
Potential Probiotic Lacticaseibacillus paracasei MJM60396 Prevents Hyperuricemia in a Multiple Way by Absorbing Purine, Suppressing Xanthine Oxidase and Regulating Urate Excretion in Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Screening of Purine Degrading Lactobacillus Strains
2.1.1. Incubation of Lactic Acid Bacteria (LAB) in Nucleoside Solution
2.1.2. HPLC Analysis
2.1.3. Degradation of Purine Compounds by the Cell Lysates of MJM60396 and MJM60662
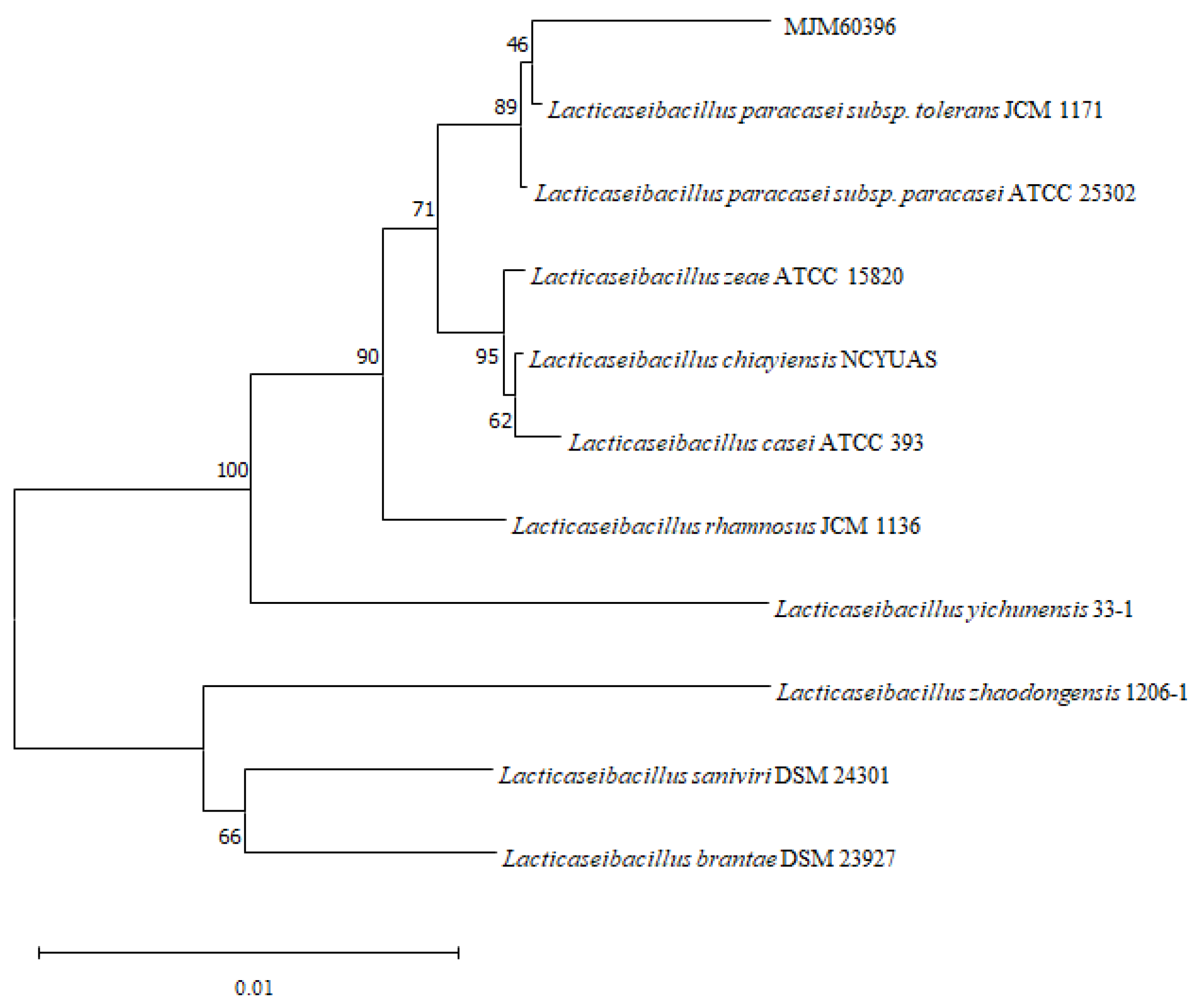
2.2. Strain Identification by 16S rDNA Sequence and Phylogenetic Analysis of MJM60396
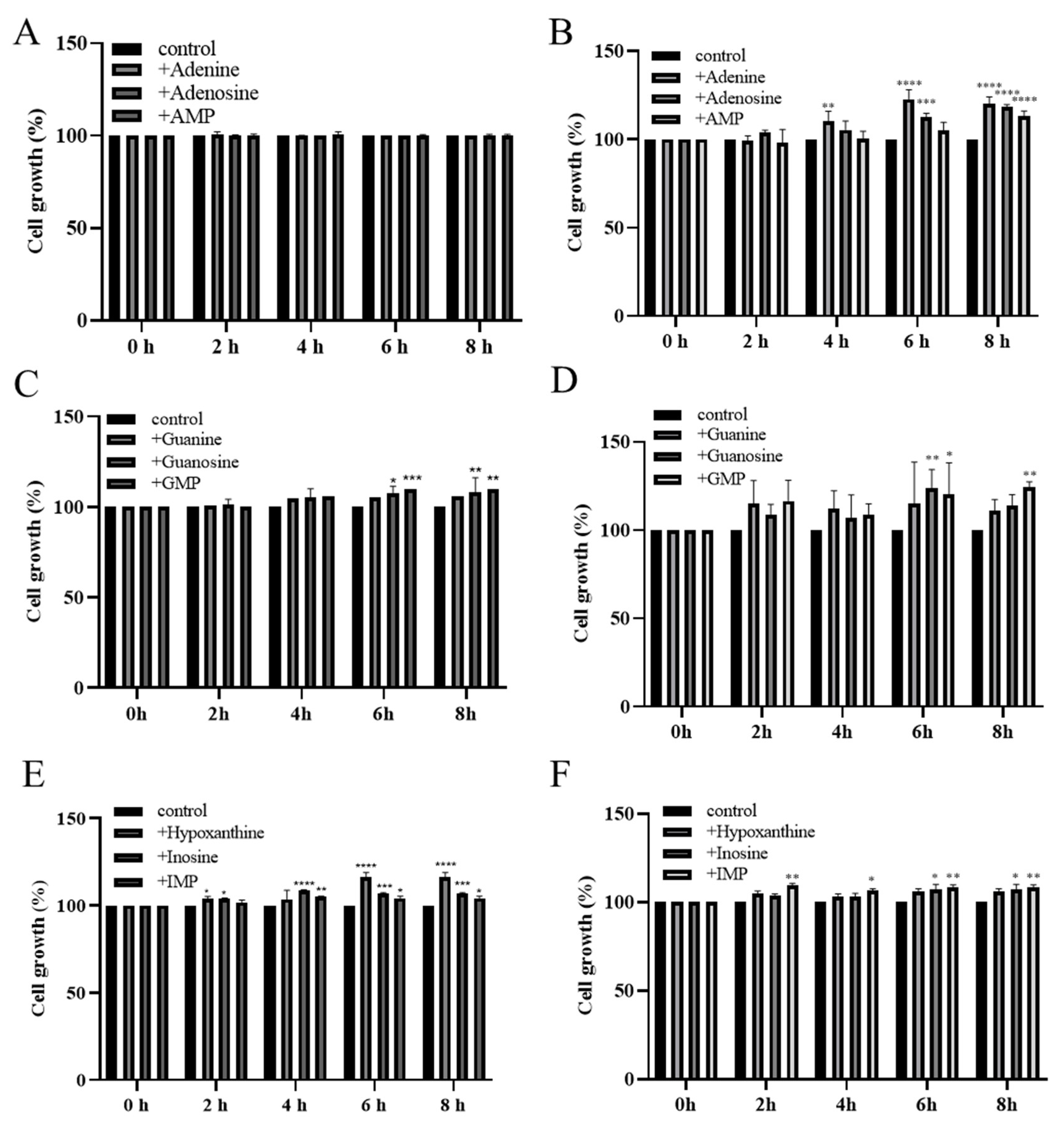
2.3. Utilization of Purine Compounds by MJM60396, MJM60662
2.4. Animal Study
2.4.1. Animal Monitoring and Treatment
2.4.2. Serum Biochemical Analysis
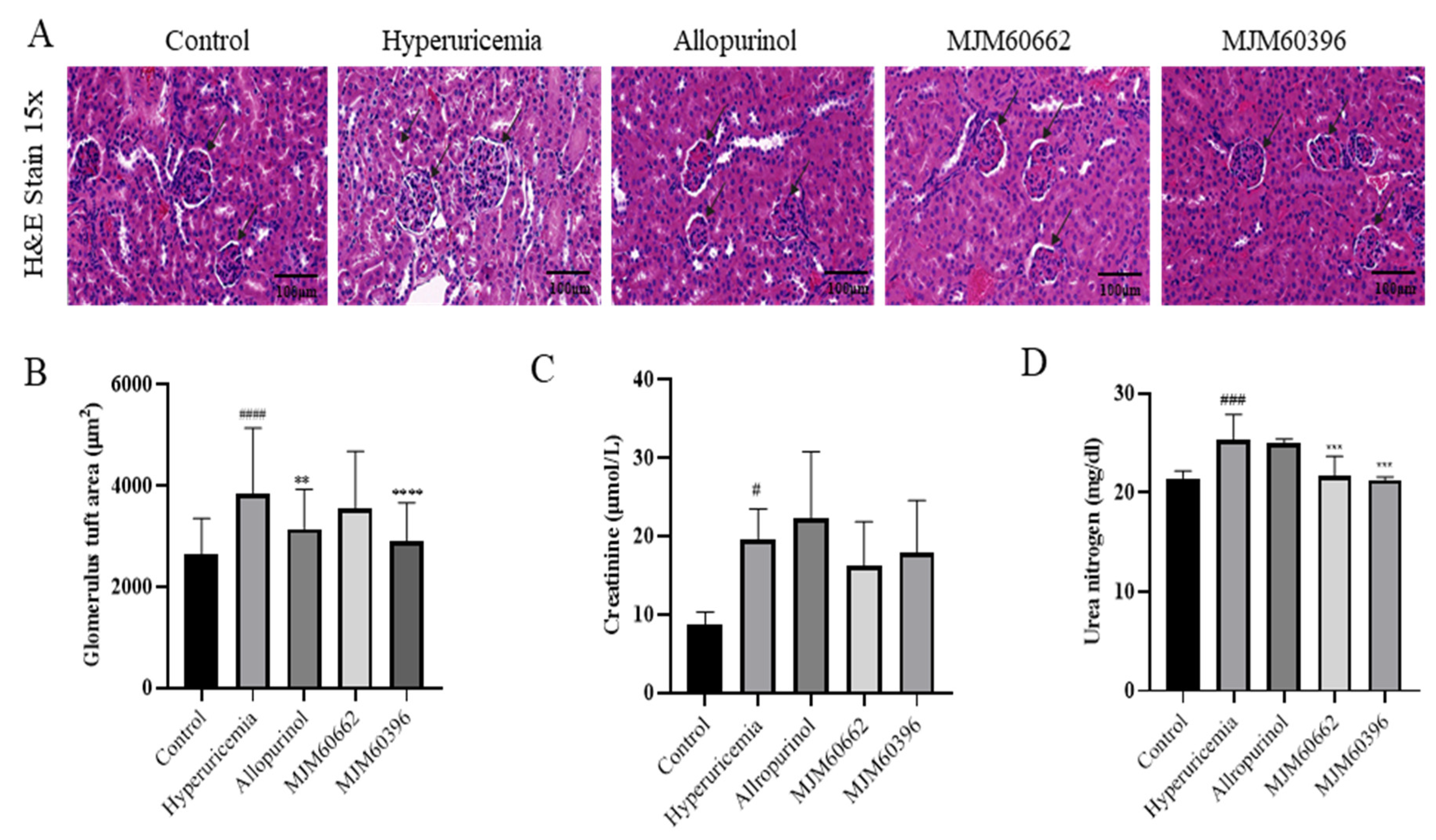
2.4.3. Kidney, Intestine Sectioning and Histopathological Assessment
2.4.4. Fecal Sample Analysis
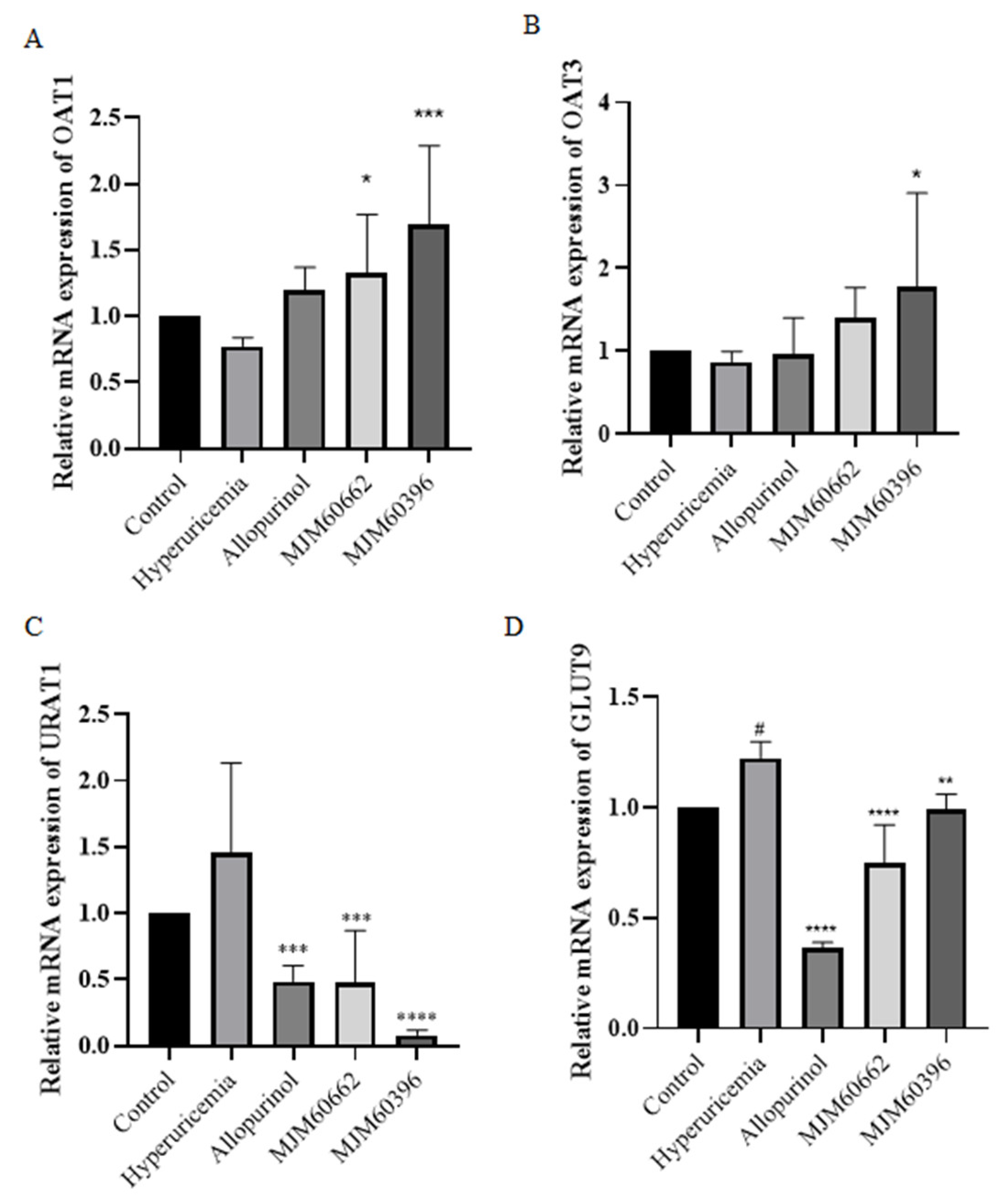
2.4.5. RNA Extraction, cDNA Synthesis, and Quantitative Real-Time PCR (qRT-PCR)
2.4.6. ZO-1 and Occludin Expression
2.5. Statistical Analysis
3. Result
3.1. Screening of Inosine, Guanosine, and Adenosine Degrading LAB Strains
3.1.1. Assimilation of Guanine and Inosine by LAB Strains
3.1.2. Degradation of Purine Compounds by Cell Lysates
3.2. Phylogenetic Analysis of MJM60396
3.3. Utilization of Purine Compounds by LAB for Growth
3.4. Animal Study
3.4.1. Effect of MJM60396 on the Body Weight and the Serum Uric Acid Level
3.4.2. Effect of LAB on the Kidney by Histological Observation and Biochemistry Analysis
3.4.3. Effect of MJM60396 on mRNA, Protein Expression of Kidney Genes Associated with Hyperuricemia
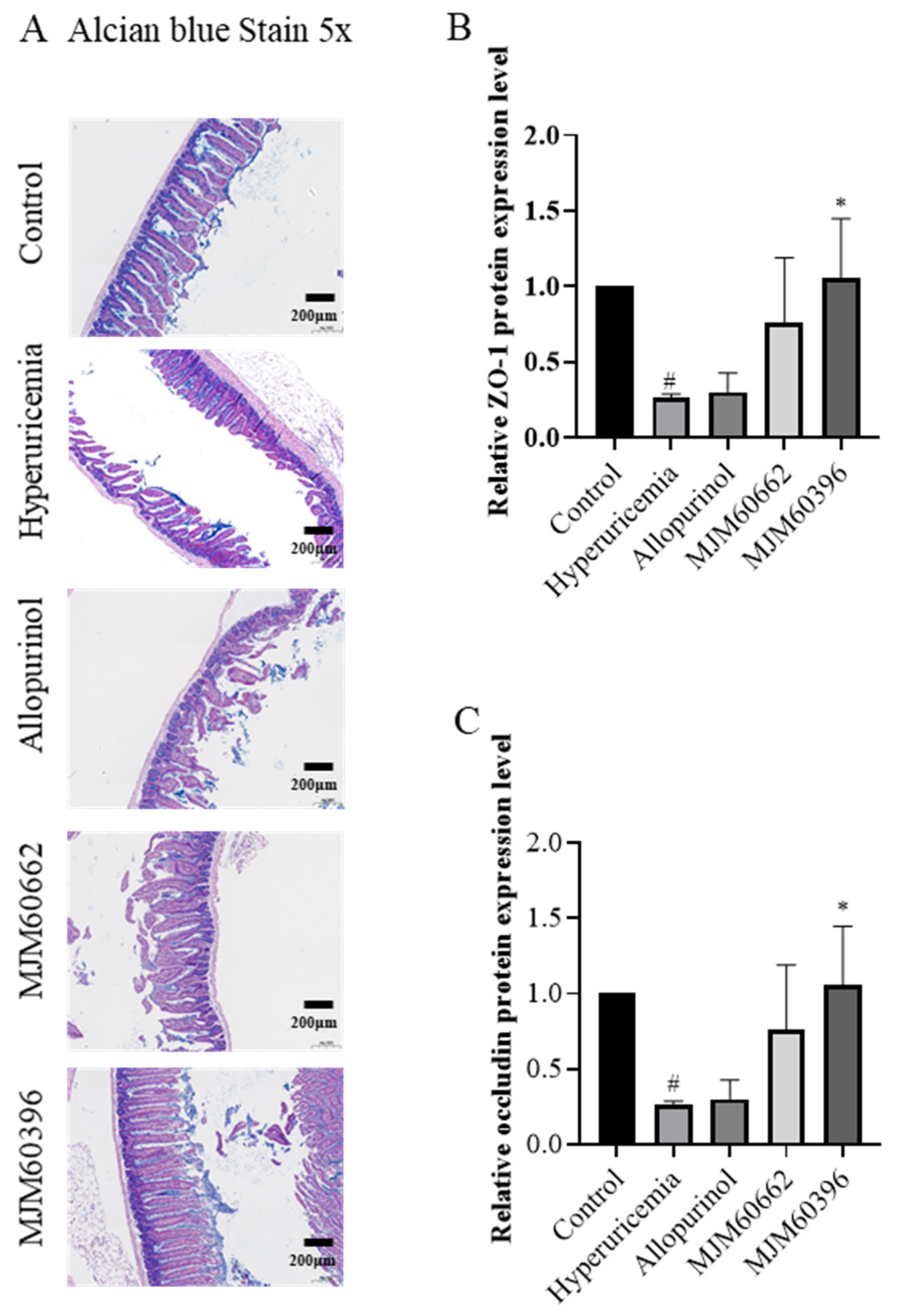
3.4.4. Alcian Blue Staining and Expression of Tight Junction Proteins in the Intestine
3.4.5. Intestinal Microbial Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ying, W.; Ze, Y. Limosilactobacillus Fermentum JL-3 Isolated from “Jiangshui” Ameliorates Hyperuricemia by Degrading Uric Acid. Available online: https://www.tandfonline.com/doi/full/10.1080/19490976.2021.1897211 (accessed on 28 October 2021).
- De Oliveira, E.P.; Burini, R.C. High Plasma Uric Acid Concentration: Causes and Consequences. Diabetol. Metab. Syndr. 2012, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.W.; Muzny, D.M.; Lee, C.C.; Caskey, C.T. Two Independent Mutational Events in the Loss of Urate Oxidase during Hominoid Evolution. J. Mol. Evol. 1992, 34, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Benedict, J.D.; Forsham, P.H.; Stetten, D. The Metabolism of Uric Acid in the Normal and Gouty Human Studied with the Aid of Isotopic Uric Acid. J. Biol. Chem. 1949, 181, 183–193. [Google Scholar] [CrossRef]
- Garrel, D.R.; Verdy, M.; PetitClerc, C.; Martin, C.; Brulé, D.; Hamet, P. Milk- and Soy-Protein Ingestion: Acute Effect on Serum Uric Acid Concentration. Am. J. Clin. Nutr. 1991, 53, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Wang, H.; Chen, T. Interactions between Intestinal Microflora/Probiotics and the Immune System. BioMed Res. Int. 2019, 2019, e6764919. [Google Scholar] [CrossRef] [Green Version]
- Delcenserie, V.; Martel, D.; Lamoureux, M.; Amiot, J.; Boutin, Y.; Roy, D. Immunomodulatory Effects of Probiotics in the Intestinal Tract. Curr. Issues Mol. Biol. 2008, 10, 37–54. [Google Scholar] [CrossRef] [Green Version]
- Hassan, A.; Din, A.U.; Zhu, Y.; Zhang, K.; Li, T.; Wang, Y.; Luo, Y.; Wang, G. Updates in Understanding the Hypocholesterolemia Effect of Probiotics on Atherosclerosis. Appl. Microbiol. Biotechnol. 2019, 103, 5993–6006. [Google Scholar] [CrossRef]
- Moumita, S.; Das, B.; Sundaray, A.; Satpathi, S.; Thangaraj, P.; Marimuthu, S.; Jayabalan, R. Study of Soy-Fortified Green Tea Curd Formulated Using Potential Hypocholesterolemic and Hypotensive Probiotics Isolated from Locally Made Curd. Food Chem. 2018, 268, 558–566. [Google Scholar] [CrossRef]
- Vivarelli, S.; Falzone, L.; Basile, M.S.; Nicolosi, D.; Genovese, C.; Libra, M.; Salmeri, M. Benefits of Using Probiotics as Adjuvants in Anticancer Therapy (Review). World Acad. Sci. J. 2019, 1, 125–135. [Google Scholar] [CrossRef]
- Guandalini, S. Probiotics for Prevention and Treatment of Diarrhea. J. Clin. Gastroenterol. 2011, 45, S149. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.-Y.; Yin, T.-L.; Zhou, J.; Xu, J.; Lu, X.-J. Gut Microbiome and Cancer Immunotherapy. J. Cell. Physiol. 2020, 235, 4082–4088. [Google Scholar] [CrossRef] [PubMed]
- Kaźmierczak-Siedlecka, K.; Daca, A.; Fic, M.; van de Wetering, T.; Folwarski, M.; Makarewicz, W. Therapeutic Methods of Gut Microbiota Modification in Colorectal Cancer Management—Fecal Microbiota Transplantation, Prebiotics, Probiotics, and Synbiotics. Gut Microbes 2020, 11, 1518–1530. [Google Scholar] [CrossRef] [PubMed]
- Yamada, N.; Saito-Iwamoto, C.; Nakamura, M.; Soeda, M.; Chiba, Y.; Kano, H.; Asami, Y. Lactobacillus Gasseri PA-3 Uses the Purines IMP, Inosine and Hypoxanthine and Reduces Their Absorption in Rats. Microorganisms 2017, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Kilstrup, M.; Hammer, K.; Ruhdal Jensen, P.; Martinussen, J. Nucleotide Metabolism and Its Control in Lactic Acid Bacteria. FEMS Microbiol. Rev. 2005, 29, 555–590. [Google Scholar] [CrossRef]
- Bolotin, A.; Wincker, P.; Mauger, S.; Jaillon, O.; Malarme, K.; Weissenbach, J.; Ehrlich, S.D.; Sorokin, A. The Complete Genome Sequence of the Lactic Acid Bacterium Lactococcus Lactis Ssp. Lactis IL1403. Genome Res. 2001, 11, 731–753. [Google Scholar] [CrossRef]
- James, A.; Ke, H.; Yao, T.; Wang, Y. The Role of Probiotics in Purine Metabolism, Hyperuricemia and Gout: Mechanisms and Interventions. Food Rev. Int. 2021. [Google Scholar] [CrossRef]
- Bobulescu, I.A.; Moe, O.W. Renal Transport of Uric Acid: Evolving Concepts and Uncertainties. Adv. Chronic Kidney Dis. 2012, 19, 358–371. [Google Scholar] [CrossRef] [Green Version]
- Sorensen, L.B. Role of the Intestinal Tract in the Elimination of Uric Acid. Arthritis Rheum. 1965, 8, 694–706. [Google Scholar] [CrossRef]
- Abramson, R.; Levitt, M. Use of Pyrazinamide to Assess Renal Uric Acid Transport in the Rat: A Micropuncture Study. Am. J. Physiol.-Leg. Content 1976, 230, 1276–1283. [Google Scholar] [CrossRef]
- Giordano, C.; Karasik, O.; King-Morris, K.; Asmar, A. Uric Acid as a Marker of Kidney Disease: Review of the Current Literature. Dis. Mark. 2015, 2015, e382918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Zhu, C. Hyperuricemia Is a Adverse Prognostic Factor for Colon Cancer Patients. Int. J. Gen. Med. 2021, 14, 3001–3006. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Li, H.; Liu, Z.; Li, C.; Chen, Y.; Jiang, C.; Yu, Y.; Tian, Z. Impaired Intestinal Barrier Function in a Mouse Model of Hyperuricemia. Mol. Med. Rep. 2019, 20, 3292–3300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, H.; Tsunoda, T.; Ooyama, K.; Sakiyama, M.; Sogo, T.; Takada, T.; Nakashima, A.; Nakayama, A.; Kawaguchi, M.; Higashino, T.; et al. Hyperuricemia in Acute Gastroenteritis Is Caused by Decreased Urate Excretion via ABCG2. Sci. Rep. 2016, 6, 31003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, Z.; Liu, S.; Zhou, Y.; Mi, S.; Liu, G.; Wu, X.; Yao, K.; Assaad, H.; Deng, Z.; Hou, Y.; et al. Chlorogenic Acid Decreases Intestinal Permeability and Increases Expression of Intestinal Tight Junction Proteins in Weaned Rats Challenged with LPS. PLoS ONE 2014, 9, e97815. [Google Scholar] [CrossRef]
- Song, C.; Zhao, X. Uric Acid Promotes Oxidative Stress and Enhances Vascular Endothelial Cell Apoptosis in Rats with Middle Cerebral Artery Occlusion. Biosci. Rep. 2018, 38, BSR20170939. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Lv, Q.; Ren, H.; Gao, L.; Zhao, P.; Yang, X.; Yang, G.; Xu, D.; Wang, G.; Yang, W.; et al. The Altered Gut Microbiota of High-Purine-Induced Hyperuricemia Rats and Its Correlation with Hyperuricemia. PeerJ 2020, 8, e8664. [Google Scholar] [CrossRef]
- Hyperuricemia Is Associated with Impaired Intestinal Permeability in Mice. Am. J. Physiol.-Gastrointest. Liver Physiol. 2019, 317, G484–G492. [CrossRef]
- Slamet, A.R.; Subarna, T.; Nurawan, A.; Samsisaputra, W.; Salmin, M. Assembly of Location Specific Technology of Cacao in West Java (Indonesia); BPTP: Lembang, Indonesia, 1998. [Google Scholar]
- Jeung, W.H.; Shim, J.-J.; Woo, S.-W.; Sim, J.-H.; Lee, J.-L. Lactobacillus Curvatus HY7601 and Lactobacillus Plantarum KY1032 Cell Extracts Inhibit Adipogenesis in 3T3-L1 and HepG2 Cells. J. Med. Food 2018, 21, 876–886. [Google Scholar] [CrossRef]
- Cheng, J.; Park, S.B.; Kim, S.H.; Yang, S.H.; Suh, J.-W.; Lee, C.H.; Kim, J.G. Suppressing Activity of Staurosporine from Streptomyces Sp. MJM4426 against Rice Bacterial Blight Disease. J. Appl. Microbiol. 2016, 120, 975–985. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.Y.; Yoon, S.J.; Han, D.H.; Gupta, H.; Youn, G.S.; Shin, M.J.; Ham, Y.L.; Kwak, M.J.; Kim, B.Y.; Yu, J.S.; et al. Lactobacillus and Pediococcus Ameliorate Progression of Non-Alcoholic Fatty Liver Disease through Modulation of the Gut Microbiome. Gut Microbes 2020, 11, 882–899. [Google Scholar] [CrossRef] [PubMed]
- Ohno, I. Relationship Between Hyperuricemia and Chronic Kidney Disease. Nucleosides Nucleotides Nucleic Acids 2011, 30, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Hyperuricemia Causes Glomerular Hypertrophy in the Rat—Abstract. Am. J. Nephrol. 2003, 23, 2–7. [CrossRef] [PubMed]
- Wen, S.; Wang, D.; Yu, H.; Liu, M.; Chen, Q.; Bao, R.; Liu, L.; Zhang, Y.; Wang, T. The Time-Feature of Uric Acid Excretion in Hyperuricemia Mice Induced by Potassium Oxonate and Adenine. Int. J. Mol. Sci. 2020, 21, 5178. [Google Scholar] [CrossRef]
- Nigam, S.K.; Wu, W.; Bush, K.T.; Hoenig, M.P.; Blantz, R.C.; Bhatnagar, V. Handling of Drugs, Metabolites, and Uremic Toxins by Kidney Proximal Tubule Drug Transporters. Clin. J. Am. Soc. Nephrol. 2015, 10, 2039–2049. [Google Scholar] [CrossRef] [Green Version]
- Habu, Y.; Yano, I.; Takeuchi, A.; Saito, H.; Okuda, M.; Fukatsu, A.; Inui, K. Decreased Activity of Basolateral Organic Ion Transports in Hyperuricemic Rat Kidney: Roles of Organic Ion Transporters, ROAT1, ROAT3 and ROCT2. Biochem. Pharmacol. 2003, 66, 1107–1114. [Google Scholar] [CrossRef]
- Li, M.; Yang, D.; Mei, L.; Yuan, L.; Xie, A.; Yuan, J. Screening and Characterization of Purine Nucleoside Degrading Lactic Acid Bacteria Isolated from Chinese Sauerkraut and Evaluation of the Serum Uric Acid Lowering Effect in Hyperuricemic Rats. PLoS ONE 2014, 9, e105577. [Google Scholar] [CrossRef]
- Kano, H.; Yamada, N.; Saito, C.; Murayama-Chiba, Y.; Asami, Y.; Ito, H. Lactobacillus Gasseri PA-3, but Not L. Gasseri OLL2996, Reduces the Absorption of Purine Nucleosides in Rats. Nucleosides Nucleotides Nucleic Acids 2018, 37, 353–360. [Google Scholar] [CrossRef]
- Yamada, N.; Iwamoto, C.; Kano, H.; Yamaoka, N.; Fukuuchi, T.; Kaneko, K.; Asami, Y. Evaluation of Purine Utilization by Lactobacillus Gasseri Strains with Potential to Decrease the Absorption of Food-Derived Purines in the Human Intestine. Nucleosides Nucleotides Nucleic Acids 2016, 35, 670–676. [Google Scholar] [CrossRef]
- Martinussen, J.; Wadskov-Hansen, S.L.L.; Hammer, K. Two Nucleoside Uptake Systems in Lactococcus Lactis: Competition between Purine Nucleosides and Cytidine Allows for Modulation of Intracellular Nucleotide Pools. J. Bacteriol. 2003, 185, 1503–1508. [Google Scholar] [CrossRef] [Green Version]
- Fox, I.H.; Wyngaarden, J.B.; Kelley, W.N. Depletion of Erythrocyte Phosphoribosylpyrophosphate in Man. N. Engl. J. Med. 1970, 283, 1177–1182. [Google Scholar] [CrossRef] [PubMed]
- Martinussen, J.; Sørensen, C.; Jendresen, C.B.; Kilstrup, M. Two Nucleoside Transporters in Lactococcus Lactis with Different Substrate Specificities. Microbiology 2010, 156, 3148–3157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitetta, L.; Coulson, S.; Thomsen, M.; Nguyen, T.; Hall, S. Probiotics, D–Lactic Acidosis, Oxidative Stress and Strain Specificity. Gut Microbes 2017, 8, 311–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, P.; Fernández de Palencia, P.; Romano, A.; Fernández, M.; Lucas, P.; Spano, G.; López, P. Biogenic Amine Production by the Wine Lactobacillus Brevis IOEB 9809 in Systems That Partially Mimic the Gastrointestinal Tract Stress. BMC Microbiol. 2012, 12, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, N.; Gu, M.J.; Kye, Y.-C.; Ju, Y.-J.; Hong, R.; Ju, D.B.; Pyung, Y.J.; Han, S.H.; Park, B.-C.; Yun, C.-H. Bacteriophage EK99P-1 Alleviates Enterotoxigenic Escherichia Coli K99-Induced Barrier Dysfunction and Inflammation. Sci. Rep. 2022, 12, 941. [Google Scholar] [CrossRef] [PubMed]
- Ben-Mahdi, M.H.; Dang, P.M.-C.; Gougerot-Pocidalo, M.-A.; O’Dowd, Y.; El-Benna, J.; Pasquier, C. Xanthine Oxidase-Derived ROS Display a Biphasic Effect on Endothelial Cells Adhesion and FAK Phosphorylation. Oxidative Med. Cell. Longev. 2016, 2016, 9346242. [Google Scholar] [CrossRef] [Green Version]
- Ni, C.; Li, X.; Wang, L.; Li, X.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Lactic Acid Bacteria Strains Relieve Hyperuricaemia by Suppressing Xanthine Oxidase Activity via a Short-Chain Fatty Acid-Dependent Mechanism. Food Funct. 2021, 12, 7054–7067. [Google Scholar] [CrossRef]
- Pereira, F.C.; Wasmund, K.; Cobankovic, I.; Jehmlich, N.; Herbold, C.W.; Lee, K.S.; Sziranyi, B.; Vesely, C.; Decker, T.; Stocker, R.; et al. Rational Design of a Microbial Consortium of Mucosal Sugar Utilizers Reduces Clostridiodes Difficile Colonization. Nat. Commun. 2020, 11, 5104. [Google Scholar] [CrossRef]
- Smith, B.J.; Miller, R.A.; Ericsson, A.C.; Harrison, D.C.; Strong, R.; Schmidt, T.M. Changes in the Gut Microbiome and Fermentation Products Concurrent with Enhanced Longevity in Acarbose-Treated Mice. BMC Microbiol. 2019, 19, 130. [Google Scholar] [CrossRef] [Green Version]
- Biddle, A.; Stewart, L.; Blanchard, J.; Leschine, S. Untangling the Genetic Basis of Fibrolytic Specialization by Lachnospiraceae and Ruminococcaceae in Diverse Gut Communities. Diversity 2013, 5, 627–640. [Google Scholar] [CrossRef]
- Geerlings, S.Y.; Kostopoulos, I.; de Vos, W.M.; Belzer, C. Akkermansia muciniphila in the Human Gastrointestinal Tract: When, Where, and How? Microorganisms 2018, 6, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derrien, M.; Belzer, C.; de Vos, W.M. Akkermansia Muciniphila and Its Role in Regulating Host Functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, C.M.A.; Connors, J.; Dunn, K.A.; Bielawski, J.P.; Comeau, A.M.; Langille, M.G.I.; Van Limbergen, J. Bacterial Taxa and Functions Are Predictive of Sustained Remission Following Exclusive Enteral Nutrition in Pediatric Crohn’s Disease. Inflamm. Bowel Dis. 2020, 26, 1026–1037. [Google Scholar] [CrossRef]
- Seregin, S.S.; Golovchenko, N.; Schaf, B.; Chen, J.; Pudlo, N.A.; Mitchell, J.; Baxter, N.T.; Zhao, L.; Schloss, P.D.; Martens, E.C.; et al. NLRP6 Protects Il10−/− Mice from Colitis by Limiting Colonization of Akkermansia Muciniphila. Cell Rep. 2017, 19, 733–745. [Google Scholar] [CrossRef]
- Zhao, G.-J.; Li, D.; Zhao, Q.; Lian, J.; Hu, T.-T.; Hong, G.-L.; Yao, Y.-M.; Lu, Z.-Q. Prognostic Value of Plasma Tight-Junction Proteins for Sepsis in Emergency Department: An Observational Study. Shock 2016, 45, 326–332. [Google Scholar] [CrossRef] [Green Version]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.-J.M.; Wells, J.M. Regulation of Human Epithelial Tight Junction Proteins by Lactobacillus Plantarum in Vivo and Protective Effects on the Epithelial Barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boffetta, P.; Nordenvall, C.; Nyrén, O.; Ye, W. A Prospective Study of Gout and Cancer. Eur. J. Cancer Prev. 2009, 18, 127–132. [Google Scholar] [CrossRef]
- Bush, K.T.; Singh, P.; Nigam, S.K. Gut-Derived Uremic Toxin Handling in Vivo Requires OAT-Mediated Tubular Secretion in Chronic Kidney Disease. JCI Insight 2020, 5, e133817. [Google Scholar] [CrossRef] [Green Version]
- Fang, C.; Chen, L.; He, M.; Luo, Y.; Zhou, M.; Zhang, N.; Yuan, J.; Wang, H.; Xie, Y. Molecular Mechanistic Insight into the Anti-Hyperuricemic Effect of Eucommia Ulmoides in Mice and Rats. Pharm. Biol. 2019, 57, 112–119. [Google Scholar] [CrossRef] [Green Version]
- Buschmann, C.T.; Tsokos, M. Deaths: Pregnancy-related deaths—Pathology. In Encyclopedia of Forensic and Legal Medicine, 2nd ed.; Payne-James, J., Byard, R.W., Eds.; Elsevier: Oxford, UK, 2016; pp. 123–127. ISBN 978-0-12-800055-7. [Google Scholar]
- Hernández-Gómez, J.G.; López-Bonilla, A.; Trejo-Tapia, G.; Ávila-Reyes, S.V.; Jiménez-Aparicio, A.R.; Hernández-Sánchez, H. In Vitro Bile Salt Hydrolase (BSH) Activity Screening of Different Probiotic Microorganisms. Foods 2021, 10, 674. [Google Scholar] [CrossRef]
- Palaniyandi, S.A.; Damodharan, K.; Suh, J.-W.; Yang, S.H. In Vitro Characterization of Lactobacillus Plantarum Strains with Inhibitory Activity on Enteropathogens for Use as Potential Animal Probiotics. Indian J. Microbiol. 2017, 57, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and Broth Dilution Methods to Determine the Minimal Inhibitory Concentration (MIC) of Antimicrobial Substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Guidance on the Assessment of Bacterial Susceptibility to Antimicrobials of Human and Veterinary Importance. EFSA J. 2012, 10, 2740. [CrossRef]
- Bove, P.; Gallone, A.; Russo, P.; Capozzi, V.; Albenzio, M.; Spano, G.; Fiocco, D. Probiotic Features of Lactobacillus Plantarum Mutant Strains. Appl. Microbiol. Biotechnol. 2012, 96, 431–441. [Google Scholar] [CrossRef]
- Damodharan, K.; Lee, Y.S.; Palaniyandi, S.; Yang, S.H.; Suh, J.-W. Preliminary Probiotic and Technological Characterization of Pediococcus Pentosaceus Strain KID7 and in Vivo Assessment of Its Cholesterol-Lowering Activity. Front. Microbiol. 2015, 6, 768. [Google Scholar] [CrossRef] [Green Version]
- Soncin, F.; Shapiro, R.; Fett, J.W. A Cell-Surface Proteoglycan Mediates Human Adenocarcinoma HT-29 Cell Adhesion to Human Angiogenin. J. Biol. Chem. 1994, 269, 8999–9005. [Google Scholar] [CrossRef]
- Tagg, J.R.; McGiven, A.R. Assay System for Bacteriocins. Appl. Microbiol. 1971, 21, 943. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′-3′) |
|---|---|---|
| OAT1 | Forward | GAGCAGAGGAAAGCAGAAGC |
| Reverse | CCCTTTAGTGCTGTGTGACG | |
| OAT3 | Forward | TACAGTTGTCCGTGTCTGCT |
| Reverse | CTTCCTCCTTCTTGCCGTTG | |
| URAT1 | Forward | AGGTCCTGACAGGTTCTGT |
| Reverse | CTCTGCCTTCCTCCTGTTGA | |
| GLUT9 | Forward | TTCGGGTCCTCCTTCCTCTA |
| Reverse | GGACACAGTCACAGACCAGA | |
| GAPDH | Forward | GGCACAGTCAAGGCTGGAATG |
| Reverse | ATGGTGGTGAAGACGCCAGTA |
| Strain No. | Strain Name * | (μmol/min) | (%) | (μmol/min) | (%) |
|---|---|---|---|---|---|
| MJM60349 | Latilactobacillus curvatus | 1.7 | 16.6 | 1.5 | 14.3 |
| MJM60355 | Latilactobacillus curvatus | 1.7 | 16.5 | 1.6 | 14.7 |
| MJM60363 | Leuconostoc mesenteroides | 4.0 | 38.2 | 3.3 | 31.3 |
| MJM60364 | Latilactobacillus sakei | 4.3 | 41.3 | 5.1 | 48.8 |
| MJM60366 | Latilactobacillus curvatus | 10.3 | 100.0 | 2.9 | 27.9 |
| MJM60367 | Leuconostoc mesenteroides | 9.6 | 92.6 | 2.7 | 25.1 |
| MJM60368 | Latilactobacillus sakei | 3.4 | 33.1 | 3.4 | 32.5 |
| MJM60370 | Leuconostoc mesenteroides | 2.7 | 25.5 | 2.3 | 22.2 |
| MJM60371 | Leuconostoc mesenteroides | 4.8 | 46.1 | 4.0 | 38.0 |
| MJM60375 | Latilactobacillus sakei | 4.0 | 39.0 | 4.2 | 40.2 |
| MJM60376 | Leuconostoc mesenteroides | 1.9 | 18.4 | 1.9 | 18.1 |
| MJM60377 | Leuconostoc mesenteroides | 1.9 | 18.6 | 1.9 | 17.7 |
| MJM60378 | Leuconostoc mesenteroides | 6.5 | 62.4 | 5.3 | 50.4 |
| MJM60341 | Lacticaseibacillus paracasei | 7.9 | 73.3 | 4.7 | 44.3 |
| MJM60385 | Lacticaseibacillus rhamnosus | 2.8 | 30.0 | 2.1 | 23.0 |
| MJM60386 | Levilactobacillus brevis | 6.9 | 75.9 | 6.3 | 67.5 |
| MJM60389 | Lactobacillus plantarum | 7.5 | 82.1 | 6.8 | 73.0 |
| MJM60391 | Levilactobacillus brevis | 4.1 | 44.8 | 3.0 | 32.5 |
| MJM60392 | Lactococcus lactis | 1.9 | 21.0 | 2.0 | 21.1 |
| MJM60396 | Lacticaseibacillus paracasei | 10.3 | 100.0 | 10.5 | 100.0 |
| MJM60662 | Lactobacillus gasseri | 10.3 | 100.0 | 10.5 | 100.0 |
| LGG | Lacticaseibacillus rhamnosus GG | 8.5 | 81.16 | 8.2 | 78.53 |
| Degradation (%) | L. paracasei MJM60396 | L. gasseri MJM60662 | ||||
|---|---|---|---|---|---|---|
| Live | Pellet | Supernatant | Live | Pellet | Supernatant | |
| Adenosine | 100 | 100 | 29 | 100 | 45 | 44 |
| Guanosine | 100 | 100 | 48 | 100 | 72 | 46 |
| Inosine | 100 | 100 | 50 | 100 | 60 | 35 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.; Werlinger, P.; Suh, J.-W.; Cheng, J. Potential Probiotic Lacticaseibacillus paracasei MJM60396 Prevents Hyperuricemia in a Multiple Way by Absorbing Purine, Suppressing Xanthine Oxidase and Regulating Urate Excretion in Mice. Microorganisms 2022, 10, 851. https://doi.org/10.3390/microorganisms10050851
Lee Y, Werlinger P, Suh J-W, Cheng J. Potential Probiotic Lacticaseibacillus paracasei MJM60396 Prevents Hyperuricemia in a Multiple Way by Absorbing Purine, Suppressing Xanthine Oxidase and Regulating Urate Excretion in Mice. Microorganisms. 2022; 10(5):851. https://doi.org/10.3390/microorganisms10050851
Chicago/Turabian StyleLee, Youjin, Pia Werlinger, Joo-Won Suh, and Jinhua Cheng. 2022. "Potential Probiotic Lacticaseibacillus paracasei MJM60396 Prevents Hyperuricemia in a Multiple Way by Absorbing Purine, Suppressing Xanthine Oxidase and Regulating Urate Excretion in Mice" Microorganisms 10, no. 5: 851. https://doi.org/10.3390/microorganisms10050851
APA StyleLee, Y., Werlinger, P., Suh, J.-W., & Cheng, J. (2022). Potential Probiotic Lacticaseibacillus paracasei MJM60396 Prevents Hyperuricemia in a Multiple Way by Absorbing Purine, Suppressing Xanthine Oxidase and Regulating Urate Excretion in Mice. Microorganisms, 10(5), 851. https://doi.org/10.3390/microorganisms10050851