
Food Is Reservoir of MDR Salmonella: Prevalence of ESBLs Profiles and Resistance Genes in Strains Isolated from Food

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Salmonella Isolation
2.2. Antibiotic Susceptibility Profile Determination
2.3. ESBL Production Evaluation by Double-Disc Test
2.4. Detection of Beta-Lactamase Genes
3. Results
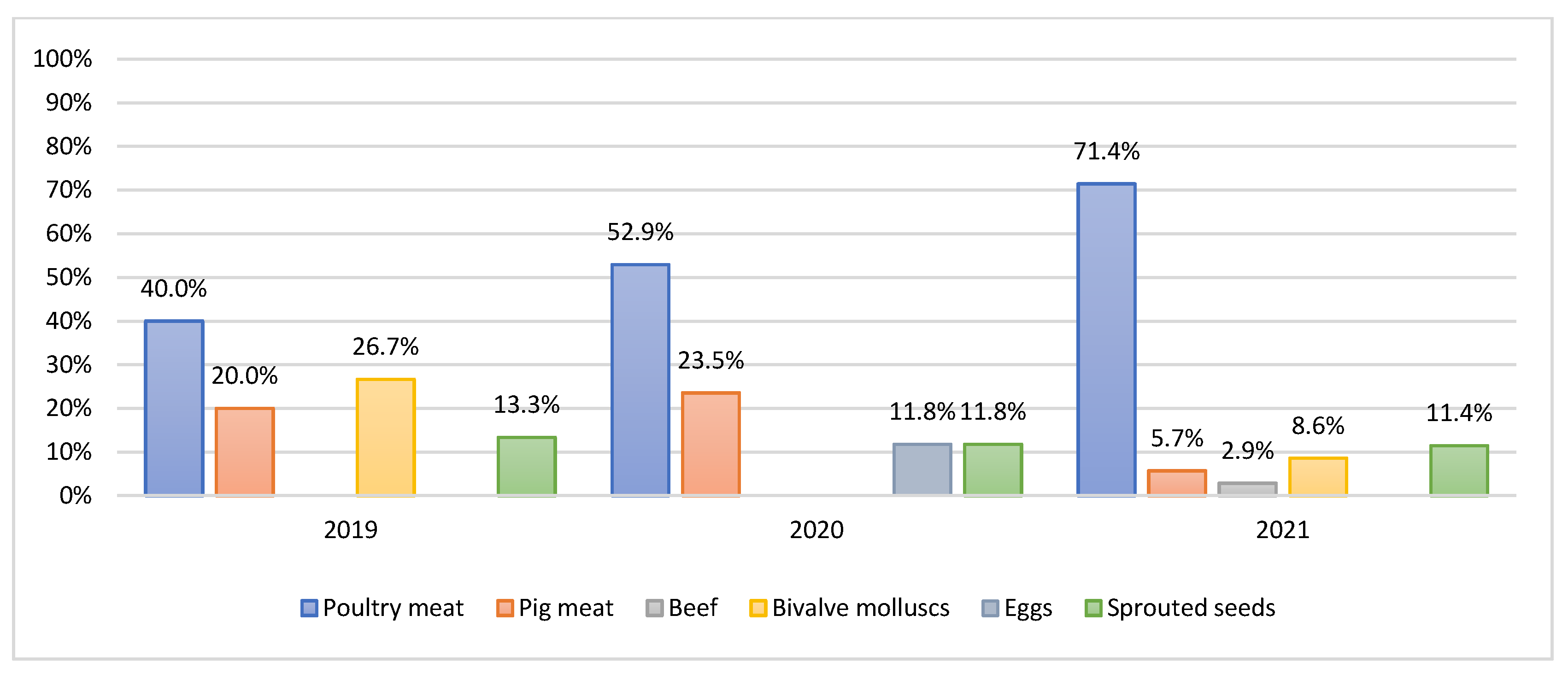
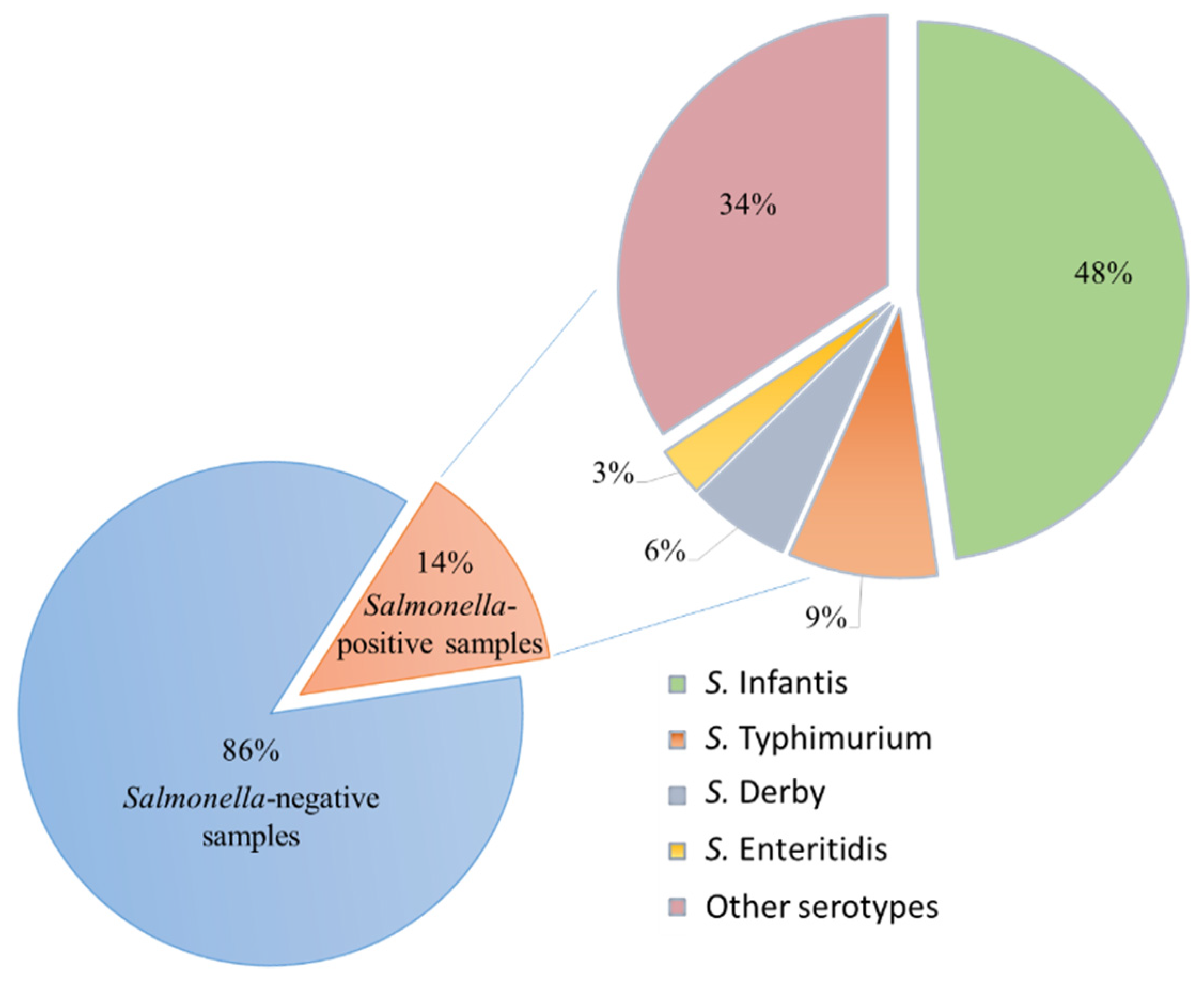
3.1. Isolation Results
3.2. Antibiotic Susceptibility and ESBL Production Test Results
3.3. Detection of Beta-Lactamase Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar] [CrossRef]
- Liu, H.; Whitehouse, C.A.; Li, B. Presence and Persistence of Salmonella in Water: The Impact on Microbial Quality of Water and Food Safety. Front. Public Health 2018, 6, 159. [Google Scholar] [CrossRef] [PubMed]
- Ehuwa, O.; Jaiswal, A.K.; Jaiswal, S. Salmonella, Food Safety and Food Handling Practices. Foods 2021, 10, 907. [Google Scholar] [CrossRef] [PubMed]
- Commission of the European Communities Regulation (EC) No 2073/2005 of 15 November 2005 on Microbiological Criteria for Foodstuffs. 2005. Available online: http://data.europa.eu/eli/reg/2005/2073/oj (accessed on 14 December 2021).
- Eng, S.-K.; Pusparajah, P.; Ab Mutalib, N.-S.; Ser, H.-L.; Chan, K.-G.; Lee, L.-H. Salmonella: A Review on Pathogenesis, Epidemiology and Antibiotic Resistance. Front. Life Sci. 2015, 8, 284–293. [Google Scholar] [CrossRef] [Green Version]
- Elnekave, E.; Hong, S.L.; Lim, S.; Boxrud, D.; Rovira, A.; Mather, A.E.; Perez, A.; Alvarez, J. Transmission of Multidrug-Resistant Salmonella Enterica Subspecies Enterica 4,[5],12:I:- Sequence Type 34 between Europe and the United States. Emerg. Infect. Dis. 2020, 26, 3034–3038. [Google Scholar] [CrossRef] [PubMed]
- Proietti, P.C.; Stefanetti, V.; Musa, L.; Zicavo, A.; Dionisi, A.M.; Bellucci, S.; Mensa, A.L.; Menchetti, L.; Branciari, R.; Ortenzi, R.; et al. Genetic Profiles and Antimicrobial Resistance Patterns of Salmonella Infantis Strains Isolated in Italy in the Food Chain of Broiler Meat Production. Antibiotics 2020, 9, 814. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2018/2019. EFSA J. 2021, 19, e06490. [Google Scholar] [CrossRef]
- Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014; ISBN 978-92-4-156474-8.
- Castro-Vargas, R.E.; Herrera-Sánchez, M.P.; Rodríguez-Hernández, R.; Rondón-Barragán, I.S. Antibiotic Resistance in Salmonella Spp. Isolated from Poultry: A Global Overview. Vet. World 2020, 13, 2070–2084. [Google Scholar] [CrossRef]
- Campos, J.; Mourão, J.; Peixe, L.; Antunes, P. Non-Typhoidal Salmonella in the Pig Production Chain: A Comprehensive Analysis of Its Impact on Human Health. Pathogens 2019, 8, 19. [Google Scholar] [CrossRef] [Green Version]
- Iredell, J.; Brown, J.; Tagg, K. Antibiotic Resistance in Enterobacteriaceae: Mechanisms and Clinical Implications. BMJ 2016, 352, h6420. [Google Scholar] [CrossRef]
- Philippon, A.; Slama, P.; Dény, P.; Labia, R. A Structure-Based Classification of Class A β-Lactamases, a Broadly Diverse Family of Enzymes. Clin. Microbiol. Rev. 2016, 29, 29–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Q.; Wu, Q.; Zhang, S.; Zhang, J.; Yang, G.; Wang, J.; Xue, L.; Chen, M. Characterization of Extended-Spectrum β-Lactamase-Producing Enterobacteriaceae From Retail Food in China. Front. Microbiol. 2018, 9, 1709. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.T.A.; Kanki, M.; Nguyen, P.D.; Le, H.T.; Ngo, P.T.; Tran, D.N.M.; Le, N.H.; Dang, C.V.; Kawai, T.; Kawahara, R.; et al. Prevalence, Antibiotic Resistance, and Extended-Spectrum and AmpC β-Lactamase Productivity of Salmonella Isolates from Raw Meat and Seafood Samples in Ho Chi Minh City, Vietnam. Int. J. Food Microbiol. 2016, 236, 115–122. [Google Scholar] [CrossRef]
- Colosi, I.A.; Baciu, A.M.; Opriș, R.V.; Peca, L.; Gudat, T.; Simon, L.M.; Colosi, H.A.; Costache, C. Prevalence of ESBL, AmpC and Carbapenemase-Producing Enterobacterales Isolated from Raw Vegetables Retailed in Romania. Foods 2020, 9, 1726. [Google Scholar] [CrossRef]
- Kim, H.-S.; Chon, J.-W.; Kim, Y.-J.; Kim, D.-H.; Kim, M.; Seo, K.-H. Prevalence and Characterization of Extended-Spectrum-β-Lactamase-Producing Escherichia Coli and Klebsiella Pneumoniae in Ready-to-Eat Vegetables. Int. J. Food Microbiol. 2015, 207, 83–86. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Biological Hazards (BIOHAZ); Koutsoumanis, K.; Allende, A.; Álvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; et al. Role Played by the Environment in the Emergence and Spread of Antimicrobial Resistance (AMR) through the Food Chain. EFSA J. 2021, 19, 6651. [Google Scholar] [CrossRef]
- ISO 6579-1:2017; Microbiology of the Food Chain—Horizontal Method for the Detection, Enumeration and Serotyping of Salmonella—Part 1: Detection of Salmonella spp. International Organization for Standardization; AFNOR: Saint-Denis, France, 2017.
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility: Supplement M100, 31st ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2021; ISBN 978-1-68440-105-5. [Google Scholar]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Guideline for the Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance, Version 2.0; 2017. Available online: https://www.eucast.org/resistance_mechanisms/ (accessed on 14 December 2021).
- Zhao, X.; Hu, M.; Zhang, Q.; Zhao, C.; Zhang, Y.; Li, L.; Qi, J.; Luo, Y.; Zhou, D.; Liu, Y. Characterization of Integrons and Antimicrobial Resistance in Salmonella from Broilers in Shandong, China. Poult. Sci. 2020, 99, 7046–7054. [Google Scholar] [CrossRef] [PubMed]
- Gargano, V.; Sciortino, S.; Gambino, D.; Costa, A.; Agozzino, V.; Reale, S.; Alduina, R.; Vicari, D. Antibiotic Susceptibility Profile and Tetracycline Resistance Genes Detection in Salmonella Spp. Strains Isolated from Animals and Food. Antibiotics 2021, 10, 809. [Google Scholar] [CrossRef] [PubMed]
- Gargano, V.; Gambino, D.; Migliore, S.; Vitale, M.; Sciortino, S.; Costa, A.; Vicari, D. Can Human Handling Increase the Presence of Multidrug Resistance (MDR) in Salmonella Spp. Isolated from Food Sources? Microorganisms 2021, 9, 2018. [Google Scholar] [CrossRef] [PubMed]
- Dhanani, A.S.; Block, G.; Dewar, K.; Forgetta, V.; Topp, E.; Beiko, R.G.; Diarra, M.S. Genomic Comparison of Non-Typhoidal Salmonella Enterica Serovars Typhimurium, Enteritidis, Heidelberg, Hadar and Kentucky Isolates from Broiler Chickens. PLoS ONE 2015, 10, e0128773. [Google Scholar] [CrossRef]
- Cantón, R.; Coque, T.M. The CTX-M β-Lactamase Pandemic. Curr. Opin. Microbiol. 2006, 9, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Alcalá, L.; Alonso, C.A.; Simón, C.; González-Esteban, C.; Orós, J.; Rezusta, A.; Ortega, C.; Torres, C. Wild Birds, Frequent Carriers of Extended-Spectrum β-Lactamase (ESBL) Producing Escherichia Coli of CTX-M and SHV-12 Types. Microb. Ecol. 2016, 72, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Liakopoulos, A.; Mevius, D.; Ceccarelli, D. A Review of SHV Extended-Spectrum β-Lactamases: Neglected Yet Ubiquitous. Front. Microbiol. 2016, 7, 1374. [Google Scholar] [CrossRef] [PubMed]
- Bush, K. Past and Present Perspectives on β-Lactamases. Antimicrob. Agents Chemother. 2018, 62, e01076-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groussin, M.; Poyet, M.; Sistiaga, A.; Kearney, S.M.; Moniz, K.; Noel, M.; Hooker, J.; Gibbons, S.M.; Segurel, L.; Froment, A.; et al. Elevated Rates of Horizontal Gene Transfer in the Industrialized Human Microbiome. Cell 2021, 184, 2053–2067.e18. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Target | Primers | Sequence (5′-3′) | Amplicon Size (bp) | Reference |
|---|---|---|---|---|
| blaTEM | blaTEM_F | ATTCTTGAAGACGAAAGGGC | 661 | [22] |
| blaTEM_R | ACGCTCAGTGGAACGAAAAC | |||
| blaCTX-M | blaCTX-M_F | CGCTTTGCGATGTGCAG | 585 | |
| blaCTX-M_R | ACCGCGATATCGTTGGT | |||
| blaSHV | blaSHV_F | CACTCAAGGATGTATTGTG | 807 | |
| blaSHV_R | TTAGCGTTGCCAGTGCTCG | |||
| blaOXA | blaOXA_F | ACACAATACATATCAACTTCGC | 590 | |
| blaOXA_R | AGTGTGTTTAGAATGGTGATC |
| ID | Food | Salmonella Serotype | Isolation Year | Resistance | ESBL Production |
|---|---|---|---|---|---|
| AL-3 | Poultry meat | S. Infantis | 2019 | AMP, CTX, NAL, SXT, TET | − |
| AL-11 | Poultry meat | S. Newport | 2019 | KAN, AMP, SXT, TET | + |
| AL-14 | Poultry meat | S. Infantis | 2019 | KAN, AMP, CTX, NAL, TET | − |
| AL-15 | Poultry meat | S. Infantis | 2019 | KAN, AMP, CTX, NAL, SXT, TET | − |
| AL-30 | Poultry meat | S. Infantis | 2020 | KAN, NAL, SXT, TET | + |
| AL-20 | Poultry meat | S. Infantis | 2020 | NAL, SXT, TET | + |
| AL-21 | Poultry meat | S. Infantis | 2020 | KAN, NAL, SXT, TET | + |
| AL-25 | Poultry meat | S. Infantis | 2020 | STR, AMP, NAL, SXT, TET | + |
| AL-26 | Poultry meat | S. Infantis | 2020 | STR, NAL, SXT, TET | + |
| AL-27 | Poultry meat | S. Infantis | 2020 | KAN, STR, NAL, SXT, TET | + |
| AL-32 | Poultry meat | S. Infantis | 2020 | KAN, STR, NAL, SXT, TET | + |
| AL-34 | Poultry meat | S. Infantis | 2021 | KAN, STR, AMP, CTX, NAL, LEVO, CHL | − |
| AL-35 | Poultry meat | S. Infantis | 2021 | KAN, STR, AMP, CTX, NAL, LEVO, CHL | − |
| AL-37 | Poultry meat | S. Agona | 2021 | STR, AMP, SXT | + |
| AL-38 | Pig meat | S. Salamae | 2021 | KAN, GEN, TOB, AMP, AMC, NAL, SXT, CHL | + |
| AL-39 | Poultry meat | S. Infantis | 2021 | KAN, SXT, TET | + |
| AL-43 | Poultry meat | S. Infantis | 2021 | KAN, AMP, STR, NAL, SXT, TET | + |
| AL-44 | Poultry meat | S. Infantis | 2021 | KAN, TOB, AMP, CTX, CRO, NAL, SXT | − |
| AL-45 | Poultry meat | S. Infantis | 2021 | AMP, CTX, CRO, NAL, SXT, TET | − |
| AL-46 | Poultry meat | S. Infantis | 2021 | STR, AMP, NAL, TET | − |
| AL-47 | Beef | S. Cerro | 2021 | AMP, AMC, CTX, CRO, NAL, SXT, TET | − |
| AL-48 | Poultry meat | S. Infantis | 2021 | KAN, GEN, TOB, AMP, AMC, CTX, CRO, NAL, SXT, TET | − |
| AL-49 | Poultry meat | S. Infantis | 2021 | KAN, AMP, AMC, CTX, CRO, NAL, SXT, TET | − |
| AL-50 | Poultry meat | S. Infantis | 2021 | KAN, AMP, AMC, CTX, CRO, SXT, TET | − |
| AL-51 | Pig meat | S. Typhimurium | 2021 | STR, AMP, TET | + |
| AL-52 | Poultry meat | S. Infantis | 2021 | AMP, CTX, CAZ, CRO, NAL, SXT, TET | + |
| AL-53 | Poultry meat | S. Kentucky | 2021 | STR, AMP, CAZ, CTX, CRO, NAL, SXT, TET | + |
| AL-56 | Poultry meat | S. Infantis | 2021 | AMP, AMC, CRO, NAL, SXT, TET | − |
| AL-57 | Poultry meat | S. Infantis | 2021 | TOB, AMP, AMC, CTX, CRO, NAL, SXT, TET | + |
| AL-58 | Poultry meat | S. Infantis | 2021 | AMP, AMC, CTX, NAL, SXT, TET | + |
| AL-59 | Poultry meat | S. Infantis | 2021 | GEN, AMP, CTX, NAL, SXT, TET | − |
| AL-60 | Poultry meat | S. Infantis | 2021 | KAN, TOB, AMP, CTX, NAL, SXT, TET | + |
| AL-63 | Poultry meat | S. Infantis | 2021 | STR, AMP, CTX, CAZ, NAL, SXT, TET, CHL | + |
| AL-65 | Poultry meat | S. Infantis | 2021 | AMP, CTX, CAZ, CRO, NAL, SXT, TET | − |
| AL-66 | Poultry meat | S. Infantis | 2021 | KAN, STR, AMP, NAL, TET | − |
| AL-67 | Poultry meat | S. Infantis | 2021 | KAN, AMP, NAL, TET | − |
| ID Strains | Food | Salmonella Serotype | ESBL Production | bla Gene Detected |
|---|---|---|---|---|
| AL-11 | Poultry meat | S. Newport | + | blaTEM, blaSHV |
| AL-30 | Poultry meat | S. Infantis | + | blaCTX-M |
| AL-20 | Poultry meat | S. Infantis | + | blaSHV |
| AL-21 | Poultry meat | S. Infantis | + | blaCTX-M |
| AL-25 | Poultry meat | S. Infantis | + | blaSHV |
| AL-26 | Poultry meat | S. Infantis | + | blaSHV |
| AL-27 | Poultry meat | S. Infantis | + | blaCTX-M |
| AL-32 | Poultry meat | S. Infantis | + | blaSHV |
| AL-37 | Poultry meat | S. Agona | + | blaSHV |
| AL-38 | Pig meat | S. Salamae | + | blaCTX-M, blaSHV |
| AL-39 | Poultry meat | S. Infantis | + | blaSHV |
| AL-43 | Poultry meat | S. Infantis | + | blaCTX-M |
| AL-51 | Pig meat | S. Typhimurium | + | blaSHV |
| AL-52 | Poultry meat | S. Infantis | + | blaSHV |
| AL-53 | Poultry meat | S. Kentucky | + | blaCTX-M |
| AL-57 | Poultry meat | S. Infantis | + | blaCTX-M |
| AL-58 | Poultry meat | S. Infantis | + | blaCTX-M, blaSHV |
| AL-60 | Poultry meat | S. Infantis | + | blaSHV |
| AL-63 | Poultry meat | S. Infantis | + | blaCTX-M, blaSHV |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gambino, D.; Gargano, V.; Butera, G.; Sciortino, S.; Pizzo, M.; Oliveri, G.; Cardamone, C.; Piraino, C.; Cassata, G.; Vicari, D.; et al. Food Is Reservoir of MDR Salmonella: Prevalence of ESBLs Profiles and Resistance Genes in Strains Isolated from Food. Microorganisms 2022, 10, 780. https://doi.org/10.3390/microorganisms10040780
Gambino D, Gargano V, Butera G, Sciortino S, Pizzo M, Oliveri G, Cardamone C, Piraino C, Cassata G, Vicari D, et al. Food Is Reservoir of MDR Salmonella: Prevalence of ESBLs Profiles and Resistance Genes in Strains Isolated from Food. Microorganisms. 2022; 10(4):780. https://doi.org/10.3390/microorganisms10040780
Chicago/Turabian StyleGambino, Delia, Valeria Gargano, Gaspare Butera, Sonia Sciortino, Mariangela Pizzo, Giuseppa Oliveri, Cinzia Cardamone, Chiara Piraino, Giovanni Cassata, Domenico Vicari, and et al. 2022. "Food Is Reservoir of MDR Salmonella: Prevalence of ESBLs Profiles and Resistance Genes in Strains Isolated from Food" Microorganisms 10, no. 4: 780. https://doi.org/10.3390/microorganisms10040780
APA StyleGambino, D., Gargano, V., Butera, G., Sciortino, S., Pizzo, M., Oliveri, G., Cardamone, C., Piraino, C., Cassata, G., Vicari, D., & Costa, A. (2022). Food Is Reservoir of MDR Salmonella: Prevalence of ESBLs Profiles and Resistance Genes in Strains Isolated from Food. Microorganisms, 10(4), 780. https://doi.org/10.3390/microorganisms10040780