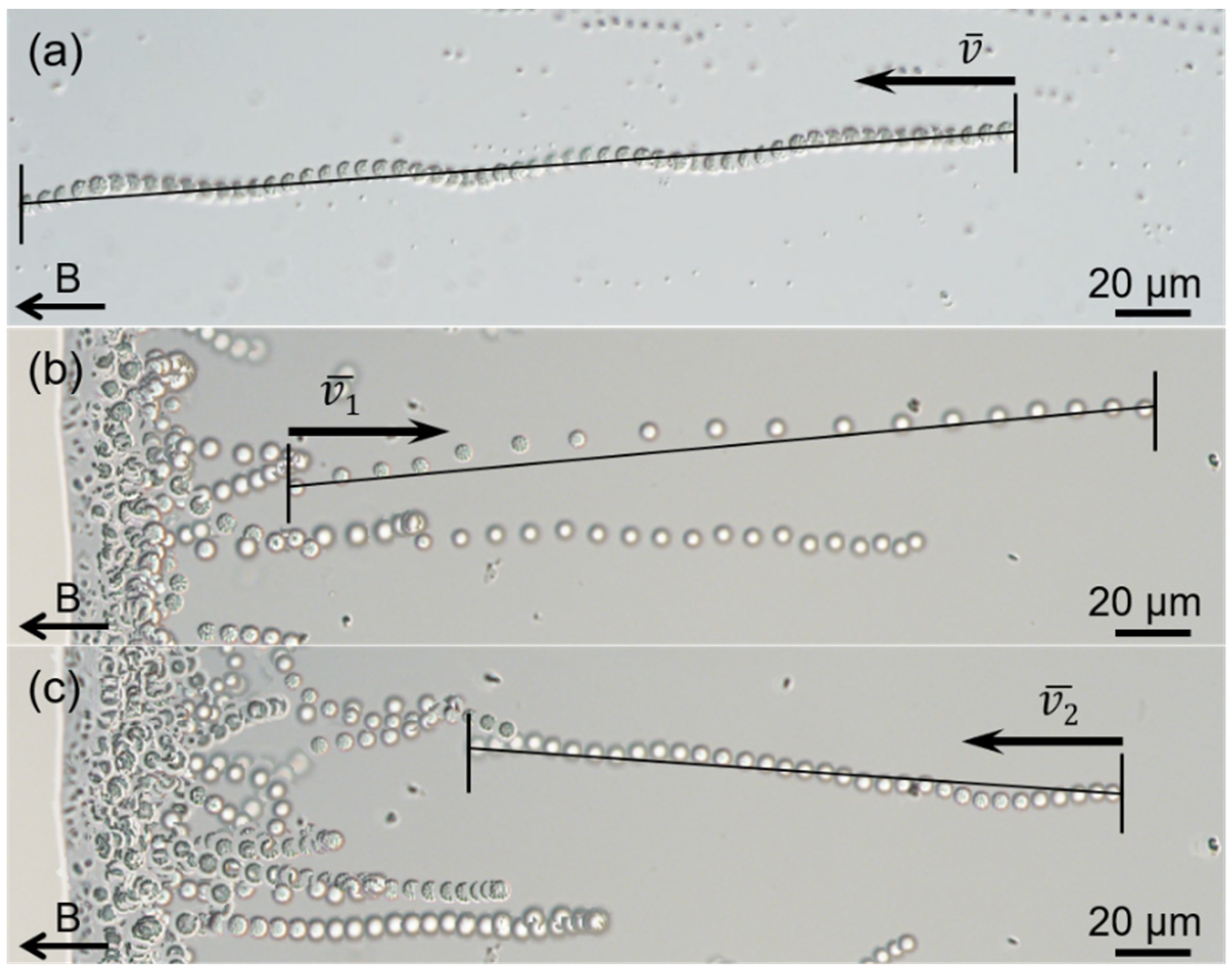
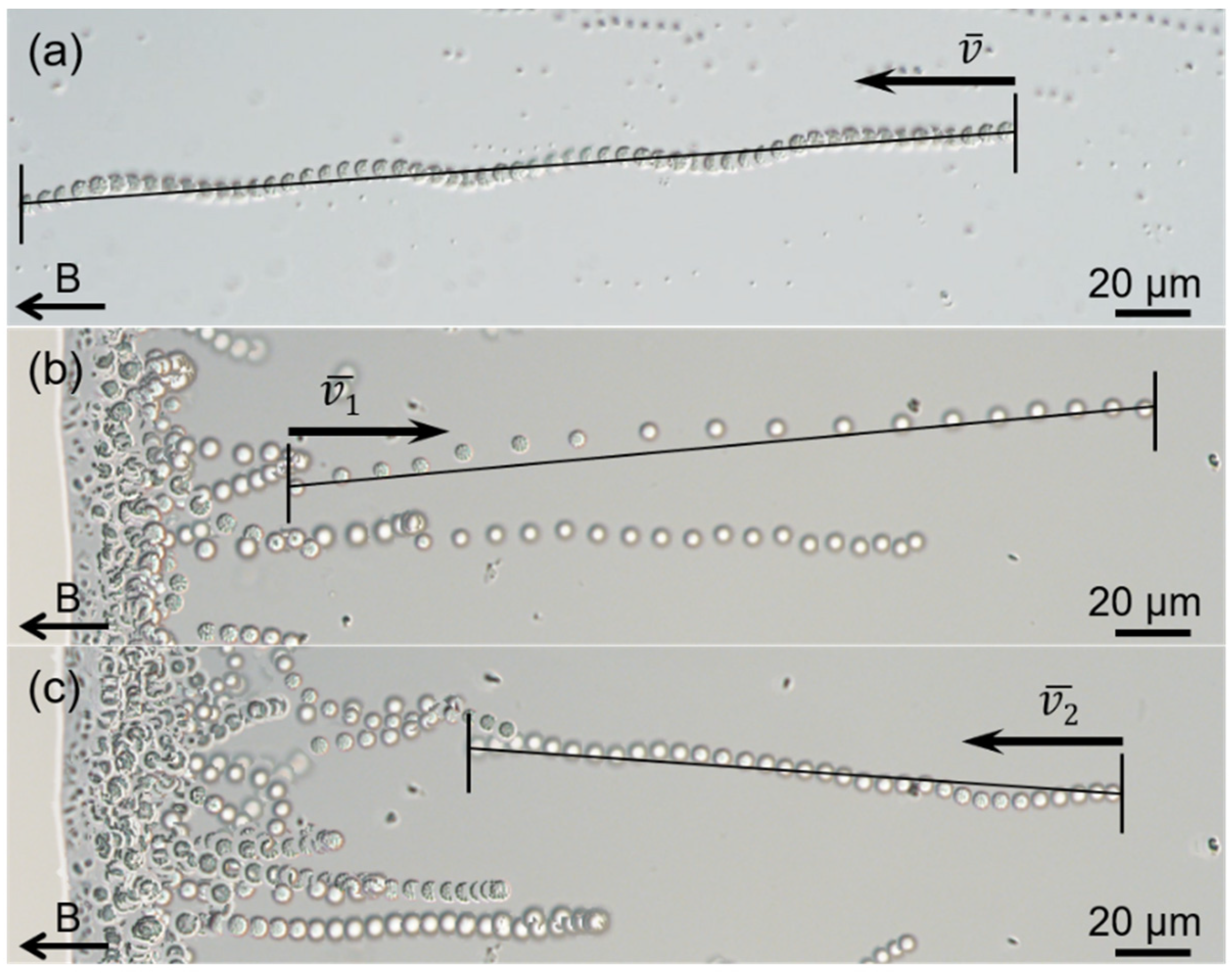
3.1. Occurrence, Structure, and Motility of the sMMPs
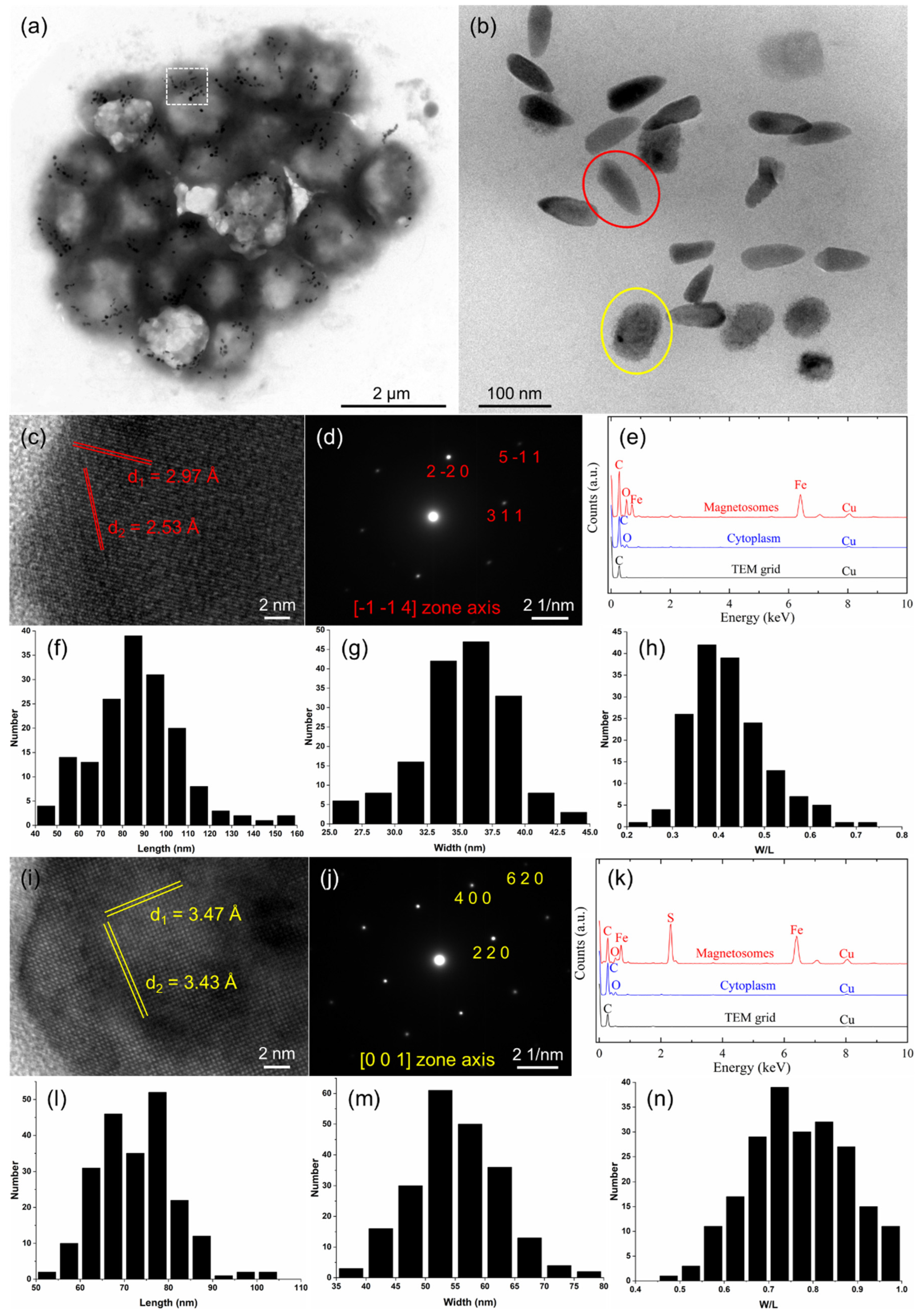
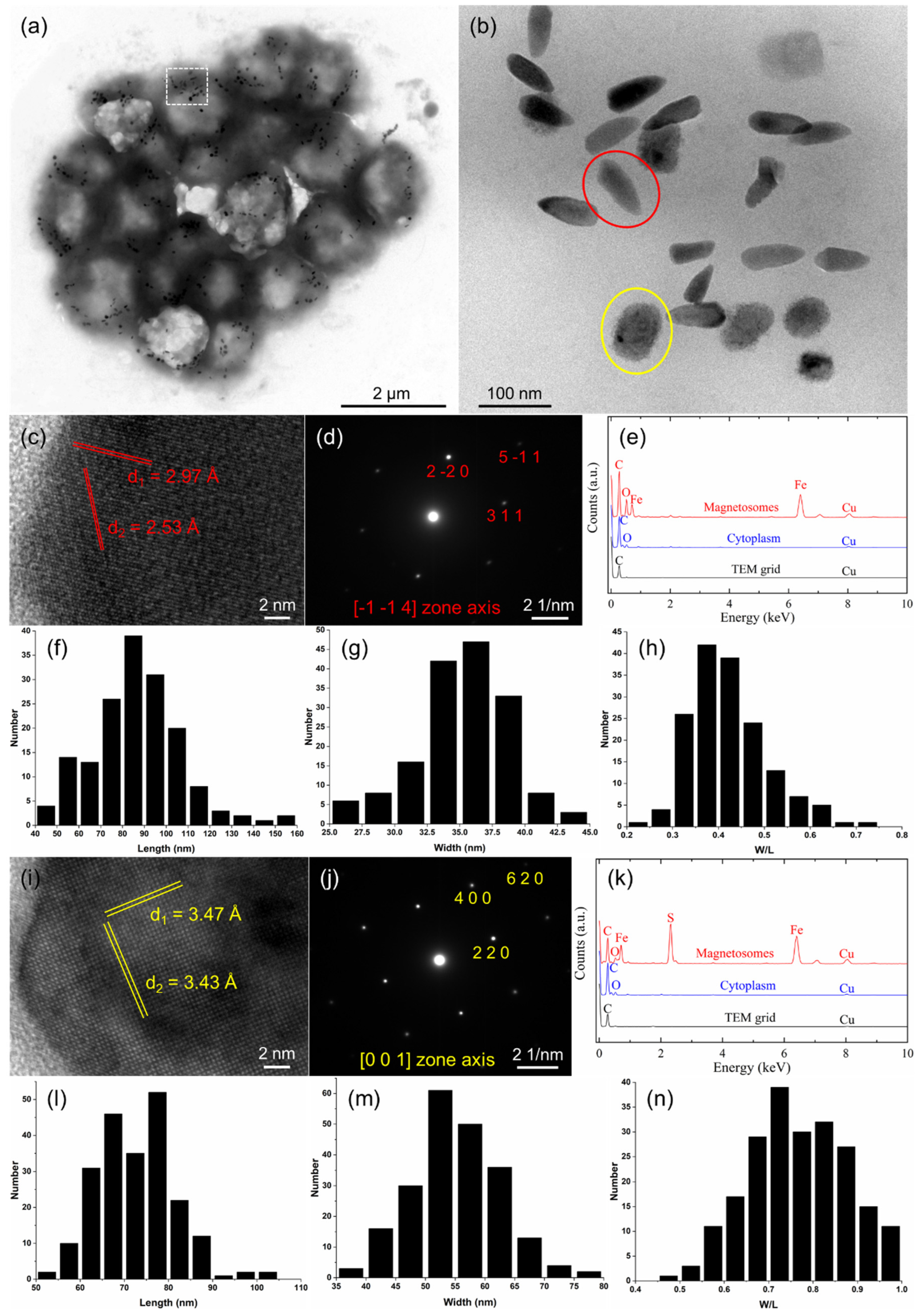
3.2. Characterization of Magnetosome Biomineralization in the sMMPs
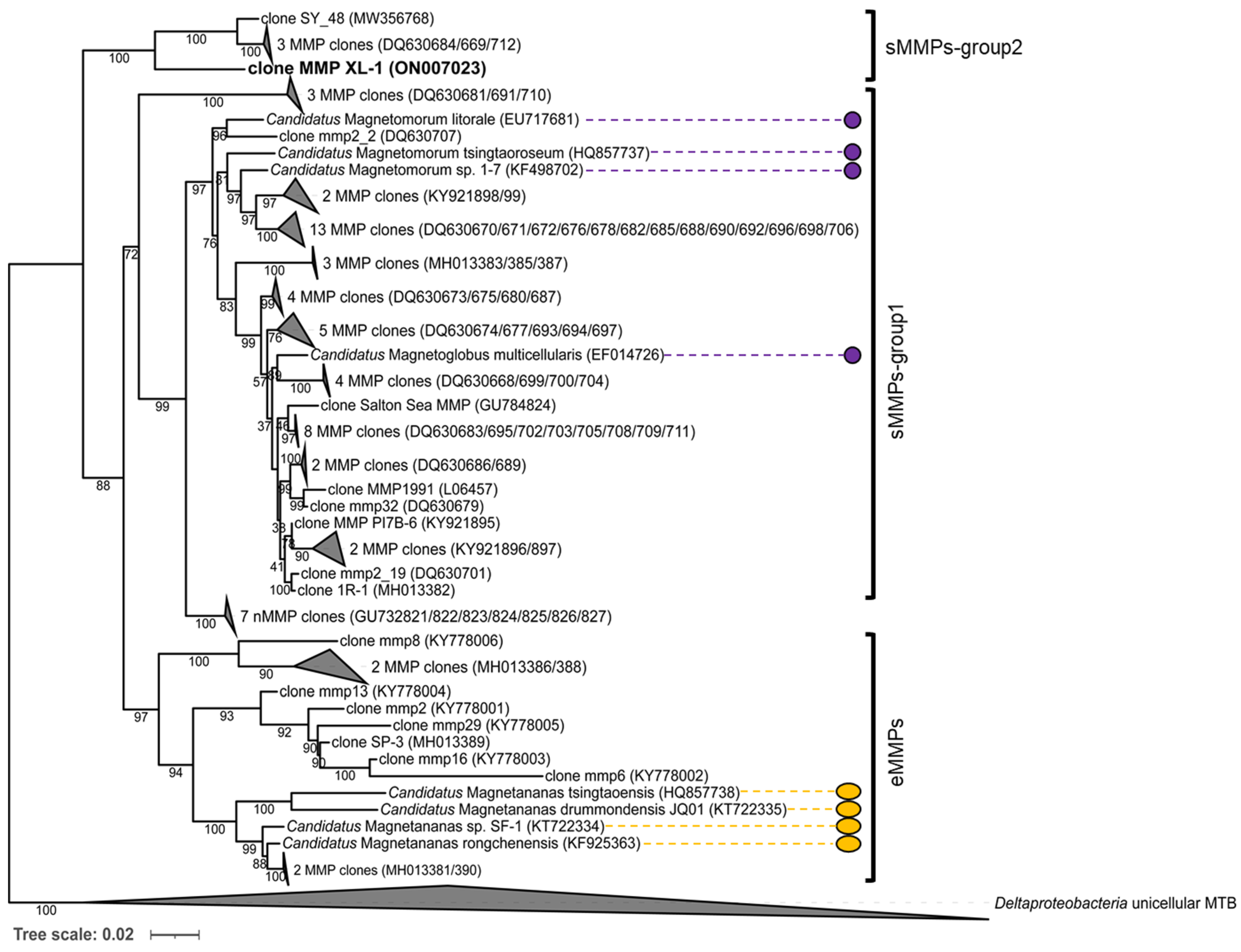
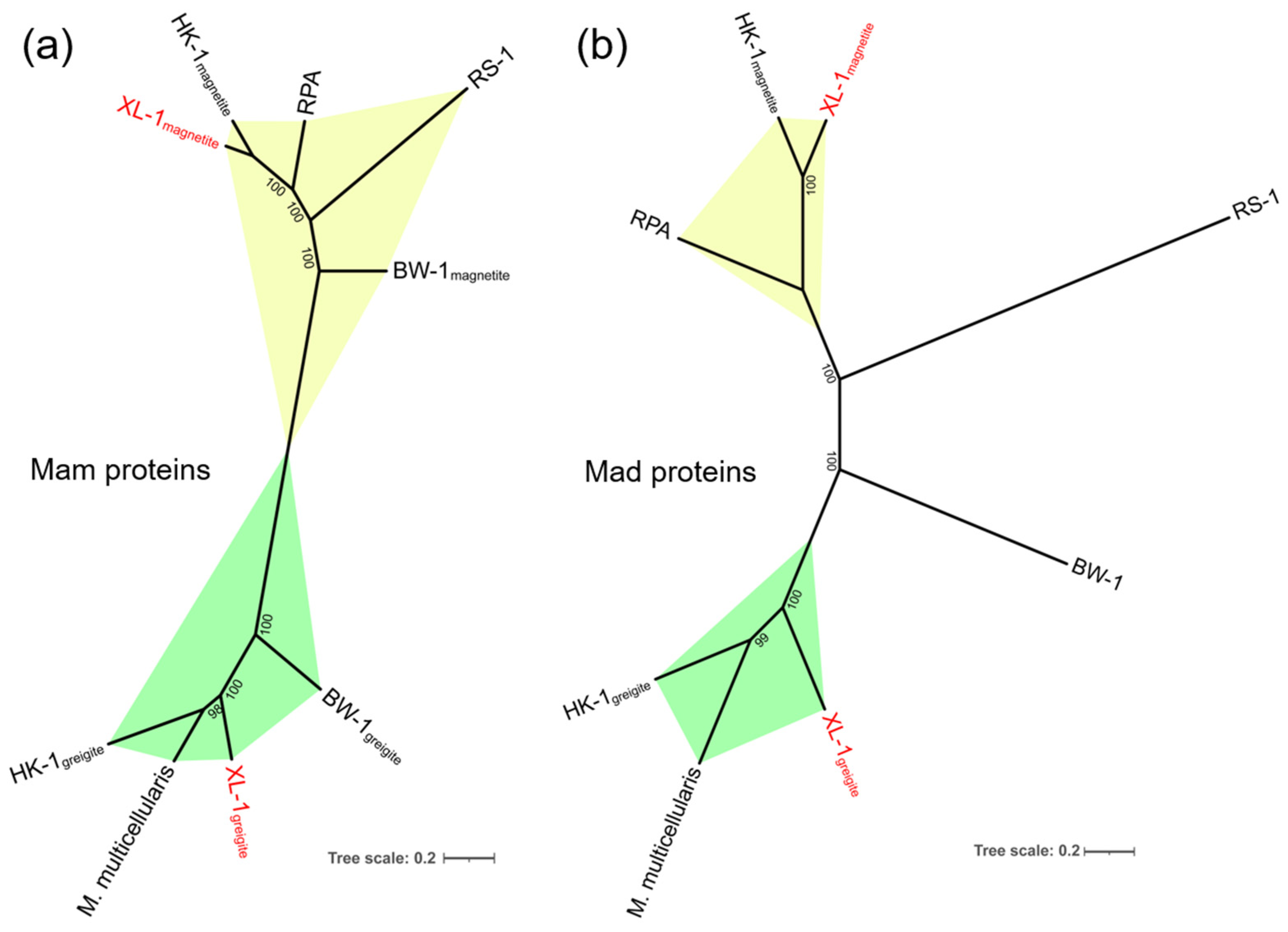
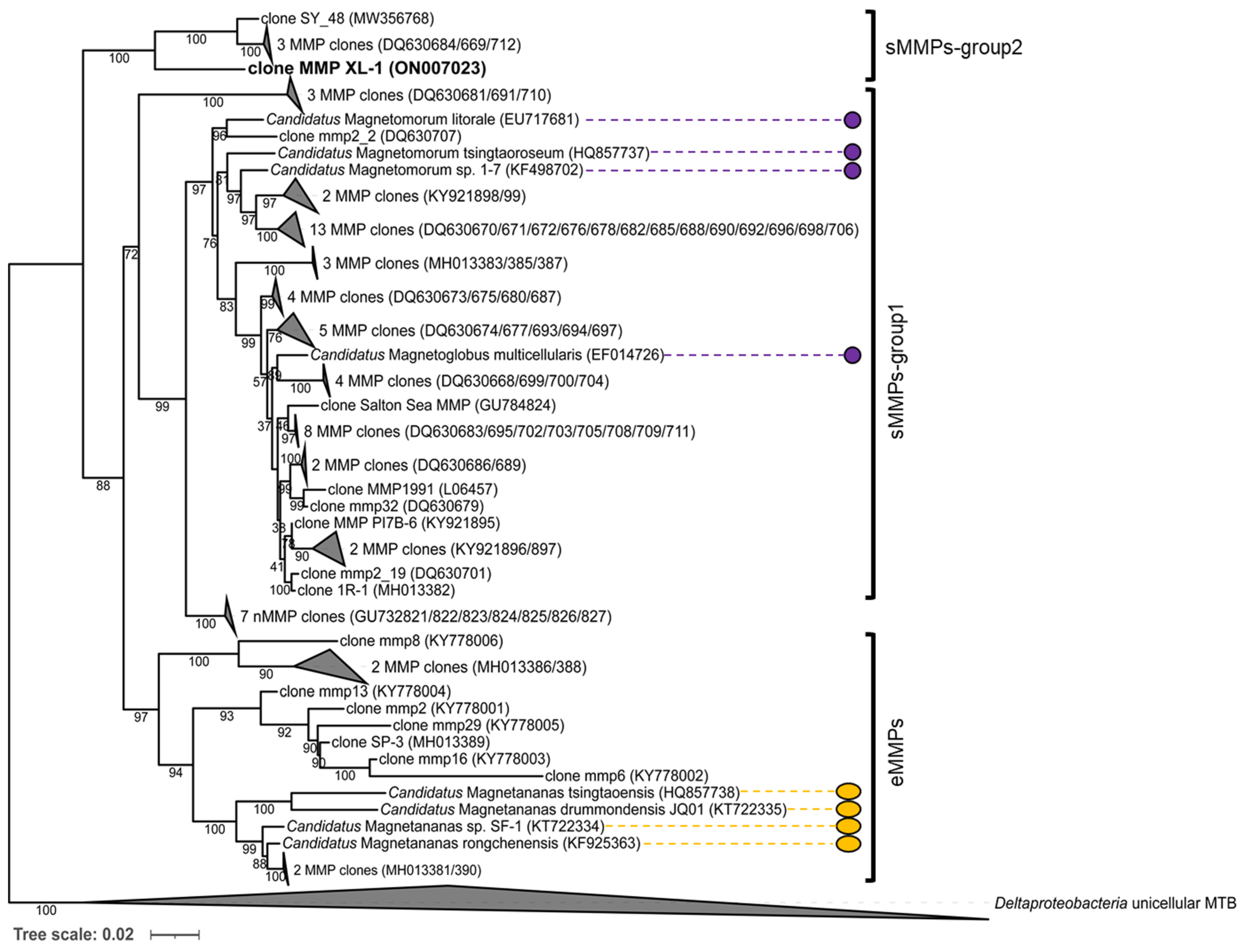
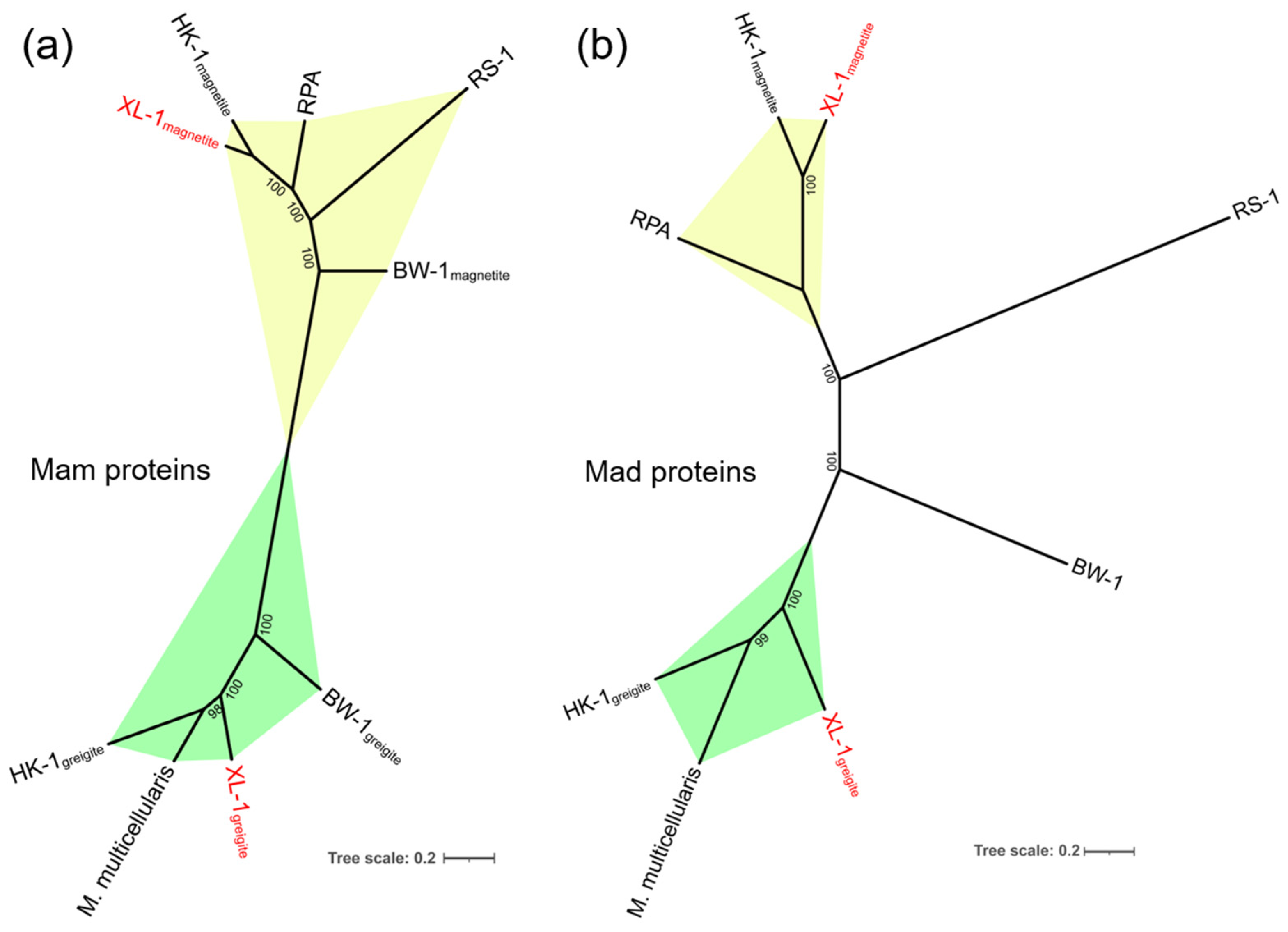
3.3. Phylogenetic Analysis of the sMMPs
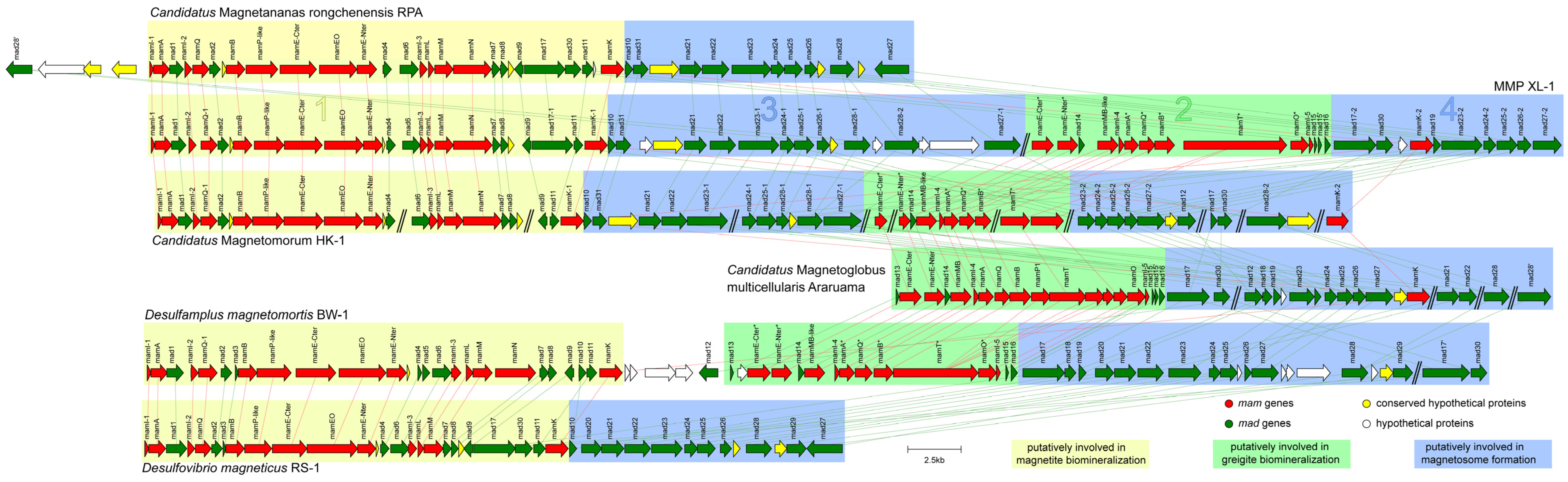
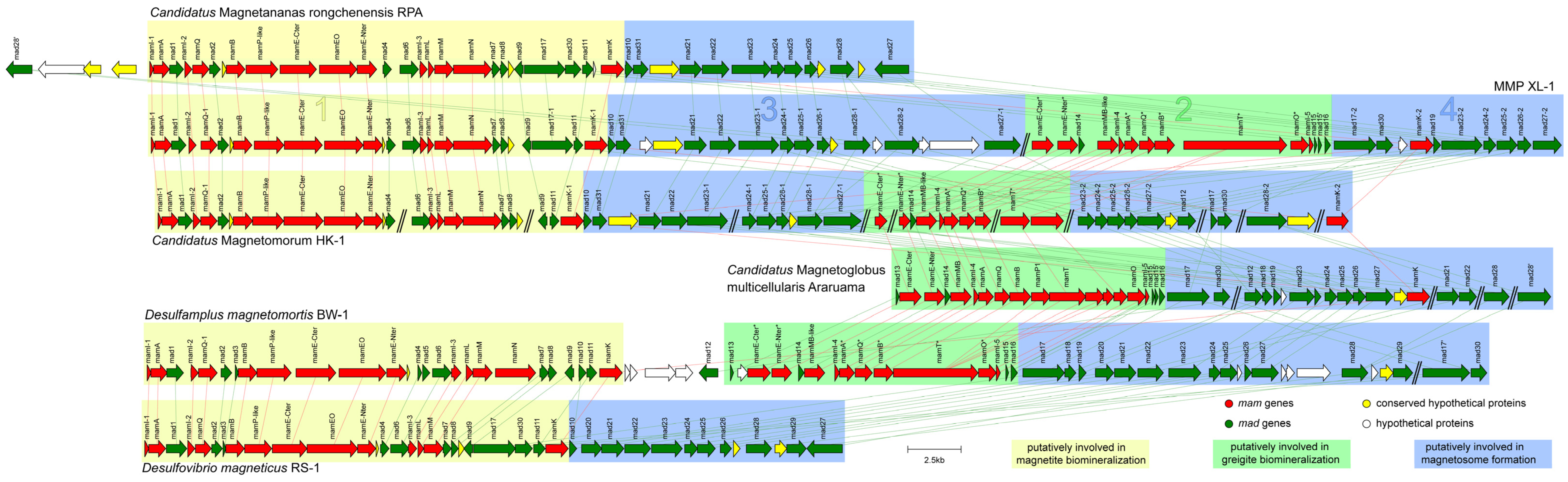
3.4. General Genomic Features of the Proposed MMP XL-1 and Comparative Genomic Analysis of Magnetosome Gene Clusters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blakemore, R. Magnetotactic bacteria. Science 1975, 190, 377–379. [Google Scholar] [CrossRef] [PubMed]
- Bazylinski, D.A.; Lefèvre, C.T.; Schüler, D. Magnetotactic bacteria. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 453–494. [Google Scholar] [CrossRef]
- Bazylinski, D.A.; Frankel, R.B. Magnetosome formation in prokaryotes. Nat. Rev. Microbiol. 2004, 2, 217–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spring, S.; Amann, R.; Ludwig, W.; Schleifer, K.H.; van Gemerden, H.; Petersen, N. Dominating role of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment. Appl. Environ. Microbiol. 1993, 59, 2397–2403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, S.L.; Sievert, S.M.; Frankel, R.B.; Bazylinski, D.A.; Edwards, K.J. Spatiotemporal distribution of marine magnetotactic bacteria in a seasonally stratified coastal salt pond. Appl. Environ. Microbiol. 2004, 70, 6230–6239. [Google Scholar] [CrossRef] [Green Version]
- Kolinko, S.; Jogler, C.; Katzmann, E.; Wanner, G.; Peplies, J.; Schüler, D. Single-cell analysis reveals a novel uncultivated magnetotactic bacterium within the candidate division OP3. Environ. Microbiol. 2012, 14, 1709–1721. [Google Scholar] [CrossRef]
- Lin, W.; Pan, Y. A putative greigite-type magnetosome gene cluster from the candidate phylum Latescibacteria. Environ. Microbiol. Rep. 2015, 7, 237–242. [Google Scholar] [CrossRef]
- Ji, B.; Zhang, S.D.; Zhang, W.J.; Rouy, Z.; Alberto, F.; Santini, C.L.; Mangenot, S.; Gagnot, S.; Philippe, N.; Pradel, N.; et al. The chimeric nature of the genomes of marine magnetotactic coccoid-ovoid bacteria defines a novel group of Proteobacteria. Environ. Microbiol. 2017, 19, 1103–1119. [Google Scholar] [CrossRef]
- Lin, W.; Pan, Y.; Bazylinski, D.A. Diversity and ecology of and biomineralization by magnetotactic bacteria. Environ. Microbiol. Rep. 2017, 9, 345–356. [Google Scholar] [CrossRef]
- Lin, W.; Zhang, W.; Zhao, X.; Roberts, A.P.; Paterson, G.A.; Bazylinski, D.A.; Pan, Y. Genomic expansion of magnetotactic bacteria reveals an early common origin of magnetotaxis with lineage-specific evolution. ISME J. 2018, 12, 1508–1519. [Google Scholar] [CrossRef] [Green Version]
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Micro. Microbiol. 2021, 71, 005056. [Google Scholar] [CrossRef]
- Waite, D.W.; Chuvochina, M.; Pelikan, C.; Parks, D.H.; Yilmaz, P.; Wagner, M.; Loy, A.; Naganuma, T.; Nakai, R.; Whitman, W.B.; et al. Proposal to reclassify the proteobacterial classes Deltaproteobacteria and Oligoflexia, and the phylum Thermodesulfobacteria into four phyla reflecting major functional capabilities. Int. J. Syst. Evol. Microbiol. 2020, 70, 5972–6016. [Google Scholar] [CrossRef] [PubMed]
- Uzun, M.; Alekseeva, L.; Krutkina, M.; Koziaeva, V.; Grouzdev, D. Unravelling the diversity of magnetotactic bacteria through analysis of open genomic databases. Sci. Data 2020, 7, 252. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Zhang, W.; Paterson, G.A.; Zhu, Q.; Zhao, X.; Knight, R.; Bazylinski, D.A.; Roberts, A.P.; Pan, Y. Expanding magnetic organelle biogenesis in the domain Bacteria. Microbiome 2020, 8, 152. [Google Scholar] [CrossRef] [PubMed]
- Farina, M.; de Barros, H.L.; Esquivel, D.M.S.; Danon, J. Ultrastructure of a magnetotactic microorganism. Biol. Cell 1983, 48, 85–88. [Google Scholar]
- Abreu, F.; Martins, J.L.; Silveira, T.S.; Keim, C.N.; de Barros, H.G.P.L.; Filho, F.J.G.; Lins, U. ‘Candidatus Magnetoglobus multicellularis’, a multicellular, magnetotactic prokaryote from a hypersaline environment. Int. J. Syst. Evol. Microbiol. 2007, 57, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Simmons, S.L.; Edwards, K.J. Unexpected diversity in populations of the many-celled magnetotactic prokaryote. Environ. Microbiol. 2007, 9, 206–215. [Google Scholar] [CrossRef]
- Wenter, R.; Wanner, G.; Schüler, D.; Overmann, J. Ultrastructure, tactic behaviour and potential for sulfate reduction of a novel multicellular magnetotactic prokaryote from North Sea sediments. Environ. Microbiol. 2009, 11, 1493–1505. [Google Scholar] [CrossRef]
- Zhou, K.; Pan, H.; Zhang, S.; Yue, H.; Xiao, T.; Wu, L. Occurrence and microscopic analyses of multicellular magnetotactic prokaryotes from coastal sediments in the Yellow Sea. Chin. J. Oceanol. Limn. 2011, 29, 246–251. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.; Zhang, W.Y.; Pan, H.M.; Li, J.H.; Yue, H.D.; Xiao, T.; Wu, L.F. Adaptation of spherical multicellular magnetotactic prokaryotes to the geochemically variable habitat of an intertidal zone. Environ. Microbiol. 2013, 15, 1595–1605. [Google Scholar] [CrossRef]
- Zhang, R.; Chen, Y.R.; Du, H.J.; Zhang, W.Y.; Pan, H.M.; Xiao, T.; Wu, L.F. Characterization and phylogenetic identification of a species of spherical multicellular magnetotactic prokaryotes that produces both magnetite and greigite crystals. Res. Microbiol. 2014, 165, 481–489. [Google Scholar] [CrossRef]
- Teng, Z.; Zhang, Y.; Zhang, W.; Pan, H.; Xu, J.; Huang, H.; Xiao, T.; Wu, L.F. Diversity and characterization of multicellular magnetotactic prokaryotes from coral reef habitats of the Paracel Islands, South China Sea. Front. Microbiol. 2018, 9, 2135. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Li, J.; Wu, L.-F.; Zhang, W.-J. Morphological and phylogenetic diversity of magnetotactic bacteria and multicellular magnetotactic prokaryotes from a mangrove ecosystem in the Sanya River, South China. J. Oceanol. Limnol. 2021, 39, 2015–2026. [Google Scholar] [CrossRef]
- Lefèvre, C.; Bernadac, A.; Pradel, N.; Wu, L.; Yu-Zhang, K.; Xiao, T.; Yonnet, J.-P.; Lebouc, A.; Song, T.; Fukumori, Y. Characterization of Mediterranean magnetotactic bacteria. J. Ocean. Univ. China 2007, 6, 355–359. [Google Scholar] [CrossRef]
- Zhou, K.; Zhang, W.-Y.; Yu-Zhang, K.; Pan, H.-M.; Zhang, S.-D.; Zhang, W.-J.; Yue, H.-D.; Li, Y.; Xiao, T.; Wu, L.-F. A novel genus of multicellular magnetotactic prokaryotes from the Yellow Sea. Environ. Microbiol. 2012, 14, 405–413. [Google Scholar] [CrossRef]
- Chen, Y.R.; Zhang, R.; Du, H.J.; Pan, H.M.; Zhang, W.Y.; Zhou, K.; Li, J.H.; Xiao, T.; Wu, L.F. A novel species of ellipsoidal multicellular magnetotactic prokaryotes from Lake Yuehu in China. Environ. Microbiol. 2015, 17, 637–647. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.R.; Zhang, W.Y.; Zhou, K.; Pan, H.M.; Du, H.J.; Xu, C.; Xu, J.H.; Pradel, N.; Santini, C.L.; Li, J.H.; et al. Novel species and expanded distribution of ellipsoidal multicellular magnetotactic prokaryotes. Environ. Microbiol. Rep. 2016, 8, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Qian, X.X.; Santini, C.L.; Kosta, A.; Menguy, N.; Le Guenno, H.; Zhang, W.; Li, J.; Chen, Y.R.; Liu, J.; Alberto, F.; et al. Juxtaposed membranes underpin cellular adhesion and display unilateral cell division of multicellular magnetotactic prokaryotes. Environ. Microbiol. 2020, 22, 1481–1494. [Google Scholar] [CrossRef]
- Keim, C.N.; Abreu, F.; Lins, U.; de Barros, H.L.; Farina, M. Cell organization and ultrastructure of a magnetotactic multicellular organism. J. Struct. Biol. 2004, 145, 254–262. [Google Scholar] [CrossRef]
- Abreu, F.; Silva, K.T.; Leão, P.; Guedes, I.A.; Keim, C.N.; Farina, M.; Lins, U. Cell adhesion, multicellular morphology, and magnetosome distribution in the multicellular magnetotactic prokaryote Candidatus Magnetoglobus multicellularis. Microsc. Microanal. 2013, 19, 535–543. [Google Scholar] [CrossRef] [PubMed]
- Kolinko, S.; Richter, M.; Glöckner, F.-O.; Brachmann, A.; Schüler, D. Single-cell genomics reveals potential for magnetite and greigite biomineralization in an uncultivated multicellular magnetotactic prokaryote. Environ. Microbiol. Rep. 2014, 6, 524–531. [Google Scholar] [CrossRef]
- Lefèvre, C.T.; Abreu, F.; Lins, U.; Bazylinski, D.A. Nonmagnetotactic multicellular prokaryotes from low-saline, nonmarine aquatic environments and their unusual negative phototactic behavior. Appl. Environ. Microbiol. 2010, 76, 3220–3227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Zhang, W.; Du, H.; Leng, X.; Li, J.H.; Pan, H.; Xu, J.; Wu, L.F.; Xiao, T. Seasonal changes in the vertical distribution of two types of multicellular magnetotactic prokaryotes in the sediment of Lake Yuehu, China. Environ. Microbiol. Rep. 2018, 10, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Du, H.J.; Chen, Y.R.; Zhang, R.; Pan, H.M.; Zhang, W.Y.; Zhou, K.; Wu, L.F.; Xiao, T. Temporal distributions and environmental adaptations of two types of multicellular magnetotactic prokaryote in the sediments of Lake Yuehu, China. Environ. Microbiol. Rep. 2015, 7, 538–546. [Google Scholar] [CrossRef]
- Abreu, F.; Morillo, V.; Nascimento, F.F.; Werneck, C.; Cantão, M.E.; Ciapina, L.P.; de Almeida, L.G.; Lefèvre, C.T.; Bazylinski, D.A.; de Vasconcelos, A.T.R.; et al. Deciphering unusual uncultured magnetotactic multicellular prokaryotes through genomics. ISME J. 2014, 8, 1055–1068. [Google Scholar] [CrossRef] [Green Version]
- Grünberg, K.; Wawer, C.; Tebo, B.M.; Schüler, D. A large gene cluster encoding several magnetosome proteins is conserved in different species of magnetotactic bacteria. Appl. Environ. Microbiol. 2001, 67, 4573–4582. [Google Scholar] [CrossRef] [Green Version]
- Murat, D.; Quinlan, A.; Vali, H.; Komeili, A. Comprehensive genetic dissection of the magnetosome gene island reveals the step-wise assembly of a prokaryotic organelle. Proc. Natl. Acad. Sci. USA 2010, 107, 5593–5598. [Google Scholar] [CrossRef] [Green Version]
- Lohße, A.; Ullrich, S.; Katzmann, E.; Borg, S.; Wanner, G.; Richter, M.; Voigt, B.; Schweder, T.; Schüler, D. Functional analysis of the magnetosome island in Magnetospirillum gryphiswaldense: The mamAB operon is sufficient for magnetite biomineralization. PLoS ONE 2011, 6, e25561. [Google Scholar] [CrossRef] [Green Version]
- Abreu, F.; Cantao, M.E.; Nicolás, M.F.; Barcellos, F.G.; Morillo, V.; Almeida, L.G.; Do Nascimento, F.F.; Lefevre, C.T.; Bazylinski, D.A.; de Vasconcelos, A.T.R.; et al. Common ancestry of iron oxide- and iron-sulfide-based biomineralization in magnetotactic bacteria. ISME J. 2011, 5, 1634–1640. [Google Scholar] [CrossRef]
- Leão, P.; Chen, Y.R.; Abreu, F.; Wang, M.; Zhang, W.J.; Zhou, K.; Xiao, T.; Wu, L.F.; Lins, U. Ultrastructure of ellipsoidal magnetotactic multicellular prokaryotes depicts their complex assemblage and cellular polarity in the context of magnetotaxis. Environ. Microbiol. 2017, 19, 2151–2163. [Google Scholar] [CrossRef]
- Nakazawa, H.; Arakaki, A.; Narita-Yamada, S.; Yashiro, I.; Jinno, K.; Aoki, N.; Tsuruyama, A.; Okamura, Y.; Tanikawa, S.; Fujita, N.; et al. Whole genome sequence of Desulfovibrio magneticus strain RS-1 revealed common gene clusters in magnetotactic bacteria. Genome. Res. 2009, 19, 1801–1808. [Google Scholar] [CrossRef] [Green Version]
- Lefèvre, C.T.; Menguy, N.; Abreu, F.; Lins, U.; Pósfai, M.; Prozorov, T.; Pignol, D.; Frankel, R.B.; Bazylinski, D.A. A cultured greigite-producing magnetotactic bacterium in a novel group of sulfate-reducing bacteria. Science 2011, 334, 1720–1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakaguchi, T.; Arakaki, A.; Matsunaga, T. Desulfovibrio magneticus sp. nov., a novel sulfate-reducing bacterium that produces intracellular single-domain-sized magnetite particles. Int. J. Syst. Evol. Microbiol. 2002, 52, 215–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flies, C.B.; Peplies, J.; Schüler, D. Combined approach for characterization of uncultivated magnetotactic bacteria from various aquatic environments. Appl. Environ. Microbiol. 2005, 71, 2723–2731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfe, R.S.; Thauer, R.K.; Pfennig, N. A ‘capillary racetrack’ method for isolation of magnetotactic bacteria. Fems. Microbiol. Ecol. 1987, 45, 31–35. [Google Scholar] [CrossRef]
- Schüler, D. The biomineralization of magnetosomes in Magnetospirillum gryphiswaldense. Int. Microbiol. 2002, 5, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Jogler, C.; Wanner, G.; Kolinko, S.; Niebler, M.; Amann, R.; Petersen, N.; Kube, M.; Reinhardt, R.; Schüler, D. Conservation of proteobacterial magnetosome genes and structures in an uncultivated member of the deep-branching Nitrospira phylum. Proc. Natl. Acad. Sci. USA 2011, 108, 1134–1139. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The Ribosomal Database Project: Improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef] [Green Version]
- Pan, H.; Zhu, K.; Song, T.; Yu-Zhang, K.; Lefèvre, C.; Xing, S.; Liu, M.; Zhao, S.; Xiao, T.; Wu, L.F. Characterization of a homogeneous taxonomic group of marine magnetotactic cocci within a low tide zone in the China Sea. Environ. Microbiol. 2008, 10, 1158–1164. [Google Scholar] [CrossRef]
- Zhang, W.Y.; Zhou, K.; Pan, H.M.; Yue, H.D.; Jiang, M.; Xiao, T.; Wu, L.F. Two genera of magnetococci with bean-like morphology from intertidal sediments of the Yellow Sea, China. Appl. Environ. Microb. 2012, 78, 5606–5611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pernthaler, J.; Glöckner, F.O.; Schönhuber, W.; Amann, R. Fluorescence in situ hybridization (FISH) with rRNA-targeted oligonucleotide probes. Method Microbiol. 2001, 30, 207–226. [Google Scholar] [CrossRef]
- Li, J.; Zhang, H.; Menguy, N.; Benzerara, K.; Wang, F.; Lin, X.; Chen, Z.; Pan, Y. Single-cell resolution of uncultured magnetotactic bacteria via fluorescence-coupled electron microscopy. Appl. Environ. Microbiol. 2017, 83, e00409-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Chen, Y.; Shi, C.; Huang, Z.; Zhang, Y.; Li, S.; Li, Y.; Ye, J.; Yu, C.; Li, Z.; et al. SOAPnuke: A MapReduce acceleration-supported software for integrated quality control and preprocessing of high-throughput sequencing data. Gigascience 2018, 7, gix118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Luo, R.; Liu, C.M.; Leung, C.M.; Ting, H.F.; Sadakane, K.; Yamashita, H.; Lam, T.W. MEGAHIT v1.0: A fast and scalable metagenome assembler driven by advanced methodologies and community practices. Methods 2016, 102, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Uritskiy, G.V.; DiRuggiero, J.; Taylor, J. MetaWRAP-a flexible pipeline for genome-resolved metagenomic data analysis. Microbiome 2018, 6, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Ji, R.; Zhang, W.; Pan, Y.; Lin, W. MagCluster: A tool for identification, annotation, and visualization of magnetosome gene clusters. Microbiol. Resour. Ann. 2022, 11, e01031-21. [Google Scholar] [CrossRef]
- Gilchrist, C.L.M.; Chooi, Y.H. Clinker & clustermap.js: Automatic generation of gene cluster comparison figures. Bioinformatics 2021, 37, 2473–2475. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.H.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefèvre, C.T.; Trubitsyn, D.; Abreu, F.; Kolinko, S.; Jogler, C.; de Almeida, L.G.P.; de Vasconcelos, A.T.R.; Kube, M.; Reinhardt, R.; Lins, U.; et al. Comparative genomic analysis of magnetotactic bacteria from the Deltaproteobacteria provides new insights into magnetite and greigite magnetosome genes required for magnetotaxis. Environ. Microbiol. 2013, 15, 2712–2735. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef]
- Keim, C.N.; Martins, J.L.; Abreu, F.; Rosado, A.S.; de Barros, H.L.; Borojevic, R.; Lins, U.; Farina, M. Multicellular life cycle of magnetotactic prokaryotes. FEMS Microbiol. Lett. 2004, 240, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Lins, U.; Keim, C.N.; Evans, F.F.; Farina, M.; Buseck, P.R. Magnetite (Fe3O4) and greigite (Fe3S4) crystals in multicellular magnetotactic prokaryotes. Geomicrobiol. J. 2007, 24, 43–50. [Google Scholar] [CrossRef]
- Moskowitz, B.M.; Bazylinski, D.A.; Egli, R.; Frankel, R.B.; Edwards, K.J. Magnetic properties of marine magnetotactic bacteria in a seasonally stratified coastal pond (Salt Pond, MA, USA). Geophys. J. Int. 2008, 174, 75–92. [Google Scholar] [CrossRef]
- Descamps, E.C.T.; Monteil, C.L.; Menguy, N.; Ginet, N.; Pignol, D.; Bazylinski, D.A.; Lefèvre, C.T. Desulfamplus magnetovallimortis gen. nov., sp. nov., a magnetotactic bacterium from a brackish desert spring able to biomineralize greigite and magnetite, that represents a novel lineage in the Desulfobacteraceae. Syst. Appl. Microbiol. 2017, 40, 280–289. [Google Scholar] [CrossRef]
- Lefèvre, C.T.; Trubitsyn, D.; Abreu, F.; Kolinko, S.; de Almeida, L.G.; de Vasconcelos, A.T.; Lins, U.; Schüler, D.; Ginet, N.; Pignol, D.; et al. Monophyletic origin of magnetotaxis and the first magnetosomes. Environ. Microbiol. 2013, 15, 2267–2274. [Google Scholar] [CrossRef]
- Lefèvre, C.T.; Wu, L.-F. Evolution of the bacterial organelle responsible for magnetotaxis. Trends Microbiol. 2013, 21, 534–543. [Google Scholar] [CrossRef]
- Kolinko, S.; Richter, M.; Glöckner, F.O.; Brachmann, A.; Schüler, D. Single-cell genomics of uncultivated deep-branching magnetotactic bacteria reveals a conserved set of magnetosome genes. Environ. Microbiol. 2016, 18, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, Y.; Liu, L.; Pan, Y.; Lin, W. Identification and genomic characterization of two previously unknown magnetotactic Nitrospirae. Front. Microbiol. 2021, 12, 690052. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteins in Magnetite Biomineralization | Accession Number | Proteins in Greigite Biomineralization | Accession Number | Identity (%) | E-Value |
|---|---|---|---|---|---|
| MamI-1 | XL1_145_00043 | MamI-4 | XL1_87_00005 | 62.50 | 2.68e-06 |
| MamA | XL1_145_00042 | MamA * | XL1_87_00006 | 46.34 | 8.62e-19 |
| MamI-2 | XL1_145_00040 | MamI-5 | XL1_87_00011 | 64.10 | 7.65e-07 |
| MamQ-1 | XL1_145_00039 | MamQ * | XL1_87_00007 | 46.28 | 3.82e-18 |
| MamB | XL1_145_00036 | MamB * | XL1_87_00008 | 58.93 | 1.22e-61 |
| MamP-like | XL1_145_00035 | MamT * | XL1_87_00009 | 55.75 | 1.65e-14 |
| MamE-Cter | XL1_145_00034 | MamE-Cter * | XL1_87_00001 | 52.70 | 4.36e-23 |
| MamEO | XL1_145_00033 | MamO * | XL1_87_00010 | 61.54 | 1.31e-49 |
| MamE-Nter | XL1_145_00032 | MamE-Nter * | XL1_87_00002 | 65.84 | 1.24e-42 |
| MamM | XL1_145_00026 | MamMB-like | XL1_87_00004 | 48.43 | 2.86e-21 |
| Mad17-1 | XL1_145_00020 | Mad17-2 | XL1_87_00015 | 72.15 | 0 |
| MamK-1 | XL1_145_00018 | MamK-2 | XL1_87_00018 | 83.81 | 1.73e-179 |
| Mad23-1 | XL1_145_00011 | Mad23-2 | XL1_87_00020 | 58.28 | 1.23e-95 |
| Mad24-1 | XL1_145_00010 | Mad24-2 | XL1_87_00021 | 52.12 | 9.87e-14 |
| Mad25-1 | XL1_145_00009 | Mad25-2 | XL1_87_00022 | 62.87 | 1.12e-47 |
| Mad26-1 | XL1_145_00008 | Mad26-2 | XL1_87_00023 | 52.73 | 4.27e-10 |
| Mad27-1 | XL1_145_00001 | Mad27-2 | XL1_87_00024 | 71.36 | 7.13e-156 |
| Morphology of MTB | Sampling Site | Cell Diameter/Size (μm) | Magnetotaxis Motility (μm/s) | “Ping-Pong” Motility | Magnetosome Characteristics | MGC | Detected sMMP Strain | References | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Excursion (μm/s) | Return (μm/s) | Type | Shape | Composition | Size (nm) | Proportion of Magnetite (%) | |||||||
| sMMP | Itaipu Lagoon, Brazil (43°04′ W, 22°57′ S) | - | - | - | - | Type-I | Bullet-shaped | Magnetite | 104 ± 29 (L) 42 ± 6 (W) | - | - | - | [67] |
| Type-II | Bullet-shaped | Magnetite | 95 ± 23 (L) 38 ± 5 (W) | 15–96 | |||||||||
| Irregularly shaped | Greigite | 72 ± 8 (L) | |||||||||||
| Type-III | Irregularly shaped | Greigite | 70 ± 8 (L) | - | |||||||||
| Araruama Lagoon, Brazil (42°13′ W, 22°50′ S) | 6.0–9.5 | 90 ± 20 | - | - | Type-III | Irregularly shaped | Greigite | 88 (L) × 71 (W) | - | Greigite biomineralization | Ca. Magnetoglobus multicellularis (EF014726) | [16,39] | |
| Wadden Sea, northern Germany (53°53.555′ N, 8°40.565′ E) | 5.7 ± 1.1 | - | - | - | Type-IV | Bullet-shaped | Greigite | 91 ± 21 (L) 40 ± 6 (W) | - | - | Ca. Magnetomorum litorale (EU717681) | [18] | |
| Wadden Sea, northern Germany (53°53.53′ N, 8°40.75′ E) | - | - | - | - | - | - | - | - | - | Magnetite and greigite biomineralization | Ca. Magnetomorum HK-1 (GCA_001292585) | [31] | |
| Yuehu Lake, China (37°21′ N, 122°33′ E) | 5.5 ± 0.8 | - | - | - | Type-I | Bullet-shaped | Magnetite | 92 ± 27 (L) 29 ± 5 (W) | - | - | - | [19] | |
| 5.6 ± 0.9 | 37 ± 20 | 124 ± 53 | 93 ± 39 | Type-II | Bullet-shaped | Magnetite | 80.1 ± 16.1 (L) 33.6 ± 3.5 (W) | 21.8–64.8 | - | Ca. Magnetomorum rongchengroseum (KF498702) | [21] | ||
| Irregularly shaped | Greigite | 63.9 ± 9.3 (L) 52.5 ± 7.5 (W) | |||||||||||
| Huiquan Bay, China (36°03′ N, 120°21′ E) | 5.5 ± 0.8 | 55 | - | - | Type-I | Bullet-shaped | Magnetite | 92 ± 20 (L) 35 ± 4 (W) | - | - | Ca. Magnetomorum tsingtaoroseum (HQ857737) | [20] | |
| Sanya Mangrove, China (18°15.242′ N, 109°30.585′ E) | 4.6 ± 0.2 | - | - | - | Type-I | Bullet-shaped | Magnetite | 78 ± 18 (L) 34 ± 4 (W) | - | - | SY_5 (MW356767) SY_48 (MW356768) | [23] | |
| Type-II | Bullet-shaped | Magnetite | 88 ± 19 (L) 34 ± 5 (W) | - | |||||||||
| Irregularly shaped | Greigite | 80 ± 19 (L) | |||||||||||
| Type-III | Irregularly shaped | Greigite | 77 ± 11 (L) | - | |||||||||
| Jinsha Bay, China (21°16.267′ N, 110°24.067′ E) | 4.78 ± 0.67 | 78.0 ± 41.4 | 223.9 ± 54.5 | 102.2 ± 19.0 | Type-II | Bullet-shaped | Magnetite | 87.0 ± 20.3 (L) 35.2 ± 3.5 (W) | 8.2–82.0 | Magnetite and greigite biomineralization | MMP XL-1 | This study | |
| Irregularly shaped | Greigite | 72.8 ± 8.7 (L) 55.2 ± 7.3 (W) | |||||||||||
| eMMP | Huiquan Bay, China (36°03′ N, 120°21′ E) | 9.6 ± 1.2 × 7.8 ± 0.9 | 99 ± 50 | - | - | Type-I | Bullet-shaped | Magnetite | 102 ± 24 (L) 38 ± 6 (W) | - | - | Ca. Magnetananas tsingtaoensis (HQ857738) | [25] |
| Yuehu Lake, China (37°21′ N, 122°34′ E) | 9.18 ± 1.01 × 7.41 ± 0.76 | 77 ± 33 | 223 ± 27 | 169 ± 27 | Type-I | Bullet-shaped | Magnetite | 115 ± 27 (L) 39 ± 5 (W) | - | Magnetite biomineralization | Ca. Magnetananas rongchenensis (KF925363) | [26,40] | |
| Type-II | Bullet-shaped | Magnetite | - | ||||||||||
| Irregularly shaped | Greigite | 102 ± 14 (L) 78 ± 13 (W) | |||||||||||
| Rod-shaped | Badwater Basin, America | - | 30 | - | - | Type-II | Bullet-shaped | Magnetite | - | - | Magnetite and greigite biomineralization | Desulfamplus magnetovallimortis BW-1 (JN252194) | [42,69] |
| Irregularly shaped | Greigite | - | |||||||||||
| Vibrio-shaped | Kameno River waterway, Japan | 3–5 × 1 | - | - | - | Type-I | Bullet-shaped | Magnetite | - | - | Magnetite biomineralization | Desulfovibrio magneticus RS-1 (NR_027575) | [41,43] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, K.; Pan, H.; Chen, J.; Liu, J.; Zhao, Y.; Chen, S.; Zhang, W.; Xiao, T.; Wu, L.-F. A Novel Isolate of Spherical Multicellular Magnetotactic Prokaryotes Has Two Magnetosome Gene Clusters and Synthesizes Both Magnetite and Greigite Crystals. Microorganisms 2022, 10, 925. https://doi.org/10.3390/microorganisms10050925
Cui K, Pan H, Chen J, Liu J, Zhao Y, Chen S, Zhang W, Xiao T, Wu L-F. A Novel Isolate of Spherical Multicellular Magnetotactic Prokaryotes Has Two Magnetosome Gene Clusters and Synthesizes Both Magnetite and Greigite Crystals. Microorganisms. 2022; 10(5):925. https://doi.org/10.3390/microorganisms10050925
Chicago/Turabian StyleCui, Kaixuan, Hongmiao Pan, Jianwei Chen, Jia Liu, Yicong Zhao, Si Chen, Wenyan Zhang, Tian Xiao, and Long-Fei Wu. 2022. "A Novel Isolate of Spherical Multicellular Magnetotactic Prokaryotes Has Two Magnetosome Gene Clusters and Synthesizes Both Magnetite and Greigite Crystals" Microorganisms 10, no. 5: 925. https://doi.org/10.3390/microorganisms10050925
APA StyleCui, K., Pan, H., Chen, J., Liu, J., Zhao, Y., Chen, S., Zhang, W., Xiao, T., & Wu, L.-F. (2022). A Novel Isolate of Spherical Multicellular Magnetotactic Prokaryotes Has Two Magnetosome Gene Clusters and Synthesizes Both Magnetite and Greigite Crystals. Microorganisms, 10(5), 925. https://doi.org/10.3390/microorganisms10050925