
Bacterial Communities of Forest Soils along Different Elevations: Diversity, Structure, and Functional Composition with Potential Impacts on CO2 Emission


Abstract
:1. Introduction
2. Materials and Methods
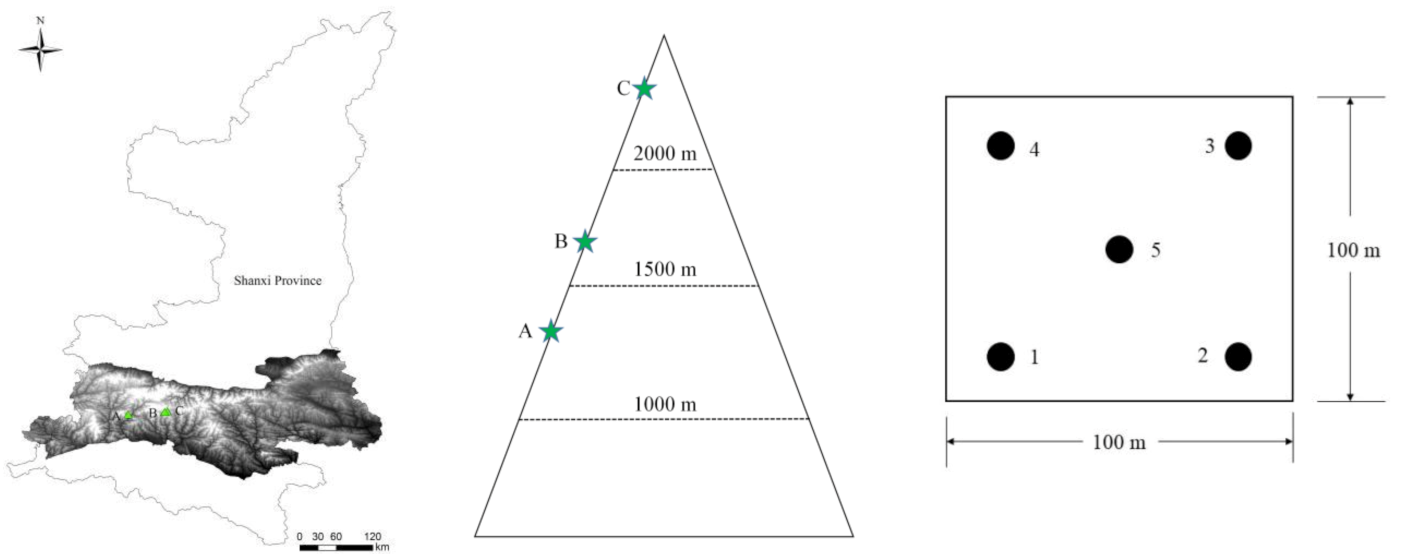
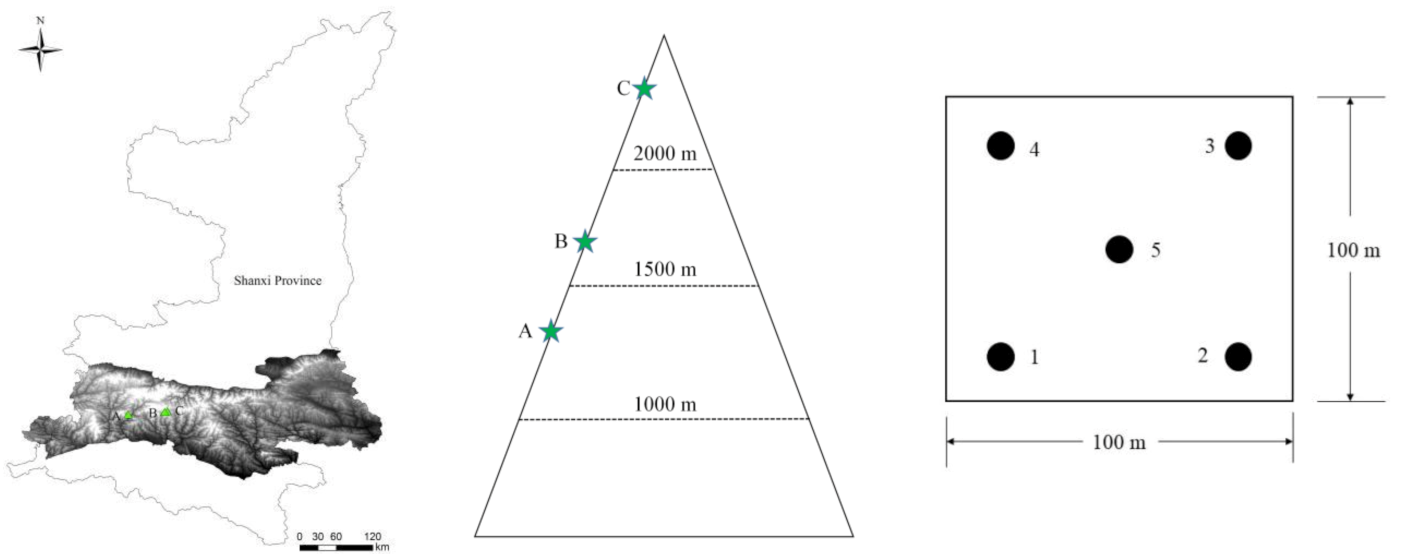
2.1. Site and Soil Sampling
2.2. Soil DNA Extraction, PCR Amplification, and Illumina Sequencing
2.3. Raw Data Processing and Functional Annotation
2.4. CO2 Efflux Measurements
2.5. Statistical Analyses
3. Results
3.1. Soil Bacterial Community Structure and α-Diversity
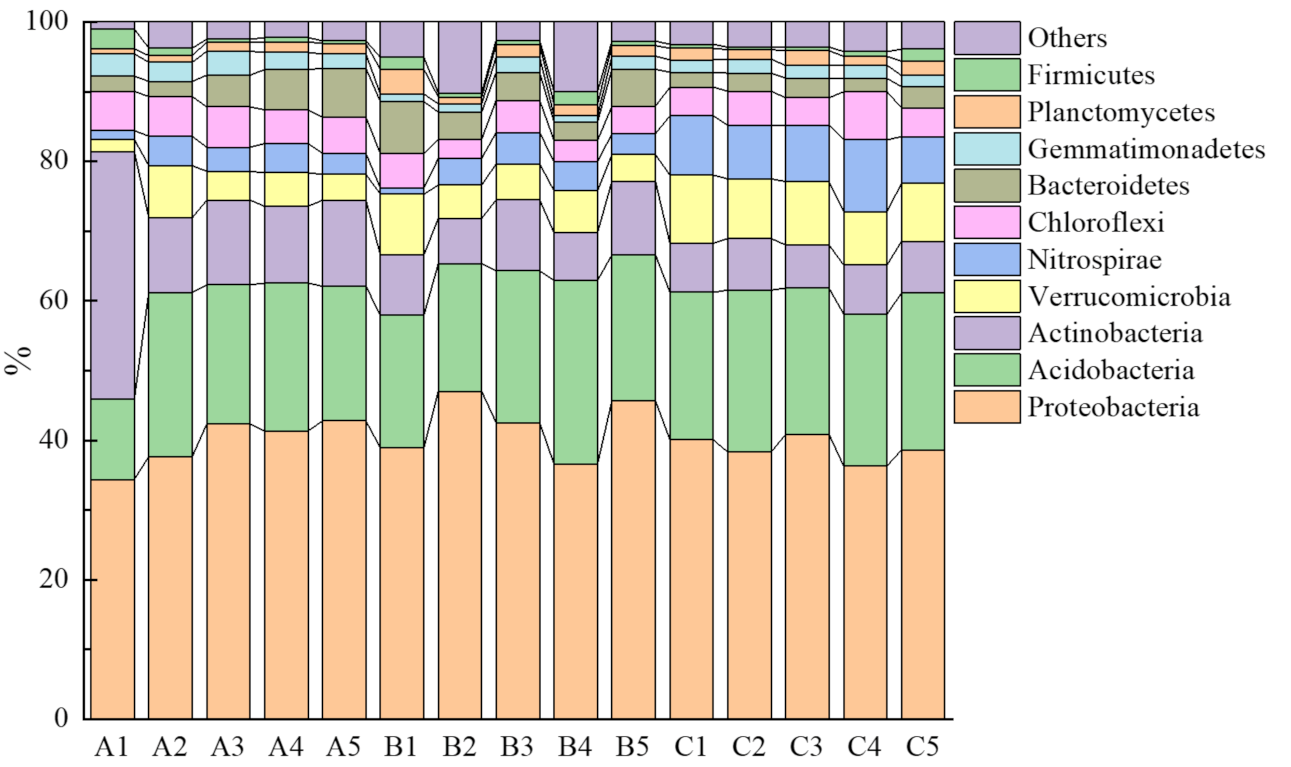
3.2. Functional Group Abundances of Bacterial Communities
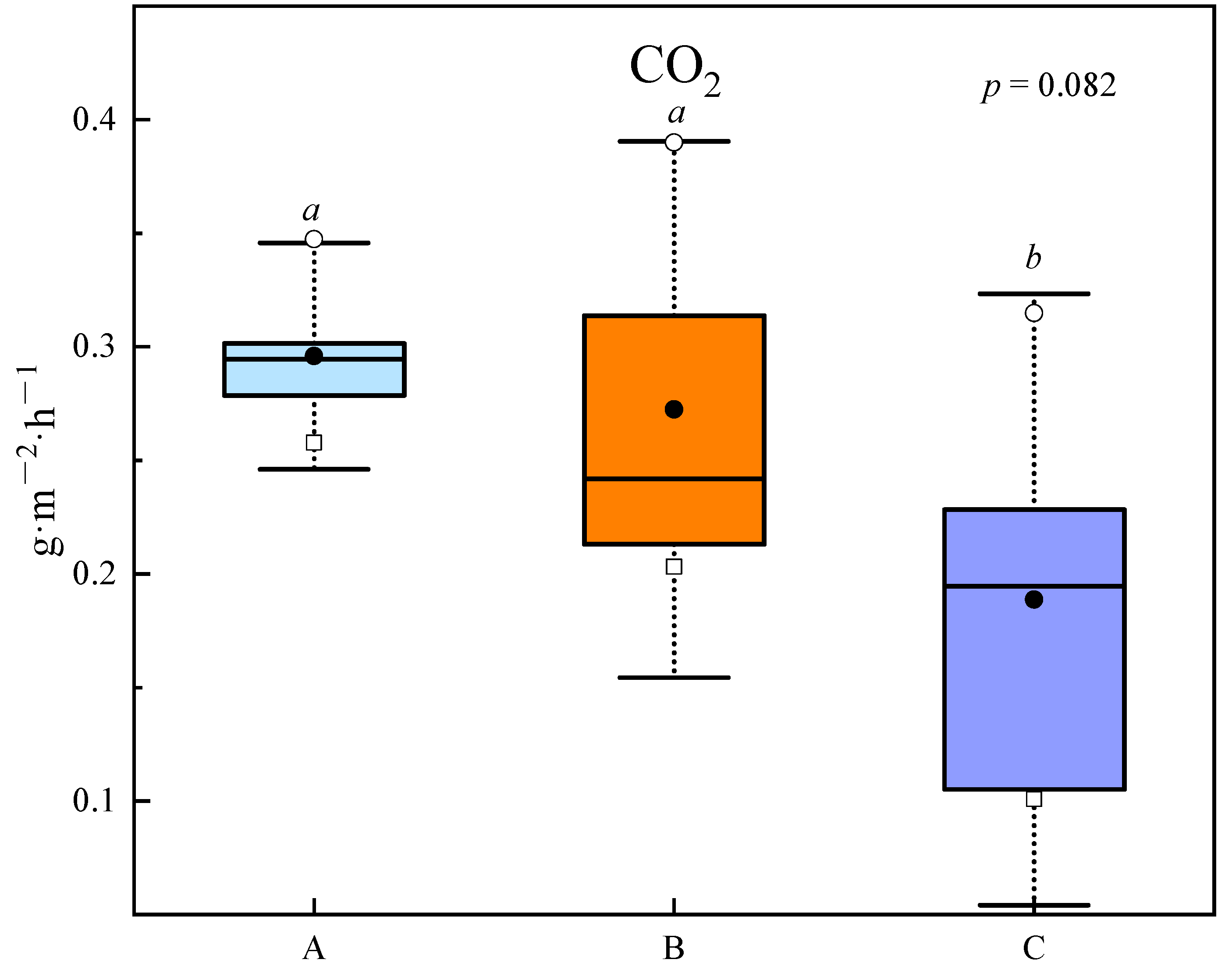
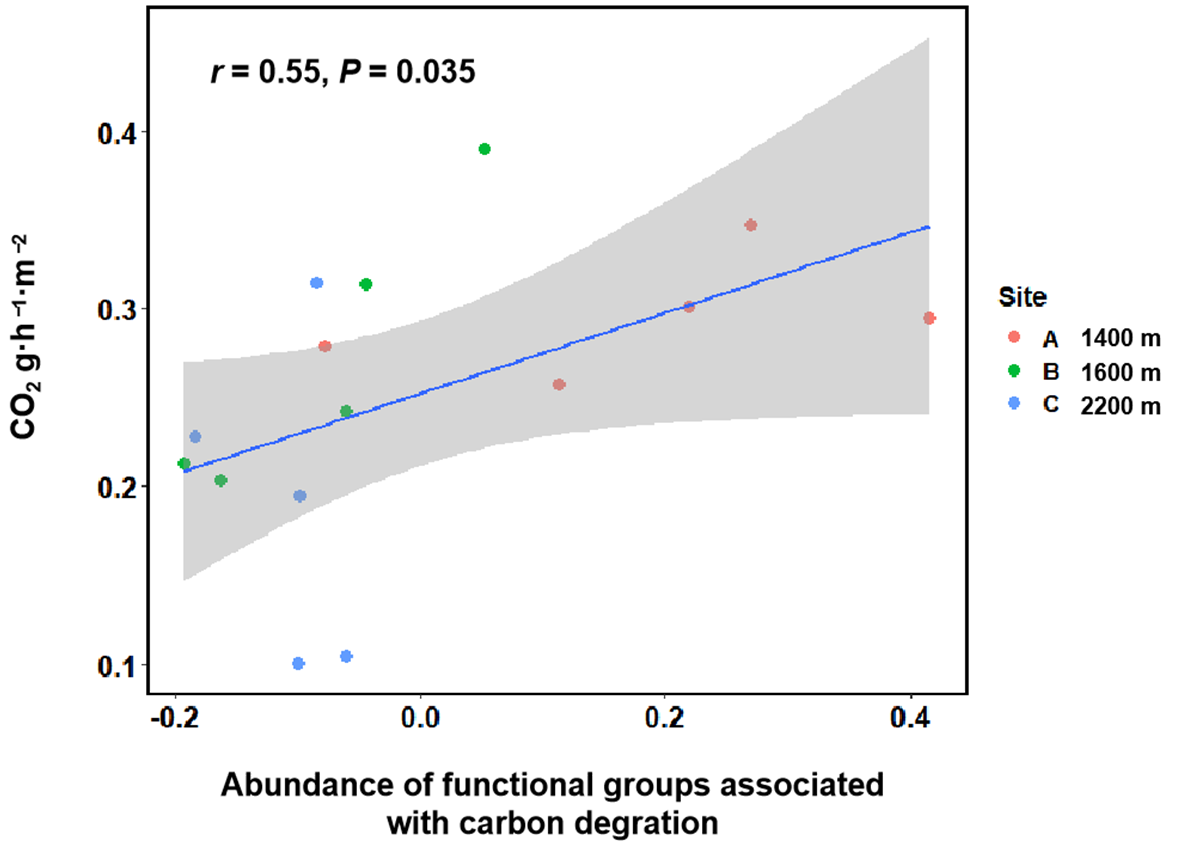
3.3. CO2 Effluxes and the Associations with Bacterial Communities and Functional Composition
3.4. Effects of Environmental Factors on Soil Bacterial Communities and Their Functional Groups
4. Discussion
4.1. Correlations between Environmental Factors, Bacterial α-Diversity and Taxonomic Composition
4.2. Correlations between Environmental Factors and Bacterial Functional Composition of Soils
4.3. Effects of Soil Bacterial Communities and Bacterial Functional Groups on CO2 Effluxes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tian, Q.; Jiang, Y.; Tang, Y.; Wu, Y.; Tang, Z.; Liu, F. Soil pH and Organic Carbon Properties Drive Soil Bacterial Communities in Surface and Deep Layers Along an Elevational Gradient. Front. Microbiol. 2021, 12, 646124. [Google Scholar] [CrossRef]
- Liu, J.; Dang, P.; Gao, Y.; Zhu, H.; Zhu, H.; Zhao, F.; Zhao, Z. Effects of tree species and soil properties on the composition and diversity of the soil bacterial community following afforestation. For. Ecol. Manag. 2018, 427, 342–349. [Google Scholar] [CrossRef]
- Kalia, V.; Gong, C.; Patel, S.; Lee, J.-K. Regulation of Plant Mineral Nutrition by Signal Molecules. Microorganisms 2021, 9, 774. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an Ecological Classification of Soil Bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Zhang, W.; Zhong, Z.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Differential responses of soil microbial biomass, diversity, and compositions to altitudinal gradients depend on plant and soil characteristics. Sci. Total Environ. 2018, 610–611, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Louca, S.; Jacques, S.M.D.S.; Pires, A.P.F.; Leal, J.S.; Gonzalez, A.L.; Doebeli, M.; Farjalla, V. Functional structure of the bromeliad tank microbiome is strongly shaped by local geochemical conditions. Environ. Microbiol. 2017, 19, 3132–3151. [Google Scholar] [CrossRef]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 1272–1277. [Google Scholar] [CrossRef]
- Wang, N.F.; Zhang, T.; Zhang, F.; Wang, E.T.; He, J.F.; Ding, H.; Zhang, B.T.; Liu, J.; Ran, X.B.; Zang, J.Y. Diversity and structure of soil bacterial communities in the Fildes Region (maritime Antarctica) as revealed by 454 pyrosequencing. Front. Microbiol. 2015, 6, 1188. [Google Scholar] [CrossRef] [Green Version]
- Fanin, N.; Gundale, M.J.; Farrell, M.; Ciobanu, M.; Baldock, J.A.; Nilsson, M.-C.; Kardol, P.; Wardle, D.A. Consistent effects of biodiversity loss on multifunctionality across contrasting ecosystems. Nat. Ecol. Evol. 2018, 2, 269–278. [Google Scholar] [CrossRef]
- Fierer, N.; McCain, C.M.; Meir, P.; Zimmermann, M.; Rapp, J.; Silman, M.R.; Knight, R. Microbes do not follow the elevational diversity patterns of plants and animals. Ecology 2011, 92, 797–804. [Google Scholar] [CrossRef] [Green Version]
- Ju, H.; Shen, G.Z.; Xu, W.T.; Zhao, C.M.; Su, L.; Wang, Y.; Xie, Z.Q.; Zhang, Q.L. The emission of CH4, CO2, and N2O in the typical forest soils of Shennongjia under the precipitation reduction. Acta Ecol. Sin. 2016, 36, 6397–6408. [Google Scholar]
- Tang, X.; Zhou, G.; Zhang, D.; Zhou, C. Soil-atmospheric exchange of CO2, CH4, and N2O in three subtropical forest ecosystems in southern China. Glob. Chang. Biol. 2006, 12, 546–560. [Google Scholar] [CrossRef]
- Patel, S.K.; Das, D.; Kim, S.C.; Cho, B.-K.; Kalia, V.C.; Lee, J.-K. Integrating strategies for sustainable conversion of waste biomass into dark-fermentative hydrogen and value-added products. Renew. Sustain. Energy Rev. 2021, 150, 111491. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, S.; Li, S.Z.; Deng, Y. Advances in molecular ecology on microbial functional genes of carbon cycle. Microbiol. China 2017, 44, 1676–1689. [Google Scholar]
- Liu, X.; Wu, P.; Shao, X.; Songer, M.; Cai, Q.; Zhu, Y.; He, X. Spatiotemporally monitoring forest landscape for giant panda habitat through a high learning-sensitive neural network in Guanyinshan Nature Reserve in the Qinling Mountains, China. Environ. Earth Sci. 2017, 76, 589. [Google Scholar] [CrossRef]
- Shi, Z.; Wang, F.; Zhang, K.; Chen, Y. Diversity and distribution of arbuscular mycorrhizal fungi along altitudinal gradients in Mount Taibai of the Qinling Mountains. Can. J. Microbiol. 2014, 60, 811–818. [Google Scholar] [CrossRef]
- Al-Shammary, A.A.G.; Kouzani, A.Z.; Kaynak, A.; Khoo, S.Y.; Norton, M.; Gates, W. Soil Bulk Density Estimation Methods: A Review. Pedosphere 2018, 28, 581–596. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Sansupa, C.; Mohamed Wahdan, S.F.; Hossen, S.; Disayathanoowat, T.; Wubet, T.; Purahong, W. Can We Use Functional Annotation of Prokaryotic Taxa (FAPROTAX) to Assign the Ecological Functions of Soil Bacteria? Appl. Sci. 2021, 11, 688. [Google Scholar] [CrossRef]
- Wang, Y.S.; Wang, Y.H. Quick measurement of CH4, CO2 and N2O emissions from a short-plant ecosystem. Adv. Atmos. Sci. 2003, 20, 842–844. [Google Scholar]
- Song, C.; Zhang, J.; Wang, Y.; Wang, Y.; Zhao, Z. Emission of CO2, CH4 and N2O from freshwater marsh in northeast of China. J. Environ. Manag. 2008, 88, 428–436. [Google Scholar] [CrossRef]
- Du, C.; Geng, Z.C. Effects of Seasonal Changes of Soil Properties on Soil Bacterial Diversity and Community Structure of Two Forest Types in Qinling Mountains. J. Northwest. For. Univ. 2021, 36, 54–61. [Google Scholar]
- He, J.; Yan, B.; Li, J.S.; Fu, G.; Qi, Y.; Ma, Y.W.; Qiao, M.P. Altitude Distribution Patterns and Regional Differences of Soil Bacterial Community in Northern Slopes in the Middle Qinling Mountains. Res. Environ. Sci. 2019, 32, 1374–1383. [Google Scholar]
- Wang, M.; Sun, H.; Xu, L.; Xu, Z. Bacterial diversity in tea plant (Camellia sinensis) rhizosphere soil from Qinling Mountains and its relationship with environmental elements. Plant. Soil 2021, 460, 403–415. [Google Scholar] [CrossRef]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; van Bodegom, P.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H.; et al. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef]
- Kim, H.-S.; Lee, S.-H.; Jo, H.Y.; Finneran, K.T.; Kwon, M.J. Diversity and composition of soil Acidobacteria and Proteobacteria communities as a bacterial indicator of past land-use change from forest to farmland. Sci. Total Environ. 2021, 797, 148944. [Google Scholar] [CrossRef]
- Xia, Y.; He, X.; Feng, Z.; Zhang, Q.; Yang, H. A comprehensive analysis of the microbial diversity in natural and engineered ecosystems based on high-throughput sequencing of 16S rRNA gene. Int. Biodeterior. Biodegrad. 2019, 140, 160–168. [Google Scholar] [CrossRef]
- Ranjard, L.; Dequiedt, S.; Jolivet, C.; Saby, N.P.A.; Thioulouse, J.; Harmand, J.; Loisel, P.; Rapaport, A.; Fall, S.; Simonet, P.; et al. Biogeography of soil microbial communities: A review and a description of the ongoing french national initiative. Agron. Sustain. Dev. 2010, 30, 359–365. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Zhang, J.; Chen, L.; Zhang, C.; Yu, Z. Temperature and Straw Quality Regulate the Microbial Phospholipid Fatty Acid Composition Associated with Straw Decomposition. Pedosphere 2016, 26, 386–398. [Google Scholar] [CrossRef]
- Griffiths, R.I.; Thomson, B.C.; James, P.; Bell, T.; Bailey, M.; Whiteley, A. The bacterial biogeography of British soils. Environ. Microbiol. 2011, 13, 1642–1654. [Google Scholar] [CrossRef]
- Ganzert, L.; Lipski, A.; Hubberten, H.-W.; Wagner, D. The impact of different soil parameters on the community structure of dominant bacteria from nine different soils located on Livingston Island, South Shetland Archipelago, Antarctica. Fems Microbiol. Ecol. 2011, 76, 476–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.H.; Chu, H.L.; Dou, Q.; Xie, Q.Z.; Tang, M.; Sung, C.K.; Wang, C.Y. Phosphorus and Nitrogen Drive the Seasonal Dynamics of Bacterial Communities in Pinus Forest Rhizospheric Soil of the Qinling Mountains. Front. Microbiol. 2018, 9, 1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagar, I.; Nimonkar, Y.; Dhotre, D.; Shouche, Y.; Ranade, D.; Dewala, S.; Prakash, O. A Microcosm Model for the Study of Microbial Community Shift and Carbon Emission from Landfills. Indian J. Microbiol. 2022, 1–9. [Google Scholar] [CrossRef]
- Shen, C.; Xiong, J.; Zhang, H.; Feng, Y.; Lin, X.; Li, X.; Liang, W.; Chu, H. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [Green Version]
- Hirao, T.; Fujii, M.; Shigyo, N.; Kojima, H.; Fukui, M. Influence of understory vegetation on soil bacterial communities and nitrogen cycling gene abundance in cool-temperate and sub-alpine forests along an elevational gradient. Pedobiologia 2021, 87–88, 150746. [Google Scholar] [CrossRef]
- Han, W.; Wang, G.; Liu, J.; Ni, J. Effects of vegetation type, season, and soil properties on soil microbial community in subtropical forests. Appl. Soil Ecol. 2021, 158, 103813. [Google Scholar] [CrossRef]
- Sierra, C.A.; Trumbore, S.E.; Davidson, E.A.; Vicca, S. Janssens Sensitivity of decomposition rates of soil organic matter with respect to simultaneous changes in temperature and moisture. J. Adv. Modeling Earth Syst. 2015, 7, 335–356. [Google Scholar] [CrossRef]
- López-Mondéjar, R.; Voříšková, J.; Větrovský, T.; Baldrian, P. The bacterial community inhabiting temperate deciduous forests is vertically stratified and undergoes seasonal dynamics. Soil Biol. Biochem. 2015, 87, 43–50. [Google Scholar] [CrossRef]
- Lin, Y.-T.; Jia, Z.; Wang, D.; Chiu, C.-Y. Effects of temperature on the composition and diversity of bacterial communities in bamboo soils at different elevations. Biogeosciences 2017, 14, 4879–4889. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.; Peng, C.; Yang, B.; Song, H.; Li, Q.; Jiang, L.; Wei, G.; Wang, K.; Wang, H.; Liu, S.; et al. Contrasting soil bacterial community, diversity, and function in two forests in China. Front. Microbiol. 2018, 9, 1693. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Chen, W.; Luo, S.; Ma, L.; Li, X.; Tian, C. Rhizosphere microbiota assemblage associated with wild and cultivated soybeans grown in three types of soil suspensions. Arch. Agron. Soil Sci. 2019, 65, 74–87. [Google Scholar] [CrossRef]
- Khan, M.A.W.; Bohannan, B.J.M.; Nüsslein, K.; Tiedje, J.M.; Tringe, S.G.; Parladé, E.; Barberán, A.; Rodrigues, J.L.M. Deforestation impacts network co-occurrence patterns of microbial communities in Amazon soils. FEMS Microbiol. Ecol. 2018, 95, fiy230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Q.; Haowei, Y.; Zhang, Z.; Van Nostrand, J.D.; Wu, L.; Guo, X.; Feng, J.; Wang, M.; Yang, S.; Zhao, J.; et al. Microbial Functional Responses Explain Alpine Soil Carbon Fluxes under Future Climate Scenarios. mBio 2021, 12, e00761-20. [Google Scholar] [CrossRef] [PubMed]
- Louca, S.; Polz, M.F.; Mazel, F.; Albright, M.B.; Huber, J.A.; O’Connor, M.I.; Ackermann, M.; Hahn, A.S.; Srivastava, D.S.; Crowe, S.A.; et al. Function and functional redundancy in microbial systems. Nat. Ecol. Evol. 2018, 2, 936–943. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, Q.; Zheng, M.; Gao, Y.; Yuan, T.; Hale, L.; Van Nostrand, J.D.; Zhou, J.; Wan, S.; Yang, Y. Microbial functional traits are sensitive indicators of mild disturbance by lamb grazing. ISME J. 2019, 13, 1370–1373. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Hu, B.X.; Ren, H.; Zhang, J. Composition and functional diversity of microbial community across a mangrove-inhabited mudflat as revealed by 16S rDNA gene sequences. Sci. Total Environ. 2018, 633, 518–528. [Google Scholar] [CrossRef]
- Liang, S.; Deng, J.; Jiang, Y.; Wu, S.; Zhou, Y.; Zhu, W. Functional Distribution of Bacterial Community under Different Land Use Patterns Based on FaProTax Function Prediction. Pol. J. Environ. Stud. 2020, 29, 1245–1261. [Google Scholar] [CrossRef]
- Galloway, J.N.; Cowling, E.B. Reactive nitrogen and the world: 200 years of change. Ambio 2002, 31, 64–71. [Google Scholar] [CrossRef]
- Li, G.; Kim, S.; Han, S.H.; Chang, H.; Du, D.; Son, Y. Precipitation affects soil microbial and extracellular enzymatic responses to warming. Soil Biol. Biochem. 2018, 120, 212–221. [Google Scholar] [CrossRef]
- Yang, H.; Li, Y.; Wu, M.; Zhang, Z.; Li, L.; Wan, S. Plant community responses to nitrogen addition and increased precipitation: The importance of water availability and species traits. Glob. Chang. Biol. 2011, 17, 2936–2944. [Google Scholar] [CrossRef]
- Wei, H. Soil Greenhouse Gas. Emission and its Influencing Factors from Temperate Deciduous Broad-Leaved Forest and Tropical Mountain Rain Forest; Northwest A&F University: Xianyang, China, 2018. [Google Scholar]
- Stein, L.Y. The Long-Term Relationship between Microbial Metabolism and Greenhouse Gases. Trends Microbiol. 2020, 28, 500–511. [Google Scholar] [CrossRef] [PubMed]
- van Leeuwen, J.P.; Djukic, I.; Bloem, J.; Lehtinen, T.; Hemerik, L.; de Ruiter, P.C.; Lair, G.J. Effects of land use on soil microbial biomass, activity and community structure at different soil depths in the Danube floodplain. Eur. J. Soil Biol. 2017, 79, 14–20. [Google Scholar] [CrossRef]
- Jin, Y.H. Variations of Soil Microbial Diversity along an Elevation Gradient in the Wuyi Mountains; Nanjing Forestry University: Nanjing, China, 2012. [Google Scholar]
- Huang, N.; Wang, L.; Song, X.-P.; Black, T.A.; Jassal, R.S.; Myneni, R.B.; Wu, C.; Song, W.; Ji, D.; Yu, S.; et al. Spatial and temporal variations in global soil respiration and their relationships with climate and land cover. Sci. Adv. 2020, 6, eabb8508. [Google Scholar] [CrossRef]
- Zhou, Y.; Jia, X.; Han, L.; Liu, Z.; Kang, S.; Zhao, Y. Fungal community diversity in soils along an elevation gradient in a Quercus aliena var. acuteserrata forest in Qinling Mountains, China. Appl. Soil Ecol. 2021, 167, 104104. [Google Scholar] [CrossRef]
- Yang, Y.; Gao, Y.; Wang, S.; Xu, D.; Yu, H.; Wu, L.; Lin, Q.; Hu, Y.; Li, X.; He, Z.; et al. The microbial gene diversity along an elevation gradient of the Tibetan grassland. ISME J. 2014, 8, 430–440. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plots | pH | Elevation (m) | Annual Temperature (°C) | Annual Precipitation (mm) | Bulk Density (g·cm−3) |
|---|---|---|---|---|---|
| A1 | 4.10 | 1446.00 | 9.77 | 649.2 | 0.27 |
| A2 | 3.99 | 1448.00 | 9.77 | 649.2 | 0.40 |
| A3 | 4.19 | 1477.00 | 9.77 | 649.2 | 0.33 |
| A4 | 3.99 | 1446.00 | 9.77 | 649.2 | 0.31 |
| A5 | 4.18 | 1448.00 | 9.77 | 649.2 | 0.21 |
| B1 | 4.55 | 1669.00 | 7.04 | 677.5 | 0.24 |
| B2 | 4.55 | 1669.00 | 7.04 | 677.5 | 0.29 |
| B3 | 4.52 | 1669.00 | 7.04 | 677.5 | 0.27 |
| B4 | 4.66 | 1669.00 | 7.04 | 677.5 | 0.27 |
| B5 | 4.50 | 1669.00 | 7.04 | 677.5 | 0.28 |
| C1 | 4.05 | 2290.00 | 5.27 | 696.7 | 0.25 |
| C2 | 4.16 | 2280.00 | 5.27 | 696.7 | 0.23 |
| C3 | 4.11 | 2289.00 | 5.27 | 696.7 | 0.27 |
| C4 | 4.00 | 2278.00 | 5.27 | 696.7 | 0.21 |
| C5 | 4.13 | 2277.00 | 5.27 | 696.7 | 0.21 |
| Variable a | Bacterial Communities | Bacterial Functional Groups | Bacterial Functional Groups Associated with Carbon Degradation | |||
|---|---|---|---|---|---|---|
| r | p | r | p | r | p | |
| pH | 0.26 | 0.041 * | 0.26 | 0.024 * | −0.02 | 0.509 |
| Elevation | 0.07 | 0.311 | 0.57 | 0.001 ** | 0.52 | 0.002 ** |
| AT | 0.32 | 0.009 ** | 0.26 | 0.021 * | 0.38 | 0.005 ** |
| AP | 0.32 | 0.007 ** | 0.28 | 0.012 * | 0.39 | 0.002 ** |
| BD | 0.05 | 0.315 | −0.15 | 0.834 | −0.07 | 0.621 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, W.; Li, Z.; Lei, J.; Liu, X. Bacterial Communities of Forest Soils along Different Elevations: Diversity, Structure, and Functional Composition with Potential Impacts on CO2 Emission. Microorganisms 2022, 10, 766. https://doi.org/10.3390/microorganisms10040766
Sun W, Li Z, Lei J, Liu X. Bacterial Communities of Forest Soils along Different Elevations: Diversity, Structure, and Functional Composition with Potential Impacts on CO2 Emission. Microorganisms. 2022; 10(4):766. https://doi.org/10.3390/microorganisms10040766
Chicago/Turabian StyleSun, Wanlong, Zhouyuan Li, Jiesi Lei, and Xuehua Liu. 2022. "Bacterial Communities of Forest Soils along Different Elevations: Diversity, Structure, and Functional Composition with Potential Impacts on CO2 Emission" Microorganisms 10, no. 4: 766. https://doi.org/10.3390/microorganisms10040766
APA StyleSun, W., Li, Z., Lei, J., & Liu, X. (2022). Bacterial Communities of Forest Soils along Different Elevations: Diversity, Structure, and Functional Composition with Potential Impacts on CO2 Emission. Microorganisms, 10(4), 766. https://doi.org/10.3390/microorganisms10040766