
Effect of Intrinsic Tannins on the Fermentation Quality and Associated with the Bacterial and Fungal Community of Sainfoin Silage



Abstract
:1. Introduction
2. Materials and Methods
2.1. Forage and Ensiling
2.2. Characteristics of Wilted and Ensiled Sainfoin
2.3. Sequencing Analysis of the Bacterial Community
2.4. Statistical Analysis
3. Results
3.1. Characteristics of Wilted and Ensiling of Sainfoin
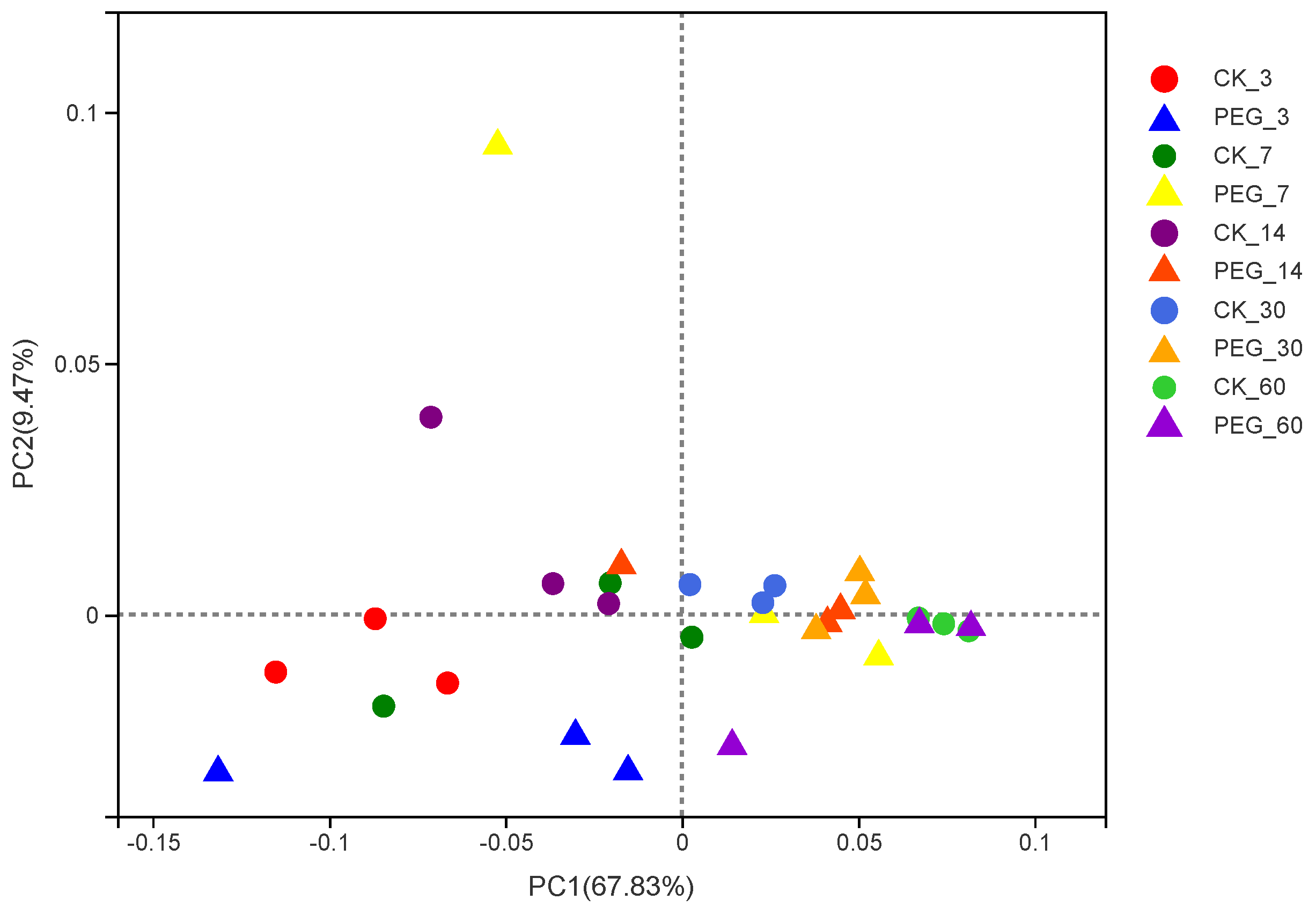
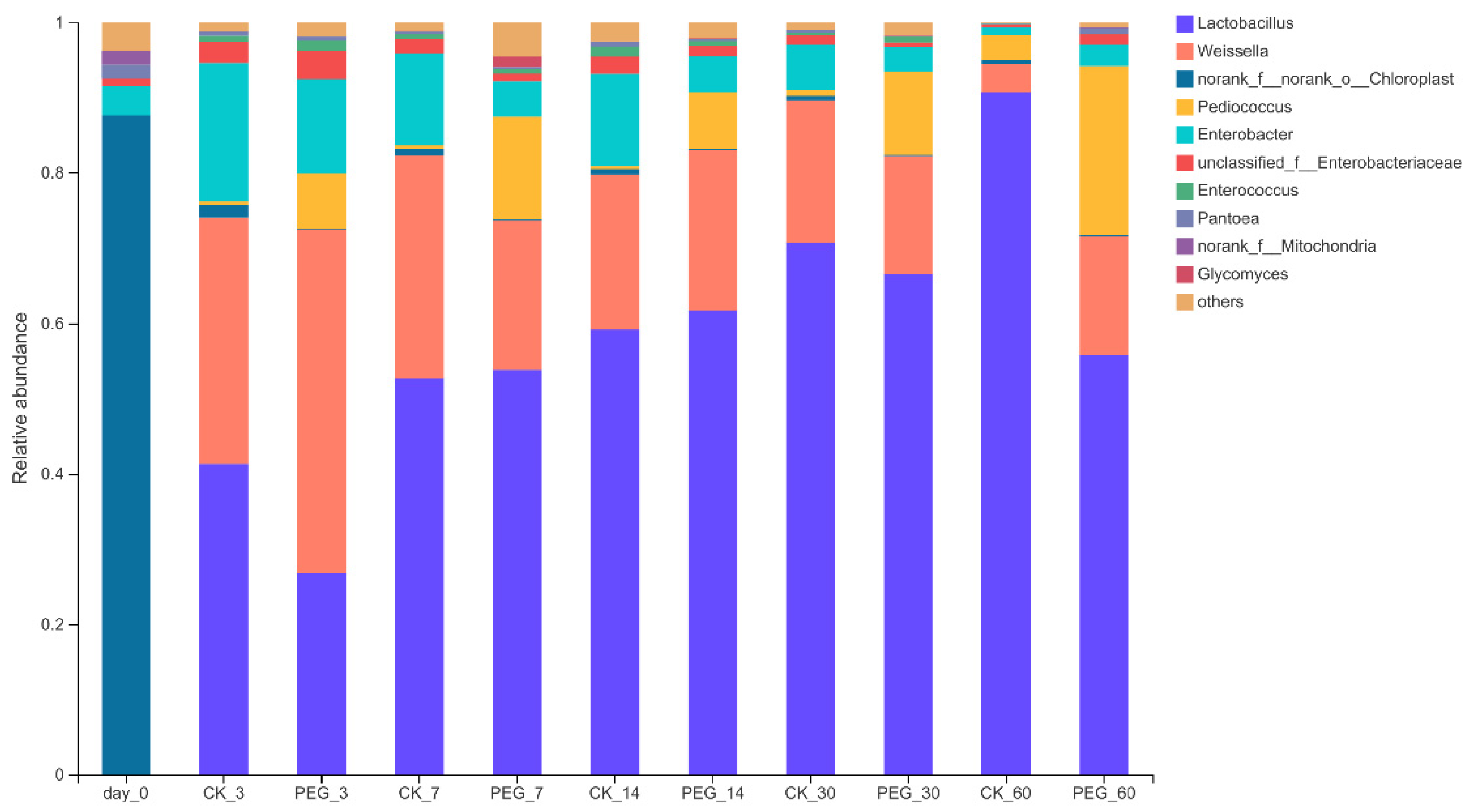
3.2. Bacterial Community of Sainfoin Silage
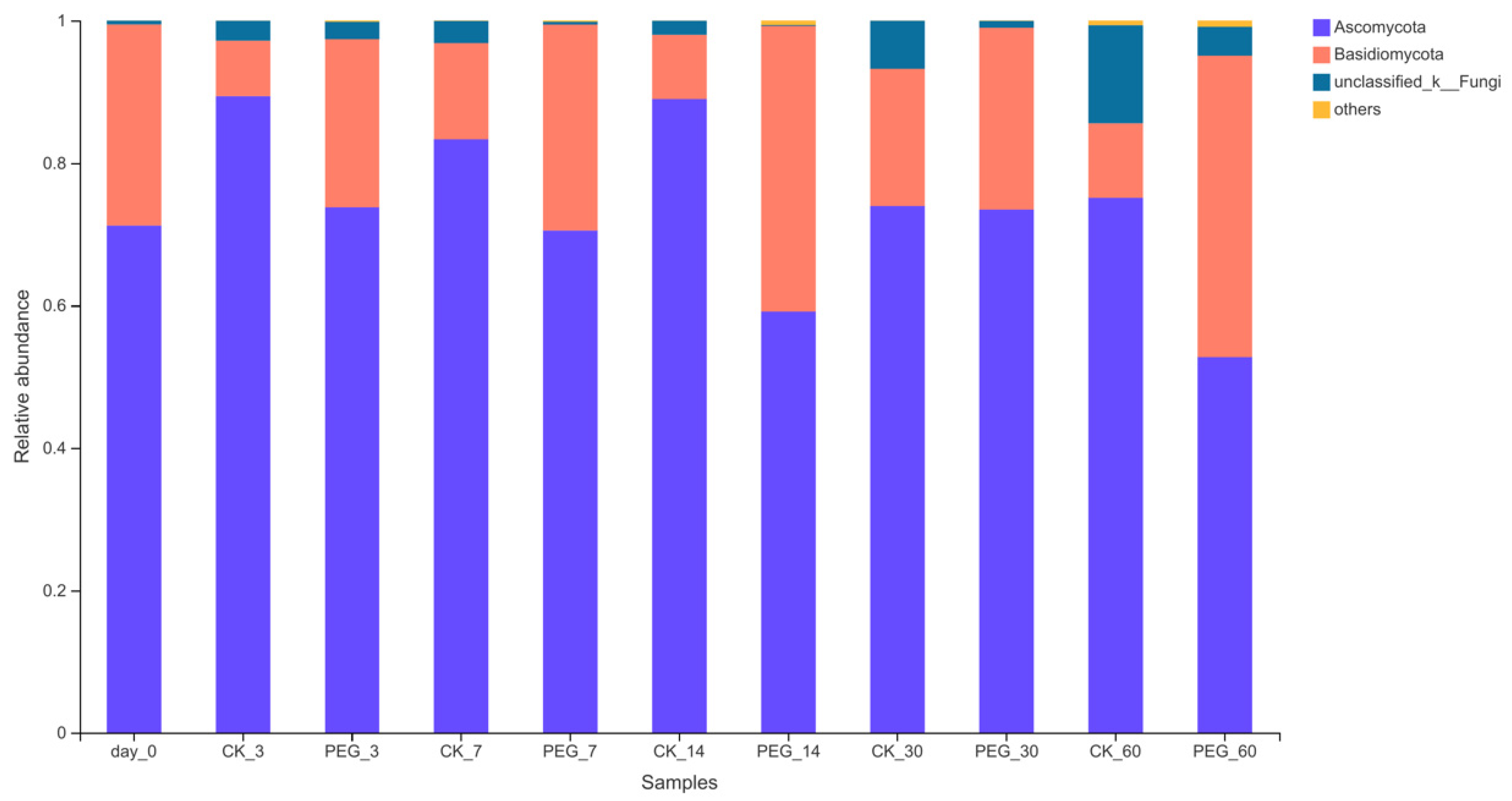
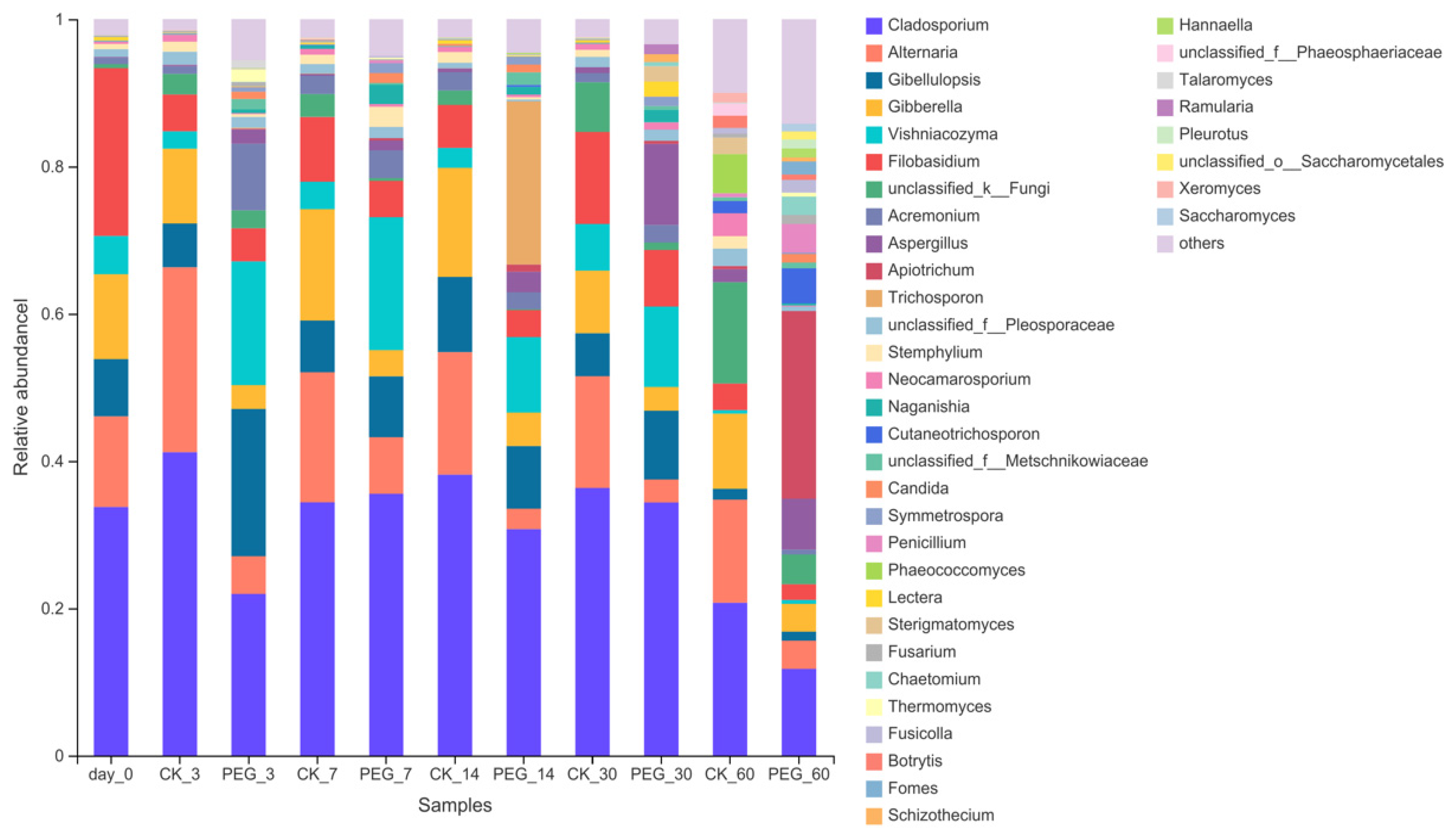
3.3. Fungi Community of Sainfoin Ensiled with or without PEG
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zeller, W.E. Activity, Purification, and Analysis of Condensed Tannins: Current State of Affairs and Future Endeavors. Crop Sci. 2019, 59, 886–904. [Google Scholar] [CrossRef]
- McAllister, T.A.; Martinez, T.; Bae, H.; Muir, A.; Yanke, L.; Jones, G. Characterization of Condensed Tannins Purified from Legume Forages: Chromophore Production, Protein Precipitation, and Inhibitory Effects on Cellulose Digestion. J. Chem. Ecol. 2005, 31, 2049–2068. [Google Scholar] [CrossRef]
- Xu, D.M.; Ding, Z.T.; Wang, M.S.; Bai, J.; Ke, W.C.; Zhang, Y.X.; Guo, X.S. Characterization of the Microbial Community, Metabolome and Biotransformation of Phenolic Compounds of Sainfoin (Onobrychis viciifolia) Silage Ensiled with or without Inoculation of Lactobacillus Plantarum. Bioresour. Technol. 2020, 316, 123910. [Google Scholar] [CrossRef] [PubMed]
- Azuhnwi, B.; Boller, B.; Martens, M.; Dohme-Meier, F.; Ampuero, S.; Gunter, S.; Kreuzer, M.; Hess, H.D. Morphology, Tannin Concentration and Forage Value of 15 Swiss Accessions of Sainfoin (Onobrychis viciifolia) as Influenced by Harvest Time and Cultivation Site. Grass Forage Sci. 2011, 66, 474–487. [Google Scholar] [CrossRef]
- Theodoridou, K.; Aufre`re, J.; Andueza, D.; Le Morvan, A.; Picard, F.; Stringano, E.; Pourrat, J.; Mueller-Harvey, I.; Baumont, R. Effect of Plant Development during First and Second Growth Cycle on Chemical Composition, Condensed Tannins and Nutritive Value of Three Sainfoin (Onobrychis viciifolia) Varieties and Lucerne. Grass Forage Sci. 2011, 66, 402–414. [Google Scholar] [CrossRef]
- Wang, Y.; McAllister, T.A.; Acharya, S. Condensed Tannins in Sainfoin: Composition, Concentration, and Effects on Nutritive and Feeding Value of Sainfoin Forage. Crop Sci. 2015, 55, 13–22. [Google Scholar] [CrossRef]
- Cavallarin, L.; Antoniazzi, S.; Borreani, G.; Tabacco, E. Effects of Wilting and Mechanical Conditioning on Proteolysis in Sainfoin (Onobrychis Viciffolia) Wilted Herbage and Silaget. J. Sci. Food Agric. 2005, 85, 831–838. [Google Scholar] [CrossRef]
- Brinkhaus, A.G.; Wyss, U.; Arrigo, Y.; Girard, M.; Bee, G.; Zeitza, J.O.; Kreuzer, M.; Dohme-Meier, F. In Vitro Ruminal Fermentation Characteristics and Utilisable CP Supply of Sainfoin and Birdsfoot Trefoil Silages and Their Mixtures with Other Legumes. Animal 2017, 11, 580–590. [Google Scholar] [CrossRef] [Green Version]
- Berard, N.C.; Holley, R.A.; McAllister, T.A.; Ominski, K.H.; Wittenberg, K.M.; Bouchard, K.S.; Bouchard, J.J.; Krause, D.O. Poten- Tial to Reduce Escherichia Coli Shedding in Cattle Feces by Using Sainfoin (Onobrychis viciifolia) Forage, Tested in Vitro and in Vivo. Appl. Environ. Microbiol. 2009, 75, 1074–1079. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.L.; Hao, Y.Q.; Jin, L.; Xu, Z.J.; McAllister, T.A. Anti-Escherichia Coli O157:H7 Properties of Purple Prairie Clover and Sainfoin Condensed Tannins. Molecules 2013, 18, 2183. [Google Scholar] [CrossRef]
- Jones, W.T.; Mangan, J.L. Complexes of the Condensed Tannins of Sainfoin (Onobrychis viciifolia) with Fraction 1 Leaf Protein and with Submaxillary Mucoprotein, and Their Reversal by Polyethylene Glycol and PH. J. Sci. Food Agric. 1977, 28, 126–136. [Google Scholar] [CrossRef]
- Peng, K.; Jin, L.; Niu, Y.D.; Huang, Q.; McAllister, T.A.; Yang, H.E.; Denise, H.; Xu, Z.; Acharya, S.; Wang, S.; et al. Condensed Tannins Affect Bacterial and Fungal Microbiomes and Mycotoxin Production during Ensiling and upon Aerobic Exposure. Appl. Environ. Microbiol. 2018, 84, e02274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, J.; Xu, D.; Wang, M.; Li, Z.; Guo, X. Effects of Antibacterial Peptide-Producing Bacillus Subtilis and Lactobacillus Buchneri on Fermentation, Aerobic Stability, and Microbial Community of Alfalfa Silage. Bioresour. Technol. 2020, 315, 123881. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Feng, Y.; Pei, J.; Li, J.; Wang, Z.; Fu, S.; Zheng, Y.; Li, Z.; Peng, Z. Effects of Lactobacillus Plantarum Additive and Temperature on the Ensiling T Quality and Microbial Community Dynamics of Cauliflower Leaf Silages. Bioresour. Technol. 2020, 307, 123238. [Google Scholar] [CrossRef] [PubMed]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Weatherburn, M.W. Phenol-Hypochlorite Reaction for Determination of Ammonia. Anal. Chem. 1967, 39, 971–974. [Google Scholar] [CrossRef]
- Terrill, T.H.; Rowan, A.M.; Douglas, G.B.; Barry, T.N. Determination of Extractable and Bound Condensed Tannin Concentrations in Forage Plants, Protein Concentrate Meals and Cereal Grains. J. Sci. Food Agric. 1992, 58, 321–329. [Google Scholar] [CrossRef]
- He, L.; Lv, H.; Xing, Y.; Chen, X.; Zhang, Q. Intrinsic Tannins Affect Ensiling Characteristics and Proteolysis of Neolamarckia Cadamba Leaf Silage by Largely Altering Bacterial Community. Bioresour. Technol. 2020, 311, 123496. [Google Scholar] [CrossRef]
- Ni, K.K.; Wang, F.F.; Zhu, B.G.; Yang, J.X.; Zhou, G.A.; Pan, Y.; Tao, Y.; Zhong, J. Effects of Lactic Acid Bacteria and Molasses Additives on the Microbial Community and Fermentation Quality of Soybean Silage. Bioresour. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef]
- Adams, R.I.; Miletto, M.; Taylor, J.W.; Bruns, T.D. Dispersal in Microbes: Fungi in Indoor Air Are Dominated by Outdoor Air and Show Dispersal Limitation at Short Distances. ISME J. 2013, 7, 1460. [Google Scholar] [CrossRef] [Green Version]
- Su, R.N.; Ni, K.K.; Wang, T.W.; Yang, X.P.; Zhang, J.; Liu, Y.Y.; Shi, W.X.; Yan, L.; Jie, C.; Zhong, J. Effects of Ferulic Acid Esterase-Producing Lactobacillus Fermentum and Cellulase Additives on the Fermentation Quality and Microbial Community of Alfalfa Silage. PeerJ 2019, 7, e7712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Pian, R.Q.; Chen, X.Y.; Lv, H.J.; Zhou, W.; Zhang, Q. Beneficial Effects of Tannic Acid on the Quality of Bacterial Communities Present in High-Moisture Mulberry Leaf and Stylo Silage. Front. Microbiol. 2020, 11, 586412. [Google Scholar] [CrossRef] [PubMed]
- Rufino-Moya, P.J.; Blanco, M.; Bertolin, J.R. Joy Effect of the Method of Preservation on the Chemical Composition T and in Vitro Fermentation Characteristics in Two Legumes Rich in Condensed Tannins. Anim. Feed Sci. Technol. 2019, 251, 12–20. [Google Scholar] [CrossRef]
- Ali, N.; Wang, S.R.; Zhao, J.; Dong, Z.H.; Li, J.F.; Nazar, M.; Shao, T. Microbial Diversity and Fermentation Profile of Red Clover Silage Inoculated with Reconstituted Indigenous and Exogenous Epiphytic Microbiota. Bioresour. Technol. 2020, 314, 123606. [Google Scholar] [CrossRef]
- McDonald, P.; Henderson, A.R.; Herson, S. The Biochemistry of Silage; Chalcombe Publications: Kent, UK, 1991; pp. 256–271. [Google Scholar]
- Chen, L.Y.; Qu, H.; Bai, S.Q.; Yan, L.J.; You, M.H.; Gou, W.L.; Li, P.; Gao, F. Effect of Wet Sea Buckthorn Pomace Utilized as an Additive on Silage Fermentation Profile and Bacterial Community Composition of Alfalfa. Bioresour. Technol. 2020, 314, 123773. [Google Scholar] [CrossRef]
- Renzetti, A.; Betts, J.W.; Fukumoto, K.; Rutherford, R.N. Antibacterial Green Tea Catechins from a Molecular Perspective: Mechanisms of Action and Structure-Activity Relationships. Food Funct. 2020, 11, 9370–9396. [Google Scholar] [CrossRef]
- Jones, G.A.; McAllister, T.A.; Muir, A.D.; Cheng, K.J. Effects of Sainfoin (Onobrychis viciifolia) Condensed Tannins on Growth and Proteolysis by Four Strains of Ruminal Bacteria. Appl. Environ. Microbiol. 1994, 60, 1374–1378. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, M.; Tomiyama, D.; Shigemune, N.; Mitani, A.; Xu, W.; Miyamoto, T. Cell Surface Hydrophobicity Contributes to Lactobacillus Tolerance to Antibacterial Actions of Catechins. Food Sci. Technol. Res. 2015, 21, 583–588. [Google Scholar] [CrossRef] [Green Version]
- Muck, R.; Nadeau, E.M.G.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L., Jr. Silage Review: Recent Advances and Future Uses of Silage Additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef]
- Cai, Y.; Kumai, S.; Ogawa, M.; Benno, Y.; Nakase, T. Characterization and Identification of Pediococcus Species Isolated from Forage Crops and Their Application for Silage Preparation. Appl. Environ. Microbiol. 1999, 65, 2901–2906. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.S.; Chen, M.Y.; Bai, J.; Zhang, J.Y.; Su, R.N.; Franco, M.; Ding, Z.T.; Zhang, X.; Zhang, Y.; Guo, X.S. Ensiling Characteristics, in Vitro Rumen Fermentation Profile, Methane Emission and Archaeal and Protozoal Community of Silage Prepared with Alfalfa, Sainfoin and Their Mixture. Anim. Feed Sci. Technol. 2021, 284, 115154. [Google Scholar] [CrossRef]
- Zhou, Y.; Drouin, P.; Lafreniere, C. Effect of Temperature (5–25 °C) on Epiphytic Lactic Acid Bacteria Populations and Fermentation of Whole-Plant Corn Silage. J. Appl. Microbiol. 2016, 121, 657–671. [Google Scholar] [CrossRef] [PubMed]
- Pahlow, G.; Muck, R.E.; Driehuis, F.; Elferink, S.J.W.H.; Spoelstra, S.F. Microbiology of Ensiling. In Silage Science and Technology Agronomy; Buxton, D.R., Muck, R., Harrison, J.H., Eds.; Asa, Cssa, Ssa: Madison, WI, USA, 2003; pp. 31–93. [Google Scholar]
- Muck, R.E. Silage Microbiology and Its Control through Additives. Rev. Bras. Zootec. 2010, 39, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Scudamore, K.A.; Livesey, C.T. Occurrence and Significance of Mycotoxins in Forage Crops and Silage: A Review. J. Sci. Food Agric. 1998, 77, 1–17. [Google Scholar] [CrossRef]
- Wali, A.; Nishino, N. Bacterial and Fungal Microbiota Associated with the Ensiling of Wet Soybean Curd Residue under Prompt and Delayed Sealing Conditions. Microorganisms 2020, 8, 1334. [Google Scholar] [CrossRef]
- Cogan, T.; Hawkey, R.; Higgie, E.; Lee, M.R.F.; Mee, E.; Parfitt, D.; Raj, J.; Roderick, S.; Walker, N.; Ward, P.; et al. Silage and Total Mixed Ration Hygienic Quality on Commercial Farms: Implications for Animal Production. Grass Forage Sci. 2017, 72, 601–613. [Google Scholar] [CrossRef] [Green Version]
- Tangni, E.K.; Wambacq, E.; Bastiaanse, H.; Haesaert, G.; Pussemier, L.; De Poorter, J.; Foucart, G.; Van Hove, F. Survey of Fungal Diversity in Silages Supplied to Dairy Cattle in Belgium over a Two-Year Period. J. Anim. Sci. Adv. 2017, 7, 1861–1873. [Google Scholar]
- Vandicke, J.; De Visschere, K.; Ameye, M.; Croubels, S.; De Saeger, S.; Audenaert, K.; Haesaert, G. Multi-Mycotoxin Contamination of Maize Silages in Flanders, Belgium: Monitoring Mycotoxin Levels from Seed to Feed. Toxins 2021, 13, 202. [Google Scholar] [CrossRef]
- Cobo-Diaz, J.F.; Legrand, F.; Le Floc, G.; Picot, A. Influence of Maize Residues in Shaping Soil Microbiota and Fusarium Spp. Communities. Microb. Ecol. 2021, 83, 702–713. [Google Scholar] [CrossRef]
- Mansfield, M.A.; Kuldau, G.A. Microbiological and Molecular Determination of Mycobiota in Fresh and Ensiled Maize Silage. Mycologia 2007, 99, 269–278. [Google Scholar] [CrossRef]
- Alonso, V.A.; Pereyra, C.M.; Keller, L.A.M.; Dalcero, A.M.; Rosa, C.A.R.; Chiacchiera, S.M.; Cavaglieri, L.R. Fungi and Mycotoxins in Silage: An Overview. J. Appl. Microbiol. 2013, 115, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Cheli, F.; Campagnoli, A.; Dell’Orto, V. Fungal Populations and Mycotoxins in Silages: From Occurrence to Analysis. Anim. Feed Sci. Technol. 2013, 183, 1–16. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Composition | Mean ± SE |
|---|---|
| Dry matter (g/kg fresh forage) | 236.5 ± 2.5 |
| Crude protein | 223.12 ± 0.61 |
| Water-soluble carbohydrates | 101.43 ± 0.89 |
| Neutral detergent fiber | 505.83 ± 5.03 |
| Acid detergent fiber | 300.66 ± 10.58 |
| Neutral detergent—insoluble protein | 129.16 ± 1.11 |
| Acid detergent—insoluble protein | 30.23 ± 0.50 |
| pH | 5.41 ± 0.01 |
| Soluble protein | 10.75 ± 0.24 |
| Extractable condensed tannins (g/kg DM) | 47.31 ± 2.46 |
| Protein-bound condensed tannins (g/kg DM) | 45.67 ± 1.91 |
| Fiber-bound condensed tannins (g/kg DM) | 16.80 ± 0.34 |
| Total condensed tannins | 109.78 ± 3.52 |
| Lactic-acid bacteria (Log10 cfu/g FM) | 6.75 ± 0.06 |
| Yeasts (Log10 cfu/g FM) | ND |
| Molds (Log10 cfu/g FM) | 5.70 ± 0.04 |
| Aerobic bacteria (Log10 cfu/g FM) | 7.11 ± 0.03 |
| Item | Treatment | Days of Ensiling | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 3 | 7 | 14 | 30 | 60 | Day | PEG | D*P | |||
| DM | CK | 22.13Aa | 21.60Aa | 21.83Aa | 21.48Aa | 21.68Aa | 0.139 | <0.01 | <0.01 | 0.173 |
| PEG | 24.16Ba | 23.28Ba | 23.73Ba | 23.46Ba | 22.68Bb | |||||
| DML (%DM) | CK | 6.43Aa | 8.67Aa | 7.72Aa | 9.19Aa | 8.32Ba | 0.403 | <0.01 | <0.01 | 0.299 |
| PEG | 8.57Ab | 11.92Aa | 10.21Aa | 11.19Aa | 14.15Aa | |||||
| CP (g/kg DM) | CK | 240.13Aa | 239.64Aa | 240.85Aa | 244.70Aa | 246.79Aa | 2.72 | <0.01 | <0.01 | <0.01 |
| PEG | 219.60Ba | 202.90Bb | 204.52Bb | 216.02Bac | 208.02Bbc | |||||
| NDF (g/kg DM) | CK | 493.37Aa | 473.01Aa | 426.64Aa | 448.55Aa | 455.10Aa | 8.54 | <0.01 | <0.01 | 0.540 |
| PEG | 420.10Aa | 351.96Bb | 348.77Ab | 350.79Bb | 348.58Bb | |||||
| ADF (g/kg DM) | CK | 299.39Ab | 271.07Ab | 322.29Aa | 300.73Ab | 274.06Ab | 4.90 | 0.105 | <0.01 | 0.011 |
| PEG | 280.77Aa | 265.25Aa | 239.75Ba | 248.98Aa | 245.87Aa | |||||
| E-CT (g/kg DM) | CK | 19.63 | 20.93 | 19.38 | 18.67 | 16.83 | 0.721 | 0.484 | - | - |
| P-CT (g/kg DM) | CK | 69.69 | 64.82 | 69.73 | 72.57 | 69.92 | 1.03 | 0.280 | - | - |
| F-CT (g/kg DM) | CK | 27.40 | 25.61 | 27.14 | 26.12 | 25.48 | 0.407 | 0.441 | - | - |
| pH | CK | 4.61Ab | 4.75Aa | 4.67Aab | 4.57Ab | 4.32Ac | 0.025 | <0.01 | 0.53 | 0.064 |
| PEG | 4.56Ab | 4.68Aa | 4.69Aab | 4.60Aa | 4.36Ac | |||||
| AN (g/kg DM) | CK | 0.64Bc | 0.94Bb | 1.13Bb | 1.42Ba | 1.60Ba | 0.064 | <0.01 | <0.01 | <0.01 |
| PEG | 1.16Ad | 1.24Ad | 1.51Ac | 1.93Ab | 2.44Aa | |||||
| WSC (g/kg DM) | CK | 46.02Aab | 41.77Aab | 37.95Ab | 35.76Abc | 31.09Ac | 0.89 | <0.01 | <0.01 | <0.01 |
| PEG | 30.89Bab | 32.86Ba | 30.64Bab | 26.47Bbc | 23.34Bc | |||||
| LA (g/kg DM) | CK | 107.74Aa | 96.25Aa | 86.33Aa | 108.99Ba | 115.36Aa | 4.37 | 0.003 | <0.01 | 0.063 |
| PEG | 102.72Ab | 123.03Ab | 127.94Ab | 158.89Aa | 162.98Aa | |||||
| AA (g/kg DM) | CK | 67.57Ab | 76.66Aab | 66.20Ab | 73.15Bb | 101.62Aa | 3.56 | <0.01 | <0.01 | <0.01 |
| PEG | 63.33Ab | 80.20Ab | 68.61Ab | 138.47Aa | 117.85Aa | |||||
| PA (g/kg DM) | CK | 4.72Aa | 1.47Ab | 0.71Ab | 1.41Ab | 1.82Bb | 0.293 | <0.01 | <0.01 | 0.135 |
| PEG | 6.10Aa | 1.89Ab | 2.28Ab | 2.75Ab | 5.50Aa | |||||
| LA to AA Ratio | CK | 1.60Aa | 1.26 Aa | 1.35 Aa | 1.50 Aa | 1.13 Aa | 0.044 | 0.014 | 0.035 | <0.01 |
| PEG | 1.61 Aa | 1.56 Aa | 1.86 Aa | 1.15 Aa | 1.45 Aa | |||||
| LAB Log10 cfu/g FM | CK | 9.04Aa | 8.82Bab | 8.70Aab | 8.06Bc | 7.13Bd | 0.089 | <0.01 | <0.01 | 0.068 |
| PEG | 9.19Aa | 9.28Aa | 8.99Aa | 8.60Ab | 7.72Ac | |||||
| Molds Log10 cfu/g FM | CK | 5.06Ba | 4.92Aa | 4.82Bab | 4.78Aab | 4.44Bb | 0.042 | <0.01 | <0.01 | 0.300 |
| PEG | 5.50Aa | 5.21Aab | 5.20Aab | 5.14Aab | 5.08Ab | |||||
| Yeasts Log10 cfu/g FM | CK | 3.94Aa | 3.82Ba | 3.87Aa | 2.85Ab | 2.79Ab | 0.072 | <0.01 | 0.195 | <0.01 |
| PEG | 3.53Abc | 4.33Aa | 3.78Ab | 3.11Acd | 2.91Ad | |||||
| AB Log10 cfu/g FM | CK | 9.15Aa | 8.80Bab | 8.64Ab | 8.15Bc | 7.24Bd | 0.086 | <0.01 | <0.01 | 0.106 |
| PEG | 9.36Aa | 9.28Aab | 8.95Ab | 8.62Ac | 7.83Ad | |||||
| Days | Treatment | Sobs | Shannon | Simpson | Ace | Chao | Good’s Coverage |
|---|---|---|---|---|---|---|---|
| Before ensiled | —— | 143 | 0.89 | 0.68 | 201.07 | 168.3 | 0.999 |
| 3 | CK | 852A | 2.78 | 0.17 | 852A | 852A | 0.999 |
| PEG | 290.33B | 2.20 | 0.23 | 439.9B | 433.4B | 0.997 | |
| 7 | CK | 329 | 2.41 | 0.21 | 456.05 | 470.25 | 0.997 |
| PEG | 552.67 | 2.42 | 0.24 | 596.04 | 604.67 | 0.999 | |
| 14 | CK | 357 | 2.61 | 0.18B | 544.22 | 488.43 | 0.997 |
| PEG | 307.67 | 2.20 | 0.26A | 423.48 | 427.06 | 0.997 | |
| 30 | CK | 410.67A | 2.23 | 0.18B | 538.13 | 554.1 | 0.999 |
| PEG | 352.67B | 2.10 | 0.31A | 501.24 | 496.38 | 0.999 | |
| 60 | CK | 229.33 | 1.88 | 0.31 | 317.79 | 295.08 | 0.999 |
| PEG | 199.67 | 1.85 | 0.28 | 289.93 | 269.35 | 0.999 |
| Days | PEG | Sobs | Shannon | Simpson | Ace | Chao | Goods Coverage |
|---|---|---|---|---|---|---|---|
| Before ensiled | N | 148 | 2.25 | 0.18 | 192.26 | 197.05 | 0.999 |
| 3 | Y | 122.67B | 2.97A | 0.10 | 127.30B | 125.66B | 0.999 |
| N | 169.67A | 2.44B | 0.13 | 193.21A | 192.22A | 0.999 | |
| 7 | Y | 79.67B | 2.84 | 0.10 | 81.87B | 80.92B | 0.999 |
| N | 165.67A | 2.63 | 0.11 | 175.83A | 177.83A | 0.999 | |
| 14 | Y | 55.00B | 2.48 | 0.11 | 58.31B | 57.72B | 0.999 |
| N | 126.00A | 2.57 | 0.18 | 130.69A | 132.56A | 0.999 | |
| 30 | Y | 36.33B | 2.69 | 0.09 | 47.63B | 39.00B | 0.999 |
| N | 125.33A | 2.61 | 0.11 | 129.43A | 130.26A | 0.999 | |
| 60 | Y | 59.00 | 3.08 | 0.11 | 66.81 | 60.50 | 0.999 |
| N | 51.00 | 2.95 | 0.08 | 51.98 | 51.61 | 0.999 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, R.; Zhang, F.; Wang, T.; Zhang, Y.; Li, X.; Chen, Y.; Ma, C. Effect of Intrinsic Tannins on the Fermentation Quality and Associated with the Bacterial and Fungal Community of Sainfoin Silage. Microorganisms 2022, 10, 844. https://doi.org/10.3390/microorganisms10050844
Huang R, Zhang F, Wang T, Zhang Y, Li X, Chen Y, Ma C. Effect of Intrinsic Tannins on the Fermentation Quality and Associated with the Bacterial and Fungal Community of Sainfoin Silage. Microorganisms. 2022; 10(5):844. https://doi.org/10.3390/microorganisms10050844
Chicago/Turabian StyleHuang, Rongzheng, Fanfan Zhang, Ting Wang, Yulin Zhang, Xiao Li, Yongcheng Chen, and Chunhui Ma. 2022. "Effect of Intrinsic Tannins on the Fermentation Quality and Associated with the Bacterial and Fungal Community of Sainfoin Silage" Microorganisms 10, no. 5: 844. https://doi.org/10.3390/microorganisms10050844
APA StyleHuang, R., Zhang, F., Wang, T., Zhang, Y., Li, X., Chen, Y., & Ma, C. (2022). Effect of Intrinsic Tannins on the Fermentation Quality and Associated with the Bacterial and Fungal Community of Sainfoin Silage. Microorganisms, 10(5), 844. https://doi.org/10.3390/microorganisms10050844